 2022, Vol. 42
2022, Vol. 42文章信息
- 闫明, 刘志萍, Mukti RamSubedi, 梁林峰, 奚为民
- YAN Ming, LIU Zhiping, Mukti Ram SUBEDI, LIANG Linfeng, XI Weimin
- 特大干旱对树木死亡的影响——以美国德克萨斯州东部森林为例
- The complex impacts of unprecedented drought on forest tree mortality: A case study of dead trees in east Texas, USA
- 生态学报. 2022, 42(3): 1034-1046
- Acta Ecologica Sinica. 2022, 42(3): 1034-1046
- http://dx.doi.org/10.5846/stxb202101150152
-
文章历史
- 收稿日期: 2021-01-15






2. Departments of Natural Resources Management, Texas Tech University, Lubbock, TX 79409, USA;
3. 山西省林业调查规划院, 太原 030002;
4. Department of Biological and Health Sciences, Texas A & M University-Kingsville, Kingsville, TX 78363, USA
2. Departments of Natural Resources Management, Texas Tech University, Lubbock, TX 79409, USA;
3. Shanxi Forestry Investigation and Planning Institute, Taiyuan 030002, China;
4. Department of Biological and Health Sciences, Texas A & M University-Kingsville, Kingsville TX 78363, USA
树木的死亡是影响森林群落结构动态变化的重要因素[1], 是森林演替和自然更新的驱动力和重要的生态过程之一[2]。树木死亡率的持续变化可以改变森林结构、组成和生态系统服务[3]。造成树木死亡的原因很多, 其中非生物因素主要包括干旱、洪水、极端温度、冰雪灾害等;生物因素主要包括树木的自然衰亡、病虫害、竞争以及人类活动等。树木死亡是一个受环境、病理、生理等因素影响的复杂过程, 同时也受到随机事件的影响[4-5]。通常, 树木死亡都是多种因素共同作用的结果[4]。特大干旱和相关自然干扰(如火灾和病虫害)的影响最大, 在多个时空尺度上影响了生态系统的结构和功能。极端干旱已经引起森林中树木的大量死亡, 并且对森林生态系统的功能和碳平衡也具有深远的影响[6]。森林树木死亡率的增加可能是由于树木对资源可利用性的增加(CO2和光照)导致的竞争加剧, 但其机制仍然未知[7]。极端气候事件在过去几十年以来一直增加, 不同的全球循环模型预测的干旱频率和严重程度以及更具可变性的降水模式也会增加[8], 对森林系统产生较大的不利影响[9]。研究森林对不同时空尺度干旱的响应可为气候变化背景下生态系统的变化提供重要的科学依据[10]。
2011年美国德克萨斯州经历了历史上最严重的一次干旱, 导致全州大约3.01亿株树木死亡。德州东部的森林死亡的树木多达6560万株[11]。Klockow等使用美国森林资源清查与分析(Forest Inventory and Analysis, FIA)数据, 研究了德州东部森林在2011年大旱后的四年各树种的死亡率并对松树死亡率和林分结构的关系进行了专门研究, 模拟了不同林分结构的松树在干旱背景下的死亡率[12]。Subedi等用FIA数据研究了德州东部干旱强度和树木死亡率和生物量损失的关系[13]。伴随着气候变暖, 类似2011年特大干旱会在德州频繁发生。因此搞清特大干旱对德州东部森林影响的中长期效应和复杂性, 对帮助林业管理部门和林地拥有者采取相应的抗旱对策, 十分重要。为了进一步探究干旱对森林树木死亡的影响格局和形成原因, 本文使用20年FIA森林清查数据, 分析调查过四次1797个样地中的209663株树木的死亡率时空变化规律和死亡差异成因, 回答三个问题: (1) 不同干旱强度和干旱长度对德州东部不同区域和类型的树木死亡的影响如何?(2) 特大干旱后德州东部森林树木的年度和调查周期水平树木死亡率对干旱的响应趋势是怎样的?(3) 造成德州东部森林树木死亡的主要原因(干旱和树木之间的竞争作用)的相对贡献率如何?
1 研究区概况美国德州东部林区为研究区(图 1)。德州东部边界约为北纬29°17′—33°57′和西经93°30′—96°27′, 面积约9×106 hm2。地貌不是特别复杂, 东北部地形比较平坦, 最高海拔约200m。土壤有排水良好的淋溶土, 也有排水不良的壤土、粘土等。该区属于亚热带气候, 夏季炎热潮湿, 冬季短暂温和。降水量从东向西逐渐减小, 温度从南向北渐高。年均降水量和年均温度分别在990-1600mm和16-23℃之间变化, 十分有利于森林的生长。

|
| 图 1 美国德克萨斯州东部研究区及森林资源清查与分析(Forest Inventory and Analysis, FIA)样地分布图 Fig. 1 Location map of the study area. Black dots in the main map indicate approximate locations of FIA sample plots |
德州东部林地面积约为4.9×106 hm2。2019年森林覆盖率为54%。优势的森林类型组为火炬松-短叶松(Loblolly-shortleaf pine, 45%), 橡树-胡桃树(Oak-hickory, 23%), 橡树-松树(Oak-pine, 12%)和橡树-北美枫香树-落羽杉(Oak-gum-cypress, 11%)等。森林含多种针叶和阔叶树种, 常见的有火炬松(Pinus taeda)、北美枫香(Liquidambar styraciflua)、星毛栎(Quercus stellata)、短叶松(Pinus echinata)、水栎(Quercus nigra)、南方红橡(Quercus falcata)、翅榆(Ulmus alata)、多花蓝果树(Nyssa sylvatica)、柳叶栎(Quercus phellos)、湿地松(Pinus elliottii)、白橡(Quercus alba)、乌桕(Triadica sebifera)、红枫(Acer rubrum)、美国红梣(Fraxinus pennsylvanica)和糖朴(Celtis laevigata)等。
持续性的炎热和间歇性的干旱是德州东部亚热带半湿润气候重要特征之一。近20年中, 德州就经历了三次干旱(1999-2000、2005-2006、2010-2012年)。2011年前后德州经历历史性的特大干旱, 平均降水量为287mm, 创下连续12个月最干旱的新纪录[14], 是历史上最干旱的年份, 大约比20世纪12个月的平均降水量(1159mm)少47%, 导致树木大面积的死亡[13]。除间歇性的干旱之外, 德州东部还存在多种自然干扰, 包括飓风、林火和病虫害等。该区南部在2005年和2008年分别遭受了两次飓风(Rita和Ike)袭击, 在飓风经过的路线上, 造成森林较大面积的树木受损和死亡[15]。
2 研究方法 2.1 森林数据树木个体、样地数据均来自于FIA项目。该项目从1928年开始, 定期对美国森林进行清查、统计和分析评估。在每2.43×107m2的面积上设置一个样地, 设计如图 2。

|
| 图 2 FIA地设计图 Fig. 2 FIA plot design |
德州东部森林资源清查与分析始于1935年(第一周期), 之后分别在1953-1955年(第二周期), 1965年(第三周期), 1975年(第四周期), 1986年(第五周期)和1992年(第六周期)进行了六轮清查。2000年全美调查标准统一后, 又在2001-2003年(第七周期), 2003-2008年(第八周期), 2008-2013年(第九周期), 2013-2019年(第十周期)共进行了四轮森林清查。2019年德州东部林区中共有样地3836个, 其中森林样地2358个。本文研究近20年德州东部森林变化和干旱影响, 选用2001-2019年调查过四次的森林样地共1797个进行统计分析。样地总面积为48946 hm2, 样地在各轮清查中树木的状态及数量见表 1。
| 第7周期 Cycle 7 |
第8周期 Cycle 8 |
第9周期 Cycle 9 |
第10周期 Cycle 10 |
|
| 活树/(株) Living tree | 47576 | 48266 | 47963 | 48117 |
| 死树/(株) Dead tree | 1571 | 3251 | 6059 | 6860 |
| 移除/(株) Remove tree | 0 | 4825 | 5815 | 6221 |
将样地内树木按照树木种组、胸径、高度、林地起源、纬度、海拔、生态区及地理区域共八个类型进行分类, 统计不同分类标准下树木的状态和数量, 根据公式(1)计算出不同清查周期之间的树木死亡率:
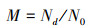
|
(1) |
式中M为周期水平上的树木死亡率, Nd为某个周期死树的数量, N0为上个周期活树的数量。此外, 还计算了周期水平上的年度树木死亡率, 方法同公式(1), 式中M为周期水平上的年度死亡率, Nd为某年死树的数量, N0为上一次调查时活树的数量。
2.2 干旱数据标准化降水蒸散指数(SPEI)作为目前最常用的干旱强度指数, 充分考虑了水分平衡对干旱的影响, 既有对温度敏感的特点, 又具备多尺度、多空间比较的优点, 根据Vicente-serrano等学者, 可根据SPEI值划分成不同的干旱等级(表 2)[16]。
| 极端干旱 Extremely drought |
中等干旱 Moderately dry |
轻度干旱 Mild dry |
正常 Normal |
轻度湿润 Slightly moist |
中等湿润 Moderate humidity |
极端湿润 Extreme humidity |
|
| SPEI | ≤-2.0 | -2.0—-1.0 | -1.0—-0.5 | -0.5-0.5 | 0.5-1.0 | 1.0-2.0 | ≥2.0 |
| SPEI: 标准化降水蒸散指数Standardized precipitation evapotranspiration index | |||||||
选取德州东部42个气象站的40年(1980-2019年)的月度气象数据, 使用温度、降水量和日照时数按公式(2)和(3)计算SPEI值[14]。详细过程参考Vicente-Serrano等[16]。
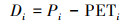
|
(2) |
式中Di为第i个月的累计水分亏缺, Pi为第i个月的降水, PETi为第i个月的潜在蒸散量。
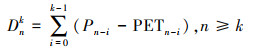
|
(3) |
式中, k为时间尺度(月), n为计算次数。
进而对42个气象站的SPEI值进行插值, 获取1797个样地上的每月SPEI值。统计一年中SPEI≤-1的月份数量, 即为该年度的干旱长度。
2.3 干旱后树木周期水平上不同死因的变化率选择第8周期作为干旱前期状态, 第9周期作为干旱后期状态, 计算不同死因导致树木死亡变化率, 算法见公式(4):
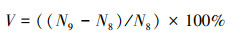
|
(4) |
式中V表示干旱后树木周期水平上不同死因的变化率, N9表示第九周期某死因树木数量, N8表示第八周期某死因树木数量。
2.4 干旱及树木间竞争指标的选择从FIA数据库中进一步提取年度树木信息, 得到每一年各样地的死亡率。干旱指标选择干旱强度(SPEI)、干旱长度(SPEI≤-1的月份数量);同时选择样地树木密度、树木基面积、林地年龄、样地调查间隔时间作为样地中树木之间的竞争因子[3-17], 林地年龄和样地调查间隔时间在FIA数据库获得。树木密度、树木基面积的算法见公式(5)和(6):
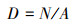
|
(5) |
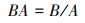
|
(6) |
D为样地中的树木密度, N为单位样地面积上所有树木的个体数, A为样地面积(hm2);BA为树木基面积, B为单位面积上所有树木的胸高处的断面积(m2)。
2.5 树木死亡率年度变化和树木间竞争指标的回归采用广义非线性混合效应模型(GNLM)公式(7),(8)和(9)预测每个样地的树木死亡率pij[18], 进一步使用广义线性模型(GLM)广义线性模型方法进行树木死亡率的年度变化与树木间竞争指标的回归。在树木间竞争指标中, 样地树木死亡率的年度变化=样地树木死亡率/样地平均调查间隔时间, 森林密度=树木株数/样地面积, 树木基面积=所有树木的基面积(DBH)之和/样地面积, 林地年龄和平均调查间隔时间从数据库中获取。
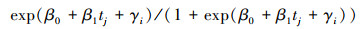
|
(7) |
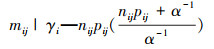
|
(8) |
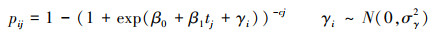
|
(9) |
nij表示第i个样地, 第j次调查的前一次调查的活树数量, mij表示相应周期的死树数量;tj表示第j次调查的时间(年), c代表重复调查间隔时间(年), 随机截距参数(gi)遵循正态分布[3]。
2.6 统计分析Excel 2016进行树木死亡率、样地密度和树木基面积的计算;使用R 4.0.3软件中的SPEI软件包计算SPEI值;One-way ANOVA方法对树木的死亡率进行单因素方差分析, LSD方法对其进行多重比较;使用Stata 15软件对树木死亡率Pij进行预测, 并对标准化后的干旱指标(干旱强度、干旱长度)和树木间的竞争指标(样地密度、树木基面积、林地年龄和调查间隔时间)进行PCA分析, 并进行GLM回归;使用Origin 2018软件作图。
3 结果 3.1 森林树木死亡率在林业调查周期水平上的时空分布特大干旱对树木死亡的影响有一定的滞后效应。干旱发生年份处于第9周期的2011年, 八类中树木第8周期的死亡率最低, 其次是第9周期, 死亡率最高是在第10周期(图 3)。松属树木在各周期的死亡率均为最小, 分别为4.60%、7.92%、8.24%, 表明松属树木相对其他树木更加耐旱;胸径小、高度低的树木在各周期的死亡率均大于相对较大的树木, 胸径小的树木死亡率, 分别为17.10%、26.00%、29.15%, 表明干旱对胸径小、高度较小的个体影响比较明显。干旱对较大树木的影响大于处于中间水平的个体, 胸径最大的个体死亡率在第10周期达到了最大为9.84%;高度最高的个体死亡率居于高度最小的树木之后, 并在9周期达到最大(9.44%)。天然林死亡率高于人工林, 人工林死亡率是在第9周期达到最高(10.26%), 天然林死亡率为13.47%。天然林在第10周期死亡率达到最为16.15%, 说明人工林抗旱能力高于天然林。

|
| 图 3 不同调查周期各种死因树木死亡率 Fig. 3 Mortality of trees of different causes of death in different cycle 横坐标8,9,10分别表示不同周期 |
此外图 3种按照空间区域类型划分结果中显示: 西海岸平原在各周期中的死亡率在第8周期死亡率达到最大(22.27%), 而黑土草原在第10周期死亡率达到最大(25.17%);德州东部的西南地区树木死亡率在后两个周期中均为最高, 死亡率达到最高发生在第10周期(17.59%)。在以纬度和海拔为划分标准时, 树木死亡率最高均发生在第10周期, 但差异不明显, 这是因为德州东部纬度跨度不太大、海拔起伏也不明显。
天气是造成干旱前后不同周期树木死亡数量差异的最主要原因(图 4)。图 4显示导致树木死亡的原因主要有三类: 天气原因、植物竞争和林地清理。在第8周期中, 因天气原因、竞争和植树造林或土地清理活动死亡的树木分别有311, 1210, 1150株;在第9周期中, 因天气原因、竞争和林地清理死亡的树木分别有1523、1774, 1513株;在第10周期中因天气原因、竞争和林地清理活动死亡的树木分别有1419、1796, 1551株。第9周期中天气原因造成的死树量约为第8周期的5倍, 且死树数量在第10周期也达到第8周期的4.5倍。

|
| 图 4 各周期不同死因造成树木死亡数量及干旱后树木周期水平上不同死因的变化率 Fig. 4 Total dead trees in last four FIA cycles grouped by damage agents and the change rate of different death causes at the tree cycle level after drought |
另外在图 4中因天气原因死亡树木的变化率(389.71%)远大于由竞争造成死亡树木的变化率(46.61%), 此外, 由火灾和疾病引起的死树的变化率也较高(400%和108.04%)。由于动物啃食所造成的死树的变化率反而降低(-5.88%), 这可能是因为在干旱年, 动物减少繁殖, 进而使得树木因动物啃食而死的数量减少。
3.2 森林树木死亡率年度水平上的变化趋势年度死亡率分析结果显示特大干旱当年直接导致大量树木死亡。图 5中显示树木的死亡数和死亡率在2011年都达到了最大值, 2012年次之, 2008年、2005年和2013年再次之。这四年致死原因中比例最大的为天气原因, 对应树木死亡率分别为5.74%、4.08%、2.99%、2.73%、2.34%。2012-2013年树木死亡率持续偏高, 为2011年的干旱的滞后效应导致。2008年和2005年的树木死亡率偏高, 则是由于2005年和2008年遭受两次飓风损害引起的。

|
| 图 5 2004-2013年度的死树死亡原因及树木死亡率 Fig. 5 Number of dead trees and mortality causes of dead trees in 2004-2013 不同小写字母表示差异显著(P < 0.05) |
不同死因树木的年度数量统计表明因虫灾和动物啃食造成的树木死亡数量最低;火灾、疾病造成的树木死亡数量略高于前者, 由竞争和天气造成的树木死亡数量最高, 虽然竞争造成的树木死亡数量相对较高, 但在2011年干旱过后, 天气原因造成的树木死亡数量最大, 约是竞争造成的两倍左右。在2011年, 竞争造成627株树木死亡, 因天气原因死亡1081株树。同年, 因火灾和疾病死亡的树木数量显著增加, 分别达到347株和159株, 并且在此后的1-2年间死树数量依然高于干旱前的水平。
3.3 树木死亡差异原因和综合贡献率分析表 3显示德州东部四个地理区域中主要的死亡贡献率为天然林, 尤其是东北部天然林贡献率达到89.22%;四个区域中主要死亡贡献率为小树;其中除其它种贡献率为70.64%, 位居第二的为松属树种, 其贡献率为61.54%, 整体而言, 松属树种死亡率不是太高, 这是因为德州东部松属树种较多;按照林地起源来划分, 无论是天然林还是人工林, 其死亡贡献率最大的都是小树, 分别达到了43.69%和70.64%;天然林和人工林中死亡贡献率情况各不相同, 在天然林中松属和枫香属树种死亡贡献率低于人工林, 其他种则是人工林高于天然林, 可见自然条件下的松属树种和枫香树种死亡数量相对较少;无论物种, 小树死亡贡献率较高, 贡献率由高到低种的顺序为: 松属、枫香、其它树种、白橡、红橡、榆属。
| 地理区域Geographical area | 林地起源Stand origin | 树木大小Tree size | ||||||||||
| 东北部 11.18 % (25.24%) |
东南部 13.01 % (40.20%) |
西北部 12.66 % (17.13%) |
西南部 13.78 % (17.43%) |
天然林 13.47 % (76.73%) |
人工林 10.26 % (23.27%) |
大树 9.63 % (15.71%) |
中树 7.78 % (34.33%) |
幼树 26.00 % (49.96%) |
||||
| 林地起源 Stand origin |
天然林 | 11.08% (71.35%) |
14.55% (70.69%) |
13.11% (86.10%) |
15.67% (89.22%) |
|||||||
| 人工林 | 11.44% (28.65%) |
10.37% (29.31%) |
10.34% (10.42%) |
6.69% (10.78%) |
||||||||
| 树木大小 Tree size |
大树 | 4.68% (8.83%) |
11.33% (16.42%) |
10.15% (16.60%) |
13.00% (23.16%) |
10.46% (18.63%) |
5.36% (6.10%) |
|||||
| 中树 | 6.09% (29.63%) |
7.84% (33.54%) |
8.39% (36.39%) |
9.85% (40.93%) |
10.00% (37.69%) |
3.56% (23.26%) |
||||||
| 幼树 | 27.13% (61.54%) |
25.52% (50.04%) |
24.22% (47.01%) |
27.42% (35.92%) |
23.29% (43.69%) |
29.17% (70.64%) |
||||||
| 树木种组 Tree species group |
松属 7.92 % (27.17%) |
6.24% (23.35%) |
8.42% (32.14%) |
7.73% (16.70) |
9.45% (31.47%) |
10.69% (22.69%) |
5.42% (41.91%) |
6.51% (32.88%) |
4.64% (30.29%) |
29.39% (23.22%) |
||
| 枫香属 15.32 % (14.46%) |
14.93% (21.52%) |
17.07% (11.33%) |
12.19% (14.19%) |
17.95% (11.72) |
13.42% (14.13%) |
26.58% (15.53%) |
6.26% (5.78%) |
9.13% (12.69%) |
28.58% (18.40%) |
|||
| 白橡 18.78 % (18.83%) |
15.49% (16.61%) |
19.87% (18.92%) |
19.41% (21.43%) |
21.03% (19.28%) |
17.62% (20.48%) |
16.27% (13.40%) |
17.63% (28.89%) |
12.96% (16.63%) |
22.28% (17.18%) |
|||
| 红橡 9.90 % (4.87%) |
6.46% (3.20%) |
13.16% (3.37%) |
9.70% (8.49%) |
11.00% (7.18%) |
9.18% (5.53%) |
21.11% (2.70%) |
7.17% (7.35%) |
8.47% (6.49%) |
21.9% (2.97%) |
|||
| 榆属 11.94 % (4.92%) |
12.03% (5.82%) |
13.78% (1.93%) |
10.30% (8.88%) |
13.44% (6.62%) |
10.96% (5.46%) |
24.86% (3.12%) |
12.14% (3.57%) |
6.07% (3.51%) |
18.85% (6.31%) |
|||
| 其他 17.65 % (29.76%) |
17.24% (29.50%) |
17.42% (32.31%) |
17.36% (30.31%) |
19.78% (23.72%) |
15.97% (31.71%) |
33.50% (23.33%) |
14.83% (21.53%) |
13.17% (30.38%) |
23.95% (31.91%) |
|||
| ()内的值为死亡贡献率 | ||||||||||||
为了探究是否样地结构的异质性影响了树木死亡率的增加, 本文使用样地的树木死亡率的年度变化与林地年龄、树木基面积、森林密度和平均调查间隔时间做了广义线性混合效应模型回归(图 6)。结果表明树木死亡率的年度变化与林地年龄、树木基面积、森林密度均不显著相关(P>0.05), 而只与平均调查间隔时间呈现一定正相关(P < 0.05)。这表明虽然不同样地的林地年龄、树木基面积和森林密度存在差异, 但不对树木的死亡率造成显著影响。

|
| 图 6 模拟的德州东部森林树木死亡率的年度变化与林地年龄、树木基面积、森林密度和平均调查间隔时间的关系 Fig. 6 Annual regression of forest sample plots and mortality in cycle 9 of eastern Texas |
对影响森林树木死亡率的六个干旱和竞争指标的主成分分析结果显示, 各指标中第一至三主成分解释贡献率分别为: 28.35%、24.27%、20.24%、累计为72.86%(表 4)。主成分特征值和贡献率是选择主成分的依据。第一主成分特征值为1.701, 方差贡献率为28.356%, 表明干旱长度有较高的正载荷, 干旱强度(SPEI)有较高的负载荷。第一主成分是由干旱强度、干旱长度组成的一个综合指标;第二主成分的特征值为1.456, 方差贡献率为24.266%, 反映数目基面积和树木密度有较高的正载荷, 说明第二主成分可用树木基面积以及树木密度作为一个指标;第三主成分的特征值为1.214, 方差贡献率为20.241%, 表现为林地年龄有较高的正载荷, 树木密度有较高的负载荷, 说明林地年龄和树木密度可作为第三主成分的一个指标。前三个主成分累计方差贡献率为72.863%, 反应了干旱、竞争对树木死亡率影响的72.863%的信息, 综合以上三个主成分的特点, 干旱强度、干旱长度、树木密度、树木基面积和林地年龄可作为判断干旱、竞争对树木死亡率影响的主要指标。在三个主成分中, 从解释贡献率来看, 干旱强度、干旱长度累计解释了28.356%, 这个比例都分别大于另外两个主成分对死亡率的解释, 而且在干旱强度和干旱长度中, 干旱长度载荷与干旱强度载荷(绝对值)相当, 表明了树木死亡主要是由干旱而不是树种间的竞争引起, 干旱长度和干旱强度对树木死亡率的影响大致相同。
| 干旱和竞争指标 Drought and competition items |
主成分Principal Component | ||
| 1 | 2 | 3 | |
| 干旱强度Drought severity | -0.906 | -0.008 | 0.084 |
| 干旱长度Drought length | 0.907 | 0.018 | -0.075 |
| 树木密度Density | -0.118 | 0.657 | -0.648 |
| 树木基面积Basal area | -0.037 | 0.897 | 0.13 |
| 林地年龄Stand age/a | 0.078 | 0.348 | 0.874 |
| 调查间隔时间Remeasurement period/a | 0.192 | 0.313 | 0.023 |
| 特征值Eigenvalues | 1.701 | 1.456 | 1.214 |
| 贡献率Contribution rate/% | 28.356 | 24.266 | 20.241 |
| 累计贡献率Cumulative rate/% | 28.356 | 52.622 | 72.863 |
Gustafson等对美国五大湖区的四个干旱敏感树种群进行了研究, 结果证明了干旱是导致树木死亡的原因之一[17]。Moore等在2011年研究了美国南部德州严重干旱过后的第二年夏天树木死亡情况, 结果表明裸子植物的死亡率(主要是松属树种)低于被子植物[11];Maggard等2016年为了研究持续干旱对树木生长的影响, 选择美国俄克拉荷马州东南部一个种植园中的火炬松作为研究对象, 研究结果表明火炬松在一定程度上具有抵抗干旱胁迫的能力[19]。松属树种为德州东部森林的优势树种, Klockow等2018年以德州东部森林树木为研究对象, 其结果也表明在经历干旱后松属树种的死亡率也低于其他属树种[12]。在本研究中, 处于干旱期的松属树种的死亡率最低, 这与前人的研究结果相同, 但究其原因则是因为德州东部松属树种居多。橡树在德州东部森林中也属于优势树种, 但是对于橡树的研究, Moore等在2016年的研究结果表明白橡树比红橡树更能适应干旱[11]。这与本文的研究结果不同, 本文的研究表明在20年的时间尺度上, 德州东部森林的红橡树的死亡率一直低于白橡树, 这可能是因为在本文的样地中, 红橡树和白橡树的年龄组成、密度不同所导致的。
Moore等2016年对德州东部的自然生长的火炬松和人工种植的火炬松以及全部的短叶松做了研究, 结果表明人工种植的火炬松通常得到集中管理, 自然生长的火炬松和短叶松很少受到管理, 因而种植的火炬树在干旱期间受影响小, 死亡率低于自然生长的火炬松和短叶松[11]。这与本文的研究结果相同, 本文的结果显示在德州东部四个清查周期, 天然林的死亡率都高于人工林的死亡率。Moore等在2011年中研究表明德州东部的东南部树木死亡率为1.3%, 其东北部树木死亡率为3.9%[11], 本研究中德州东部四个地理区域的树木死亡率差异并不十分明显, 但均在11.18%—13.78%之间, 这是由于本研究是对德州东部所有树木包括幼树进行死亡率计算的结果, 由于小树死亡率高, 提高了的树木死亡率。从年度水平上树木死亡率来看, 在2005年、2008年和2011年德州东部发生了特大扰动事件, 在这之后的几年森林树木死亡率依然很高, 说明干旱的会使树木死亡滞后, 这与Edgar等的研究相同[20]。在本文中, 2005年和2008年树木死亡数量也受到飓风的影响, 从而使得2005年和2008年的森林死亡率升高, 与Harcombe等的研究结果一致[15]。
树木在生长过程中会逐渐对外界的干扰具有更强的抵抗力, 所以随着树木胸径的增大, 死亡率会逐渐降低[21]。Lindenmayer等在对全球大型老树的研究中发现, 大型老树很脆弱, 在干旱发生后极易死亡[22]。Mueller等对1996年和2002年发生在美国亚利桑那州北部的干旱进行了研究, 结果表明干旱影响到了较大的松树[23];Nepstad等在亚马逊地区研究了干旱对大树和藤本植物的影响, 表明热带地区死亡的大树的数量比死亡的小树多[24]。董蕾等从生理角度阐述了气候变化背景下碳饥饿与水分代谢、植物死亡关系, 表明干旱会使得植物体内水分代谢与碳代谢会发生失衡现象[25]。这可能是由于树木随个体长大进入老年期, 逐渐衰老, 其生理功能逐渐衰退加之干旱的影响, 导致树木死于“碳饥饿”[26]。本文的研究结果显示在遭受干旱时胸径大、高度高的这类大树的死亡率略高于中等尺度胸径和高度的个体, 但本研究中小树的死亡率最高。原因之一是本文包括了幼树, 死亡率估算偏高。为更好的量化干旱带来的实际后果, 在今后的研究中应加强对森林生物量和碳储量变化的分析。
Peng等2011年对加拿大北部森林进行了研究, 结果显示在海拔在500-1200m的范围间干旱发生后, 海拔是影响树木死亡的一个因素, 并且死亡率随海拔增高而增加[18];同样van Mantgem等2009年在对美国西部树木死亡率的研究中也证明了在1000-2000m的海拔范围间, 树木的死亡率随海拔升高逐渐增大[3]。这与本研究的结果不一致。本文发现在干旱前和干旱期, 海拔低于100m的树木死亡率都大于海拔高于100m的值, 且在干旱期间更为明显。这可能是因为: 低海拔的地方温度较高海拔的地方高, 在温度高的地方干旱效应有所增强造成的;而在干旱后期则表现为海拔100m以上的树木死亡率更高, 这可能是由于干旱的滞后效应导致的。
4.2 树木死亡差异的形成原因和干旱和植物竞争对树木死亡影响的复杂性本研究中, 在2004-2013年不同死因造成的树木死亡数量各不相同, 由虫灾、动物啃食导致树木死亡的数量较少, 而致死数量级较大的原因有天气、竞争、疾病、火灾。Lugo和Oliva等从生理角度研究了病原体与树木的营养关系, 研究结果发现在干旱后, 树木疾病可加速树木的死亡[27-28], 这与本研究的结果一致, 本研究中干旱过后的2011-2013年, 因疾病致死的树大幅增多;此外, 火灾致死的树木在2011年达到了最多, 图 4中显示了因干旱引起火灾和疾病导致树木的死亡急剧增加, 说明在天气干旱时, 火灾的发生频率会增多, 并且造成了树木对疾病的抵抗力下降, 也会导致树木因染病而死亡。朴世龙等的研究中指出干旱会使得火灾频率或强度增加, 造成生态系统碳汇功能的下降[29], 这与本研究的结果一致;本研究图 4中还显示由于动物啃食所造成的死树的变化率在大旱期间降低, 这是因为在遇到干旱年份的时候, 很多动物都会减少繁殖, 进而使得树木因动物啃食而死亡的数量也减少, Martin等的研究针对热带鸣禽的生存率和繁殖率之间的关系进行了研究, 最终总结出了天气干旱会导致动物繁殖率降低、生存率提高的结论, 这项结论同样适用于很多其它动物[30], 这与本研究结果一致。
Laurance等研究了亚马逊的热带森林死亡率, 研究结果显示树种间对资源(如CO2和光照等)的竞争加剧而导致树木死亡率增高[31]。这与本研究的结果不同, 这可能是由于Laurance等的研究对象为亚马逊地区的原始热带雨林, 群落的物种组成复杂, 群落内的树木大小及密度和美国德州东部的亚热带森林群落差异较大造成的。本研究的结果支持van Mantgem等美国西部树木死亡率的增加不是由于竞争作用的加剧造成的, 而可能是由于区域性的干旱和随之而来的水分亏缺造成的结论[3]。在年度水平上, 特大干旱发生的2011年和随后的2012年, 因干旱造成的树木死亡远多于因植物间竞争造成的树木死亡(图 5), 而其他年份中因竞争导致的树木死亡数量较天气原因则略高一些。主成分分析表明干旱对树木死亡率的影响较大, 而竞争因素中的密度对其影响较小, 在干旱期, 树木的死亡率与林地年龄、树木基面积、森林密度均无关系, 仅与平均调查间隔时间相关, 所以干旱比竞争对树木死亡的影响更大。
主成分分析表明干旱长度和干旱强度对树木死亡率的影响基本相当, 与Gustafson等在美国五大湖地区的四个干旱敏感树种的干旱胁迫措施和死亡率之间关系的结论不同。他们发现干旱长度对树木的影响要比干旱强度大[17]。这可能是由于他们研究地区于美国五大湖地区, 对干旱敏感的四个树种的观测时间尺度较短, 而本文针对20年来德州东部林区的多个树种, 两个研究区气候差异很大, 森林类型各异, 而且不同树种耐受干旱的程度也有所不同, 都会导致研究结果有所差异。
5 结论在调查周期水平上, 旱后的树木死亡率明显高于旱前, 有明显的滞后效应, 且存在较显著的树种和区域分异。由于滞后效应, 树木死亡率还表现为在干旱后期死亡率略高于旱中。在年度水平上, 树木在干旱发生的2011年死亡率最高, 干旱之后的2年树木死亡率仍然较高, 可见干旱对树木死亡的直接的和延续影响。干旱发生后, 疾病、火灾等造成树木死亡的数量也明显增多, 说明干旱还带来了一系列的其它干扰因素的间接影响。多种干扰相互叠加和耦合, 进一步造成了树木的延续死亡。干旱相较树木间竞争对树木死亡的影响大, 干旱强度和干旱长度对树木死亡的影响基本相当。树木干旱期间所形成的死亡格局分析表明树木所在地理区域、林地起源、树木大小和物种之间分布情况各不相同, 形成了德州东部森林死亡的复杂格局。本研究所揭示了干旱对德州东部森林影响的中长期延续效应和树木死亡过程的复杂性, 为更准确认识旱后森林生态系统变化提供了科学依据, 有助于该区及邻近区域抗旱措施制定和森林可持续经营管理。
致谢: 感谢山西大学张峰教授对本文数量分析部分提出的重要建议;感谢Texas A & M University-Kingsville对此项研究提供了前期研究经费和大力支持。| [1] |
Lewis S L, Phillips O L, Baker T R, Lloyd J, Malhi Y, Almeida S, Higuchi N, Laurance W F, Neill D A, Silva J N M, Terborgh J, Lezama A T, Martinez R V, Brown S, Chave J, Kuebler C, Vargas P N, Vinceti B. Concerted changes in tropical forest structure and dynamics: evidence from 50 South American long-term plots. Philosophical Transacions of the Royal Society B: Biological Sciences, 2004, 359(1443): 421-436. DOI:10.1098/rstb.2003.1431 |
| [2] |
王利伟, 李步杭, 叶吉, 白雪娇, 原作强, 邢丁亮, 蔺菲, 师帅, 王绪高, 郝占庆. 长白山阔叶红松林树木短期死亡动态. 生物多样性, 2011, 19(2): 260-270. |
| [3] |
van Mantgem P J, Stephenson N L, Byrne J C, Daniels L D, Franklin J F, Fulé P Z, Harmon M E, Larson A J, Smith J M, Taylor A H, Veblen T T. Widespread increase of tree mortality rates in the western United States. Science, 2009, 323(5913): 521-524. DOI:10.1126/science.1165000 |
| [4] |
Franklin J F, Shugart H H, Harmon M E. Tree death as an ecological process. BioScience, 1987, 37(8): 550-556. DOI:10.2307/1310665 |
| [5] |
Yang Y Q, Titus S J, Huang S M. Modeling individual tree mortality for white spruce in Alberta. Ecological Modelling, 2003, 163(3): 209-222. DOI:10.1016/S0304-3800(03)00008-5 |
| [6] |
Choat B, Brodribb T J, Brodersen C R, Duursma R A, López R, Medlyn B E. Triggers of tree mortality under drought. Nature, 2018, 558(7711): 531-539. DOI:10.1038/s41586-018-0240-x |
| [7] |
Van Mantgem P J, Stephenson N L. Apparent climatically induced increase of tree mortality rates in a temperate forest. Ecology Letters, 2007, 10(10): 909-916. DOI:10.1111/j.1461-0248.2007.01080.x |
| [8] |
Klein T, Hartmann H. Climate change drives tree mortality. Science, 2018, 362(6416): 758. |
| [9] |
McDowell N G, Michaletz S T, Bennett K E, Solander K C, Xu C G, Maxwell R M, Middleton R S. Predicting chronic climate-driven disturbances and their mitigation. Trends in Ecology & Evolution, 2018, 33(1): 15-27. |
| [10] |
孔冬冬, 张强, 顾西辉, 王月, 李华贞. 植被对不同时间尺度干旱事件的响应特征及成因分析. 生态学报, 2016, 36(24): 7908-7918. |
| [11] |
Moore G W, Edgar C B, Vogel J G, Washington-Allen R A, March R G, Zehnder R. Tree mortality from an exceptional drought spanning mesic to semiarid ecoregions. Ecological Applications, 2016, 26(2): 602-611. DOI:10.1890/15-0330 |
| [12] |
Klockow P A, Vogel J G, Edgar C B, Moore G W. Lagged mortality among tree species four years after an exceptional drought in east Texas. Ecosphere, 2018, 9(10): e02455. DOI:10.1002/ecs2.2455 |
| [13] |
Subedi M R, Xi W M, Edgar C B, Rideout-Hanzak S, Yan M. Tree mortality and biomass loss in drought-affected forests of East Texas, USA. Journal of Forestry Research, 2021, 32(1): 67-80. DOI:10.1007/s11676-020-01106-w |
| [14] |
Subedi M R, Xi W M, Edgar C B, Rideout-Hanzak S, Hedquist B C. Assessment of geostatistical methods for spatiotemporal analysis of drought patterns in East Texas, USA. Spatial Information Research, 2019, 27(1): 11-21. DOI:10.1007/s41324-018-0216-9 |
| [15] |
Harcombe P A, Leipzig L E M, Elsik I S. Effects of hurricane rita on three long-term forest study plots in east Texas, USA. Wetlands, 2009, 29(1): 88-100. DOI:10.1672/08-64.1 |
| [16] |
Vicente-Serrano S M, Beguería S, López-Moreno J I. A multiscalar drought index sensitive to global warming: the standardized precipitation evapotranspiration index. Journal of Climate, 2010, 23(7): 1696-1718. DOI:10.1175/2009JCLI2909.1 |
| [17] |
Gustafson E J, Sturtevant B R. Modeling forest mortality caused by drought stress: implications for climate change. Ecosystems, 2013, 16(1): 60-74. DOI:10.1007/s10021-012-9596-1 |
| [18] |
Peng C H, Ma Z H, Lei X D, Zhu Q, Chen H, Wang W F, Liu S R, Li W Z, Fang X Q, Zhou X L. A drought-induced pervasive increase in tree mortality across Canada's boreal forests. Nature Climate Change, 2011, 1(9): 467-471. DOI:10.1038/nclimate1293 |
| [19] |
Maggard A, Will R, Wilson D, Meek C. Response of mid-rotation loblolly pine (Pinus taeda L.) physiology and productivity to sustained, moderate drought on the western edge of the range. Forests, 2016, 7(9): 203. |
| [20] |
Edgar C B, Westfall J A, Klockow P A, Vogel J G, Moore G W. Interpreting effects of multiple, large-scale disturbances using national forest inventory data: a case study of standing dead trees in east Texas, USA. Forest Ecology and Management, 2019, 437: 27-40. DOI:10.1016/j.foreco.2019.01.027 |
| [21] |
Blundell A G, Peart D R. Growth strategies of a shade-tolerant tropical tree: the interactive effects of canopy gaps and simulated herbivory. Journal of Ecology, 2001, 89(4): 608-615. DOI:10.1046/j.0022-0477.2001.00581.x |
| [22] |
Lindenmayer D B, Laurance W F, Franklin J F. Global decline in large old trees. Science, 2012, 338(6112): 1305-1306. DOI:10.1126/science.1231070 |
| [23] |
Mueller R C, Scudder C M, Porter M E, Trotter III R T, Gehring C A, Whitham T G. Differential tree mortality in response to severe drought: evidence for long-term vegetation shifts. Journal of Ecology, 2005, 93(6): 1085-1093. DOI:10.1111/j.1365-2745.2005.01042.x |
| [24] |
Nepstad D C, Tohver I M, Ray D, Moutinho P, Cardinot G. Mortality of large trees and lianas following experimental drought in an amazon forest. Ecology, 2007, 88(9): 2259-2269. DOI:10.1890/06-1046.1 |
| [25] |
董蕾, 李吉跃. 植物干旱胁迫下水分代谢、碳饥饿与死亡机理. 生态学报, 2013, 33(18): 5477-5483. |
| [26] |
Lugo A E, Scatena F N. Background and catastrophic tree mortality in tropical moist, wet, and rain forests. Biotropica, 1996, 28(4): 585-599. DOI:10.2307/2389099 |
| [27] |
Oliva J, Stenlid J, Martínez-Vilalta J. The effect of fungal pathogens on the water and carbon economy of trees: implications for drought-induced mortality. New Phytologist, 2014, 203(4): 1028-1035. DOI:10.1111/nph.12857 |
| [28] |
张鹏霞, 叶清, 欧阳芳, 彭龙慧, 刘兴平, 郭跃华, 曾菊平. 气候变暖、干旱加重江西省森林病虫灾害. 生态学报, 2017, 37(2): 639-649. |
| [29] |
朴世龙, 张新平, 陈安平, 刘强, 连旭, 王旭辉, 彭书时, 吴秀臣. 极端气候事件对陆地生态系统碳循环的影响. 中国科学: 地球科学, 2019, 49(9): 1321-1334. |
| [30] |
Martin T E, Mouton J C. Longer-lived tropical songbirds reduce breeding activity as they buffer impacts of drought. Nature Climate Change, 2020, 10(10): 953-958. DOI:10.1038/s41558-020-0864-3 |
| [31] |
Laurance W F, Oliveira A A, Laurance S G, Condit R, Nascimento H E M, Sanchez-Thorin A C, Lovejoy T E, Andrade A, D'Angelo S, Ribeiro J E, Dick C W. Pervasive alteration of tree communities in undisturbed Amazonian forests. Nature, 2004, 428(6979): 171-175. DOI:10.1038/nature02383 |