 2022, Vol. 42
2022, Vol. 42文章信息
- 张世虎, 张悦, 马晓玉, 王聪, 马群, 杨雪纯, 徐婷, 马越, 郑智
- ZHANG Shihu, ZHANG Yue, MA Xiaoyu, WANG Cong, MA Qun, YANG Xuechun, XU Ting, MA Yue, ZHENG Zhi
- 大气氮沉降影响草地植物物种多样性机制研究综述
- Mechanisms underlying loss of plant biodiversity by atmospheric nitrogen deposition in grasslands
- 生态学报. 2022, 42(4): 1252-1261
- Acta Ecologica Sinica. 2022, 42(4): 1252-1261
- http://dx.doi.org/10.5846/stxb202011072845
-
文章历史
- 收稿日期: 2020-01-10
- 网络出版日期: 2021-11-04

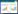




大气氮沉降是指大气中的氮元素以NHx(包括NH3、RNH2和NH4+)和NOx的形式, 降落到陆地和水体的过程[1]。氮素是植物生长必需的矿质元素, 氮沉降的增加直接促进植物迅速生长, 提高陆地生态系统的净初级生产力。同时, 氮沉降也导致土壤酸化, 改变土壤矿质养分的有效性和微生物群落结构与功能, 进而间接影响植物生长, 驱动陆地生态系统结构与功能的改变。全球大气氮沉降总量从1860年15 Tg上升到2005年187 Tg, 预计到2050年将会进一步增加至200 Tg[2]。近30年来, 我国大气氮素沉降量大量增加, 已高于北美任何地区, 与西欧20世纪80年代氮沉降高峰时的数量相当, 成为继北美、西欧的全球第三大氮沉降集中区[3]。最近评估, 我国氮沉降趋于稳定, 但氮沉降组分发生改变, 主要表现为氨氮沉降减少, 硝氮沉降增加[4]。在自然生态系统中, 植物与环境长期协同进化过程中, 已经形成了各种互补性地利用不同时空及其化学形态的矿质养分, 孕育了丰富的物种, 维持了生态系统稳定性[5]。但在地质历史上前所未有的大气氮沉降背景下, 生态系统的富营养化将对生源要素的生物地球化学循环和物种的综合适应策略产生深刻影响, 导致草地生态系统结构与功能的改变, 尤其是长期氮沉降导致草地生态系统物种多样性降低, 极大地改变了生态系统稳定性。
早期研究认为, 氮沉降降低草地植物物种多样性是不同物种或功能群之间相互竞争排斥或外源性氮输入限制群落更新所导致的[6-7]。近年来, 越来越多研究认为, 氮添加导致草地植物物种多样性降低是各种因素综合作用的结果[8]。然而, 由于研究区域土壤属性、草地植被类型、不同植物功能群或不同植物物种对氮素本身的响应的差异及其不同物种的功能属性的差异, 氮添加导致物种多样性降低的机制仍然存在一定的争议。因此, 全面系统地总结氮沉降导致草地植物物种多样性降低的生物学机制, 将有助于进一步揭示氮沉降降低草地植物物种多样性的内在规律, 同时也有利于在氮沉降背景下合理利用草地资源, 为草地生态系统的可持续利用提供理论依据。为此, 本文综述了氮沉降导致草地植物物种多样性降低已报道的主要机制, 主要包括以下几个方面: (1) 资源竞争;(2) 群落更新限制;(3) 土壤酸化及其离子毒害;(4) 营养失衡;(5) 氮素本身的毒害;(6) 次生胁迫。最后, 针对我国温带草原生态系统土壤和植被特征, 讨论了我国草地生态系统结构与功能在氮富集背景下的研究策略。
1 大气氮沉降降低草地物种多样性的可能机制大气氮沉降导致物种丧失在全球不同的草地生态系统中普遍存在。英国洛桑试验站早在1856年就开始了包括氮素在内的草地生态系统养分添加的长期定位试验, 持续至今已有160多年的连续观测结果[9-10], 该试验首次观察到长期氮素添加改变了草地植被群落结构和降低了物种多样性, 并发现氮添加导致物种丢失与土壤酸化相关的现象[10]。在欧洲6个国家不同土壤类型温带半自然草地的氮添加试验研究发现, 氮添加速率与草地植物物种多样性存在显著的负相关, 而且氮添加速率在每年0-40 kg/hm2范围导致物种多样性降低最为明显[11]。Stevens等[12]通过分析英国68个不同自然草地植物物种多样性与自然氮沉降速率的关系, 得出长期自然氮沉降速率与草地物种多样性呈显著负相关的结论, 而且每年2.5 kg/hm2的氮沉降导致每4 m2草地中有一个物种丧失。20世纪80年代, Tilman在美国Cedar Creek开展了氮添加对草地物种多样性和生产力影响的长期研究, 发现即使是低剂量的氮添加也会降低植物物种多样性[13]。近年来, 在我国蒙古高原、青藏高原、黄土高原原位氮添加试验同样也导致草地物种多样性降低[14-16]。目前, 针对氮沉降导致全球草地物种丧失的普遍现象, 各国学者提出了多种可能机制, 主要包括资源竞争排斥、群落更新限制、土壤酸化、离子毒害、养分失衡、氮素本身对植物毒害、次生胁迫等。
1.1 资源竞争排斥假说在资源限制的生态系统中, 物种通过竞争权衡(tradeoff)实现资源利用方式分化, 达到对不同时空和化学形态的限制性资源优化利用, 从而维持生态系统较高的物种多样性和稳定性[17]。生态位维度理论认为, 外源性养分添加降低限制性养分的数量或维度, 减少物种间潜在权衡机会, 降低地下资源的异质性, 从而导致物种多样性丧失[18]。因此, 资源竞争和生态位维度理论把养分添加降低草地植物物种多样性归因于外源性养分输入导致土壤资源异质性的降低。无论是美国高草草原[19]、还是英国酸性土壤草地[19]以及我国高寒草地生态系统[20], 限制性养分添加降低了草地植物物种多样性。通过限制性养分原位添加试验综合分析发现, 增加限制性养分降低了土壤资源的生态位维度和多样性, 加速群落内部物种的周转速率, 最终导致植物物种多样性的降低[18]。而且, 最近Farrer和Suding[8]利用温室控制试验证明, 资源竞争仍然是导致物种多样性丧失的主要诱因。因此, 无论在区域还是全球尺度上、在原位还是室内控制试验都证明了资源竞争限制是导致物种丧失的主要诱因。但这些原位试验仅通过试验处理后物种多样性数目与资源添加数目相关分析得出的结论, 在试验过程中并未考虑土壤资源的量变过程以及群落内部不同物种利用养分的策略, 而且室内试验仅仅通过密度控制试验, 无法说明具体的何种资源导致的物种丧失。因此, 有必要结合室内和野外原位试验, 充分考虑草地植物群落中不同物种或功能群利用土壤养分的策略, 全面系统地探讨植物利用土壤养分的策略以及养分添加对植物养分利用策略的影响, 有针对性地长期监测养分添加后土壤中限制性养分的量变过程, 为衡量生态位维度的变化提供令人信服的佐证, 最终从草地植物本身利用多种限制性资源的策略角度揭示外源性养分添加对植物适应机制的影响。
在大多数草地生态系统中, 氮素是主要的限制性资源之一。氮添加导致不同物种或功能群对限制性资源的竞争, 主要包括对地上的光竞争、地下水分和养分的竞争、地上和地下资源的总体竞争。其中, 光竞争理论一直被认为是解释氮沉降诱导物种丧失的主要机制。该理论认为, 氮添加促进一些喜氮植物的快速生长, 造成对群落中其它物种的遮荫, 从而导致冠层以下物种因受光限制而丧失[21-22]。Hautier等[22]通过对种植在室内培养箱中的6种草地植物进行氮添加处理, 发现在培养箱底部补光可以有效逆转施肥对物种多样性的负面影响, 这一结果为光竞争假说提供了有力佐证。同时, DeMalach[23]通过原位控制试验证明光竞争仍然是解释氮添加降低草地物种多样性的主要机制, 为光竞争理论提供了野外控制试验的证据。Simkim等[24]认为, 在草地生态系统中, 氮沉降引起的光竞争排斥是解释物种丢失的主要机制, 其主要原因是草地植物物种由于长期适应高光照, 对外源性氮输入引起的暂时性遮荫很难适应, 从而引起矮杆喜光植物物种丢失。
但也有人对光竞争引起植物物种丧失的机制提出质疑。主要是Dickson和Foster[25]的研究团队, 他们通过比较施肥、草地遮荫、刈割等对草地植物群落的影响, 发现施氮导致草地植物物种丧失与光竞争无关, 并认为温室和原位草地生态系统光强的差别可能是造成Hautier等[22]和Dickson和Foster[25]结论差异的主要原因。在Hautier等[22]的研究中, 培养箱植株冠层的光强只有470 μmol m-2 s-1, 而在实际草地植株冠层的光强高达1500 μmol m-2 s-1, 因此培养箱中种植的植物本身已经受到弱光胁迫, 在植株冠层下方补光能有效减小弱光胁迫, 改变植物的生长, 降低物种丧失。同时, 在内蒙古典型草原, 通过刈割改善光照后, 发现刈割对氮添加导致的物种多样性降低没有显著的影响[26]。目前, 关于光竞争假设遇到的瓶颈是所有与光竞争假设相关的证据并未考虑植物本身的光合属性。因此, 在未来的研究中, 应结合不同植物的光合相关功能性状将有助于进一步深化光竞争理论。
除了对地上光资源的竞争, 群落内部物种间对地下资源的竞争可能也与氮沉降导致草地植物多样性降低有关[27]。地下资源竞争理论认为, 养分添加提高了土壤养分分布的异质性或斑块状分布, 能优先获取土壤“斑块”中养分的物种最终成为群落中的优势种, 从而改变了群落的组成和降低物种多样性[27]。同时, 也有人认为, 地上和地下资源的总体竞争是导致氮添加导致草地生态系统物种丧失的主要原因[25]。但外源性氮素输入引起的植物对地上和地下资源的竞争必会引起植物碳分配格局的改变, 对于植物体本身而言, 碳分配格局是衡量植物自身资源分配均衡的主要指标, 碳分配格局的改变意味破坏植物自身资源分配的稳态, 无论向哪个方向的改变, 对植物本身而言都是不利的, 能稳定自身资源分配平衡的物种或许最终才能在竞争中取胜。因此, 从植物调节自身资源分配的内稳机制出发, 并结合植物吸收地上和地下资源的生理生态特征, 将有助于解析长期氮沉降降低草地植物物种多样性的潜在机制。
1.2 群落更新限制新物种定植速率和已有物种灭绝速率的动态平衡决定局域(local) 物种多样性, 因此群落更新限制假说认为, 氮添加导致物种多样性降低是由于降低了局域物种的定植速率, 增加了已有物种的灭绝速率所致, 其基本解释为长期氮添加促进了地面凋落物积累, 从而抑制种子萌发和幼苗定植[7]。此外, 喜氮植物的快速生长造成对其它临近植物遮荫, 从而导致喜光植物的幼苗长期受光资源限制而死亡, 加速了物种灭绝速率[7]。这一理论最早是Tilman[7]根据北美Cedar Greek的4个氮添加野外控制试验平台11年连续监测数据所提出的, 并得到了逐步的完善和拓展;例如Herry等[28]的研究结论是氮添加驱动的光限制降低幼苗的成活率影响新物种定植, 而Price和Morgan[29]认为, 短期氮添加提高本地物种的灭绝速率和降低外来物种的定值速率, 从而限制了植物群落更新。此外, 长期氮沉降也降低了草地土壤种子库丰度[30], 这也许直接抑制群落幼苗物种多样性, 最终影响植物群落物种组成和更新。和北美草地生态系统一样, 氮添加也提高我国北方草地本地物种的灭绝速率和降低外来物种的定植速率[31-32], 而且施氮频度对新物种定植速率具有重要影响, 其主要原因是施氮频度的降低提高了土壤中铵态氮浓度, 可能对新生幼苗产生毒害[31]。这说明和北美草地一样, 氮添加限制我国北方草地植物群落的更新, 究其成因机制明显不同。但这些结论都是来自数学统计相关分析的推论, 缺乏真正生物学过程的研究。一般而言, 对于生产力较低的草地生态系统, 地面凋落物很少, 光照充分, 生长在该生态系统的植物物种进化出适应凋落物少和高光照的特征, 因此氮添加导致凋落物的增加和光照强度的限制也许是导致物种丧失的重要机制, 但针对生产力和凋落物较高的草地生态系统, 这种机制是否起作用需要进一步的研究。
1.3 土壤酸化及其离子毒害土壤是植物生长和发育的载体, 任何影响土壤物属性的因素都会影响植物生长。大气氮沉降一方面增加土壤氮含量, 同时也作为酸源, 降低土壤pH, 导致土壤酸化[1]。土壤pH主要由土壤母质决定, 它通过控制土壤的矿质养分有效性, 影响植物生长[33]。大气氮沉降对英国酸性草地植物群落物种多样性影响的研究表明, 土壤酸化可能是导致大气氮沉降引起物种丧失的主要原因[34]。但近期在内蒙古草原对氮添加长期监测发现, 短期氮添加并未显著降低土壤pH, 但植物物种多样性出现了明显降低, 说明除了土壤酸化外, 氮富集导致物种丢失可能还存在其它机制[35]。
无论是在酸性还是碱性土壤草地, 植物物种都经过长期自然选择, 高度适应了其土壤特性。从理论上推测, 氮沉降通过酸化土壤, 改变土壤矿质养分的稳态和有毒金属离子的释放, 造成植物毒害, 进而影响草地物种多样性[1]。此外, 氮沉降通过酸化土壤改变土壤矿质养分有效性, 影响植物生长和物种多样性的研究主要来自欧洲酸性草地[36-38]。酸性土壤中Al3+、Mn2+和Fe3+浓度较高, 对植物产生潜在的毒害作用。铝毒害是酸性土壤中制约植物生长的主要环境因子[35]。当土壤pH < 5时, 对植物有毒害的Al3+在土壤中活化[39]。Al3+的毒害作用主要是抑制根系生长, 最终导致根系吸收水分和养分功能丧失, 影响植物的正常生长[37-38]。虽然有研究结果表明, 大气氮沉降引发的草地物种多样性降低与土壤中Al3+含量呈正相关关系[38], 但在氮富集条件下保留的物种耐铝毒的机制仍不清楚。由于酸性土壤中的本底Mn2+含量相对较高, 酸性草地植物可能已经具备耐高锰的能力, 而且氮富集导致的土壤酸化对进一步增加其含量的幅度较小[40], 所以未见有酸性草地植物遭受锰毒害的报导。另外, 土壤酸化会使土壤释放更多的Fe3+, 但双子叶植物不能直接吸收Fe3+, 需要借助植物根系的铁还原酶将Fe3+还原为Fe2+才能被植物吸收[33], 所以也没有在酸性土壤中报道铁毒害的现象。
与酸性土壤草地不同, 我国蒙古高原、青藏高原、黄土高原草地土壤属于偏碱性钙质性土壤, 生长在其上的植物物种高度适应了其土壤养分状态, 即土壤的Fe3+、Mn2+、磷等有效性低, 而盐基阳离子(Ca2+、Mg2+、K+)含量较高, 土壤缓冲酸化能力较强。大量调查与野外氮添加控制试验表明, 氮富集同样能导致我国蒙古高原、青藏高原、黄土高原草地土壤酸化和物种多样性降低[14-16]。在内蒙古典型草原, 施氮(16 kg hm-2 a-1和32 kg hm-2 a-1)8年后土壤pH由7.05分别降至6.18和5.79, 而植物物种多样性则分别降低了35% 和39%[35]。进一步研究发现, 氮添加酸化土壤的同时, 显著增加了土壤中Mn2+、Fe3+、Cu2+的含量, 其中Mn2+离子含量的增加最为明显[41]。土壤Mn2+的含量主要由土壤pH和氧化还原电位决定, 我国蒙古高原、青藏高原、黄土高原具有高pH和高氧化还原电位。因此, 其土壤具有低Mn2+含量的特征。氮富集诱导的土壤Mn2+活化, 使长期生长在低Mn2+土壤环境的植物面临巨大锰毒害的危险。与酸性土壤中的铝毒害不同, 植物过量的累积锰会影响铁的吸收, 最终导致光合速率的降低[42]。双子叶植物和单子叶禾草对铁的吸收存在不同机制, 因此氮富集导致的土壤酸化促进土壤Mn2+的释放, 使双子叶植物叶片累积过量Mn造成毒害, 影响Fe的吸收及其光合等生理过程, 这已经成为温带碱性钙质草地氮沉降导致植物多样性降低的一个重要机制[35](图 1)。

|
| 图 1 氮沉降导致土壤酸化降低草地物种多样性的图解 Fig. 1 Diagram on evoked-nitrogen reduction in plant diversity ↑和↓分别代表增加和降低; MT和IT为金属传递通道和离子通道的缩写; N, C, Pn分别为氮、碳、光合的缩写 |
在自然生态系统中, 植物物种与其环境协同进化过程中形成了稳定的化学计量关系, 当大规模外源性养分输入草地生态系统后, 打破了已有的稳定化学计量关系, 引起土壤和植物体内养分失衡, 可能诱导一些物种在植物群落中丧失。氮沉降主要影响土壤和植物体内氮磷平衡, 最终影响植物群落物种组成[43]。在磷限制的草地生态系统中, 土壤中较低的磷含量往往限制低量的氮沉降引起的快速型植物的生长, 从而减轻氮沉降对物种多样性的影响, 但就大规模的氮沉降而言, 较低的土壤磷含量往往引起植物体内氮磷比大幅增加, 一些植物因磷限制而在植物群落中丧失, 从而降低了植物群落的物种多样性[43]。与氮限制草地生态系统相比, 磷限制草地生态系统孕育更多的物种, 特别是濒危物种、杂类草物种多样性明显高于氮限制生态系统[44-46]。在荷兰和比利时, 70%的濒危物种生活在磷限制草地生态系统[44], 其主要原因在于: 磷在土壤中存在形式的多样性, 不同植物或功能群物种形成获取磷的不同策略, 比如根系分泌磷酸活性酶、酸化根际、形成菌根或排根, 产生了生态位分化, 最终形成较大适应低磷生态系统的物种库。此外, 根据植物功能性状, 对磷敏感的植物往往具有株型低矮、抗逆性强、生长缓慢等特征[45-46], 土壤中有效磷的增加促进生长较快的植物迅速生长对其产生光竞争排斥[46]。Soons等[47]通过整合全球氮、磷养分添加试验数据, 仍然认为氮添加(并非磷添加)是导致植物物种多样性丧失主要驱动因子。
在我国蒙古高原、青藏高原、黄土高原草地生态系统, 土壤中有效磷的含量较低, 长期生活在该土壤中的植物已经形成适应低磷的机制, 比如: 在内蒙古典型草原生长共优势种植物冷蒿(Artemisia frigida)和克氏针茅(Stipa krylovii), 对磷具有不同利用策略[48], 但连续12年以每年160 kg/hm2的氮添加显著提高土壤中有效磷含量[49], 这是否与氮沉降导致的物种丧失有一定的联系, 目前尚未进行系统研究。此外, 目前磷对植物物种多样性的影响的报道主要集中在欧洲酸性土壤的草地生态系统, 是否在我国北方碱性土壤草原也存在这种现象, 有待于进一步调查或长期监测研究。已有大量的研究报道, 在澳大利亚和南非磷贫瘠的土壤中生长的植物, 当外源性磷输入增加土壤中有效磷含量时, 这些植物出现了叶片衰老和坏死, 通常把这种现象称为“磷毒”[49]。磷毒的形成机制在生理和分子水平进行了广泛的研究, 但目前尚未达成共识[49]。植物磷毒也可能解释在磷限制生态系统比磷丰富生态系统物种多样性更高的现象。但目前尚未有在氮添加水平下土壤有效磷的增加对草地生态系统植物物种产生磷毒害的报道。
1.5 氮素本身对植物的毒害在自然生态系统, 共存物种由于长期竞争导致种内和种间利用养分资源的分化, 形成不同植物以不同方式利用土壤中不同化学形态养分资源的格局, 从而促进物种共存, 保持物种多样性, 维持生态系统的稳定性。大规模的大气氮沉降增加土壤有效氮含量, 改变了土壤中不同形态氮的比例。因此, 植物能灵活改变氮的利用方式的物种有可能在竞争中取胜[50]。植物优先吸收的氮素形态与土壤中各种形态氮的有效性有关[51]。例如, 在北极冻原、温带草地、北方针叶林等土壤贫瘠的生态系统中的植物能吸收有机氮[52]。在北极冻原, 土壤中氮素不仅在时空上出现分化, 而且植物群落中优势种往往利用最丰富化学形态氮, 最终几乎形成群落中物种丰度与土壤中氮丰度相对应的关系[53], 氮形态分化一方面促进物种共存, 另一方面提高了氮利用效率。然而, 在温带草地生态系统, 植物主要吸收无机态氮[54], 特别是硝态氮[55], 这也许是在温带草地近似中性的土壤环境中, 硝态氮是土壤中氮的主要存在形式。
在我国, 铵态氮是大气氮沉降的主要形态[4]。当氮素以铵态氮的形式进入土壤往往会引起土壤酸化, 从而抑制其硝化过程, 导致土壤中铵态氮积累, 提高土壤中铵态氮比例。土壤中主要形态氮及其不同形态氮比例影响植物生长, 而且长期生长在不同生境中的植物对土壤中不同形态氮利用具有一定的选择性。一般而言, 适应酸性土壤生长的嫌钙和低氧化还原势土壤条件下生长的植物主要利用铵态氮, 而在pH较高的石灰性土壤上生长喜钙植物主要吸收硝态氮[50]。在我国北方温带草地生态系统, 大多数土壤属于钙质的偏碱性土壤, 由此推测在其生长的大多数植物主要吸收利用硝态氮。但这些植物是通过何种策略应对以铵态氮沉降为主的氮沉降, 目前缺乏系统研究。
一直以来, 铵态氮对草地植物物种多样性影响的研究主要集中在欧州欧石楠土壤和酸性土壤中, 而我国北方草地土壤属于偏碱性的石灰性土壤, 土壤中铵态氮浓度相对较低。但长期氮添加导致土壤酸化, 是否铵态氮也会对草地植物造成毒害需要长期系统的研究。尽管在我国草地生态系统已经有大量的原位氮添加试验平台, 但研究不同形态氮对草地植物影响的报道相对较少。到目前为止, 仅有Song等[56]报道6年的不同形态氮添加对我国青藏高原高寒草地生态系统物种多样性没有显著的影响, 但不同功能群的物种对不同形态氮的响应不同。尽管这份研究首次在国内报道不同形态氮对草地植物群落的影响, 但并未考虑在研究地点土壤碱性的特征(pH=8), 而且该试验也没有使用硝化抑制剂, 在这种条件下施用的铵态氮很快通过硝化作用转化成硝态氮, 铵态氮在土壤中保留时间很短, 因此该研究结果可能低估了铵态氮对植物群落的影响。此外, 已有的研究发现, 在北极冻原, 土壤中氮矿化速率极低, 许多植物形成了能吸收有机氮策略[53], 是否在我国青藏高原草地植物物种也具有这种氮吸收策略, 目前尚未报道。因此, 一方面采用合理的方法抑制铵态氮在土壤中硝化过程, 合理评价不同形态氮对我国北方草地生态系统、不同功能群及其物种的影响, 以便解析不同植物及其功能群物种对氮沉降的不同时空响应, 将有助于进一步阐明氮素本身对草地植物多样性的影响;另一方面, 采用同位素标记不同形态氮, 研究不同区域、不同草地类型植物物种或功能群对氮素吸收利用策略, 对认识氮沉降背景下草地植物群落结构与功能的变化具有重要意义。
1.6 次生胁迫大规模的大气氮沉降通过降低植物对病虫害的抵抗能力和改变土壤微生物群落组成等间接影响植物物种的多样性[1]。首先, 氮沉降提高了植物群落生产力、增加了植物体内蛋白质含量和改变了蛋白质组成、降低了植物体内钙含量、改变次生代谢产物含量, 最终不仅提高了病原菌及其昆虫食物资源的质量和数量, 而且降低植物对病虫害的防御和抵抗能力[57]。因此, 长期氮添加可能会导致草地植物群落病虫害的爆发, 间接降低草地植物物种多样性。Bobbink等[58]认为大气氮沉降是欧洲欧石楠荒漠中帚石楠甲虫(Lochmaea suturalis) 频繁爆发的主要原因, 但这与对我国内蒙古草原亚洲小飞蝗(Oedaleus asiaticus) 爆发的原因恰恰相反, 如在内蒙古温带草原, 重度放牧导致植物体内氮含量下降, 从而引起喜食低氮的亚洲小飞蝗爆发[59]。与禾草相比, 杂类草叶片中蛋白含量更高, 硅和钙的含量相对较低, 因此杂类草也许更易遭受虫害。在美国高草草原长期氮添加和昆虫原位控制试验研究发现, 控制昆虫增加了群落中杂类草相对丰度, 导致禾草和杂类草的比率增加[60], 这也可能是氮添加后期一些杂类草进一步丧失有关。在内蒙古典型草原12年的氮肥添加发现, 每年20 kg/hm2氮添加明显降低杂类草冷蒿叶片中的钙含量[26], 但是否在我国北方温带草原, 长期氮沉降是否导致虫害发生, 从而降低地上植物物种多样性, 目前尚未报道。
此外, 大规模的大气氮沉降也会降低草地植物对病原菌的抵抗能力, 从而间接降低植物物种多样性;其主要原因是氮添加提高植物叶片中氮含量, 为病原菌生长、繁殖提供养料, 或者氮沉降降低了植物体内防御性化合物的含量[61]。此外, 不同功能群物种对长期氮添加引发的病害具有不同的响应, 例如在美国高草草原, 长期氮添加增加碳四禾草体内蛋白含量, 从而提高真菌病害侵染和繁殖, 降低了碳四禾草叶片寿命[62], 因此这也许是长期氮添加导致在美国碳四禾草优势种逐渐被碳三禾草所取代的原因之一。氮沉降往往降低土壤和宿主植物体内菌根真菌的多样性性和生物量, 导致菌根真菌为宿主植物供给养分的能力下降, 进而间接影响地上植物的群落组成[63]。在内蒙古典型草原连续12年氮添加降低优势种杂类草冷蒿根内丛枝菌根真菌的多样性和侵染率, 但优势种禾草丛枝菌根真菌对氮添加没有任何响应[64]。因此, 揭示不同功能群或物种对菌根真菌和氮添加的响应机制, 将有助于进一步阐明菌根真菌在植物物种丧失过程中的作用。
2 结论与展望大气氮沉降降低草地植物物种多样性已成为普遍现象, 就其成因机制目前尚未达成共识, 因此有必要开展系统综合的动态研究。目前已报道的可能机制主要包括氮添加引起的不同功能群植物对地上和地下资源的竞争、植物群落更新限制、土壤酸化诱导的金属离子毒害、氮素本身对植物的影响、外源性氮输入对植物及其土壤生态系统养分稳态的扰动、氮沉降引起的对植物病害、虫害、土壤微生物群落的影响间接作用于地上植物群落的过程。但在以往氮沉降对草地植物物种多样性的研究中, 缺乏在氮沉降背景下土壤过程、植物功能性状、植物生理生态过程的时空变化的系统整合研究, 导致难以区分众多土壤和生物学过程变化在草地生态系统的结构与功能对氮沉降响应中的因果关系, 因此在以后的研究中应该优先考虑以下几个方面: 首先, 关注氮沉降对土壤变化过程的影响, 研究氮添加对土壤pH、矿质元素有效性变化、不同植物矿质养分利用策略、植物根系属性、光合特性等影响, 在物种、功能群和群落水平, 探讨草地植物多样性对氮沉降的响应, 揭示氮沉降对草地生态系统生物地球化学循环、物种适应对策、生物多样性维持的作用机理, 阐明在氮沉降背景下土壤圈-植物圈相互作用过程在草地生态系统多样性与生产力维持中的作用机制, 为草地生态系统的可持续利用提供理论依据;其次, 与欧洲酸性土壤草地生态系统相比, 我国北方草地属于钙质土壤, 土壤pH相差甚远, 因此尽管外源性氮输入导致土壤酸化, 但很少达到欧洲草地土壤酸化的程度, 所以在土壤酸化未达到铝离子对植物毒害之前, 土壤中锰离子浓度的剧增已经对一些植物产生毒害, 但这一过程还需要进一步长期监测金属离子毒害的动态过程, 而且需要挖掘不同植物物种、功能群以及同一植物地上和地下部分对金属离子响应差异的生理生态机制;再次, 氮沉降对草地生态系统物种多样性的影响是一个长期的动态过程, 是外源性氮输入引起的各种因子变化综合作用的结果, 但每种机制也许在不同时期或不同的草地生态系统中占主导地位, 因此有必要在将来的研究中探讨驱动不同时空草地生态系统物种多样性对氮沉降响应的主要机制。总之, 氮沉降降低草地生态系统植物物种多样性是多尺度、多机制共同作用的结果, 涉及地上与地下资源、短期氮素本身的影响和长期氮素积累形成次生胁迫、种间和种内、功能群之间、植物本身的进化适应机制及其功能性状等综合作用的结果, 全面系统地考虑这些因素在不同时空中相对贡献和绝对贡献, 有待于阐明其生物学机制。
| [1] |
Bobbink R, Hicks K, Galloway J, Spranger T, Alkemade R, Ashmore M, Bustamante M, Cinderby S, Davidson E, Dentener F, Emmett B, Erisman J W, Fenn M, Gilliam F, Nordin A, Pardo L, De Vries W. Global assessment of nitrogen deposition effects on terrestrial plant diversity: a synthesis. Ecological Applications: A Publication of the Ecological Society of America, 2010, 20(1): 30-59. DOI:10.1890/08-1140.1 |
| [2] |
Galloway J N, Townsend A R, Erisman J W, Bekunda M, Cai Z C, Freney J R, Martinelli L A, Seitzinger S P, Sutton M A. Transformation of the nitrogen cycle: recent trends, questions, and potential solutions. Science, 2008, 320(5878): 889-892. DOI:10.1126/science.1136674 |
| [3] |
Liu X J, Zhang Y, Han W X, Tang A H, Shen J L, Cui Z L, Vitousek P, Erisman J W, Goulding K, Christie P, Fangmeier A, Zhang F S. Enhanced nitrogen deposition over China. Nature, 2013, 494(7438): 459-462. DOI:10.1038/nature11917 |
| [4] |
Yu G R, Jia Y L, He N P, Zhu J X, Chen Z, Wang Q F, Piao S L, Liu X J, He H L, Guo X B, Zhang W, Li P, Ding G A, Goulding K. Stabilization of atmospheric nitrogen deposition in China over the past decade. Nature Geoscience, 2019, 12(6): 424-429. DOI:10.1038/s41561-019-0352-4 |
| [5] |
Loreau M, Naeem S, Inchausti P, Bengtsson J, Grime J P, Hector A, Hooper D U, Huston M A, Raffaelli D, Schmid B, Tilman D, Wardle D A. Biodiversity and ecosystem functioning: current knowledge and future challenges. Science, 2001, 294(5543): 804-808. DOI:10.1126/science.1064088 |
| [6] |
Grime J P. Competitive exclusion in herbaceous vegetation. Nature, 1973, 242(5396): 344-347. DOI:10.1038/242344a0 |
| [7] |
Tilman D. Species richness of experimental productivity gradients: how important is colonization limitation?. Ecology, 1993, 74(8): 2179-2191. DOI:10.2307/1939572 |
| [8] |
Farrer E C, Suding K N. Teasing apart plant community responses to N enrichment: the roles of resource limitation, competition and soil microbes. Ecology letters, 2016, 19(10): 1287-1296. DOI:10.1111/ele.12665 |
| [9] |
Storkey J, Macdonald A J, Poulton P R, Scott T, Köhler I H, Schnyder H, Goulding K W T, Crawley M J. Grassland biodiversity bounces back from long-term nitrogen addition. Nature, 2015, 528(7582): 401-404. DOI:10.1038/nature16444 |
| [10] |
Silvertown J, Poulton P, Johnston E, Edwards G, Heard M, Biss P M. The Park Grass Experiment 1856-2006:its contribution to ecology. Journal of Ecology, 2006, 94(4): 801-814. DOI:10.1111/j.1365-2745.2006.01145.x |
| [11] |
Bobbink R. Plant Species Richness and the Exceedance of Empirical Nitrogen Critical Loads: An Inventory. Report Landscape Ecology. Bilthoven: Utrecht University/RIVM, 2004.
|
| [12] |
Stevens C J, Dise N B, Mountford J O, Gowing D J. Impact of nitrogen deposition on the species richness of grasslands. Science, 2004, 303(5665): 1876-1879. DOI:10.1126/science.1094678 |
| [13] |
Clark C M, Tilman D. Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands. Nature, 2008, 451(7179): 712-715. DOI:10.1038/nature06503 |
| [14] |
Zhang Y H, Lü X T, Isbell F, Stevens C, Han X, He N P, Zhang G M, Yu Q, Huang J H, Han X G. Rapid plant species loss at high rates and at low frequency of N addition in temperate steppe. Global Change Biology, 2014, 20(11): 3520-3529. DOI:10.1111/gcb.12611 |
| [15] |
Zhang D Y, Peng Y F, Li F, Yang G B, Wang J, Yu J C, Zhou G Y, Yang Y H. Trait identity and functional diversity co-drive response of ecosystem productivity to nitrogen enrichment. Journal of Ecology, 2019, 107(5): 2402-2414. DOI:10.1111/1365-2745.13184 |
| [16] |
何玉惠, 刘新平, 谢忠奎. 氮素添加对黄土高原荒漠草原草本植物物种多样性和生产力的影响. 中国沙漠, 2015, 35(1): 66-71. |
| [17] |
Tilman D. Resource Competition and Community Structure. Princeton: Princeton University Press, 1982.
|
| [18] |
Harpole W S, Sullivan L L, Lind E M, Firn J, Adler P B, Borer E T, Chase J, Fay P A, Hautier Y, Hillebrand H, MacDougall A S, Seabloom E W, Williams R, Bakker J D, Cadotte M W, Chaneton E J, Chu C J, Cleland E E, D'Antonio C, Davies K F, Gruner D S, Hagenah N, Kirkman K, Knops J M H, La Pierre K J, McCulley R L, Moore J L, Morgan J W, Prober S M, Risch A C, Schuetz M, Stevens C J, Wragg P D. Addition of multiple limiting resources reduces grassland diversity. Nature, 2016, 537(7618): 93-96. DOI:10.1038/nature19324 |
| [19] |
Harpole W S, Tilman D. Grassland species loss resulting from reduced niche dimension. Nature, 2007, 446(7137): 791-793. DOI:10.1038/nature05684 |
| [20] |
Ren Z W, Li Q, Chu C J, Zhao L Q, Zhang J Q, Ai D, Yang Y B, Wang G. Effects of resource additions on species richness and ANPP in an alpine meadow community. Journal of Plant Ecology, 2010, 3(1): 25-31. DOI:10.1093/jpe/rtp034 |
| [21] |
Newman E I. Competition and diversity in herbaceous vegetation. Nature, 1973, 244(5414): 310-310. DOI:10.1038/244310a0 |
| [22] |
Hautier Y, Niklaus P A, Hector A. Competition for light causes plant biodiversity loss after eutrophication. Science, 2009, 324(5927): 636-638. DOI:10.1126/science.1169640 |
| [23] |
DeMalach N, Zaady E, Kadmon R. Light asymmetry explains the effect of nutrient enrichment on grassland diversity. Ecology Letters, 2017, 20(1): 60-69. DOI:10.1111/ele.12706 |
| [24] |
Simkin S M, Allen E B, Bowman W D, Clark C M, Belnap J, Brooks M L, Cade B S, Collins S L, Geiser L H, Gilliam F S, Jovan S E, Pardo L H, Schulz B K, Stevens C J, Suding K N, Throop H L, Waller D M. Conditional vulnerability of plant diversity to atmospheric nitrogen deposition across the United States. Proceedings of the National Academy of Sciences, 2016, 113(15): 4086-4091. DOI:10.1073/pnas.1515241113 |
| [25] |
Dickson T L, Foster B L. Fertilization decreases plant biodiversity even when light is not limiting. Ecology Letters, 2011, 14(4): 380-388. DOI:10.1111/j.1461-0248.2011.01599.x |
| [26] |
Tian Q Y, Liu N N, Bai W M, Li L H, Chen J Q, Reich P B, Yu Q, Guo D L, Smith M D, Knapp A K, Cheng W X, Lu P, Gao Y, Yang A, Wang T Z, Li X, Wang Z W, Ma Y B, Han X G, Zhang W H. A novel soil manganese mechanism drives plant species loss with increased nitrogen deposition in a temperate steppe. Ecology, 2016, 97(1): 65-74. DOI:10.1890/15-0917.1 |
| [27] |
Rajaniemi T K. Why does fertilization reduce plant species diversity? Testing three competition-based hypotheses. Journal of Ecology, 2002, 90(2): 316-324. DOI:10.1046/j.1365-2745.2001.00662.x |
| [28] |
Henry M, Stevens H, Bunker D E, Schnitzer S A, Carson W P. Establishment limitation reduces species recruitment and species richness as soil resources rise. Journal of Ecology, 2004, 92(2): 339-347. DOI:10.1111/j.0022-0477.2004.00866.x |
| [29] |
Price J N, Morgan J W. Vegetation dynamics following resource manipulations in herb-rich woodland. Plant Ecology, 2007, 181(1): 29-37. |
| [30] |
Basto S, Thompson K, Phoenix G, Sloan V, Leake J, Rees M. Long-term nitrogen deposition depletes grassland seed banks. Nature Communications, 2015, 6(1): 6185. DOI:10.1038/ncomms7185 |
| [31] |
Zhang Y H, Stevens C J, Lu X T, He N P, Huang J H, Han X G. Fewer new species colonize at low frequency N addition in a temperate grassland. Functional Ecology, 2016, 30(7): 1247-1256. DOI:10.1111/1365-2435.12585 |
| [32] |
Xu Z W, Wan S Q, Ren H Y, Han X G, Li M H, Cheng W X, Jiang Y. Effects of water and nitrogen addition on species turnover in temperate grasslands in northern China. PLoS One, 2012, 7(6): e39762. DOI:10.1371/journal.pone.0039762 |
| [33] |
Marschner H. Mineral Nutrition of Higher Plants. 2nd ed. London: Academic Press, 1995.
|
| [34] |
Maskell L C, Smart S M, Bullock J M, Thompson K, Stevens C J. Nitrogen deposition causes widespread loss of species richness in British habitats. Global Change Biology, 2010, 16(2): 671-679. DOI:10.1111/j.1365-2486.2009.02022.x |
| [35] |
Tian Q Y, Yang L Y, Ma P F, Zhou H R, Liu N N, Bai W M, Wang H, Ren L F, Lu P, Han W W, Schultz P A, Bever J D, Zhang F S, Lambers H, Zhang W H. Below-ground-mediated and phase-dependent processes drive nitrogen-evoked community changes in grasslands. Journal of Ecology, 2020, 108(5): 1874-1887. DOI:10.1111/1365-2745.13415 |
| [36] |
Roem W J, Berendse F. Soil acidity and nutrient supply ratio as possible factors determining changes in plant species diversity in grassland and heathland communities. Biological Conservation, 2000, 92(2): 151-161. DOI:10.1016/S0006-3207(99)00049-X |
| [37] |
Bowman W D, Cleveland C C, Halada L, Hreško J, Baron J S. Negative impact of nitrogen deposition on soil buffering capacity. Nature Geoscience, 2008, 1(11): 767-770. DOI:10.1038/ngeo339 |
| [38] |
Horswill P, O'Sullivan O, Phoenix G K, Lee J A, Leake J R. Base cation depletion, eutrophication and acidification of species-rich grasslands in response to long-term simulated nitrogen deposition. Environmental Pollution, 2008, 155(2): 336-349. DOI:10.1016/j.envpol.2007.11.006 |
| [39] |
Tyler G. Soil chemistry and plant distributions in rock habitats of southern Sweden. Nordic Journal of Botany, 1996, 16(6): 609-635. DOI:10.1111/j.1756-1051.1996.tb00279.x |
| [40] |
Rengel Z, Zhang W H. Role of dynamics of intracellular calcium in aluminium-toxicity syndrome. New Phytologist, 2003, 159(2): 295-314. DOI:10.1046/j.1469-8137.2003.00821.x |
| [41] |
Tian Q Y, Liu N N, Bai W M, Li L H, Zhang W H. Disruption of metal ion homeostasis in soils is associated with nitrogen deposition-induced species loss in an Inner Mongolia steppe. Biogeosciences, 2015, 12(11): 3499-3512. DOI:10.5194/bg-12-3499-2015 |
| [42] |
Millaleo R, Reyes-Díaz M, Alberdi M, Ivanov A G, Krol M, Hüner N P A. Excess manganese differentially inhibits photosystem I versus II in Arabidopsis thaliana. Journal of Experimental Botany, 2013, 64(1): 343-354. DOI:10.1093/jxb/ers339 |
| [43] |
Güsewell S. N: P ratios in terrestrial plants: variation and functional significance. New Phytologist, 2004, 164(2): 243-266. DOI:10.1111/j.1469-8137.2004.01192.x |
| [44] |
Wassen M J, Venterink H O, Lapshina E D, Tanneberger F. Endangered plants persist under phosphorus limitation. Nature, 2005, 437(7058): 547-550. DOI:10.1038/nature03950 |
| [45] |
Ceulemans T, Merckx R, Hens M, Honnay O. Trait-based analysis of the role of phosphorus vs. nitrogen enrichment in plant species loss across North-west European grasslands. Journal of Applied Ecology, 2011, 48(5): 1155-1163. DOI:10.1111/j.1365-2664.2011.02023.x |
| [46] |
Ceulemans T, Merckx R, Hens M, Honnay O. Plant species loss from European semi-natural grasslands following nutrient enrichment-is it nitrogen or is it phosphorus?. Global Ecology and Biogeography, 2013, 22(1): 73-82. DOI:10.1111/j.1466-8238.2012.00771.x |
| [47] |
Soons M B, Hefting M M, Dorland E, Lamers L P M, Versteeg C, Bobbink R. Nitrogen effects on plant species richness in herbaceous communities are more widespread and stronger than those of phosphorus. Biological Conservation, 2017, 212: 390-397. DOI:10.1016/j.biocon.2016.12.006 |
| [48] |
刘娜娜, 田秋英, 张文浩. 内蒙古典型草原优势种冷蒿和克氏针茅对土壤低磷环境适应策略的比较. 植物生态学报, 2014, 38(9): 905-915. |
| [49] |
Lambers H, Raven J A, Shaver G R, Smith S E. Plant nutrient-acquisition strategies change with soil age. Trends in Ecology & Evolution, 2008, 23(2): 95-103. |
| [50] |
Ashton I W, Miller A E, Bowman W D, Suding K N. Niche complementarity due to plasticity in resource use: plant partitioning of chemical N forms. Ecology, 2010, 91(11): 3252-3260. DOI:10.1890/09-1849.1 |
| [51] |
Lambers H, Chapin III F S, Pons T L. Plant Physiological Ecology. New York: Springer, 1998.
|
| [52] |
Von Felten S, Hector A, Buchmann N, Niklaus P A, Schmid B, Scherer-Lorenzen M. Belowground nitrogen partitioning in experimental grassland plant communities of varying species richness. Ecology, 2009, 90(5): 1389-1399. DOI:10.1890/08-0802.1 |
| [53] |
McKane R B, Johnson L C, Shaver G R, Nadelhoffer K J, Rastetter E B, Fry B, Giblin A E, Kielland K, Kwiatkowski B L, Laundre J A, Murray G. Resource-based niches provide a basis for plant species diversity and dominance in arctic tundra. Nature, 2002, 415(6867): 68-71. DOI:10.1038/415068a |
| [54] |
Harrison K A, Bol R, Bardgett R D. Preferences for different nitrogen forms by coexisting plant species and soil microbes. Ecology, 2007, 88(4): 989-999. DOI:10.1890/06-1018 |
| [55] |
Kahmen A, Renker C, Unsicker S B, Buchmann N. Niche complementarity for nitrogen: An explanation for the biodiversity and ecosystem functioning relationship?. Ecology, 2006, 87(5): 1244-1255. DOI:10.1890/0012-9658(2006)87[1244:NCFNAE]2.0.CO;2 |
| [56] |
Song M H, Yu F H, Ouyang H, Cao G M, Xu X L, Cornelissen J H C. Different inter-annual responses to availability and form of nitrogen explain species coexistence in an alpine meadow community after release from grazing. Global Change Biology, 2012, 18(10): 3100-3111. DOI:10.1111/j.1365-2486.2012.02738.x |
| [57] |
Kleijn D, Bekker R M, Bobbink R, De Graaf M C C, Roelofs J G M. In search for key biogeochemical factors affecting plant species persistence in heathland and acidic grasslands: a comparison of common and rare species. Journal of Applied Ecology, 2008, 45(2): 680-687. DOI:10.1111/j.1365-2664.2007.01444.x |
| [58] |
Bobbink R, Hornung M, Roelofs J G M. The effects of air-borne nitrogen pollutants on species diversity in natural and semi-natural European vegetation. Journal of Ecology, 1998, 86(5): 717-738. DOI:10.1046/j.1365-2745.1998.8650717.x |
| [59] |
Cease A J, Elser J J, Ford C F, Hao S G, Kang L, Harrison J F. Heavy livestock grazing promotes locust outbreaks by lowering plant nitrogen content. Science, 2012, 335(6067): 467-469. DOI:10.1126/science.1214433 |
| [60] |
La Pierre K J, Smith M D. Soil nutrient additions increase invertebrate herbivore abundances, but not herbivory, across three grassland systems. Oecologia, 2016, 180(2): 485-497. DOI:10.1007/s00442-015-3471-7 |
| [61] |
Dueck T A, Elderson J. Influence of NH3 and SO2 on the growth and competitive ability of Arnica montana L. and Viola canina L. New Phytologist, 1992, 122(3): 507-514. DOI:10.1111/j.1469-8137.1992.tb00080.x |
| [62] |
Mitchell C E, Reich P B, Tilman D, Groth J V. Effects of elevated CO2, nitrogen deposition, and decreased species diversity on foliar fungal plant disease. Global Change Biology, 2003, 9(3): 438-451. DOI:10.1046/j.1365-2486.2003.00602.x |
| [63] |
Johnson N C, Rowland D L, Corkidi L, Allen E B. Plant winners and losers during grassland N-eutrophication differ in biomass allocation and mycorrhizas. Ecology, 2008, 89(10): 2868-2878. DOI:10.1890/07-1394.1 |
| [64] |
Zheng Z, Ma P F, Li J, Ren L F, Bai W M, Tian Q Y, Sun W, Zhang W H. Arbuscular mycorrhizal fungal communities associated with two dominant species differ in their responses to long-term nitrogen addition in temperate grasslands. Functional Ecology, 2018, 32(6): 1575-1588. DOI:10.1111/1365-2435.13081 |