 2022, Vol. 42
2022, Vol. 42文章信息
- 吕同汝, 蒋勇军, 吴泽, 茆杨, 邱菊, 吴超
- LÜ Tongru, JIANG Yongjun, WU Ze, MAO Yang, QIU Ju, WU Chao
- 亚热带岩溶区典型常绿和落叶树种的蒸腾特征及其对环境因子的响应
- Transpiration characteristics of typical evergreen and deciduous tree species in subtropical karst areas and their responses to environmental factors
- 生态学报. 2022, 42(3): 1047-1058
- Acta Ecologica Sinica. 2022, 42(3): 1047-1058
- http://dx.doi.org/10.5846/stxb202101020006
-
文章历史
- 收稿日期: 2021-01-02
- 网络出版日期: 2021-10-09






植物蒸腾是水分通过植物体从土壤转移到大气的过程[1], 它受太阳辐射、气温、相对湿度和风速等蒸腾驱动因子的推动, 也受土壤对植物供水能力的影响[2]。理解蒸腾的影响机制并予以量化, 是研究区域植被耗水情况和水量平衡动态变化的重要基础[3]。亚热带岩溶区虽水热条件较好, 但旱、雨季分明, 75%—85%的降水主要集中在雨季[4-5]。另外由于岩溶区的土层浅薄且不连续, 土壤持水能力较低, 植物常受到水分胁迫的影响(尤其是在旱季)[6]。然而, 岩溶区适生树种往往能够吸收基岩裂隙或表层岩溶带的水分, 通过灵活的水分利用策略来应对水分胁迫, 并保持着一定水平的蒸腾量来维持自身的生存和生长[7]。因此, 研究亚热带岩溶区植物蒸腾特征有助于理解植物水分利用机制[8], 对进一步分析植物在岩溶生态系统水循环中的作用具有重要意义。
当前, 亚热带岩溶区关于植物蒸腾特征的研究主要以常绿树种为研究对象, 如黄玉清等[4]、吴鹏等[8]和李成龙等[9]分别对青冈栎(Cyclobalanopsis glauca)、小果润楠(Machilus microcarpa)和天峨槭(Acer wangchii)的树干液流和蒸腾特征进行了分析, 发现其研究样树的日蒸腾量表现为夏季高冬春低的特征, 且日变化过程呈现典型的单峰型曲线。亚热带岩溶区森林植被以常绿树种为主, 但也生长着部分落叶树种。如落叶树种刺槐, 它作为世界上引种最成功的三大树种之一, 适应能力强, 耐贫瘠和干旱[10]。不同叶片习性的树种在气孔导度、储水能力、导水能力和木材密度等方面存在差异[11], 而植物的生理特征差异会对其蒸腾能力和水分利用策略产生重要影响。生长在亚热带岩溶区的落叶树种, 其蒸腾特征及对环境因子的响应是否有别于乡土常绿树种, 对此尚缺乏足够认识。因此, 有必要对两种不同生活型树种的蒸腾特征及其影响因子进行对比分析, 以更好地了解亚热带岩溶区内常绿和落叶树种的蒸腾耗水规律及其对环境因子的响应差异。
基于此, 本研究以重庆中梁山岩溶槽谷内次生林为研究对象, 利用热耗散技术连续监测适生常绿树种女贞(Ligustrum lucidum)和落叶树种刺槐(Robinia pseudoacacia)的树干液流, 分析不同生活型树种的蒸腾特征及其对气象因子和土壤水分变化的响应。旨在探讨: (1)在亚热带岩溶区, 不同时间尺度下, 影响典型常绿和落叶树种整树蒸腾量主要有哪些环境因子;(2)常绿和落叶树种的蒸腾特征是否存在差异及其原因。以揭示亚热带岩溶区树木的蒸腾耗水规律, 为脆弱生态系统的植被恢复与造林树种的选择提供理论依据。
1 材料与方法 1.1 研究区概况研究区(图 1)位于重庆市北碚区中梁山岩溶槽谷(106°23′15″—106°28′05″E, 29°40′30″—29°48′10″N), 属于亚热带季风气候区, 夏季炎热, 冬季温和, 年均温达18 ℃;年降水量达1000-1200 mm, 年内分配不均匀, 有明显的旱雨季之分, 4-9月降雨量占全年降水量的75%左右。中梁山总体由东北-西南向的三岭两槽构成, 其山岭下覆抗侵蚀能力强的砂岩和页岩(下三叠统飞仙关组: T1f和上三叠统须家河组: T3xj), 槽谷下覆易被侵蚀的石灰岩和白云岩(下三叠统嘉陵江组: T1j和中三叠统雷口坡组: T2l)[12]。研究区地表多裸露碳酸盐岩, 土壤主要为嘉陵江组发育而来的石灰土, 土层薄且厚度不均, 土层厚度约5-50 cm, 主要乔木树种为女贞、刺槐和香樟(Cinnamomum camphora)等。

|
| 图 1 研究区位置、水文地质、地质剖面(改自Liu等[5])以及树干液流监测图 Fig. 1 Study area location, hydrogeology, geological profile (modified from Liu et al. [5]), and sap flow monitoring pictures T1f: 下三叠统飞仙关组Feixianguan formation of lower triassic;T1j: 下三叠统嘉陵江组Jialingjiang formation of lower triassic;T2l: 中三叠统雷口坡组Leikoupo formation of middle triassic;T3xj: 上三叠统须家河组Xujiahe formation of upper triassic |
在研究区内随机选取径级适中, 茎叶健康的优势树种女贞、刺槐各3棵作为监测样树, 监测样树基本特征见表 1。于2018年7月至2019年5月运用热耗散探针法(SF-G探针式植物茎流测量系统, 德国Ecomatik公司)对选定的女贞和刺槐样树进行树干液流速率监测。根据两树种边材厚度和探针规格, 本研究中3棵女贞均采用5 cm长度探针, 3棵刺槐均采用3 cm长度探针。探针垂直插入树干北面(以避免阳光直射)距地面130 cm高度位置, 探针间隔10-15 cm。上探针为加热探针, 下探针为参比探针, 根据恒定功率下(0.2 W)探针间的温差, 计算液流速率, 每15 min记录一次数据。传感器用铝箔缠裹, 以防止光照和雨水等外界因素干扰。根据Granier建立的经验公式[13]计算液流速率:
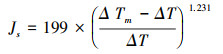
|
(1) |
| 树种 Species |
样树序号 Sample tree No. |
树高 Height/m |
胸径 Diameter at breast height/cm |
边材厚度 Sapwood thickness/cm |
边材面积 Sapwood area/cm2 |
| 女贞L. lucidum | 1 | 6.4 | 15.9 | 5.4 | 177.3 |
| 2 | 8.5 | 16.6 | 4.8 | 171.9 | |
| 3 | 7.3 | 15.8 | 4.8 | 166.8 | |
| 刺槐R. pseudoacacia | 4 | 9.5 | 22.6 | 3.9 | 220.5 |
| 5 | 11.2 | 25.5 | 3.6 | 239.7 | |
| 6 | 9.0 | 19.6 | 2.9 | 153.4 |
式中, Js(g m-2s-1)为液流速率;ΔT(℃)为上下探针间温差;ΔTm(℃)为液流速率接近0时, 上下探针间的最大温差。
运用生长锥钻取了监测样树的树芯, 将其带回实验室用水浸泡, 至清晰区别边材和心材的边界;然后用游标卡尺量取心材和树皮厚度, 根据下式求得边材面积As(cm2):
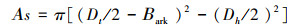
|
(2) |
式中, Dt(cm)为整个树干的宽度(整根树芯的长度), Dh(cm)为心材宽度;Bark(cm)为树皮厚度。
整树蒸腾量(ET, kg/h)通常由液流速率与边材面积的乘积而得[7]:
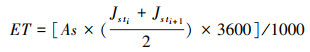
|
(3) |
式中, Jst i、Jst i+1(g m-2s-1)为i时刻和i+1时刻的液流速率;3600为由秒到小时的时间转化系数;1000为克到千克的转换系数。
通过对女贞和刺槐物候的调查和分析[14-15], 可知4月处于两树种展叶期, 女贞新旧叶更替, 刺槐新叶展叶;7月为两树种生长盛期;11月女贞冠层仍较茂密, 刺槐部分叶片脱落;2月女贞部分落叶, 刺槐叶片已几乎落尽。因此本研究选取2018年7月和11月、2019年2月和4月(分别代表夏、秋、冬和春4个季节), 每月5日至24日的整树蒸腾量和环境因子进行监测和分析。
1.3 环境因子的监测和获取 1.3.1 气象数据的获取气象数据来自研究区附近的自动气象监测站(DAVIS Vantage Pro2型, 天津气象仪器公司), 实时监测气象要素为气温(T, ℃)、降水量(P, mm)、太阳辐射强度(Rs, W/m2)、相对湿度(RH, %)和风速(Ws, m/s)等。饱和水汽压亏缺(VPD, kPa)由T和RH通过经验公式[16]计算得出:
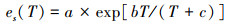
|
(4) |
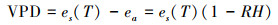
|
(5) |
式中, es(T)代表T温度下的饱和水汽压(kPa), a、b、c为参数, 分别取值为: 0.611、17.502和240.97, T为温度(℃), ea为实际水汽压(kPa), RH为相对湿度(%)。
1.3.2 土壤含水率(SMC, %)测定采用数据采集器(RR—1016型, 北京雨根科技有限公司), 分别在0-20 cm和20-40 cm土壤深度(SMC20和SMC40)安装相应的传感器(AV-EC5), 量程和精度分别为0-100%和± 0.1%, 每15 min记录一次数据。
1.3.3 根系分布调查在研究区内选取与监测样树胸径、冠幅相当, 所处坡度、土层厚度相近的女贞和刺槐, 自树干基部的主根向四周小心挖掘, 保留须根, 完整根系用白色胶布标记, 并用卷尺测量根系深度。
1.4 数据分析本文使用Excel 2010对数据进行初步处理。昼夜的划分依据为太阳辐射强度大小, 晴、雨天的划分依据为日照时数和降雨量。采用SPSS 26.0对整树蒸腾量和各环境因子进行皮尔逊相关分析, 采用Origin 2019完成作图。
2 结果与分析 2.1 不同时间尺度下树木蒸腾量与环境因子的变化特征 2.1.1 蒸腾量和环境因子的季节变化特征女贞与刺槐的整树蒸腾量(ET)表现出明显的季节差异, 春夏季ET显著高于秋冬季ET(图 2)。各季节ET均值如下: 女贞表现为夏季(1.29 kg/h)>春季(0.57 kg/h)>冬季(0.15 kg/h)>秋季(0.13 kg/h), 刺槐表现为夏季(0.90 kg/h)>春季(0.31 kg/h)>秋季(0.16 kg/h)>冬季(0.04 kg/h)。

|
| 图 2 树木蒸腾量及其环境影响因子的季节差异 Fig. 2 Seasonal differences of tree transpiration and environmental factors SMC20: 0-20 cm土壤含水率0-20 cm soil moisture content;SMC40: 20-40 cm土壤含水率20-40 cm soil moisture content |
气象因子和土壤含水率(SMC)也存在明显的季节差异(图 2)。春夏季的Rs、T、VPD和P均显著高于秋冬季;Ws的季节差异不大, 春夏季略高于秋冬季。四个季节中, 春夏季SMC高于秋冬季SMC, 且一年中SMC40均高于SMC20。SMC的变化与降雨事件关系密切, 降雨后SMC明显增加, 尤其是强降雨事件发生时, SMC表现为迅速增长, 而后逐渐减小;SMC20比SMC40对降雨的响应速度更快, 也更为敏感。
2.1.2 晴雨天蒸腾量与环境因子的变化特征对比典型晴天(7月20日, 2月5日)和雨天(7月7日, 2月10日)两树种的蒸腾量和环境因子的日变化特征(图 3)发现: 在晴天, 两树种的蒸腾量与各气象因子出现明显单峰日变化, 且它们几乎同时上升, 其中T和VPD的峰值出现时间一般滞后于Rs (3.5±0.5) h。ET在7月(雨季)的日变化曲线表现为宽峰型, 当VPD和T等因子升高到一定数值后, ET便维持在一个较高的稳定状态;而ET在2月(旱季)的日变化曲线存在树种上的差异, 女贞蒸腾量(ETL)表现出明显的单峰日变化, 而刺槐蒸腾量(ETR)日内起伏较小。

|
| 图 3 典型晴雨天的树木蒸腾量与环境因子的日变化 Fig. 3 Diurnal variation of tree transpiration and environmental factors in typical sunny and rainy days |
与晴天相比, 雨天T、Rs和VPD的日变化幅度显著减小, Ws的日内变化没有明显规律。降雨事件造成了T、Rs和VPD形成一段显著的低值, 雨后各因子的值有所升高。同时, 降雨活动使得SMC小幅度升高(2月10日雨量有限, 仅对较浅表层土壤水分有影响)。两树种对降雨的蒸腾响应呈现相反的变化特征: ETL受到降雨抑制, 在雨后0.5-1 h开始上升, 而ETR受到降雨的促进作用, 在降雨发生时快速升高。
2.1.3 蒸腾量和环境因子的昼夜变化特征为了解白天和夜间两树种蒸腾耗水量的差异, 我们定义Rs=0 W/m2为夜间, 计算白天和夜间蒸腾量及夜间蒸腾量占全天蒸腾量的比值(图 4)。结果表明: 两树种的夜间蒸腾量占日蒸腾总量35%以下, 研究期内女贞的单日夜间蒸腾累积量平均为1.08 kg, 刺槐的累积量高于女贞, 为1.56 kg。夜间蒸腾量占比存在季节差异, 女贞表现为秋(19.45%)>冬(17.39%)>春(13.01%)>夏(5.63%), 刺槐表现为冬(33.91%)>秋(27.32%)>春(21.45%)>夏(13.70%)。由此可见, 在蒸腾量小的秋冬季, 两种树的夜间蒸腾量占比均明显高于蒸腾量大的夏季。

|
| 图 4 树木昼夜蒸腾量及夜间蒸腾量占比 Fig. 4 Day and night transpiration of trees and the proportion of night transpiration ETL: 女贞蒸腾量Transpiration of L.lucidum;ETR: 刺槐蒸腾量Transpiration of R.pseudoacacia |
通过分析各季节夜间降雨量占日降雨量的比重, 可以看出, 除夏季外, 研究区各季节夜雨量占比均达60%—70%, 存在典型的夜雨现象。同时, 夜雨量占比与女贞和刺槐的夜间蒸腾量占比的季节变化规律具有一致性, 且各季节刺槐夜间蒸腾量占比均高于女贞(图 5)。

|
| 图 5 夜雨量占比和夜间蒸腾量占比的季节变化 Fig. 5 The proportion of night rainfall and night transpiration in different seasons |
将各季节女贞和刺槐的ET与T、P、Rs和SMC20等环境因子进行相关分析(表 2), 结果表明: 在四个季节里, Rs、T和VPD是影响女贞蒸腾量的主要因子, SMC20和SMC40与ETL的相关性较低;而对于刺槐而言, 春夏季蒸腾量的主要影响因子与女贞一致, 为Rs、T和VPD, 而在秋冬季, 以上3个因子对其影响减弱, SMC20和SMC40成为ETR的主要影响因子。
| 蒸腾量 ET |
季节 Season |
太阳辐 射强度 Rs |
气温 T |
水汽压亏缺 VPD |
风速 Ws |
降水量 P |
0-20 cm 土壤含水率 SMC20 |
20-40 cm 土壤含水率 SMC40 |
| 女贞蒸腾量ETL | 春季 | 0.925** | 0.653** | 0.638** | 0.274** | -0.110** | -0.122** | 0.114** |
| 夏季 | 0.873** | 0.651** | 0.609** | 0.459** | -0.072** | -0.155** | -0.005 | |
| 秋季 | 0.826** | 0.391** | 0.687** | 0.247** | 0.002 | 0.115** | 0.110** | |
| 冬季 | 0.842** | 0.646** | 0.712** | 0.358** | -0.043 | 0.157** | 0.059** | |
| 刺槐蒸腾量ETR | 春季 | 0.489** | 0.429** | 0.306** | 0.173** | 0.221** | 0.417** | 0.276** |
| 夏季 | 0.689** | 0.728** | 0.696** | 0.509** | -0.042 | -0.348** | -0.096** | |
| 秋季 | -0.046* | -0.351** | 0.152** | -0.007 | 0.208** | 0.636** | 0.457** | |
| 冬季 | 0.073** | -0.238** | -0.211** | -0.054* | -0.017 | 0.421** | 0.598** | |
| ET: 蒸腾量Transpiration;ETL: 女贞蒸腾量Transpiration of L.lucidum;ETR: 刺槐蒸腾量Transpiration of R.pseudoacacia;Rs: 太阳辐射强度Solar radiation intensity;T: 气温Air temperature;VPD: 水汽压亏缺Vapor pressure deficiency;Ws: 风速Wind speed;P: 降雨量Rainfall;SMC20: 0-20 cm土壤含水率0-20 cm soil moisture content;SMC40: 20-40 cm土壤含水率20-40 cm soil moisture content;** P < 0.01, * P < 0.05 | ||||||||
将研究期内的典型晴天和雨天整树蒸腾量和各环境因子进行相关分析(表 3), 结果表明: 在晴天, Rs、T、VPD仍是影响ET的主要因子, Ws对ET的影响也较大。SMC20与ET呈显著负相关, 一定程度上限制了树木蒸腾的进行。而在雨天, ETL与ETR和环境因子的相关性具有明显差异。就女贞而言, 其蒸腾量的主要影响因子与晴天相同, 为Rs、VPD和T, 上、下层土壤含水率对其影响也较大;然而, 刺槐蒸腾量主要影响因子为SMC20和SMC40, Rs、VPD、T和Ws对ETR的影响较小。
| 类型 Types |
蒸腾量 ET |
太阳辐射 强度Rs |
气温 T |
水汽压 亏缺 VPD |
风速 Ws |
降水量 P |
0-20 cm 土壤含水率 SMC20 |
20-40 cm 土壤含水率 SMC40 |
| 晴天Clear day | ETL | 0.806** | 0.678** | 0.706** | 0.536** | - | -0.311** | 0.085** |
| ETR | 0.465** | 0.836** | 0.768** | 0.393** | - | -0.564** | -0.013 | |
| 雨天Rainy day | ETL | 0.854** | 0.572** | 0.799** | 0.012 | -0.075** | 0.254** | 0.281** |
| ETR | 0.361** | 0.316** | 0.256** | 0.018 | 0.189** | 0.560** | 0.443** | |
| 白天Daytime | ETL | 0.809** | 0.788** | 0.754** | 0.416** | -0.059** | 0.139** | 0.424** |
| ETR | 0.556** | 0.819** | 0.739** | 0.369** | 0.034* | 0.213** | 0.447** | |
| 夜间Nighttime | ETL | 0.008 | 0.661** | 0.689** | 0.099** | -0.034 | 0.201** | 0.290** |
| ETR | 0.039 | 0.590** | 0.519** | 0.135** | 0.143** | 0.279** | 0.320** | |
| ** P < 0.01, * P < 0.05 | ||||||||
由于环境因子和植物自身生理特性的昼夜变化, 环境因子在白天和夜间对整树蒸腾量的影响存在差异(表 3)。白天太阳辐射强, 气温高, 相应地Rs、T和VPD也对ET有着重要影响, 且Ws也显著影响着ET;在夜间, 太阳辐射缺失, 气温较低, Rs对ET几乎没有影响, 同时T、VPD和Ws对ET的影响减小, 但VPD和T仍是影响ET的两大主要因子。不论昼夜, ETL均与P呈负相关, 而ETR与P呈正相关, 两树种对降雨活动表现出完全相反的响应;SMC与ET有较大相关性, 且ETR对SMC的相关程度高于ETL。
2.3 女贞和刺槐的叶片和根系特征根系挖掘结果表明(图 6): 两树种的根系均在岩石裂隙中发育, 部分须根盘覆在岩石表面。女贞的根系表现为纵向根系多于横向根系, 在0-50 cm深度内, 较粗的主根发达, 而须根较少, 在50 cm附近须根逐渐增多。而刺槐的根系多分布于0-35 cm土层深度, 且表层须根较多。从图 6左上角的叶片照片可以看出, 女贞的叶片宽大肥厚, 而刺槐叶片小且薄。

|
| 图 6 女贞(左)和刺槐(右)的叶片性状与根系分布 Fig. 6 Leaf characteristics and root distribution of L.lucidum(left) and R.pseudoacacia(right) (左上角为对应树种的叶片照片) (the upper left corner is the leaf photo of the corresponding tree species) |
植物蒸腾受蒸腾驱动因子和土壤供水的综合影响, 蒸腾驱动因子包括太阳辐射、空气温度、湿度和风速, 它们驱动着蒸腾过程;土壤对植物的供水能力主要通过土壤含水率来体现[14, 17-18]。在季节尺度下, 太阳辐射强度、气温和水汽压亏缺是影响树木蒸腾量的主要因子(表 2), 这与多数人的研究结果一致[7, 19-20];相较于秋冬季, 树木蒸腾量在春夏季对环境因子的响应更为显著, 两树种春夏季ET明显高于秋冬季ET。此结果与李成龙等[9]的研究结果一致, 这可能是因为春夏季Rs、T和VPD等较大, 蒸腾驱动强烈, ET对环境因子变化能迅速产生响应, 而秋冬季蒸腾需求较小, ET对环境因子的响应较弱。
在晴天, T、VPD和Rs显著驱动着ET发生变化, 同时风进一步加快了气孔的水分损失, 但由于VPD和气孔导度有着高度关联性, 当T达到高值, 且VPD>1.5 kPa后, 气孔控制将会增强, 以减少过多的水分消耗[11], 所以ET在日出后明显增长, 并在午间前后维持在一个较高的稳定状态, 午后则随Rs和VPD的降低, ET逐渐下降。而在雨天, 蒸腾驱动因子弱化, 湿润的环境使得叶片气孔导度较低[8], 而此时土壤水分供应充足, 但由于两树种的生理差异, 使得它们对环境因子的响应差异较大, 在降雨时, 女贞表现为蒸腾活动受到抑制, 而刺槐蒸腾量迅速增加(图 3)(具体的不同树种蒸腾情况对比见下节)。
就昼夜差别看, 植物蒸腾在白天主要受Rs、T和VPD的影响, 而夜间则几乎不受这类因子影响[20]。在本研究中, 因白天蒸腾需求强烈, 且Rs、T和VPD与ET的相关性较强, 所以蒸腾量明显高于夜间;在夜间, Rs和T等蒸腾驱动因子弱化甚至缺失, 而SWC成为影响夜间蒸腾的重要因子, 这与王艳兵等[21]的结果一致, 即夜间蒸腾很大程度上取决于土壤的补水能力, 所以亚热带部分树种在夜间能维持着低水平的蒸腾。另外, 夜间蒸腾量占比存在季节差异, 秋冬季明显高于春夏季(图 4), 这是因为两树种在秋冬季夜间仍进行着一定量的蒸腾, 但日蒸腾总量本身较小, 所以夜间蒸腾量占日蒸腾总量比重较大, 这与张婕等[22]对元宝枫(Acer truncatum)的夜间液流研究结果一致。
3.2 亚热带岩溶区典型常绿和落叶树种的蒸腾特征差异亚热带岩溶区的树种主要为常绿树种, 如女贞、青冈栎等, 也生长着部分落叶树种, 如刺槐、朴树(Celtis sinensis)等[5, 7, 23]。常绿和落叶树种在形态、生理等方面存在差异。形态差别一方面体现在叶片的物候变化及叶片性状上[11, 24], 亚热带常绿树种通常叶面积指数较高[25], 叶片厚度较大, 而落叶树种多在秋冬季节发生落叶, 以减少水分和养分消耗;另一方面, 形态差别体现在根系展布和根深上。本研究中, 女贞叶片大且肥厚, 叶面气孔发达, 利于水分蒸腾;其根系表现为二态分布, 多条主根延伸至基岩以下, 即表层根系从土壤层中吸收水分, 在土壤干旱时, 深层根系又能吸收表层岩溶带或地下水分[5]。Peng等[26]利用稳定同位素探究岩溶区女贞水分来源, 也发现女贞在雨季有60.8%的水分来源于土壤水, 而在旱季水分来源中, 土壤水仅占比34.6%, 而基岩裂隙水占比44.7%, 在旱季, 女贞能利用裂隙水维持其生长。因此, 在冬季, 女贞仍能维持一定水平的蒸腾量。然而, 刺槐叶片小且薄, 单叶蒸腾量有限, 同时表层根系发达, 须根众多(图 6), 而植物生理吸水主要通过须根进行[27], 其对表层土壤水的依赖更大。因而, 在环境因子相同的背景下, ETL在各季节中普遍高于ETR, 尤其在冬季, 刺槐的蒸腾量极低, 此时刺槐蒸腾水源主要依赖上、下两层土壤水分(表 2), 且推测此阶段耗水主要通过枝条或树干直接蒸发[28]。对生长在半干旱黄土丘陵区的刺槐进行研究也发现其吸水能力较强的有效根系主要分布在20-30 cm土层, 落叶期后, 刺槐的液流速率明显降低[14], 和我们在岩溶区得出的规律基本一致。值得注意的是, 秋季ETR略高于ETL, 这是因为在11月5日—7日(总降雨量22.5 mm)和11月16-18日(总降雨量7.1 mm)发生了较大降水事件, 导致刺槐在这几日的蒸腾量明显升高。在11月7日, ETR甚至达到ETL的3倍以上, 而此时期女贞的蒸腾作用却受到一定程度的抑制。同时因本研究中各季节的蒸腾量取用平均值, 所以出现秋季刺槐蒸腾量略高于女贞蒸腾量的情况。
有研究表明, 落叶树种因细胞壁缺乏弹性, 所以比常绿树种对土壤干旱有更大的气孔敏感性, 能更快地对土壤水分变化做出反应[11, 29]。在本研究中, 降雨活动发生时, 土壤含水率迅速升高, Rs、T和VPD等蒸腾驱动因子的影响弱化, 但上、下层土壤含水率对蒸腾量的影响却显著提高(表 3), 这可能是因为刺槐在雨天能通过降低叶片水势增强水分吸收, 更易受到土壤水分变化的影响, 且根系横向展布的刺槐在吸收表层土壤水分方面具有更大的优势[30-31], 导致雨天整树蒸腾量快速上升(图 3)。而对于女贞而言, 虽叶片气孔发达, 但气孔导度与VPD有很强的正相关关系, 在VPD较低的雨天(VPD < 1 kPa), 其叶片气孔关闭[32], 蒸腾需求显著减小, 以至于雨天女贞蒸腾量表现为较低的稳定状态。
判定夜间液流是用于蒸腾还是茎干补水, 重要依据便是VPD的作用大小[33-34]。因为本研究中的蒸腾量是由树干液流速率进一步求得, 所以ET-VPD的关系一定程度上也能反映Js-VPD的情况。在夜间, VPD与两树种蒸腾量呈显著相关(表 3), 可知亚热带岩溶区女贞和刺槐的夜间液流均主要用于蒸腾。通过分析四个季节夜间降雨量占比情况, 发现研究区存在典型的夜雨现象[35], 夜雨量占比与女贞和刺槐夜间蒸腾量占比的季节变化规律具有一致性。另外, ETR在夜间与降雨量相关性显著, 刺槐对降雨事件也具有快速响应的特征, 导致刺槐在各季节的夜间蒸腾量及其占比高于女贞(图 4, 5)。由此可知, 在蒸腾驱动因子较弱的夜晚, 相较于女贞, 刺槐吸收水分、促进蒸腾的能力更强[29]。但本研究主要针对女贞和刺槐两个树种, 而同一生活型下不同树种的边材面积、导水能力等特征仍存在差别。因此, 在未来的研究中, 还需将更多树种纳入到研究中, 以更好地评估亚热带岩溶区适生树种的蒸腾耗水特征, 从而为植被恢复和造林树种的选择提供合理建议。
4 结论(1) 不同时间尺度下, 亚热带岩溶区内女贞和刺槐的整树蒸腾量变化特征及其影响因子存在明显差异。在各季节中, Rs、T和VPD显著影响着ET, 女贞蒸腾量表现为夏季(1.29 kg/h)>春季(0.57 kg/h)>冬季(0.15 kg/h)>秋季(0.13 kg/h), 刺槐蒸腾量表现为夏季(0.90 kg/h)>春季(0.31 kg/h)>秋季(0.16 kg/h)>冬季(0.04 kg/h)。在晴天, 两树种整树蒸腾量的主要影响因子为T、VPD和Rs, ET存在明显单峰日变化特征, 当T达到高值, 且VPD>1.5 kPa后, 气孔控制将会增强, 蒸腾速率逐渐减小;而雨天整树蒸腾量更多地受到SMC20和SMC40的影响。白天整树蒸腾量与Rs、T和VPD的相关性较强, 且蒸腾量显著高于夜间。秋冬季夜间蒸腾量占比明显高于春夏季。
(2) 亚热带岩溶区常绿树种女贞和落叶树种刺槐的蒸腾耗水特征存在显著差异。在相同的环境条件下, ETL在各季节中普遍高于ETR。由于常绿和落叶树种的生理特征不同, 在降雨时女贞叶片气孔关闭, ETL受到抑制;而刺槐能通过降低叶片水势增强水分吸收, 同时其表层根系发达, 便于快速吸收土壤水分, ETR显著提升。刺槐对降雨事件的敏感性和多夜雨的区域气候相联系, 造成刺槐的夜间蒸腾量和夜间蒸腾占比高于女贞, 说明在蒸腾驱动因子较弱的雨天和夜间, 刺槐的水分利用能力更强。
| [1] |
Di N, Xi B Y, Clothier B, Wang Y, Li G D, Jia L M. Diurnal and nocturnal transpiration behaviors and their responses to groundwater-table fluctuations and meteorological factors of Populus tomentosa in the North China Plain. Forest Ecology and Management, 2019, 448: 445-456. DOI:10.1016/j.foreco.2019.06.009 |
| [2] |
Jiao L, Lu N, Fang W W, Li Z S, Wang J, Jin Z. Determining the independent impact of soil water on forest transpiration: a case study of a black locust plantation in the Loess Plateau, China. Journal of Hydrology, 2019, 572: 671-681. DOI:10.1016/j.jhydrol.2019.03.045 |
| [3] |
曹恭祥, 王云霓, 郭中, 季蒙, 王彦辉, 徐丽宏. 六盘山南侧华北落叶松人工林蒸腾对土壤水分和潜在蒸散的响应. 应用生态学报, 2020, 31(10): 3376-3384. |
| [4] |
黄玉清, 张中峰, 何成新, 赵平, 袁维园, 焦继飞, 尤业明. 岩溶区青冈栎整树蒸腾的季节变化. 应用生态学报, 2009, 20(2): 256-264. |
| [5] |
Liu J C, Shen L C, Wang Z X, Duan S H, Wu W, Peng X Y, Wu C, Jiang Y J. Response of plants water uptake patterns to tunnels excavation based on stable isotopes in a karst trough valley. Journal of Hydrology, 2019, 571: 485-493. DOI:10.1016/j.jhydrol.2019.01.073 |
| [6] |
Liu M X, Xu X L, Wang D B, Sun A Y, Wang K L. Karst catchments exhibited higher degradation stress from climate change than the non-karst catchments in southwest China: an ecohydrological perspective. Journal of Hydrology, 2016, 535: 173-180. DOI:10.1016/j.jhydrol.2016.01.033 |
| [7] |
Huang Y Q, Li X K, Zhang Z F, He C X, Zhao P, You Y M, Mo L. Seasonal changes in Cyclobalanopsis glauca transpiration and canopy stomatal conductance and their dependence on subterranean water and climatic factors in rocky karst terrain. Journal of Hydrology, 2011, 402(1/2): 135-143. |
| [8] |
吴鹏, 杨文斌, 崔迎春, 赵文君, 舒德远, 侯贻菊, 丁访军. 喀斯特区天峨槭(Acer wangchii)树干液流特征及其与环境因子的相关分析. 生态学报, 2017, 37(22): 7552-7567. |
| [9] |
李成龙, 刘延惠, 丁访军, 舒德远, 崔迎春, 赵文君, 侯贻菊, 吴鹏. 茂兰喀斯特森林小果润楠蒸腾特征及影响因素. 南京林业大学学报: 自然科学版, 2019, 43(3): 51-58. |
| [10] |
张蒙蒙. 干旱胁迫下3个刺槐类型植物叶脉网络结构与光合特性的变化[D]. 合肥: 安徽农业大学, 2019.
|
| [11] |
Siddiq Z, Chen Y J, Zhang Y J, Zhang J L, Cao K F. More sensitive response of crown conductance to VPD and larger water consumption in tropical evergreen than in deciduous broadleaf timber trees. Agricultural and Forest Meteorology, 2017, 247: 399-407. DOI:10.1016/j.agrformet.2017.08.028 |
| [12] |
Cao M, Wu C, Liu J C, Jiang Y J. Increasing leaf δ13C values of woody plants in response to water stress induced by tunnel excavation in a karst trough valley: implication for improving water-use efficiency. Journal of Hydrology, 2020, 586: 124895. DOI:10.1016/j.jhydrol.2020.124895 |
| [13] |
Granier A. Evaluation of transpiration in a Douglas-fir stand by means of sap flow measurements. Tree Physiology, 1987, 3(4): 309-320. DOI:10.1093/treephys/3.4.309 |
| [14] |
吴芳. 黄土丘陵半干旱区刺槐、侧柏人工林耗水规律及影响因素研究[D]. 杨凌: 西北农林科技大学, 2011.
|
| [15] |
李成龙. 茂兰喀斯特森林主要树种蒸腾耗水规律及其生理生态响应[D]. 贵阳: 贵州大学, 2019.
|
| [16] |
Huang L, Zhang Z S. Effect of rainfall pulses on plant growth and transpiration of two xerophytic shrubs in a revegetated desert area: tengger Desert, China. CATENA, 2016, 137: 269-276. DOI:10.1016/j.catena.2015.09.020 |
| [17] |
Song X W, Lyu S, Wen X F. Limitation of soil moisture on the response of transpiration to vapor pressure deficit in a subtropical coniferous plantation subjected to seasonal drought. Journal of Hydrology, 2020, 591: 125301. DOI:10.1016/j.jhydrol.2020.125301 |
| [18] |
倪广艳, 赵平, 朱丽薇, 牛俊峰, 赵秀华, 曾小平. 荷木整树蒸腾对干湿季土壤水分的水力响应. 生态学报, 2015, 35(3): 652-662. |
| [19] |
Tie Q, Hu H C, Tian F Q, Guan H D, Lin H. Environmental and physiological controls on sap flow in a subhumid mountainous catchment in North China. Agricultural and Forest Meteorology, 2017, 240-241: 46-57. DOI:10.1016/j.agrformet.2017.03.018 |
| [20] |
Han C, Chen N, Zhang C K, Liu Y J, Khan S, Lu K L, Li Y G, Dong X X, Zhao C M. Sap flow and responses to meteorological about the Larix principis-rupprechtii plantation in Gansu Xinlong mountain, northwestern China. Forest Ecology and Management, 2019, 451: 117519. DOI:10.1016/j.foreco.2019.117519 |
| [21] |
王艳兵, 德永军, 熊伟, 王彦辉, 李振华, 刘千. 华北落叶松夜间树干液流特征及生长季补水格局. 生态学报, 2013, 33(15): 1375-1385. |
| [22] |
张婕, 蔡永茂, 陈立欣, 陈左司南, 张志强. 北京山区元宝枫夜间液流活动特征及影响因素. 生态学报, 2019, 39(9): 3210-3223. |
| [23] |
Nie Y P, Chen H S, Wang K L, Yang J. Water source utilization by woody plants growing on dolomite outcrops and nearby soils during dry seasons in karst region of Southwest China. Journal of Hydrology, 2012, 420-421: 264-274. DOI:10.1016/j.jhydrol.2011.12.011 |
| [24] |
Zhang Y J, Meinzer F C, Qi J H, Goldstein G, Cao K F. Midday stomatal conductance is more related to stem rather than leaf water status in subtropical deciduous and evergreen broadleaf trees. Plant, Cell & Environment, 2013, 36(1): 149-158. |
| [25] |
Cristiano P M, Campanello P I, Bucci S J, Rodriguez S A, Lezcano O A, Scholz F G, Madanes N, Di Francescantonio D, Carrasco L O, Zhang Y J, Goldstein G. Evapotranspiration of subtropical forests and tree plantations: a comparative analysis at different temporal and spatial scales. Agricultural and Forest Meteorology, 2015, 203: 96-106. DOI:10.1016/j.agrformet.2015.01.007 |
| [26] |
Peng X D, Dai Q H, Ding G J, Shi D M, Li C L. The role of soil water retention functions of near-surface fissures with different vegetation types in a rocky desertification area. Plant and Soil, 2019, 441(1/2): 587-599. |
| [27] |
Polverigiani S, McCormack M L, Mueller C W, Eissenstat D M. Growth and physiology of olive pioneer and fibrous roots exposed to soil moisture deficits. Tree Physiology, 2011, 31(11): 1228-1237. DOI:10.1093/treephys/tpr110 |
| [28] |
倪广艳. 外来树种的耗水特征及其对环境水资源的影响. 生态学报, 2021, 41(3): 934-942. |
| [29] |
Wu Y Z, Huang M B, Warrington D N. Black locust transpiration responses to soil water availability as affected by meteorological factors and soil texture. Pedosphere, 2015, 25(1): 57-71. DOI:10.1016/S1002-0160(14)60076-X |
| [30] |
Nie Y P, Chen H S, Wang K L, Ding Y L. Rooting characteristics of two widely distributed woody plant species growing in different karst habitats of southwest China. Plant Ecology, 2014, 215(10): 1099-1109. DOI:10.1007/s11258-014-0369-0 |
| [31] |
吴旭, 陈云明, 唐亚坤. 黄土丘陵区刺槐和侧柏人工林树干液流特征及其对降水的响应. 植物生态学报, 2015, 39(12): 1176-1187. DOI:10.17521/cjpe.2015.0114 |
| [32] |
Domingo F, Gutiérrez L, Brenner A J, Aguilera C. Limitation to carbon assimilation of two perennial species in semi-arid south-east spain. Biologia Plantarum, 2002, 45(2): 213-220. DOI:10.1023/A:1015136421445 |
| [33] |
Fisher J B, Baldocchi D D, Misson L, Dawson T E, Goldstein A H. What the towers don't see at night: nocturnal sap flow in trees and shrubs at two AmeriFlux sites in California. Tree Physiology, 2007, 27(4): 597-610. DOI:10.1093/treephys/27.4.597 |
| [34] |
Daley M J, Phillips N G. Interspecific variation in nighttime transpiration and stomatal conductance in a mixed New England deciduous forest. Tree Physiology, 2006, 26(4): 411-419. DOI:10.1093/treephys/26.4.411 |
| [35] |
范江琳, 曹萍萍, 冯良敏, 王佳津. 1961-2018年四川盆地夜雨特征分析. 高原山地气象研究, 2019, 39(4): 24-30. |