 2022, Vol. 42
2022, Vol. 42文章信息
- 全为民, 张云岭, 齐遵利, 许敏, 范瑞良, 王桃妮, 李楠楠, 孙兆跃, 周海生, 李春, 张秀文
- QUAN Weimin, ZHANG Yunling, QI Zunli, XU Min, FAN Ruiliang, WANG Taoni, LI Nannan, SUN Zhaoyue, ZHOU Haisheng, LI Chun, ZHANG Xiuwen
- 河北唐山曹妃甸-乐亭海域自然牡蛎礁分布及生态意义
- Distribution and ecological status of natural oyster reefs on the coast of Caofeidian-Leting, Tangshan, Hebei Province
- 生态学报. 2022, 42(3): 1142-1152
- Acta Ecologica Sinica. 2022, 42(3): 1142-1152
- http://dx.doi.org/10.5846/stxb202005101172
-
文章历史
- 收稿日期: 2020-05-10






2. 唐山海洋牧场实业有限公司, 唐山 063611;
3. 河北农业大学海洋学院, 秦皇岛 066003
2. Tangshan Marine Ranching Industrial Limited Company, Tangshan 063611, China;
3. Ocean College, Hebei Agriculture University, Qinhuangdao 066003, China
牡蛎礁是由大量牡蛎固着生长于硬底物表面所形成的一种生物礁系统, 被喻为温带地区的“珊瑚礁”[1-3]。除为人类提供大量鲜活牡蛎以供食用外, 牡蛎礁具有净化水体、维持生物多样性、提供鱼类生境、固碳和防止岸线侵蚀等生态功能[3-5]。牡蛎礁曾广泛分布于全球亚热带和温带海区的多数海湾、河口以及部分海岸线的潮间带和潮下带。但过度捕捞、环境污染、生境破坏和病害侵染使全球野生牡蛎资源量急剧下降, 牡蛎礁生境遭受破坏或丧失, 过去100多年来全球牡蛎礁面积约下降了85%, 从而改变了近岸生态系统的结构与功能[4-5]。
牡蛎礁作为重要的海岸带栖息生境之一, 曾在我国温带和亚热带海区的潮间带和浅水潮下带有着广泛分布[6-12], 如20世纪80年代我国地质学者曾在我国南北沿海的许多河口和海湾内记录到牡蛎礁分布, 然而这些研究更多关注牡蛎礁的地理分布、海岸地貌发育和古环境, 而很少研究牡蛎礁的生态现状和生态系统服务价值。2000年以后, 国内才开始关注牡蛎礁的生态功能[1], 并对天津大神堂[13]和江苏小庙洪[9-10]等少数现存活体牡蛎礁的生态现状进行零星的调查研究, 结果显示这些牡蛎礁退化严重, 大部分自然牡蛎礁已经消失[10, 13-15]。截止目前, 国内有关牡蛎礁的生态学研究仍然有限, 无法全面掌握目前我国自然牡蛎礁的地理分布及生态现状, 制约着我国海岸带的保护与修复。
渤海湾曾是我国牡蛎礁的主要分布区, 但由于海岸带历史变迁等原因, 大量牡蛎礁被埋入地下成为古牡蛎礁, 而该区现存活体牡蛎礁的报道较少[6, 13]。《滦州志》卷九·《封域(下)·古迹漕运附》载: “知府孙维城复议海运故道直捷, 而所经蚕沙绿洋各口蛎房山石臼坨……”, 所描述的“蛎房山”即为牡蛎礁;据历史资料和当地渔民描述, 该海域自然牡蛎礁分布于溯河河口以东、小青河河口以西的低潮线以下海域, 后来牡蛎礁退化或消失。近年来, 在原有牡蛎礁分布海域又能捕捞到大量活体牡蛎, 极大的野生牡蛎捕捞量引起社会的广泛关注, 拯救并保护这一珍贵的自然资源及其生境势在必行。在实施自然牡蛎礁的保护与修复措施前, 亟待解决以下3个科学问题: (1)该自然牡蛎礁的地理分布区域及分布范围?(2)该自然牡蛎礁中牡蛎种群及礁体动物群落的生态现状如何?(3)该自然牡蛎礁提供多大的生态系统服务价值?为回答以上科学问题, 本研究于2019年3月对该自然牡蛎礁的空间分布、生态环境、牡蛎生物学和礁体动物群落开展了调查, 并评估了该自然牡蛎礁的生态系统服务价值, 研究结果可为该自然牡蛎礁的保护与修复提供基础信息。
1 材料与方法 1.1 研究地点研究地点位于河北省唐山市曹妃甸-乐亭海域(图 1), 是介于曹妃甸工业区与京唐港之间的半封闭海湾, 具有离岸沙岛、深槽和浅滩等多种地貌单元。海区潮汐型态系数为0.77, 属于不规则半日潮, 平均高潮位为0.81 m, 平均低潮位为-0.73 m, 平均潮差为1.54 m[16]。根据历史文献、现场调查和牡蛎捕捞统计资料, 初步确定溯河口(SRE)、溯河(SR)和捞鱼尖(LYJ)3个自然牡蛎礁分布区。共设置12个生态调查站位, 其中SR牡蛎礁3个站位、SRE牡蛎礁5个站位和LYJ牡蛎礁4个站位(图 1)。

2019年3月24—30日执行野外生态调查, 调查内容包括生态环境(水温、盐度、总悬浮颗粒物(TSS)、叶绿素a(Chla)和浮游植物)、牡蛎礁分布、牡蛎生物学(种类、密度、生物量、壳高(SH)、肥满度(CI)和遗传多样性)和礁体动物群落(种类、密度和生物量)。
在高潮位时, 采用YSI水质分析仪现场测定表层海水水温和盐度;用采水器采集1000 mL表层海水, 等分成两份, 现场用装有0.45 μm微孔滤膜的便携式真空泵抽滤, 滤膜冰冻保存后带回实验室分析测定, 海水Chla和TSS浓度分别采用分光光度计法和重量法测定, 另每站取1000 mL表层海水用鲁哥式溶液现场固定后带回室内镜检计数浮游植物。
采用社会调查、潜水调查和水下摄像等方法调查自然牡蛎礁的地理分布, 具体方法为: 首先和当地牡蛎捕捞从业人员进行访谈, 初步获取牡蛎重点采捕区的位置和范围, 再提取每个区域牡蛎采捕渔船的GPS导航航迹数据, 最后采用潜水调查和水下摄像进行验证, 综合上述信息确定牡蛎礁地理位置和分布范围。在SRE和SR, 由潜水员下潜至牡蛎礁表面进行采样, 每个站位采集1个0.5 m×0.5 m样方内(深度10cm)所有的活体牡蛎、牡蛎壳和生物样品, 装于网目0.5 mm的聚乙烯网袋中带回岸上分检。在LYJ, 低潮位时水深仅约0.4m, 直接手工采样。
回岸后分捡每个样品中的活牡蛎和礁体动物。记录活牡蛎的个体数、测定其总重(精确至0.1 g);从每个调查区样品中随机选取不少于50个牡蛎, 用游标卡尺测量壳高(SH, 精确至1mm), 带部分牡蛎样品回室内分析种类和肥满度。礁体动物(不包括牡蛎)样品用75%乙醇固定保存后, 带回室内鉴定至最低的分类单元, 并计数和称重。
基于外壳形态, 选取96个牡蛎样品进行线粒体16S rDNA基因序列分析, 据此分析牡蛎的种类和遗传多样性。从每个调查区样品中随机选取不少于20个牡蛎, 剥取牡蛎的软体部分, 测定湿肉重(WTW, g), 然后将软体部分放入烘箱中(60℃)烘干至恒重(约24 h), 并测定其干肉重(FTW, g), 依据下列公式计算牡蛎的肥满度(Condition index, CI):
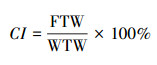
|
将测得的16S rDNA序列同GenBank中登录的牡蛎序列进行系统发育分析, 采用Phylip 3.5软件的邻接法构建系统树, 利用Donax trunculus作为外群, 置信号由1000次重复检验。
牡蛎或礁体动物的密度(个/m2)统计为每1m2牡蛎礁中的个体数, 生物量(g/m2或kg/m2)为每1 m2牡蛎礁中的总鲜重。运用一维方差分析(1-way ANOVA)检验牡蛎指标(密度、生物量和SH)或礁体动物群落指标(密度和生物量)在3个分布区(SR, SRE和LYJ)间的差异(Tukey后检验)。如数据没有通过方差同质性和方差齐性检验, 则采用一维排序方差分析(1-way ANOVA on rank, Ducnn后检验)。另采用Pearson相关性检验牡蛎指标与礁体动物群落指标之间的相关关系;所有统计分析及制图均在SigmaPlot 10.0软件中完成。
1.4 生态系统服务价值评估根据北美自然牡蛎礁的研究结果[3], 牡蛎礁的生态系统服务价值标准为10.6万美元hm-2 a-1。据此标准, 本研究评估了该自然牡蛎礁的生态系统服务价值。
2 结果与分析 2.1 环境指标调查期间海水温度介于4.1—10.9 ℃, 平均为8.2℃;海水盐度介于30.8—34.2, 平均为33.4;TSS浓度介于12.2—43.7 mg/L, 平均为27.8 mg/L;Chla浓度介于0.84—3.45 mg/m3, 平均为2.37 mg/m3。共采集到浮游植物2门21种, 其中硅藻17种和甲藻4种, 浮游植物丰度介于(0.1—23.94)×104个细胞/L, 平均为3.29×104个细胞/L, SRE中浮游植物丰度高于SR和LYJ, 主要优势种为旋链角毛藻Chaetoceros curvisetus。
| 指标Factors | 溯河口海域(SRE) | 溯河(SR) | 捞鱼尖(LYJ) | |||||
| 范围Range | 平均Mean | 范围Range | 平均Mean | 范围Range | 平均Mean | |||
| 水温Water temperature/℃ | 4.1—10.9 | 8.4 | 9.8—10.2 | 10.1 | 6.5—6.8 | 6.7 | ||
| 盐度Salinity | 30.8—33.1 | 32.0 | 31.0—32.8 | 32.4 | 34.0—34.2 | 34.1 | ||
| 悬浮物TSS/(mg/L) | 12.2—40.0 | 28.4 | 34.1—43.7 | 39.0 | 17.9—22.3 | 19.3 | ||
| 叶绿素Chla/(mg/m3) | 2.33—3.45 | 2.80 | 3.20—3.41 | 3.33 | 0.84—1.13 | 1.03 | ||
| 浮游植物 Phytoplakton/(×104个细胞/L) |
0.86—23.94 | 6.17 | 0.33—0.90 | 0.62 | 0.10—0.47 | 0.28 | ||
自然牡蛎礁主要分布于以下3个区域(图 1)。①SRE牡蛎礁: 主要分布于老龙沟潮汐通道两则0—5 m水深的浅水区, 有6个斑块状分布区, 单个牡蛎礁面积介于0.17—3.62 km2之间, 总面积约10 km2, 礁体发育较好, 礁体高度约0.3—0.5 m, 是牡蛎的主要采捕区。②SR牡蛎礁: 分布于溯河河道及溯河河口段, 面积约0.5 km2, 尤以河床底部的牡蛎礁发育较好。③LYJ牡蛎礁: 位于石臼坨和腰坨西侧, 面积约5 km2, 单个礁体斑块面积介于0.2—1.0 m2之间, 盖度约5%, 处于发育早期, 牡蛎与大型海藻共生。
2.3 牡蛎指标经PCR扩增, 获得了96个牡蛎的16S rDNA基因片段扩增产物(图 2), 可比16S rDNA序列为450 bp左右, 共检测到20个单倍型, 其中92个为长牡蛎Crassostrea gigas、3个为侏儒牡蛎Nanostrea fluctigera、1个为巨蛎属未知种Crassostrea sp.(图 3)。长牡蛎有17个单倍型、单倍型多样性为0.641、核苷酸多样性为0.00278;侏儒牡蛎有2个单倍型、单倍型多样性为0.667、核苷酸多样性为0.00147。

|
| 图 2 部分牡蛎样本的线粒体16S rDNA扩增电泳图谱 Fig. 2 Amplified products of mitochondrial 16S rDNA gene of partial oyster specimens 1—22为牡蛎样本, M为DL2000分子标记 |

|
| 图 3 基于96个牡蛎样本16S rDNA数据构建的系统发育树 Fig. 3 Phylogenetic trees constructed from 16S rDNA sequence of 96 oyster specimens CFDLT: 曹妃甸-乐亭海域的牡蛎样本Oyster specimens from Caofeidian-Leting coast |
SRE中牡蛎密度介于2764—3912个/m2, 平均为(3251±223)个/m2;SR中牡蛎密度介于1008—1200个/m2, 平均为(1135±63)个/m2;LYJ中牡蛎密度介于104—280个/m2, 平均为(173±41)个/m2;牡蛎密度的大小排序为: SRE>SR>LYJ (图 4, Tukey后检验, P<0.05);牡蛎生物量的大小排序为: SR>SRE>LYJ (图 4, Tukey后检验, P<0.05)。

|
| 图 4 3个牡蛎礁区间牡蛎密度和生物量的比较 Fig. 4 Comparisons on oyster densities and biomass among three oyster reef zones 误差棒为±1标准误, 不同小写字母表示显著性差异(P<0.05) |
图 5显示了3个自然牡蛎礁分布区中牡蛎的壳高—频率分布。SR中牡蛎SH(中值SH: 58 mm)显著大于SRE(中值SH: 40 mm)和LYJ(中值SH: 33 mm)(Dunn后检验, P<0.05), 而SRE和LYJ间没有差异(Dunn后检验, P>0.05)。

|
| 图 5 3个牡蛎礁区间牡蛎壳高—频率分布的比较 Fig. 5 Comparisons on oyster SH-frequency distributions among three oyster reef zones |
SRE中牡蛎CI值介于9.28%—18.21%, 平均为(14.59±0.38)%;SR中牡蛎CI值介于13.09%—21.09%, 平均为(17.28±0.53)%;LYJ中牡蛎CI值介于10.18%—18.45%, 平均为(14.15±0.42)%;牡蛎CI值的大小排序为: SR>SRE=LYJ (Tukey后检验, P<0.05)
2.4 礁体动物群落指标通过定性和定量采样, 在牡蛎礁内记录到49种礁体动物, 其中软体动物16种、节肢动物16种、环节动物8种、棘皮动物5种、腔肠动物2种、星形动物和脊索动物各1种(表 2)。
| 类群Taxa | 物种Species | 类群Taxa | 物种Species | |
| 软体动物 | 菲律宾蛤仔Ruditapes philippinarum | 节肢动物Arthropoda | 东方拟银杏蟹Paractaea orientalis | |
| Mollusks | 栖缝蛤Hiatella sp. | 方蟹Grapsidae | ||
| 史氏背尖贝Notoacmea schrenckii | 美丽瓷蟹Porcellana pulchra | |||
| 双纹须蚶Barbatia bistrigata | 日本蟳Charybdis japonica | |||
| 凸壳肌蛤Musculus senhousia | 绒毛近方蟹Hemigrapsus penicillatus | |||
| 栉孔扇贝Chlamys farreri | 特异大权蟹Macromedaeus distinguendus | |||
| 中华孔螂Poromya sinica | 小相手蟹Nanosesarma minutum | |||
| 布尔小笔螺Mitrella burchardi | 长指近方蟹Hemigrapsus longitarsis | |||
| 单齿螺Monodonta labio | 葛氏长臂虾Palaemon gravieri | |||
| 丽核螺Mitrella bella | 鲜明鼓虾Alpheus distinguendus | |||
| 脉红螺Rapana venosa | 口虾蛄Oratosquilla oratoria | |||
| 托氏琩螺Umbonium thomasi | 盖鳃水虱Idotea sp. | |||
| 微小螺Elachisina sp. | 钩虾Gammaridae | |||
| 秀丽织纹螺Nassarius festivus | 日本大螯蜚Grandidierella japonica | |||
| 疣荔枝螺Thais clavigera | 团水虱Sphaeroma sp. | |||
| 朝鲜鳞带石鳖Lepidozona coreanica | 泥藤壶Balanus uliginosus | |||
| 环节动物 | 多齿围沙蚕Perinereis nuntia | 棘皮动物 | 海地瓜Acaudina molpadioides | |
| Annelids | 覆瓦哈鳞虫Harmothoe imbricata | Echinodermata | 多棘海盘车Asterias amurensis | |
| 尖刺缨虫Potamilla acuminata | 金氏真蛇尾Ophiura kinbergi | |||
| 内刺盘管虫Hydroides ezoensis | 滩栖阳遂足Amphiura vadicola | |||
| 琴蜇虫Lanice conchilega | 马粪海胆Hemicentrotus pulcherrimus | |||
| 肾刺缨虫Potanmilla reniformis | 腔肠动物 | 海笔Pennatula phosphorea | ||
| 索沙蚕Lumbrineris sp. | Coelenterata | 侧花海葵Anthopleura sp. | ||
| 长吻沙蚕Glycera chirori | 星虫动物Sipuncula | 革囊星虫Phascolosoma sp. | ||
| 脊索动物 Chordata |
柄海鞘Styela clava |
SRE中礁体动物密度介于404—552个/m2, 平均为(493±26)个/m2;SR中礁体动物密度介于164—320个/m2, 平均为(228±47)个/m2;LYJ中礁体动物密度介于28—44个/m2, 平均为(35±5)个/m2。平均总密度的大小排序为: SRE>SR>LYJ (图 6, Tukey后检验, P<0.05);平均总生物量的排序为: SRE显著高于LYJ(图 6, Dunn后检验, P<0.05), 而SR介于SRE和LYJ之间(图 6, Dunn后检验, P>0.05)。

|
| 图 6 3个牡蛎礁区间大型底栖动物群落总密度和总生物量的比较 Fig. 6 Comparisons on total densities and biomass of benthic macroinvertebrates among three oyster reef zones 误差棒为标准误, 不同小写字母表示显著性差异(P<0.05) |
牡蛎密度与礁体动物密度和生物量呈现显著的正相关性(表 3, 图 7, P<0.05), 而牡蛎生物量与礁体动物密度和生物量之间的相关性并不显著(表 3, P>0.05)。
| 指标 Factors |
牡蛎密度 Oyster density |
牡蛎生物量 Oyster biomass |
礁体动物密度 Benthic macroinvertebrate density |
| 牡蛎生物量Oyster biomass | 0.204 (0.526) | ||
| 礁体动物密度Benthic macroinvertebrate density | 0.918 (<0.001) | 0.178 (0.600) | |
| 礁体动物生物量Benthic macroinvertebrate biomass | 0.864 (<0.001) | 0.042 (0.903) | 0.920 (<0.001) |

|
| 图 7 3个牡蛎礁区中牡蛎密度和礁体动物群落总密度和总生物量之间的线性回归关系 Fig. 7 Linear regressions between oyster densities and benthic macroinvertebrate communities metrics (densities and biomass) across three oyster reef zones |
由于SR牡蛎礁面积较小和LYJ牡蛎礁中牡蛎密度较低, 仅评估SRE牡蛎礁的生态系统服务价值, 如该自然牡蛎礁未受破坏, 其生态系统服务价值将达到1.06×108美元/a, 按当前汇率6.51计折合成人民币约6.90×108元/年。
3 讨论 3.1 自然牡蛎礁的地理分布渤海湾沿岸曾是我国牡蛎礁的集中分布区, 如在天津沿岸发现了许多成片的古牡蛎礁[18];除天津大神堂有活体牡蛎礁分布外, 在渤海湾其它海域未见活体牡蛎礁的报道。从史书记载, 曹妃甸-乐亭海域历史上分布有大量牡蛎礁, 但没有这片牡蛎礁地理分布和生态现状的基础信息。受人力和资金限制, 本研究并未采用声呐勘测手段精确调查自然牡蛎礁的地理分布, 但通过社会统计和现场调查等多种方法初步描述了该海域自然牡蛎礁的地理分布, 估算牡蛎礁总面积高达15 km2, 表明该牡蛎礁是截至目前为止我国面积最大的自然活体牡蛎礁。这是继曹妃甸海草场[17]后在该海域的又一重大发现, 进一步补充了我国牡蛎礁地理分布及生态现状的基础信息, 研究成果对于曹妃甸-乐亭海域海岸带管理和保护具有重要意义。
本研究发现3个自然分布区中以溯河口牡蛎礁发育最好、牡蛎密度最高和生物多样性最为丰富, 原因可能是该牡蛎礁分布于老龙沟潮汐通道两侧的浅水区, 水深约1—5 m, 老龙沟的涨潮流携带而来的丰富浮游生物为牡蛎生长和牡蛎礁发育提供了充足的饵料[19]。与历史分布区相比, 该海域现代牡蛎礁分布区整体向南发育, 原因可能是西南侧实施的曹妃甸围填海工程减弱了溯河口海域的水动力, 从而为牡蛎附着生长和牡蛎礁发育提供更为稳定的水动力环境, 减弱的水动力环境和增加的水体滞留时间更有利于牡蛎幼体的附着与生长[20-21], 加快了现代牡蛎礁向外侧发育。
3.2 牡蛎和礁体动物多样性本研究采用线粒体16S rDNA基因序列变异分析发现, 自然牡蛎礁内分布有长牡蛎和侏儒牡蛎, 其中以长牡蛎在数量上占绝对优势, 是造礁的牡蛎种类。长牡蛎原产于日本、韩国和中国江苏连云港以北沿海, 为世界性养殖种;侏儒牡蛎分布于印度一西太平洋浅水区, 我国最早报道于东海北部水深47 m海区[22], 有关该种的报道非常少, 本研究首次在曹妃甸-乐亭海域记录到这一物种, 为阐明该种的生物地理格局提供了基础信息。
牡蛎密度和大小是表征牡蛎礁生态现状的通用指标[23]。本研究发现该海域是以长牡蛎为造礁种的活体牡蛎礁, 3个自然牡蛎礁区间牡蛎密度差异很大, 尤以溯河口海域中牡蛎密度最高, 平均密度达到(3251 ±223)个牡蛎/m2, 明显高于国内外自然或人工修复牡蛎礁中的牡蛎密度[24-28];3个牡蛎礁区中牡蛎壳高均呈现单峰的正态分布, 分布有一定数量的牡蛎成体和稚贝, 表明该自然牡蛎礁拥有1个健康的、可持续的牡蛎种群;如果加以有效保护, 该牡蛎礁将持续发挥显著的生态系统服务功能。
定居性礁体动物群落是评估牡蛎礁生态现状、生物多样性和生境价值的重要指标之一[23]。本研究报道的礁体动物密度明显低于江苏海门蛎蚜山牡蛎礁[9, 28]、长江口牡蛎礁[27]和美国东海岸牡蛎礁[24-26], 原因可能是调查期间该海域平均水温低于10℃, 许多底栖动物仍未经历春季的资源补充;但牡蛎礁内动物密度仍高于曹妃甸近岸海域底栖动物密度[29]。另外, 本研究也发现牡蛎密度与礁体动物群落总密度和总生物量之间呈现显著的正相关关系, 原因是牡蛎密度越高, 其聚集生长形成的礁体结构越复杂, 而生境结构的复杂性与其维持的礁体动物丰度之间有显著的正相关性[30]。
3.3 牡蛎礁保护与生态修复策略牡蛎礁是生态系统服务价值最高的海洋生境之一。基于北美自然牡蛎礁的评估结果表明[3], 牡蛎礁的生态系统服务价值约为10.6万美元hm-2 a-1, 高于红树林(8.2万美元hm-2 a-1)、海草场(3.1万美元hm-2 a-1)和永久湿地(2.1万美元hm-2 a-1)。如对该自然牡蛎礁加以保护并免遭破坏, 其生态系统服务价值将高达6.90×108元/年;以鲜活牡蛎平均单价10元/kg计算, 则该牡蛎礁产生的生态系统服务价值相当于每年生产0.69×108kg牡蛎的经济产值, 按牡蛎礁面积10 km2计算, 则每1 m2牡蛎礁的生态系统服务价值相当于6.9 kg可捕规格牡蛎的经济产值(69元)。
自2018年春季开始, 该自然牡蛎礁持续面临较高的捕捞压力, 主要捕捞工具为牡蛎专用拖网, 该网具网口下杠有钢制耙, 拖网作业除捕捞活体牡蛎外, 也破坏了牡蛎礁生境结构, 调查期间观察到大量牡蛎壳随活体牡蛎一同被捕捞, 生境结构的破坏不仅降低了牡蛎幼体的附着基数量, 也减少了活体牡蛎礁面积, 从而对自然牡蛎礁造成长期的负面影响。随着近两年的持续捕捞, 该海域牡蛎资源量急剧下降, 2018年拖网渔获中活体牡蛎的比例达到70%, 至2019年调查时下降至20%, 表明该牡蛎礁的生态现状不容乐观, 亟待加强保护和修复。
曹妃甸-乐亭海域水流较缓, 具有广阔的潮间带滩涂和潮下带浅水区, 生境类型异质多样, 分布有大面积的自然牡蛎礁和海草场, 水体初级生产力高, 生物资源十分丰富[16]。建议以溯河口海域自然牡蛎礁和海草场为核心区域, 通过投放小型的适合牡蛎附着生长的人工礁体(牡蛎壳、混凝土模块)[31], 快速修复当地的牡蛎礁生境及其生物资源, 建设一个牡蛎礁国家级海洋牧场示范区, 重建鱼类重要栖息生境, 发挥牡蛎礁和海草场的鱼类育幼场功能, 养护海洋渔业资源, 修复与优化海洋生态系统, 探索以经济鱼类、蟹类和螺类为产出的海洋牧场利用模式。
致谢: 感谢唐山市曹妃甸蚕沙口村鱼满仓渔业捕捞专业合作社杨连革和朱振江同志在海上调查采样中提供的帮助, 感谢上海市水产研究所安传光博士鉴定礁体动物样品。| [1] |
全为民, 沈新强, 罗民波, 陈亚瞿. 河口地区牡蛎礁的生态功能及恢复措施. 生态学杂志, 2006, 25(10): 1234-1239. DOI:10.3321/j.issn:1000-4890.2006.10.016 |
| [2] |
孙兆跃, 范瑞良, 隋延鸣, 唐保军, 姜伟, 修玉娜, 李楠楠, 李雪英, 全为民. 全为民. 3种无脊椎动物对近江牡蛎Crassostrea ariakensis和熊本牡蛎C.sikamea的捕食研究. 生态学报, 2021, 41(7): 2895-2901. |
| [3] |
McLeod I M, Zu Ermgassen P S E, Gillies C L, Hancock B, Humphries A. Chapter 25-can bivalve habitat restoration improve degraded estuaries?//Wolanski E, Day J W, Elliott M, Ramachandran R, eds. Coasts and Estuaries. Amsterdam: Elsevier, 2019, 427-442.
|
| [4] |
Jackson J B C, Kirby M X, Berger W H, Bjorndal K M, Botsford L W, Bourque B J, Bradbury R H, Cooke R, Erlandson J, Estes J A, Hughes T P, Kidwell S, Lange C B, Lenihan H S, Pandolfi J M, Peterson C H, Steneck R S, Tegner M J, Warner R R. Historical overfishing and the recent collapse of coastal ecosystems. Science, 2001, 293(5530): 629-637. DOI:10.1126/science.1059199 |
| [5] |
Beck M W, Brumbaugh R D, Airoldi L, Carranza A, Coen L D, Crawford C, Defeo O, Edgar G J, Hancock B, Kay M C, Lenihan H S, Luckenbach M W, Toropova C L, Zhang G F, Guo X M. Oyster reefs at risk and recommendations for conservation, restoration, and management. Bioscience, 2011, 61(2): 107-116. DOI:10.1525/bio.2011.61.2.5 |
| [6] |
房恩军, 李雯雯, 于杰. 渤海湾活牡蛎礁(Oyster reef)及可持续利用. 现代渔业信息, 2007, 22(11): 12-14. DOI:10.3969/j.issn.1004-8340.2007.11.003 |
| [7] |
耿秀山, 傅命佐, 徐孝诗, 李培英. 现代牡蛎礁发育与生态特征及古环境意义. 中国科学B辑, 1991(8): 867-875. |
| [8] |
顾勇, 齐德利, 葛云健, 于蓉, 张忍顺. 江苏小庙洪牡蛎礁生态评价与保护区建设. 海洋科学, 2005, 29(3): 42-47. DOI:10.3969/j.issn.1000-3096.2005.03.010 |
| [9] |
全为民, 安传光, 马春艳, 黄厚见, 成伟, 王云龙, 沈新强, 陈亚瞿. 江苏小庙洪牡蛎礁大型底栖动物多样性及群落结构. 海洋与湖沼, 2012, 43(5): 992-1000. |
| [10] |
全为民, 周为峰, 马春艳, 冯美, 周振兴, 唐峰华, 吴祖立, 范瑞良, 王云龙, 包小松, 沈辉, 成伟. 江苏海门蛎岈山牡蛎礁生态现状评价. 生态学报, 2016, 36(23): 7749-7757. |
| [11] |
俞鸣同, 藤井昭二, 坂本亨. 福建深沪湾牡蛎礁的成因分析. 海洋通报, 2001, 20(5): 24-30. DOI:10.3969/j.issn.1001-6392.2001.05.004 |
| [12] |
姚庆元. 福建金门岛东北海区牡蛎礁的发现及其古地理意义. 台湾海峡, 1985(1): 108-109. |
| [13] |
孙万胜, 温国义, 白明, 李彤, 张萍, 董学鹏, 于洁. 天津大神堂浅海活牡蛎礁区生物资源状况调查分析. 河北渔业, 2014(9): 23-26, 76-76. DOI:10.3969/j.issn.1004-6755.2014.09.008 |
| [14] |
范昌福, 裴艳东, 田立柱, 王福, 刘志广, 王宏. 渤海湾西部浅海区活牡蛎礁调查结果及资源保护建议. 地质通报, 2010, 29(5): 660-667. DOI:10.3969/j.issn.1671-2552.2010.05.004 |
| [15] |
张忍顺. 江苏小庙洪牡蛎礁的地貌-沉积特征. 海洋与湖沼, 2004, 35(1): 1-7. DOI:10.3321/j.issn:0029-814X.2004.01.001 |
| [16] |
季荣耀, 陆永军, 左利钦. 渤海湾曹妃甸深槽形成机制及稳定性分析. 地理学报, 2011, 66(3): 348-355. |
| [17] |
周毅, 许帅, 徐少春, 岳世栋, 顾瑞婷, 张晓梅, 许敏, 张玉, 张云岭, 张振海. 中国温带海域新发现较大面积(大于0.5 km2)海草床: Ⅱ声呐探测技术在渤海唐山沿海海域发现中国面积最大的鳗草海草床. 海洋科学, 2019, 43(8): 50-55. |
| [18] |
岳军, Dong Y, 张宝华, 韩芳, 赵希涛, 刘冬雁, 段焕春, 张百鸣, 赵长荣. 渤海湾西北岸的几道牡蛎礁. 地质学报, 2012, 86(8): 1175-1187. DOI:10.3969/j.issn.0001-5717.2012.08.001 |
| [19] |
黎刚, 孙祝友. 曹妃甸老龙沟动力地貌体系及演化. 海洋地质与第四纪地质, 2011, 31(1): 11-19. |
| [20] |
Chang A L, Deck A K, Sullivan L J, Morgan S G, Ferner M C. Upstream-downstream shifts in peak recruitment of the native Olympia oyster in San Francisco bay during wet and dry years. Estuaries and Coasts, 2018, 41(1): 65-78. DOI:10.1007/s12237-016-0182-1 |
| [21] |
Grossman S K, Grossman E E, Barber J S, Gamblewood S K, Crosby S C. Distribution and transport of Olympia oyster Ostrea lurida larvae in northern Puget Sound, Washington. Journal of Shellfish Research, 2020, 39(2): 215-233. DOI:10.2983/035.039.0204 |
| [22] |
徐凤山, 黄修明. 中国近海牡蛎超科的新记录. 海洋科学集刊, 1993(34): 175-179. |
| [23] |
Quan W M, Humphries A T, Shen X Q, Chen Y Q. Oyster and associated benthic macrofaunal development on a created intertidal oyster (Crassostrea ariakensis) reef in the Yangtze river estuary, China. Journal of Shellfish Research, 2012, 31(3): 599-610. DOI:10.2983/035.031.0302 |
| [24] |
Luckenbach M W, Coen L D, Ross P G Jr, Stephen J A. Oyster reef habitat restoration: relationships between oyster abundance and community development based on two studies in Virginia and South Carolina. Journal of Coastal Research, 2005(40): 64-78. |
| [25] |
Erbland P J, Ozbay G. A comparison of the macrofaunal communities inhabiting a Crassostrea virginica oyster reef and oyster aquaculture gear in Indian River Bay, Delaware. Journal of Shellfish Research, 2008, 27(4): 757-768. DOI:10.2983/0730-8000(2008)27[757:ACOTMC]2.0.CO;2 |
| [26] |
Rodney W S, Paynter K T. Comparisons of macrofaunal assemblages on restored and non-restored oyster reefs in mesohaline regions of Chesapeake Bay in Maryland. Journal of Experimental Marine Biology and Ecology, 2006, 335(1): 39-51. DOI:10.1016/j.jembe.2006.02.017 |
| [27] |
Quan W M, Zhu J X, Ni Y, Shi L Y, Chen Y Q. Faunal utilization of constructed intertidal oyster (Crassostrea rivularis) reef in the Yangtze River estuary, China. Ecological Engineering, 2009, 35(10): 1466-1475. DOI:10.1016/j.ecoleng.2009.06.001 |
| [28] |
Quan W M, Fan R L, Li N N, Sun Z Y, Jiang W, Wang T N, Shi W J, Li X Y. Seasonal and temporal changes in the Kumamoto oyster Crassostrea sikamea population and associated benthic macrofaunal communities at an intertidal oyster reef in China. Journal of Shellfish Research, 2020, 39(2): 207-214. DOI:10.2983/035.039.0203 |
| [29] |
梁淼, 姜倩, 李德鹏, 柳圭泽, 陈兆林, 崔雷, 于大涛. 曹妃甸近岸海域大型底栖动物群落特征. 水产科学, 2019, 38(4): 479-491. |
| [30] |
Quan W M, Fan R L, Wang Y L, Humphries A T. Long-term oyster recruitment and growth are not influenced by substrate type in China: Implications for sustainable oyster reef restoration. Journal of Shellfish Research, 2017, 36(1): 79-86. DOI:10.2983/036.036.0110 |
| [31] |
全为民, 冯美, 周振兴, 吴祖立, 唐峰华, 王云龙, 包小松, 沈辉, 成伟. 江苏海门蛎岈山牡蛎礁恢复工程的生态评估. 生态学报, 2017, 37(5): 1709-1718. |