 2022, Vol. 42
2022, Vol. 42文章信息
- 叶国辉, 楚彬, 胡桂馨, 张飞宇, 华铣泽, 周富斐, 牛钰杰, 唐庄生, 花立民
- YE Guohui, CHU Bin, HU Guixin, ZHANG Feiyu, HUA Xianze, ZHOU Fufei, NIU Yujie, TANG Zhuangsheng, HUA Limin
- 高原鼢鼠干扰强度对祁连山东段高寒草甸大型土壤动物功能群特征及空间分布的影响
- Effects of plateau zokor (Myospalax baileyi) disturbance intensity on the characteristic and distribution of soil macro-fauna functional groups in alpine meadow in eastern Qilian Mountain
- 生态学报. 2022, 42(3): 1088-1097
- Acta Ecologica Sinica. 2022, 42(3): 1088-1097
- http://dx.doi.org/10.5846/stxb202006021422
-
文章历史
- 收稿日期: 2020-06-02
- 网络出版日期: 2021-10-11






大型土壤动物指栖息在陆地土壤中, 体长≥2 mm, 且超过90%的物种可被人眼识别的土壤无脊椎动物[1-2]。土壤动物作为陆地生态系统地下分解者和消费者, 通过食物网参与了植物-土壤界面的物质循环, 对调控生态系统过程和维持系统稳定具有重要作用[3]。由于土壤动物体型较小, 在土壤中大多处于幼虫发育阶段且受生活史动态影响, 导致种一级分类鉴定尚存在一定难度。而功能群是基于物种生活史、形态或对特定环境有相似反应的一类物种[4-5]。功能群不仅可有效解释物种对生态系统过程的响应机理, 而且对物种分类水平要求不高, 可简化群落物种研究的复杂性[6]。因此, 基于功能群研究可提高土壤动物时空动态变化的可预测性, 更加敏感反应生态系统结构和功能变化对土壤动物组成和空间分布的影响。
土壤动物群落组成受生物和非生物因子的共同影响[7]。生物过程会影响群落结构组成, 其中扩散限制和生态漂变是影响土壤动物群落组成的重要因素[8-9]。有研究表明动物群落组成可能由于资源短缺, 受低层自下而上的控制或受顶层捕食者自上而下营养级联效应的影响[10-11]。此外, 生态位理论表明环境可以影响物种分布, 环境过滤器使得物种分布在特定的生境中[12]。由于土壤动物有较差的环境耐受性和较弱的迁移能力, 其物种组成和空间分布格局对环境改变极为敏感[13-14]。在陆地生态系统中, 生物干扰常引起局域生境异质性增加, 可以影响土壤动物类群结构和多样性[15-16]。高原鼢鼠(Myospalax baileyi)是高寒草甸生态系统中营地下生活的植食性啮齿动物, 其采食、掘洞和推土造丘等行为造成草地生境破碎化。同时, 高原鼢鼠是高寒草甸生态系统重要的生物干扰源之一, 对植物-土壤界面的生态学过程具有显著影响[17], 进而可引起土壤动物多样性及空间分布的差异性[18], 且不同土壤动物类群对高寒草甸生态系统变化响应的敏感性不同[19]。
目前对于土壤动物功能群研究, 主要集中在环境因素对功能群的影响, 而忽略生物种群间的相互作用[20], 导致研究结果难以完全解释土壤动物空间格局高变异性的综合影响。因此, 在研究干扰对土壤动物功能群组成和分布影响时, 还需结合土壤动物功能群相互作用来综合分析。基于上述考虑, 本研究提出科学假设, 即高原鼢鼠干扰通过改变植物土壤环境因子来调控土壤动物功能类群间关系, 进一步影响土壤动物功能群组成及其分布格局。据此, 本研究依据高原鼢鼠新旧鼠丘数量作为干扰强度划分标准, 设立4个不同干扰强度小区。通过调查各小区草地大型土壤动物、植物群落组成以及土壤理化性质, 比较高原鼢鼠不同干扰强度下高寒草甸土壤动物功能群多样性和稳定性变化, 分析各功能群空间分布对环境因子变化的响应特征, 最终探究高原鼢鼠干扰对土壤动物群落功能和分布格局的影响机理。
1 材料与方法 1.1 研究区概况研究点位于祁连山东部高寒草甸区, 行政隶属于甘肃省天祝县。地理位置37°12′N, 102°46′E, 海拔2937 m。年均温零下0.1℃, 全年无绝对无霜期。植物生长季节从5月到9月约为120天。植被类型为高寒草甸, 优势种为多年生禾本科植物和莎草科植物, 如垂穗披碱草(Elymus nutans)、异针茅(Stipa aliena)、矮生蒿草(Kobresia humilis)和线叶嵩草(Kobresia capillifolia)。研究区域为冬季放牧草地(8.57只羊/hm2), 调查期间无家畜采食[21]。
1.2 试验设计本试验以空间尺度代替时间尺度的方法。依据高原鼢鼠新旧鼠丘数量划分干扰强度[22-23]。由于高原鼢鼠有很强的栖息地选择性, 难以找到同放牧强度、地形、植被和土壤, 特别是新旧鼠丘空间分布和数量一致的试验样地。参考梯度代替重复小区方法[22, 24]。本研究通过增设样方数量来代替重复小区从而确保调查数据的可靠性。本研究依据新旧鼠丘数量调查结果设置极重度(ED: 72个新鼠丘/hm2, 910个旧鼠丘/hm2)、重度(HD, 60个新鼠丘/hm2, 637个旧鼠丘/hm2)、中度(MD: 52个新鼠丘/hm2, 308个旧鼠丘/hm2)和轻度(LD: 28个新鼠丘/hm2, 100个旧鼠丘/hm2)4个干扰小区。小区大小为50 m × 50 m, 小区间隔大于50 m。各干扰小区内等距均匀设置9个调查样方, 期间避开鼠丘取样。于2018年6月下旬进行植物群落、大型土壤动物调查和土壤取样。
1.3 调查取样 1.3.1 植物群落调查在各干扰小区采样点采用0.5 m×0.5 m样方记录植物物种数目和密度。将各植物物种划分为莎草和杂类草类群, 室内烘箱75 ℃烘干至恒重, 称取地上生物量干重。
1.3.2 大型土壤动物群落调查在植物调查样方位置, 取长宽0.5 m×0.5 m, 深度0.15 m的土壤立方体。采用手捡方法挑出土壤立方体内大型土壤动物, 用75%乙醇保存。室内物种鉴定依据《中国土壤动物检索图鉴》[1]统计到科并计数。根据大型土壤动物食性将其分为植食性(Z)、捕食性(B)、腐食性(F)和杂食性(ZA)功能群[25]。
1.3.3 土壤理化性质测定土壤理化性质调查设置在土壤动物调查样方旁。野外测定0—15 cm土层土壤含水量、温度和紧实度。土壤化学性质取样采用直径为7 cm的土钻取0—15 cm土层土样, 装入自封袋带回实验室进行风干, 分别过0.25 mm和1 mm土壤筛进行土壤化学指标分析。室内土壤pH、土壤有机质、全氮、全磷和全钾测定参考《土壤理化分析》[26]
1.4 数据分析 1.4.1 功能群特征指数功能群丰富度用样方各大型土壤动物科类群数的数量表示。用功能群下各科级类群密度计算Shannon多样性指数(H′) 和Pielou均匀度指数(J)[27]。
优势度划分依据土壤动物个体占据总个体数10%以上为优势类群;占总个体数1%—10%为常见类群;占总个体数1%以下为稀有类群[28]。
1.4.2 群落稳定性指数和相似性指数群落稳定性指数[29]: 采用群落物种数与个体数之比(St/Si)和捕食类群物种数与植食性类群物种数之比(Sn/Sp)表示。
Sorenson相似性系数[30]:
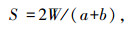
|
式中, W为样地A和B共有类群数量;a为样地A的类群数;b为样地B的类群数。Sorenson相似性系数的判别标准: 当S<0.24时为极不相似, S为0.25—0.49时中等不相似, S为0.50—0.74时中等相似, S为0.75—1.00极相似。
1.4.3 数据统计方法数据利用Excel 2013进行初步处理, 采用SPSS19.0进行方差分析, 用LSD方法进行多重比较。采用Canoco 4.5软件进行物种与环境变量多元排序。分析过程中对功能群丰度进行log(x+1)转换, 用功能群数据做除趋势对应分析(DCA)[31]。计算DCA排序轴梯度长度小于3采用冗余分析(RDA)。为评估环境因子对大型土壤动物功能群分布影响贡献值, 计算各环境因子的总效应和净效应[32]。
2 结果与分析 2.1 高原鼢鼠干扰下大型土壤动物功能群组成本试验共捕获植食性(Z)、捕食性(B)、腐食性(F)和杂食性(ZA)功能类群大型土壤动物累计1172只(表 1), 其中植食性(Z)类群为优势功能类群, 累计捕获932只分属6目17科21个类群。在LD、MD、HD和ED干扰区植食性类群个体数量占总捕获量的23.81%、16.55%、14.33%和24.83%。捕食性(B)、腐食性(F)和杂食性(ZA)功能群为高寒草甸土壤动物常见功能类群。捕食性(B)类群累计捕获82只, 分属4目8科。在LD、MD、HD和ED干扰区捕食性(B)类群占总捕获量的2.13%、1.02%、1.37%和2.47%。腐食性(F)类群和杂食性类群累计捕获158只, 分属2目3科。大型土壤动物组成调查结果显示, 植食性(Z)和捕食性(B)功能群具有较高的丰富度和多度, 是本研究区域高原鼢鼠栖息地的主要功能类群。
| 功能群 Functional groups |
类群数Group number | 个体数Individual number | 占比 Percent/% |
优势度 Dominance |
|||||||
| LD | MD | HD | ED | LD | MD | HD | ED | ||||
| 植食性Herbivore group | 13 | 14 | 12 | 19 | 279 | 194 | 168 | 291 | 79.52 | +++ | |
| 捕食性Predator group | 4 | 3 | 5 | 7 | 25 | 12 | 16 | 29 | 7.00 | ++ | |
| 腐食性Detritivore group | 2 | 2 | 1 | 2 | 16 | 58 | 3 | 35 | 9.56 | ++ | |
| 杂食性Omnivore group | 1 | 0 | 1 | 0 | 0 | 5 | 0 | 41 | 3.92 | ++ | |
| “+++”表示优势类群, 占总捕获量10%以上;“++”表示常见类群, 占总捕获量1%—10%;“+”表示稀有类群, 占总捕获量1%以下;ED: 极重度干扰Extremely disturbance;HD: 重度干扰Heavily disturbance;MD: 中度干扰Moderately disturbance;LD: 轻度干扰Lightly disturbance | |||||||||||
高原鼢鼠干扰对高寒草甸大型土壤动物功能群多样性存在一定影响(图 1)。植食性和捕食性功能群丰度、丰富度和Shannon多样性指数总体呈先下降后上升趋势, 表现为极重度干扰区植食性类群丰富度和Shannon多样性指数显著高于中度和重度干扰区(P<0.05)。高原鼢鼠干扰对捕食性功能群特征指数和植食性功能群丰度和均匀度指数无显著影响(P>0.05)。高原鼢鼠干扰下高寒草甸腐食性土壤动物功能群丰度变化明显, 中度干扰区腐食性类群丰度显著高于轻度和重度干扰区(P<0.05)。由于腐食性和杂食性功能群中类群丰富度和个体数量较少, 因此未分析高寒草甸腐食性和杂食性土壤动物功能群的丰富度、均匀度和Shannon多样性指数。

|
| 图 1 高原鼢鼠干扰下土壤动物功能群丰度、类群丰富度、均匀度和Shannon指数 Fig. 1 Abundance, group richness, evenness and Shannon indices of soil functional macro-fauna community under the plateau zokor disturbance 不同小写字母表示差异显著(P<0.05) |
高原鼢鼠干扰对高寒草甸大型土壤动物功能类群稳定性无显著影响(表 2)。各干扰区Sn/Sp的比值排序为ED>HD>LD>MD, 干扰区间Sn/Sp比值差异不显著(P>0.05), 但ED干扰区捕食性类群与植食性类群的比值(Sn/Sp)较高, 表明捕食类群对植食性类群的控制作用较强。土壤动物群落内部相互制约关系复杂, 其群落稳定性和抵抗外界干扰的能力也比较高, 表明其所处生态系统具有较高的稳定性和对外界不良干扰因素的抵御能力。不同干扰区中St /Si的比值排序为HD>ED>LD>MD, 干扰区间不存在显著差异(P>0.05), 表明土壤动物分布比较均衡。
| 高原鼢鼠干扰等级 Distrubance density |
稳定性 Relative stability value |
土壤动物功能类群相似性 Similarity indexes |
|||||
| Sn/Sp | St/Si | LD | MD | HD | ED | ||
| LD | 0.15 ± 0.040a | 0.33 ± 0.041a | * | 0.29 | 0.89 | 0.73 | |
| MD | 0.12 ± 0.037a | 0.32 ± 0.024a | 0.87 | * | 0.25 | 0.40 | |
| HD | 0.17 ± 0.063a | 0.41 ± 0.054a | 0.76 | 0.76 | * | 0.83 | |
| ED | 0.23 ± 0.053a | 0.37 ± 0.065a | 0.77 | 0.77 | 0.57 | * | |
| 对角线之上为捕食性类群相似性系数, 对角线之下为植食性类群相似性系数; Sn/Sp:为群落捕食类群物种数与相食性类群物种数之比; St/Si:为群落性种数与个人数之比; 稳定性栏中数值为平均值±标准误, 同列中相同字母表示差异不显著(P>0.05);土壤动物功能类群相似性栏中, *形成的对角线之上为捕食性类群相似性系数,对角线之下为植食性类群相似性系数 | |||||||
分析高原鼢鼠干扰对植食性和捕食性类群群落相似性指数影响发现, 各干扰区间植食性类群相似性系数S值均大于0.50, 表明高原鼢鼠不同干扰强度下高寒草甸植食性类群群落结构中等相似。中度(MD)干扰区捕食性类群与其它干扰区捕食性类群群落相似指数S值均小于0.50, 表明中度(MD)干扰区捕食性类群与其它干扰区捕食性类群群落结构中等不相似。
2.4 高原鼢鼠干扰下大型土壤动物功能类群分布与环境因子的关系土壤环境因子与土壤动物功能类群排序相关性(图 2), 轴1反映土壤温度(ST)变化, 轴2反映土壤全钾(TK)等其它土壤因子的综合变化。高寒草甸植食性(Z)、腐食性(F)和杂食性(ZA)土壤动物类群主要受土壤温度(ST)影响, 其次受土壤紧实度(SC)和全氮(TN)的影响(轴1方向);而捕食性(B)类群主要受土壤全钾(TK)和含水量(WC)影响(轴2方向)。在土壤环境因子综合影响下, 高原鼢鼠干扰区间土壤动物功能类群分布聚集情况不同(图 2)。轻度(LD)、中度(MD)和重度(HD)干扰区土壤动物功能类群分布重叠程度高, 土壤动物类群样点分布相对集中, 表明土壤环境异质性较低。极重度(ED)干扰区土壤动物类群样点分布相对分散, 聚集程度低, 表明土壤环境异质性较高。同时, 植食性(Z)、捕食性(B)、腐食性(F)和杂食性(ZA)土壤动物类群倾向分布于极重度干扰土壤环境条件。

|
| 图 2 土壤动物功能类群与土壤因子排序 Fig. 2 RDA biplot between soill factors and soil macro-fauna functional communities Z: 植食性Herbivore group; B: 捕食性Predator group; F: 腐食性Detritivore group; ZA: 杂食性Ominivore group; SC: 土壤紧实度Soil compaction; WC: 土壤含水量Soil moisture content; ST: 土壤温度Soil temperature; TOC: 土壤有机质Soil organic matter; TP: 土壤全磷Soil total P; TN: 土壤全氮Soil total N; TK: 土壤全钾Soil total K; pH: 土壤pH Soil pH; ED: 极重度干扰Extremely disturbance; HD: 重度干扰Heavily disturbance; MD: 中度干扰Moderately disturbance;LD: 轻度干扰Lightly disturbance |
植物环境因子与土壤动物功能类群排序相关性(图 3), 轴1反映莎草生物量(SB)变化, 轴2反映植被均匀度(Ev)等其它植物因子的综合变化。高寒草甸植食性(Z)和杂食性(ZA)土壤动物功能群主要受莎草生物量(SB)影响, 其次受植物均匀度(Ev)和Shannon指数(Sh)的影响(轴1方向);捕食性(B)和腐食性(F)类群主要受植物物种丰富度(Rh)、群落高度(He)和均匀度(Ev)影响(轴2方向)。在植物因子综合影响下, 高原鼢鼠干扰区间土壤动物功能群分布聚集程度不同(图 3)。轻度(LD)、中度(MD)和重度(HD)干扰区土壤动物功能群样点分布聚集程度高, 栖息地植被环境异质性低。极重度(ED)干扰区土壤动物类群样点分布相对分散, 聚集程度低, 植被环境异质性高。同时, 植食性(Z)、捕食性(B)和杂食性(ZA)土壤动物类群倾向分布于极重度干扰植被环境条件。

|
| 图 3 土壤动物功能类群与植物因子排序 Fig. 3 RDA biplot between plant factors and soil macro-fauna functional communities Ev: 植物均匀度指数Plant evenness index;Rh: 植物丰富度指数Plant richness index;Sh: 植物香农多样性指数Shannon′s diversity index;He: 植物群落高度Plant community height;SB: 莎草类植物地上生物量Cyperales above-ground biomass;FB: 杂类草植物地上生物量Forb above-ground biomass |
结合蒙特卡罗置换检验, 运用冗余分析(RDA)和偏冗余分析(Partial RDA)方法, 评估植物土壤因子对大型土壤动物功能类群分布影响贡献值(表 3)。RDA结果表明, 土壤温度(ST)、紧实度(SC)、全钾(TK)和全氮(TN)对大型土壤动物功能类群分布方差解释量达到显著水平(P<0.05), 而土壤有机质(TOC)、全磷(TP)和含水量(WC)等因子对大型土壤动物功能类群分布方差解释无显著贡献(P>0.05), 土壤环境因子累计解释总效应达到71.7%。Partial RDA结果表明, 土壤温度(ST)和全钾(TK)对大型土壤动物功能类群分布方差解释量达到显著水平(P<0.05), 土壤环境因子累计解释净效应达到40.7%。
| 环境因子 Envirment factor |
总效应/% Gross effect |
F | 净效应/% Net effect |
F |
| SC | 10.6 | 3.571* | 3.0 | 1.372 |
| ST | 22.6 | 8.752** | 13.6 | 6.160** |
| WC | 4.5 | 1.415 | 2.6 | 1.168 |
| TK | 9.5 | 3.140* | 10.3 | 4.665** |
| TOC | 5.8 | 1.846 | 5.9 | 2.696 |
| TN | 13.2 | 4.555** | 1.6 | 0.727 |
| TP | 3.1 | 0.971 | 3.1 | 1.394 |
| pH | 2.6 | 0.791 | 0.6 | 0.901 |
| He | 2.7 | 0.845 | 4.2 | 1.804 |
| SB | 10.3 | 3.431* | 9.5 | 4.058* |
| FB | 6.0 | 1.903 | 3.3 | 1.419 |
| Rh | 2.6 | 0.799 | 3.3 | 1.399 |
| Ev | 14.1 | 4.917** | 13.8 | 5.888** |
| Sh | 5.3 | 1.674 | 3.7 | 1.591 |
| * 表示在0.05水平(双侧) 上显著相关, ** 表示在0.01水平(双侧) 上显著相关; SC: 土壤紧实度Soil compaction; WC: 土壤含水量Soil moisture content; ST: 土壤温度Soil temperature; TK: 土壤全钾Soil total K; TOC: 土壤有机质Soil organic matter; TP: 土壤全磷Soil total P; TN: 土壤全氮Soil total N; pH: 土壤pH Soil pH; He: 植物群落高度Plant community height; SB: 莎草类植物地上生物量Cyperales above-ground biomass; FB: 杂类草植物地上生物量Forb above-ground biomass; Ev: 植物均匀度指数Plant evenness index; Rh: 植物丰富度指数Plant richness index; Sh: 植物Shannon′s多样性指数Shannon′s diversity index | ||||
植物因子对大型土壤动物功能类群分布影响贡献值(表 3)。RDA结果表明, 植物均匀度(Ev)和莎草生物量(SB)对大型土壤动物功能类群分布方差解释量达到显著水平(P<0.05), 植物因子对大型土壤动物功能类群分布解释累计解释总效应达到41%。Partial RDA结果表明, 植物因子累计解释净效应达到37.8%。上述结果表明土壤因子(显著贡献总效应55.9%) 对大型土壤动物功能类群分布方差解释量大于植物因子(显著贡献总效应24.4%)。
3 讨论 3.1 大型土壤动物功能群特征对高原鼢鼠干扰响应高原鼢鼠采食、造丘等行为造成草地环境异质性, 为草地生物栖息创造更多的生态条件, 有利于生态位分化, 使得更多物种共存[23]。有研究认为土壤动物群落维持机制主要受扩散、环境过滤和生物间作用的综合调控[33], 并在研究尺度上存在差异[34]。通常区域尺度上气候条件和地理屏障限制物种扩散进而影响物种分布格局[35], 而小尺度范围生物间相互作用尤为重要[36]。本研究为了消除研究尺度的影响, 并考虑到高原鼢鼠干扰可提供生境资源的多样性, 将土壤动物类群根据其食性生态位划分为四类功能群。在陆地生态系统中, 植食性类群作为广布种占据主要优势[19, 37-38], 而捕食性类群处于土壤食物网顶端对其他土壤动物群落组成和分布发挥重要调控作用, 因此本研究主要探讨了高原鼢鼠干扰对两者功能群多样性和稳定性影响。
本研究结果表明土壤动物植食性类群Shannon指数和丰富度对高原鼢鼠扰动响应较为敏感, 但不同干扰强度下丰度变异较小。研究表明, 植食性土壤动物类群对草地生态系统环境变化比较敏感[19]。在高寒草甸生态系统中, 高原鼢鼠的掘洞、造丘活动造成草地微地形变化, 是导致草地生境复杂性和异质性增强的主要干扰因子[39]。生境的复杂性增加了植食类群食物资源和栖息地微生境的多样性。因此, 植食性土壤动物类群多样性对高原鼢鼠干扰响应更为明显。一般植食性动物服从“下行控制”, 而捕食性动物服从“上行控制”[40]。但是, 由于资源限制造成植食性类群对植物资源的竞争, 使得其服从“上行控制”, 即资源控制消费者多度[40]。有研究表明大型土壤动物对地上植物群落结构和凋落物质量响应明显[41], 且土壤动物密度随植物盖度和凋落物输入而增加[42]。由于本研究区冬季持续放牧, 导致夏季调查期间草地凋落物残存量很少。并且, 高原鼢鼠干扰下植物群落结构和生产力变化差异小[18], 使得植物资源对植食性土壤动物类群丰度影响较小。其次, 本研究发现高原鼢鼠不同干扰强度下捕食性类群多样性无明显变化。由于高原鼢鼠不同干扰强度下植食性类群丰度无显著变化, 且捕食性类群受“上行控制”影响。因此, 捕食性类群丰度受高原鼢鼠干扰影响较小。另外, 高原鼢鼠不同干扰区域植食性和捕食性土壤动物类群分布比较均衡, 且植食性类群高的群落相似性和稳定性。因此, 捕食性类群多样性和相似性等受高原鼢鼠不同干扰强度影响变异较小。综合了环境过滤和生物营养级“上行控制”效应, 本研究发现高原鼢鼠干扰对捕食-植食类群种间调控关系影响较小。
3.2 高原鼢鼠干扰对高寒草甸土壤动物功能群空间分布影响高寒草甸植物以及土壤环境因子变化可以影响土壤动物的群落组成和空间分布[43-44]。本研究发现高原鼢鼠干扰导致土壤因子变化相比植物因子对土壤动物功能类群, 特别是植食性类群的空间分布影响更大。高原鼢鼠干扰可以造成草地微地形突变[39, 45-46], 这种突变首先影响土壤因子变化, 而植物因子变化明显存在滞后效应。本研究发现高原鼢鼠扰动引起小尺度土壤因子空间变异导致土壤动物功能群差异分布, 其中土壤温度、紧实度对植食性类群分布影响最大, 而腐食性类群受限于土壤全钾和土壤含水量的影响。伴随高原鼢鼠干扰强度增大, 其掘洞覆土行为降低了土壤表层紧实度[47], 且鼠丘数量增多造成丘间“郁闭度”升高可以显著降低土壤温度[23]。因此, 高原鼢鼠干扰改变土壤物理环境, 造成土壤动物功能类群组成和多样性空间分布差异。其次土壤全钾、全氮、有机质等土壤因子均可以影响土壤动物密度和类群数[19, 48], 但高原鼢鼠干扰对高寒草甸土壤营养以及含水量变化影响较小[18], 因此相对土壤化学因子, 植食性类群明显受限于土壤物理因子影响。不同功能群表现出资源利用效率的差异和生态位分化是对周围环境改变的适应策略[49]。植物资源多样性变化可以直接影响土壤动物的群落构成和分布, 本研究表明莎草类生物量和物种均匀度明显会影响植食性类群和腐食性类群空间分布。但由于植物因子对高原鼢鼠扰动响应迟滞, 其空间变异较小, 对高寒草甸土壤动物功能群的分布影响相比土壤因子较小。综上所述, 高原鼢鼠干扰下土壤物理环境因子成为高寒草甸土壤动物功能类群空间分布的主导因素。
4 结论土壤动物群落构建机制和空间格局具有尺度效应。小尺度生境下环境因子只能解释功能群组成分布部分变异, 更多变异受功能群间相互作用影响。本研究区高寒草甸植食性大型土壤动物功能群为优势类群, 高原鼢鼠干扰下高寒草甸土壤动物功能类群组成分布明显受环境因子制约和营养级联效应调控, 表现出土壤温度、紧实度以及莎草类地上生物量构成的土壤-植被环境异质性以及植食性类群对捕食性类群调控作用共同影响土壤动物功能群群落特征及分布。
| [1] |
尹文英. 中国土壤动物检索图鉴. 北京: 科学出版社, 1998: 393-701.
|
| [2] |
Gholami S, Sayad E, Gebbers R, Schirrmann M, Joschko M, Timmer J. Spatial analysis of riparian forest soil macrofauna and its relation to abiotic soil properties. Pedobiologia, 2016, 59(1/2): 27-36. |
| [3] |
董炜华, 李晓强, 宋扬. 土壤动物在土壤有机质形成中的作用. 土壤, 2016, 48(2): 211-218. |
| [4] |
孙国钧, 张荣, 周立. 植物功能多样性与功能群研究进展. 生态学报, 2003, 23(7): 1430-1435. DOI:10.3321/j.issn:1000-0933.2003.07.023 |
| [5] |
胡楠, 范玉龙, 丁圣彦, 廖秉华. 陆地生态系统植物功能群研究进展. 生态学报, 2008, 28(7): 3302-3311. |
| [6] |
Bai Y F, Wu J G, Pan Q M, Huang J H, Wang Q B, Li F S, Buyantuyev A, Han X G. Positive linear relationship between productivity and diversity: evidence from the Eurasian Steppe. Journal of Applied Ecology, 2007, 44(5): 1023-1034. DOI:10.1111/j.1365-2664.2007.01351.x |
| [7] |
Scheu S, Schaefer M. Bottom-up control of the soil macrofauna community in a beechwood on limestone: manipulation of food resources. Ecology, 1998, 79(5): 1573-1585. DOI:10.1890/0012-9658(1998)079[1573:BUCOTS]2.0.CO;2 |
| [8] |
Bahram M, Kohout P, Anslan S, Harend H, Abarenkov K, Tedersoo L. Stochastic distribution of small soil eukaryotes resulting from high dispersal and drift in a local environment. The ISME Journal, 2016, 10(4): 885-896. DOI:10.1038/ismej.2015.164 |
| [9] |
Powell J R, Karunaratne S, Campbell C D, Yao H Y, Robinson L, Singh B K. Deterministic processes vary during community assembly for ecologically dissimilar taxa. Nature Communications, 2015, 6(1): 8444. DOI:10.1038/ncomms9444 |
| [10] |
Hunter M D, Price P W. Playing chutes and ladders: heterogeneity and the relative roles of bottom-up and top-down forces in natural communities. Ecology, 1992, 73(3): 724-732. |
| [11] |
Power M E. Top-down and bottom-up forces in food webs: do plants have primacy. Ecology, 1992, 73(3): 733-746. DOI:10.2307/1940153 |
| [12] |
Lososová Z, Šmarda P, Chytrý M, Purschke O, Pyšek P, Sádlo J, Tichý L, Winter M. Phylogenetic structure of plant species pools reflects habitat age on the geological time scale. Journal of Vegetation Science, 2015, 26(6): 1080-1089. DOI:10.1111/jvs.12308 |
| [13] |
Meloni F, Civieta B F, Zaragoza J A, Moraza M L, Bautista S. Vegetation pattern modulates ground arthropod diversity in semi-arid mediterranean steppes. Insects, 2020, 11(1): 59. DOI:10.3390/insects11010059 |
| [14] |
Tao Y, Wang Z Q, Ma C, He H S, Xu J W, Jin Y H, Wang H X, Zheng X X. Vegetation heterogeneity effects on soil macro-arthropods in an alpine tundra of the Changbai Mountains, China. Plants, 2019, 8(10): 418. DOI:10.3390/plants8100418 |
| [15] |
刘继亮, 李锋瑞. 坡向和微地形对大型土壤动物空间分布格局的影响. 中国沙漠, 2008, 28(6): 1104-1112. |
| [16] |
Franklin E N, Morais J W, dos Santos E M R. Density and biomass of Acari and Collembola in primary forest, secondary regrowth and polycultures in central Amazonia. Andrias, 2001, 15: 141-153. |
| [17] |
Hu L, Zi H B, Ade L J, Lerdau M, Wang C T. Effects of zokors (Myospalax baileyi) on plant, on abiotic and biotic soil characteristic of an alpine meadow. Ecological Engineering, 2017, 103: 95-105. DOI:10.1016/j.ecoleng.2017.03.010 |
| [18] |
叶国辉, 楚彬, 胡桂馨, 张飞宇, 华铣泽, 周富斐, 花立民. 高原鼢鼠干扰下高寒草甸大型土壤动物多样性对环境因子的响应. 生态学报, 2021, 41(2): 792-802. |
| [19] |
张洪芝, 吴鹏飞, 崔丽巍. 高寒草甸大型土壤动物群落结构特征及其与环境的关系. 土壤学报, 2012, 49(6): 1267-1273. |
| [20] |
臧润国, 张志东. 热带森林植物功能群及其动态研究进展. 生态学报, 2010, 30(12): 3289-3296. |
| [21] |
Chu B, Ye G H, Yang S W, Zhou F F, Zhang F Y, Zhou J W, Hua L M. Effect of plateau zokor (Myospalax fontanierii)disturbance on plant community structure and soil properties in the eastern Qinghai-Tibet Plateau, China. Rangeland Ecology and Management, 2021, 73(4): 520-530. |
| [22] |
于海玲, 樊江文, 李愈哲, 史文娇. 高原鼢鼠干扰对三江源区高寒草甸群落特征的影响. 应用生态学报, 2018, 29(6): 1902-1910. |
| [23] |
Niu Y J, Zhou J W, Yang S W, Chu B, Zhu H M, Zhang B, Fang Q E, Tang Z S, Hua L M. Plant diversity is closely related to the density of zokor mounds in three alpine rangelands on the Tibetan Plateau. Peer J, 2019, 7: e6921. DOI:10.7717/peerj.6921 |
| [24] |
杨子翰, 陈泰祥, 郑巧燕, 卫万荣. 高原鼢鼠干扰对高寒草甸植物群落特征的影响. 草业学报, 2020, 29(5): 13-20. |
| [25] |
Doblas-Miranda E, Sánchez-Piñero F, González-Megías A. Soil macroinvertebrate fauna of a Mediterranean arid system: Composition and temporal changes in the assemblage. Soil Biology and Biochemistry, 2007, 39(8): 1916-1925. DOI:10.1016/j.soilbio.2007.02.009 |
| [26] |
中国科学院南京土壤研究所. 土壤理化分析. 上海: 上海科学技术出版社, 1978: 271-393.
|
| [27] |
吴鹏飞, 张洪芝, 崔丽巍, 钟红梅, 王永. 大型土壤动物群落对高寒草甸退化的响应. 土壤学报, 2013, 50(4): 786-799. |
| [28] |
Zheng X X, Tao Y, Wang Z Q, Ma C, He H, Yin X Q. Soil macro-fauna respond to environmental variations along a coastal-inland gradient. Peer J, 2020, 8: e9532. DOI:10.7717/peerj.9532 |
| [29] |
高宝嘉, 张执中, 李镇宇. 封山育林对昆虫群落结构及多样性稳定性影响的研究. 生态学报, 1992, 12(1): 1-7. |
| [30] |
Rempala G A, Seweryn M. Methods for diversity and overlap analysis in T-cell receptor populations. Journal of Mathematical Biology, 2013, 67(6/7): 1339-1368. |
| [31] |
Ter Braak C J F, Šmilauer P. CANOCO Reference Manual and User's Guide to CANOCO for Windows: Software for Canonical Community Ordination (Version 4.5). Ithaca, NY: Microcomputer Power, 2002.
|
| [32] |
Lososová Z, Chytrý M, Cimalová S, Kropáč Z, Otýpková Z, Pyšek P, Tichý L. Weed vegetation of arable land in central Europe: Gradients of diversity and species composition. Journal of Vegetation Science, 2004, 15(3): 415-422. DOI:10.1111/j.1654-1103.2004.tb02279.x |
| [33] |
Emerson B C, Gillespie R G. Phylogenetic analysis of community assembly and structure over space and time. Trends in Ecology & Evolution, 2008, 23(11): 619-630. |
| [34] |
Borcard D, Legendre P. All-scale spatial analysis of ecological data by means of principal coordinates of neighbour matrices. Ecological Modelling, 2002, 153(1/2): 51-68. |
| [35] |
Dong X Y, Li B, He F Z, Gu Y, Sun M Q, Zhang H M, Tan L, Xiao W, Liu S R, Cai Q H. Flow directionality, mountain barriers and functional traits determine diatom metacommunity structuring of high mountain streams. Scientific Reports, 2016, 6(1): 24711. DOI:10.1038/srep24711 |
| [36] |
Gascón S, Arranz I, Cañedo-Argüelles M, Nebra A, Ruhí A, Rieradevall M, Caiola N, Sala J, Ibàñez C, Quintana X D, Boix D. Environmental filtering determines metacommunity structure in wetland microcrustaceans. Oecologia, 2016, 181(1): 193-205. DOI:10.1007/s00442-015-3540-y |
| [37] |
刘鹏飞, 红梅, 美丽, 王文东, 赵乌英嘎, 李志新. 不同玉米秸秆还田量的土壤大型动物夏季群落动态特征. 土壤学报, 2020, 57(3): 760-772. |
| [38] |
张苗苗, 王咏雪, 田阔, 田嘉琦, 孙鹏, 水柏年. 沿浦湾秋茄种植前后大型底栖动物生态位和功能群变化. 中国水产科学, 2019, 26(5): 949-958. |
| [39] |
Niu Y J, Yang S W, Zhu H M, Zhou J W, Chu B, Ma S J, Hua R, Hua L M. Cyclic formation of zokor mounds promotes plant diversity and renews plant communities in alpine meadows on the Tibetan Plateau. Plant and Soil, 2020, 446(1/2): 65-79. |
| [40] |
Begon M, Townsend C R, Harper J L. 生态学-从个体到生态系统(第四版). 李博, 张大勇, 王德华, 译. 北京: 北京高等教育出版社, 2016: 550-556.
|
| [41] |
Decaëns T, Dutoit T, Alard D, Lavelle P. Factors influencing soil macrofaunal communities in post pastoral successions of western France. Applied Soil Ecology, 1998, 9(1/3): 361-367. |
| [42] |
Frouz J, Prach K, Pižl V, Háněl L, Starý J, Tajovský K, Materna J, Balík V, Kalčík J, Řehounková K. Interactions between soil development, vegetation and soil fauna during spontaneous succession in post mining sites. European Journal of Soil Biology, 2008, 44(1): 109-121. DOI:10.1016/j.ejsobi.2007.09.002 |
| [43] |
邵珍珍, 吴鹏飞. 小型表栖节肢动物群落对高寒湿地退化的响应. 生态学报, 2019, 39(19): 6990-7001. |
| [44] |
龙伟, 高艳美, 吴鹏飞. 若尔盖高寒草甸退化对表栖节肢动物群落的影响. 生态学杂志, 2018, 37(1): 128-138. |
| [45] |
宗文杰, 江小雷, 严林. 高原鼢鼠的干扰对高寒草地植物群落物种多样性的影响. 草业科学, 2006, 23(10): 68-72. DOI:10.3969/j.issn.1001-0629.2006.10.017 |
| [46] |
楚彬, 马素洁, 周延山, 姬程鹏, 周建伟, 周睿, 田永亮, 花立民. 祁连山东段高原鼢鼠(Myospalax baileyi)土丘空间分布格局及其与环境因子的空间关联性. 生态学报, 2018, 38(3): 964-974. |
| [47] |
周延山, 花立民, 楚彬, 刘丽, 姬程鹏, 田永亮. 高原鼢鼠繁殖特性与其栖息草地质量的关系. 兽类学报, 2017, 37(1): 87-96. |
| [48] |
李雨, 吴鹏飞, 龙伟, 马金豪. 高寒地区种植不同种类牧草对土壤节肢动物群落的影响. 生态学报, 2019, 39(20): 7697-7708. |
| [49] |
Yamada T, Zuidema P A, Itoh A, Yamakura T, Ohkubo T, Kanzaki M, Tan S, Ashton P S. Strong habitat preference of a tropical rain forest tree does not imply large differences in population dynamics across habitats. Journal of Ecology, 2007, 95(2): 332-342. DOI:10.1111/j.1365-2745.2006.01209.x |