 2022, Vol. 42
2022, Vol. 42文章信息
- 韦光富, 包玉, 王志泰, 陈信同, 于子涵, 葛虹艺
- WEI Guangfu, BAO Yu, WANG Zhitai, CHEN Xintong, YU Zihan, GE Hongyi
- 多山城市遗存山体植物多样性斑块效应研究
- Study on patch effect of plant diversity in urban remnant mountains of multi-mountain city
- 生态学报. 2022, 42(24): 10011-10028
- Acta Ecologica Sinica. 2022, 42(24): 10011-10028
- http://dx.doi.org/10.5846/stxb202202080314
-
文章历史
- 收稿日期: 2022-02-08





2. 贵州大学风景园林规划设计研究中心, 贵阳 550025
2. Landscape Architecture Planning and Design Research Center, Guizhou University, Guiyang 550025, China
中国是一个多山的国家, 山地约占国土面积的67%, 分布在山地区域的城市约占全国城市总数的一半以上[1]。多山地区在山间平缓地带发展起来的城市, 随着城市不断的扩张, 周边不容易开发建设的山体, 先后被城市建设用地隔离、包围, 最终镶嵌于城市建成环境中, 形成岛屿状城市遗存自然山体残余生境[2—3]。城市规模与格局的变化直接或间接地影响城市及其区域的生态环境, 使原本脆弱的城市生态环境不断趋于恶化[4]。城市内部和周围生物多样性的分布格局急剧变化及植物多样性下降的问题不断突出[5—7]。城市化对城市植物多样性的影响主要表现为城市植物同质化趋势明显, 城市植物多样性保护面临外来物种的入侵和乡土物种的消失问题[8—9]。因此, 在快速城市化的背景下, 如何保护和维护城市生物多样性, 使其在城市建成环境中发挥更好的生态系统服务功能, 已成为当前研究的热点问题。
基于海洋岛屿研究的“岛屿生物地理学理论”认为物种丰富度受到生境斑块的空间特性的影响, 特别是斑块大小和孤立程度[10]。已有大量的研究证实了陆地生态系统中斑块大小和隔离程度等是植物物种丰富度的可靠预测因子[11—14]。而已有陆地生态系统的相关研究主要以荒漠草地斑块或森林斑块为主要研究对象[15—17]。由于研究区域的不同, 残余斑块的大小和形状对其物种丰富度的影响具有一定的差异[18—21]。Raus等[22]研究表明斑块表面积是物种丰富度的最有影响力的变量。Loke等[23]认为, 分形维数、粗糙度指标可以揭示生境及其相关群落之间的重要关系。有研究表明, 不同坡度和不同曲率的地形对生境条件和植物多样性格局有较大的影响[24]。在山地和草原环境中, 从DEM中提取地形表征因子(如坡度、坡向、曲率和海拔)作为斑块的生境特征, 被广泛用于模拟植物物种的分布[25—28]。综上所述, 斑块的形态指标会对植物多样性产生影响, 然而由于研究视角、区域和对象的不同, 研究结果迵异。
以贵州高原为中心的中国南方喀斯特地区是世界喀斯特发育最典型、最复杂、景观与生态类型最多的一个片区, 也是典型的生态脆弱地区[29—31], 以孤峰和峰林为主的特殊地貌形态, 使得该区域城市扩展过程中大量规模不等的喀斯特山丘以岛屿或类岛屿状的(半)自然残余生境遗存在城市建成环境中, 形成了喀斯特“城市遗存山体”(Urban Remnant Mountains, URMs)生境[2, 32]。城市遗存山体斑块作为多山城市的重要生态斑块, 是城市中宝贵的自然资源, 可以为城市提供多种生态系统服务[33], 特别是在维持本土生物多样性方面具有十分重要的意义[34]。与一般的残余生境不同, 城市遗存山体不仅具有突起于地面的三维立体特征, 而且长期处于人工干扰环境中, 其植物多样性在响应各种城市人工干扰过程中, 是否存在特殊的斑块效应, 关于这一科学问题的相关研究非常薄弱。综上所述, 本研究以典型的多山城市建成区为研究区, 以其中城市遗存山体为研究对象, 分析城市遗存山体植物多样性的斑块效应。旨在探讨以下问题:(1)城市遗存山体的植物群落物种多样性是否存在斑块效应?(2)城市遗存山体斑块的哪些特征指标对其植物多样性的影响显著?
1 材料与方法 1.1 研究区概况与研究对象贵阳市位于贵州省中部, 地处北纬26°11′—26°55′、东经106°07′—107°17′之间。总地势西南高、东北低, 最低处海拔880 m、最高处海拔1659 m。该地区属于以山地和丘陵为主的丘原盆地地区, 其中山地面积4218 km2, 丘陵面积2842 km2。通过贵阳市中心城区建成区遥感影像解译测算, 截至2020年底, 贵阳市中心城区建成区面积为410.11 km2, 常住人口488.19万人。近年来, 快速城市化发展使得贵阳市区土地利用和景观格局变化更加强烈, 形成了“城在山中, 山在城间”的独特景观格局。建成区内镶嵌有539座规模、形状和相对高度各异的城市遗存山体, 大多以孤峰和峰林的形式存在。城市与山体镶嵌景观结构, 使得城市遗存山体的植物群落持续受到城市人工环境的影响[35]。本研究选取了贵阳市建成区内不同大小和形状的23个城市遗存山体样山为研究对象(图 1)。

|
| 图 1 研究区及研究对象 Fig. 1 Study area and objects ST1—ST23为山体编号 |
按东、南、西和北四方向法, 以各样本山体山顶为中心向山脚延伸, 每方向设置3个样点(山顶、山腰、山脚处各一个), 共12个样点。因为部分山体被开挖, 岩石祼露严重, 一些无法取样的样点被移除, 23座样本山体共设置276个有效样点。各样点的植物群落调查样地设置参照《植物社会学理论与方法》[36], 确定最小样地面积为30 m×30 m(900 m2)。各样地设置嵌套型样方, 乔、灌、草分别设置5个调查样方。其中, 乔木样方大小10 m×10 m, 灌木样方大小3 m×3 m, 草本样方大小1 m×1 m(图 2)。样地调查记录内容主要包括: (1)植物种类、多度等信息;(2)生境因子如海拔、坡向、坡度、坡位等。

|
| 图 2 城市遗存山体样地、样方设置示意图 Fig. 2 Schematic diagram of urban remnant mountains (URMs) sample site and sample plot setting |
植物多样性指数选取Shannon-Wiener指数(H′)、Simpson指数(D)和Pielou指数(Jh), 量化植物物种数量、结构和分布均匀程度;Margalef物种丰富度指数(R), 反映植物群落中物种的丰富程度[37]。分别对城市遗存山体植物群落的整体植物(含乔、灌、草和藤等各种植物)、乔木层、灌木层、草本层进行植物物种多样性测算。计算公式为[38]:
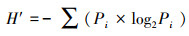
|
(1) |
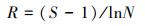
|
(2) |

|
(3) |

|
(4) |
式中, Pi=n/N, Pi为i物种在群落中所占个体的比例, n为i物种的个体数, N为全部物种的个体总数, S为群落中的物种总数。
1.2.3 城市遗存山体斑块特征指标测算以2020年研究区Pleiades卫星影像图(0.5 m空间分辨率, 含30 m分辨率DEM数字高程图)为数据源, 数据由地理空间数据云(http://www.gscloud.cn/)下载获取。利用ENVI 5.3平台对数据进行几何校正、影像裁剪等预处理。通过目视解译结合实地调查验证, 将城市遗存山体景观类型分为16类:乔灌林地、人工园林、人工水体、耕地、绿化边坡、硬化边坡、建筑物、构筑物、硬质铺装、墓地、公路、踩踏道路、硬质道路、人工裸地、泥裸地和岩裸地, 用于计算城市遗存山体景观破碎度、斑块形状、分维数指数。基于ArcGIS 10.2软件建立研究区空间属性数据库。结合城市遗存山体斑块特征, 选择了斑块面积(Pa)、表面积(Sa)、斑块形状指数(PSI)、分维数(Fd)、相对高度(Rh)、破碎度(F)、地表粗糙度(SR)、平均坡度(As)、平面曲率(Hc)、剖面曲率(Pc)指标, 构建城市遗存山体斑块特征指标体系[39]。城市遗存山体斑块特征指标获取及计算方法见表 1。
| 指标 Indices |
获取及计算方法 Methods of obtaining and calculating |
描述及意义 Description and significance |
| 斑块面积Patch area (Pa) | ArcMap 10.2软件→表面分析→ | 所选区域的斑块总面积。 |
| 表面积Surface area (Sa) | 面积和体积 | 所选区域的斑块三维表面积。 |
| 平均坡度 Average slope (As) |
ArcMap 10.2软件→表面分析→坡度 | 该点的切平面与水平地面的夹角, 表示地表面在该点的倾斜程度。 |
| 平面曲率 Horizon curvature (Hc) |
ArcMap 10.2软件→表面分析→曲率 | 平面曲率是指在地形表面上, 具体到任何一点, 指过该点的水平面沿水平方向切地形表面所得的曲线在该点的曲率值[40]。若平面曲率值是正值, 则地形表面是凸面;平面曲率若是负值, 地形表面则是凹面。 |
| 剖面曲率 Profile curvature (Pc) |
剖面曲率是地面上任意一点地表坡度的变化率, 或称为高程变化的二次导数。可以反映局部地形结构, 在地表过程模拟、水土保持等领域都有重要的应用价值[41]。 | |
| 相对高度 Relative height (Rh) |
Rh=Hmax-Hmin | 式中, Rh指某一点的地形相对高度, Hmax指分析窗口内高程的最大值, Hmin指分析窗口内高程的最小值。相对高度是反应地表起伏状况的宏观地形因子, 它的定义是一个特定分析区域内, 所有点的高程值中最大高程值和最小高程值的差[42]。 |
| 地表粗糙度 Surface roughness (SR) |
SR=1/cos(Slope×3.1415926/180) | 反映地表起伏变化与侵蚀程度的指标, 是特定的区域内地球表面积与其投影面积之比[39]。地表粗糙度反映坡面凹凸不平的程度。地表粗糙度对土壤侵蚀的增加、减少作用同时存在。 |
| 斑块形状指数 Patch shape Index (PSI) |
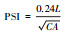 |
式中, PSI表示斑块形状指数, CA表示斑块面积, L表示斑块周长。斑块形状指数是某一不规则的斑块形状与相同面积的规则的图形例如圆或正方形之间的偏离程度[43]。 |
| 分维数 Fractal dimension (Fd) |
Fd=2ln(P/4)/lnA | 式中, Fd表示分维数;P为斑块周长;A为斑块面积。Fd值越大, 表明斑块形状越复杂, Fd值的理论范围为1.0—2.0, 1.0代表形状最简单的正方形斑块, 2.0表示等面积下周边最复杂的斑块[44]。 |
| 破碎度 Fragmentation (F) |
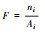
|
式中:F表示破碎度;Ai为景观类型i的面积;A为景观总面积;ni为景观类型i的斑块数。指景观被分割的破碎程度, 它在一定程度上反映了人为因素对景观的干扰程度。F值越大, 表示景观越破碎[45]。 |
在ArcMap 10.2软件中提取平均坡度、三维表面积、斑块面积、相对高度、平面曲率、剖面曲率和地表粗糙度等斑块地形因子指标。运用Fragstats 4.2计算软件对景观指数PSI、F和Fd指标进行计算;在Excel 19.0软件中进行植物群落调查数据库构建和物种多样性各指数计算。采用SPSS 20.0软件进行单因素方差分析和最小显著性差异法(LSD), 比较不同坡向、坡位的植物多样性差异;通过Spearman相关分析, 比较城市遗存山体植物物种多样性指数与城市遗存山体斑块指数之间的关系。采用Origin.2019软件作图, 图中数据为均值±标准差。
2 结果与分析 2.1 城市遗存山体斑块特征由图 3可以看出除了SR、Fd指数外, 其它8个斑块特征指数在23个城市遗存山体间存在明显差异。Pa、Sa、PSI和Fd指数的数值在ST6最高。Hc和Pc指数在ST11最高, 而其它斑块特征指数在不同的城市遗存山体均有较大的差异。SR和Rh指数在ST13最高, F和As指数分别在ST2和ST7最高。Pa、Sa和PSI指数在ST5最低, Rh、As和Hc指数在ST2最低, F指数在ST17最低, SR指数在ST7最低, Fd指数在ST3最低, Pc指数在ST22最低。

|
| 图 3 城市遗存山体斑块特征指标差异 Fig. 3 The difference of patch characteristics indices among URMs |
23座样本城市遗存山体植物群落物种多样性测算结果见表 2。不同城市遗存山体的植物群落整体植物、乔木层、灌木层和草本层的H′、R、D和Jh指数均有显著差异, 且H′指数整体上呈现为草本层>灌木层>乔木层>整体植物, R和D指数表现为整体植物>草本层>灌木层>乔木层, 而Jh指数则表现为乔木层>灌木层>草本层>整体植物。群落的整体植物水平上, 多样性指数H′、R和D在ST16最高, Jh指数在ST15最高;多样性指数H′、R和Jh在ST23最低, D指数在ST3最低。乔木层水平上, 多样性指数H′在ST8最高, R和D在ST22最高, Jh指数在ST15最高;其中ST5的多样性指数H′、R和D最低, ST1的Jh指数最低。灌木层水平上, 多样性指数H′、R和D在ST19最高, 在ST21最低, Jh指数在ST7最高, 在ST9最低。草本层水平上, 多样性指数H′、R和D在ST8最高, Jh指数在ST13最高;多样性指数H′和R在ST2最低, 多样性指数Jh和D在ST23最低。
| 群落组成水平 Levels of community composition |
编号 number |
Shannon-Wiener指数(H′) Shannon-Wiener index |
Margalef物种 丰富度指数(R) Margalef species richness index |
Pielou指数(Jh) Pielou index |
Simpson指数(D) Simpson index |
| 整体植物 | ST1 | 0.91±0.15bcde | 7.00±1.64bcd | 0.33±0.04bcdef | 0.81±0.09cdefg |
| Whole plant | ST2 | 0.90±0.14bcd | 5.85±1.49a | 0.35±0.04cdefgh | 0.83±0.06efghi |
| ST3 | 0.99±0.11fghij | 8.21±1.38ghi | 0.33±0.04bcdef | 0.33±0.03a | |
| ST4 | 1.00±0.20ghij | 7.90±3.16defgh | 0.35±0.03cdefgh | 0.84±0.08ghij | |
| ST5 | 0.98±0.19defghi | 7.89±1.95defg | 0.33±0.05bcde | 0.82±0.12defgh | |
| ST6 | 0.96±0.11defgh | 7.98±1.74efgh | 0.32±0.03b | 0.82±0.05efgh | |
| ST7 | 1.02±0.13hij | 8.13±1.78gh | 0.35±0.03efgh | 0.85±0.06hij | |
| ST8 | 1.05±0.17jk | 8.39±2.21ghi | 0.36±0.03gh | 0.86±0.07ijk | |
| ST9 | 0.98±0.17efghi | 7.83±2.18defg | 0.33±0.04bcd | 0.83±0.09fghi | |
| ST10 | 1.01±0.18ghij | 8.67±1.86hij | 0.33±0.05bcd | 0.83±0.10fghi | |
| ST11 | 0.89±0.17bc | 6.69±1.79abc | 0.33±0.05bc | 0.80±0.10cdef | |
| ST12 | 0.90±0.15bc | 6.70±1.52abc | 0.32±0.04b | 0.79±0.09cde | |
| ST13 | 1.10±0.12kl | 9.00±1.61jk | 0.36±0.03h | 0.88±0.04k | |
| ST14 | 0.96±0.16cdefgh | 7.34±1.69cdef | 0.33±0.04bcdef | 0.83±0.08efghi | |
| ST15 | 0.94±0.20cdefgh | 7.09±2.01bcde | 0.80±0.12i | 0.80±0.12cdefg | |
| ST16 | 1.15±0.12l | 11.01±1.94l | 0.36±0.03h | 0.88±0.05k | |
| ST17 | 1.09±0.11k | 9.11±1.71jk | 0.36±0.03gh | 0.88±0.05jk | |
| ST18 | 0.94±0.25cdef | 7.87±2.22defg | 0.32±0.07b | 0.78±0.17cd | |
| ST19 | 1.05±0.16jk | 9.58±2.13k | 0.34±0.04cdefg | 0.85±0.07hijk | |
| ST20 | 1.00±0.13ghij | 8.05±2.02fgh | 0.35±0.03defgh | 0.85±0.05hijk | |
| ST21 | 0.86±0.17bc | 6.32±1.77ab | 0.32±0.04b | 0.78±0.09c | |
| ST22 | 1.04±0.14ijk | 8.75±1.99ij | 0.35±0.03fgh | 0.86±0.06ijk | |
| ST23 | 0.79±0.19a | 6.09±1.81a | 0.28±0.05a | 0.71±0.14b | |
| 乔木层 | ST1 | 1.32±0.56abcd | 0.94±0.45abcd | 1.12±0.30a | 0.51±0.20ab |
| Tree layer | ST2 | 1.39±0.46bcd | 1.00±0.35bcde | 1.16±0.20abc | 0.54±0.15ab |
| ST3 | 1.46±0.50bcd | 1.05±0.42cdef | 1.23±0.17bcdef | 0.57±0.15b | |
| ST4 | 1.51±0.51d | 1.18±0.39def | 1.25±0.16cdef | 0.59±0.15b | |
| ST5 | 1.15±0.40a | 0.79±0.28a | 1.18±0.21abcde | 0.48±0.15a | |
| ST6 | 1.20±0.42ab | 0.99±0.46abcde | 1.30±0.15f | 0.52±0.13ab | |
| ST7 | 1.47±0.52cd | 0.96±0.40abcd | 1.26±0.17def | 0.58±0.15b | |
| ST8 | 1.53±0.63d | 1.19±0.53ef | 1.21±0.20abcdef | 0.59±0.18b | |
| ST9 | 1.45±0.52bcd | 0.98±0.49abcde | 1.15±0.20ab | 0.55±0.15ab | |
| ST10 | 1.50±0.33d | 0.97±0.33abcd | 1.24±0.16cdef | 0.59±0.10b | |
| ST11 | 1.28±0.51abc | 0.98±0.43abcde | 1.20±0.23abcde | 0.51±0.17ab | |
| ST12 | 1.47±0.40cd | 1.12±0.47def | 1.27±0.11ef | 0.59±0.11b | |
| ST13 | 1.44±0.45bcd | 1.00±0.43bcde | 1.29±0.16f | 0.58±0.13b | |
| ST14 | 1.28±0.48abc | 0.99±0.43abcde | 1.22±0.18abcdef | 0.52±0.14ab | |
| ST15 | 1.15±0.48ab | 1.00±0.38bcde | 1.32±0.09f | 0.50±0.13ab | |
| ST16 | 1.37±0.48abcd | 1.00±0.42bcde | 1.24±0.15cdef | 0.56±0.15b | |
| ST17 | 1.35±0.51abcd | 0.85±0.37ab | 1.20±0.20abcde | 0.55±0.16ab | |
| ST18 | 1.27±0.52abc | 1.04±0.53cde | 1.26±0.20def | 0.52±0.16ab | |
| ST19 | 1.33±0.47abcd | 1.07±0.42def | 1.29±0.15f | 0.55±0.15ab | |
| ST20 | 1.26±0.52ab | 0.9±0.40abc | 1.13±0.27a | 0.52±0.19ab | |
| ST21 | 1.18±0.53ab | 0.84±0.50ab | 1.18±0.26abcd | 0.48±0.17a | |
| ST22 | 1.52±0.53d | 1.24±0.57f | 1.29±0.15f | 0.60±0.14b | |
| ST23 | 1.25±0.45ab | 1.02±0.39bcde | 1.27±0.21def | 0.53±0.15ab | |
| 灌木层 | ST1 | 1.86±0.63cde | 1.38±0.56def | 1.21±0.20defg | 0.64±0.16def |
| Shrub layer | ST2 | 1.57±0.76bc | 1.00±0.44ab | 1.05±0.30ab | 0.55±0.22abc |
| ST3 | 2.34±0.56gh | 1.81±0.66hi | 1.21±0.13defgh | 0.74±0.11gh | |
| ST4 | 1.89±1.01de | 1.43±0.94ef | 1.11±0.22abc | 0.60±0.25cde | |
| ST5 | 2.51±0.41hij | 1.87±0.48i | 1.27±0.08fghi | 0.78±0.06hi | |
| ST6 | 2.45±0.44hi | 1.75±0.45hi | 1.22±0.11efgh | 0.76±0.09hi | |
| ST7 | 2.65±0.65ij | 1.87±0.59i | 1.34±0.34i | 0.76±0.12hi | |
| ST8 | 1.56±0.66bc | 1.08±0.45abc | 1.21±0.18defgh | 0.58±0.18bcd | |
| ST9 | 1.47±0.59ab | 1.03±0.50ab | 1.04±0.24a | 0.53±0.19ab | |
| ST10 | 1.75±0.59cd | 1.13±0.51bcd | 1.20±0.22cdef | 0.61±0.18cde | |
| ST11 | 1.73±0.57cd | 1.28±0.46cde | 1.18±0.23cdef | 0.63±0.14def | |
| ST12 | 1.58±0.61bc | 1.06±0.46abc | 1.18±0.21cdef | 0.59±0.17bcd | |
| ST13 | 2.30±0.59fgh | 1.75±0.62hi | 1.29±0.10ghi | 0.74±0.11gh | |
| ST14 | 1.98±0.70de | 1.47±0.58efg | 1.08±0.24ab | 0.64±0.19def | |
| ST15 | 1.85±0.63cde | 1.21±0.52bcde | 1.22±0.15defgh | 0.65±0.15def | |
| ST16 | 2.70±0.46jk | 2.23±0.63j | 1.25±0.12fgh | 0.81±0.08hi | |
| ST17 | 2.12±0.50efg | 1.58±0.48fgh | 1.17±0.15cde | 0.66±0.25ef | |
| ST18 | 2.44±0.71hi | 1.90±0.66i | 1.24±0.18efgh | 0.74±0.12gh | |
| ST19 | 2.90±0.52k | 2.38±0.61j | 1.30±0.12hi | 0.82±0.10i | |
| ST20 | 2.10±0.60ef | 1.68±0.60ghi | 1.18±0.19cdef | 0.69±0.15fg | |
| ST21 | 1.27±0.57a | 0.86±0.43a | 1.07±0.30ab | 0.49±0.20a | |
| ST22 | 2.36±0.49h | 1.83±0.59i | 1.21±0.14defg | 0.74±0.10gh | |
| ST23 | 1.78±0.56cd | 1.12±0.40bcd | 1.14±0.24bcd | 0.61±0.27cde | |
| 草本层 | ST1 | 2.24±0.59abc | 1.69±0.62bc | 1.07±0.21bcde | 0.70±0.15abc |
| Herb layer | ST2 | 2.11±0.41a | 1.17±0.48a | 1.17±0.15fgh | 0.71±0.08abcd |
| ST3 | 2.65±0.63efg | 1.95±0.44cde | 1.11±0.20defg | 0.75±0.08cdef | |
| ST4 | 2.66±0.57efg | 2.11±0.85def | 1.11±0.14defg | 0.78±0.10efgh | |
| ST5 | 2.43±0.67bcde | 1.74±0.67bc | 1.09±0.24cdef | 0.73±0.15cde | |
| ST6 | 2.47±0.32cde | 1.87±0.53bcd | 1.04±0.10bcd | 0.74±0.06cde | |
| ST7 | 2.20±0.51ab | 1.61±0.58b | 1.07±0.16cde | 0.70±0.11abc | |
| ST8 | 3.19±0.50i | 2.94±0.75h | 1.18±0.12gh | 0.84±0.07h | |
| ST9 | 2.67±0.62efg | 2.07±0.80def | 1.09±0.16def | 0.77±0.12efg | |
| ST10 | 2.78±0.82fgh | 2.43±0.69g | 1.08±0.31cde | 0.75±0.15cde | |
| ST11 | 2.12±0.82a | 1.60±0.59b | 0.99±0.30ab | 0.73±0.17cde | |
| ST12 | 2.49±0.60cde | 1.88±0.62bcd | 1.05±0.18bcd | 0.74±0.13cde | |
| ST13 | 2.99±0.49hi | 2.27±0.67fg | 1.22±0.09h | 0.83±0.06h | |
| ST14 | 2.34±0.66abcd | 1.69±0.59bc | 1.07±0.20bcde | 0.71±0.14bcd | |
| ST15 | 2.50±0.72cde | 1.98±0.69cdef | 1.07±0.23bcde | 0.72±0.17bcde | |
| ST16 | 2.80±0.65fgh | 2.72±0.75h | 1.10±0.23defg | 0.81±0.08fgh | |
| ST17 | 2.88±0.39gh | 2.41±0.55g | 1.14±0.10efg | 0.81±0.06gh | |
| ST18 | 2.35±0.76abcd | 1.93±0.63cde | 1.00±0.27bc | 0.67±0.27ab | |
| ST19 | 2.62±0.57ef | 2.26±0.71fg | 1.07±0.15cde | 0.76±0.10defg | |
| ST20 | 2.53±0.45de | 1.92±0.63cde | 1.13±0.12efg | 0.76±0.08efg | |
| ST21 | 2.31±0.66abcd | 1.85±0.70bcd | 1.00±0.15abc | 0.70±0.14abc | |
| ST22 | 2.63±0.60ef | 2.16±0.69ef | 1.12±0.15defg | 0.76±0.11defg | |
| ST23 | 2.16±0.62a | 1.63±0.61b | 0.93±0.20a | 0.65±0.16a | |
| 表中的数据为均值±标准差;同一列不同字母表示有显著性差异(P < 0.05) | |||||
图 4和图 5结果表明, 不同坡位和坡向群落整体植物、乔木层、灌木层和草本层的植物多样性指数H′、R、D和Jh均无显著性差异。

|
| 图 4 不同坡位植物群落各层次植物多样性指标 Fig. 4 Plant diversity indices of each level of plant community at different slope position 图中同一指标不同字母表示有显著性差异(P < 0.05) |

|
| 图 5 不同坡向植物群落各层次植物多样性指标 Fig. 5 Plant diversity indices of each level of plant community in different slope direction |
由图 6可知, 在城市遗存山体总体水平上, 群落的整体植物多样性指数与山体斑块特征的F指数呈显著负相关, 群落的整体植物的H′、Jh和D指数与山体斑块特征的Pc指数呈显著负相关, Jh指数与Hc指数呈显著负相关。乔木层水平上, 均匀度指数(Jh)与山体斑块特征的Pa和Sa指数呈显著正相关。灌木层和草本层的R指数与山体斑块特征的F指数呈显著负相关。草本层的Jh指数与Hc指数呈显著负相关, H′、R和D指数与Pc指数呈显著负相关。总体上, 植物群落的整体植物和草本层多样性指数与山体斑块特征的Pa、Sa、SR、Fd、Rh和As指数无相关性, 与山体斑块特征的F、Hc和Pc指数有相关性;乔木层的H′、R和D指数与斑块指标无相关, Jh指数与山体斑块特征的Pa和Sa指数存在相关关系;灌木层的R指数与山体斑块特征的F指数有相关性, 与其他斑块指标无相关, 多样性指数H′、Jh和D指数与斑块指标无相关。

|
| 图 6 不同植物水平植物多样性指数与城市遗存山体斑块指数的相关分析 Fig. 6 Correlation Analysis between plant diversity indices of different plant levels and URMs patch indices *为P < 0.05, **为P < 0.01;H′:Shannon-Wiener指数Shannon-Wiener index;D:Simpson指数Simpson index;Jh:Pielou指数Pielou index;R:Margalef物种丰富度指数Margalef species richness index; Pa:斑块面积Patch area;Sa:表面积Surface area;PSI:斑块形状指数Patch shape Index;SR:地表粗糙度Surface roughness;Fd: 分维数Fractal dimension;Hc: 平面曲率Horizon curvature;Pc: 剖面曲率Profile curvature;As: 平均坡度Average slope;Rh: 相对高度Relative height;F: 破碎度Fragmentation |
不同坡位的植物多样性指数与城市遗存山体斑块特征指数之间存在响应关系(图 7)。植物群落整体植物在山脚、山腰和山顶的植物多样性各指数均与F指数呈显著负相关, 山腰处植物多样性各指数均与Pc指数呈显著负相关, Jh和D指数与Hc指数呈显著负相关。乔木层水平上, 山脚处R和D指数与Hc指数呈显著负相关, D指数与Pc指数呈显著负相关, 山腰处植物多样性指数与城市遗存山体斑块指数无相关性, 山顶处H′和D指数与PSI、Fd和F指数呈显著负相关, R指数与Hc和Pc指数呈显著负相关, Jh指数与Pa和Sa指数呈显著正相关。灌木层水平上, 山脚处Jh指数与As指数呈显著正相关, 与其他斑块指标无相关, 山腰处植物多样性指数与城市遗存山体斑块指数无相关性, 山顶处R、Jh和D指数与F指数呈显著负相关, 与其他斑块指标无相关。草本层水平上, 山脚处H′、Jh和D指数与F指数呈显著负相关, 山脚和山腰处H′、R和D指数与Pc指数呈显著负相关, 山腰处R和Jh指数分别与F和Hc呈显著负相关, 山顶处R指数与Rh、Hc和Pc指数呈显著负相关, H′指数与Hc和Pc指数呈显著负相关。总体上, 植物多样性各指数与城市遗存山体斑块指数响应强度在坡位上由高到低顺序为山顶>山脚>山腰。其中山体斑块特征的Pa、Sa和As指数与植物多样性指数呈显著正相关, 山体斑块特征的F、Rh、Fd、PSI、Hc和Pc指数与植物多样性指数呈显著负相关, 山体斑块特征的SR指数与植物多样性无显著相关。

|
| 图 7 不同坡位植物多样性指数与城市遗存山体斑块指数的相关分析 Fig. 7 Correlation analysis between plant diversity indices at different slope position and URMs patch indices |
不同坡向的植物多样性与城市遗存山体斑块指数之间存在响应关系(图 8)。东坡的整体植物Jh指数与SR和As指数呈显著负相关, 乔木层的Jh指数与Pa和Sa指数呈显著正相关, 东坡和西坡的灌木层多样性指数与城市遗存山体斑块指数无显著相关, 草本层的Jh指数与As指数呈显著负相关。南坡的整体植物Jh指数与PSI、Fd和Hc指数呈显著负相关, 草本层的H′和Jh指数分别与Pc和Hc指数呈显著负相关, 整体植物、灌木层的H′和R指数与F指数呈显著负相关;西坡和北坡的整体植物多样性各指数均与F指数呈显著负相关, 西坡整体植物的H′指数与Hc指数呈显著负相关, H′和R指数与Pc指数呈显著负相关, 乔木层的H′、R和D指数与Pc和Hc指数呈显著负相关, 草本层的多样性指数与Pc指数呈显著负相关, H′、Jh和D指数与Hc指数呈显著负相关;北坡的乔木层的H′和R指数与SR指数呈显著负相关, 灌木层的Jh指数和草本的R指数分别与Hc和F指数有相关性, 草本层的H′、R和D指数与Pc呈显著负相关。总体上, 不同坡向的植物多样性各指数与城市遗存山体斑块指数响应强度由高到低顺序为西坡>北坡>南坡>东坡。不同坡向的植物多样性H′、R、Jh和D指数与Pa、Sa、F、Fd、SR、PSI、As、Hc和Pc指数有相关性, 与Rh指数无相关性。

|
| 图 8 不同坡向植物多样性指数与城市遗存山体斑块指数的相关分析 Fig. 8 Correlation analysis between plant diversity indices of different slope direction and patch indices of URMs |
植物群落与地形因子、人类活动、景观环境密切相关[46—48]。本研究结果表明, 城市遗存山体的植物多样性与大部分斑块特征指标呈显著相关, 证实了城市遗存山体的植物群落多样性中存在斑块效应。总体上, 不同层次的植物多样性受到不同斑块指标的影响, 主要体现为植物群落整体植物和草本层水平上物种多样性与F、Hc和Pc指数呈显著负相关, 灌木层水平的R指数与F指数呈显著负相关, 说明生境的破碎化不仅减小了种群的面积、阻碍基因的流动和种群的自由扩散, 还改变环境的物理化学性质, 对斑块内部生物物种多样性产生多方面的影响[49—51]。曾佩枫等表示Hc是一个地区地形聚集和分离程度, 水流经过表面时汇集的可能性[41], 物种多样性与地表径流、土壤侵蚀呈极显著相关性, 物种多样性低, 地表径流量很大, 而物种多样性高, 地表径流量较小[52]。本研究表明植物群落整体物种、灌木层和草本层物种多样性可能受地表径流、土壤侵蚀的影响较大。乔木层水平的Jh指数与Pa和Sa指数显著正相关, 根据“生物多样性的分布格局与理论”能量是影响生物多样性的主要因素[53], 说明城市遗存山体的面积和表面积与大气接触面积越大, 吸收能量越多, 消耗自身能量越少, 对乔木层物种多样性的维持更有利。不同坡位植物多样性与除SR指数外的绝大多数山体斑块指数有相关性, 其响应强度由高到低顺序为山顶>山脚>山腰。相关研究也证实了植物多样性会受地形、人为或自然干扰的影响[54—55], 城市遗存山体山顶水土流失严重、岩石暴露明显, 而山腰和山脚汇集了上坡位冲刷的水土, 随着坡位下降, 山体土壤状越来越好, 故植物多样性高, 且受地形特征影响的程度也较小。不同坡向的植物多样性与城市遗存山体斑块指数响应强度由高到低顺序为西坡>北坡>南坡>东坡。在高楼林立的城市人工环境中, 高大建筑的阴影将会使城市遗存山体南坡由阳坡变为阴坡, 城市照明可能会对山体北坡植物光环境产生影响, 所以城市遗存山体凸曲面三维特征, 使其在城市人工干扰场中的斑块特征效应更为复杂, 后期需要针对光照环境开展更深入的研究才能揭示不同坡向上的斑块特征效应。
城市是社会-经济-自然复合生态系统, 人类是城市环境中的主导因素, 城市环境一旦建成后将是一个持续不断的人工干扰场[56]。城市遗存山体被镶嵌入城后, 长期受到各种尺度上直接或间接的干扰, 各种干扰通过生境斑块作用于其植物多样性的方式、途径和程度各不相同。所以要厘清城市遗存山体植物多样性对城市干扰响应的斑块效应, 还需要开展大量的实证研究, 验证各种假设条件, 如城市遗存山体的功能定位, 各种干扰程度和人为扰动持续的时间等情景下, 城市遗存山体植物多样性的响应机理, 这将是今后深入研究的方向。
3.2 多山城市遗存山体斑块及植物多样性保护策略城市遗存自然山体是城市自然系统的核心, 是多山城市得天独厚的后发资源优势, 发挥着多种无可替代的生态服务功能[57], 也是开展城市建成环境中残余生境生态学理论相关研究的非常重要的理想场所[58]。然而在内部致密化的城市建设过程中, 城市景观格局的剧烈变化, 对城市遗存山体斑块的侵占和破坏依然严重[35, 59—60], 围绕城市遗存山体进行的各种城市建设和低效的公园化利用也严重破坏了山体生态斑块, 致使城市遗存山体生态系统受到强烈的干扰和胁迫, 严重影响了其生态过程及生态系统服务[3, 61]。本研究结果表明:(1)Pa和Sa这些面积相关的山体斑块特征指标与物种多样性呈正相关, 这一定程度上印证了岛屿生物地理学中的种-面积关系, 即斑块越大, 物种多样性水平越高。说明城市遗存山体的斑块面积大小是其维持植物多样性的关键因子。建议在城市国土空间规划和城市绿地生态系统空间配置上, 将人工园林绿地与城市遗存自然山体相结合, 在一些小型山体斑块周围适当布局人工园林绿地, 形成保护缓冲区, 增大城市遗存山体斑块生境面积。(2)城市遗存山体斑块的F、Hc和Pc指数对其植物多样性呈负相关, 说明过度的开发利用导致城市遗存山体斑块内部破碎化, 工程性破坏如山体开挖使平面曲率和剖面曲率增加, 不仅毁坏了植被, 而且加剧了水土流失和滑坡等问题, 进而对城市遗存山体植物多样性产生负面影响。建议正确认识城市遗存山体的生态价值, 从长远角度权衡经济发展和生态保护之间的利弊, 控制对城市遗存山体开发利用的强度, 严禁工程性措施对山体的侵占和破坏。(3)不同坡位植物多样性各指数与城市遗存山体斑块指数响应强度由高到低顺序为山顶>山脚>山腰。调研时发现山顶水土流失严重、岩石裸露明显以及土壤较为瘠薄。建议将城市遗存山体山顶部位做为生态修复的关键部位, 深入开展调查和研究, 制定切实可行的水土保持和修复策略, 恢复山顶部分植被。(4)不同坡向的植物多样性指数与城市遗存山体斑块指数响应强度由高到低顺序为西坡>北坡>南坡>东坡。一般情况下, 在城市复杂的人工环境中, 高层建筑对城市遗存山体的光环境改变最为明显和直接。在调研中发现部分山体因其南边有紧邻的高层建筑, 使其原本是阳坡的南坡变为常年被建设阴影覆盖的阴坡;而北坡却因邻近的建筑和道路等照明而改变夜间光环境。按照一般生态学理论, 南坡和北坡的响应理应更为明显, 这与本研究的结果却不一致。说明城市遗存山体的植物群落生态过程在城市人工环境中持续不断的各种干扰下的响应十分复杂。本研究只是在现象层面探索了城市遗存山体植物多样性对城市人工干扰环境的响应特征, 但其机理机制层面需要更为深入和广泛的研究才能揭示清楚。建议持续深入地开展城市遗存山体生态学基础理论相关研究, 为城市遗存山体生态保护和可持续生态过程的维持提供更为科学的支撑和依据。
4 结论城市遗存山体是城市建成环境中具有明显突起的三维斑块特征的生态斑块, 其植物群落整体植物、乔木层、灌木层和草本层的多样性指数均有显著差异, 存在明显的斑块效应。在城市遗存山体总体水平上, 植物多样性与斑块特征中的F、Pc、Hc、Pa和Sa指数有相关性。不同坡位植物多样性与城市遗存山体斑块指数响应强度由高到低顺序为山顶>山脚>山腰。不同坡向的植物多样性与城市遗存山体斑块指数响应强度由高到低顺序为西坡>北坡>南坡>东坡。城市遗存山体斑块的F、Hc和Pc指数对其植物多样性的影响显著。本研究一定程度上揭示了城市遗存山体植物群落多样性的影响因素, 然而全面深入地揭示城市遗存山体植物多样性的斑块特征效应, 还需要从干扰类型、程度和时间等各方面开展大量的实证研究。多山城市内大量的城市遗存山体不仅能够向城市建成环境提供不可替代的重要的生态系统功能, 更为研究人工干扰场中残余生境生态学提供了天然的理想场所。因此, 在城市化过程中应加强对城市遗存山体生境的保护, 尽可能保持其自然或近自然的生态过程, 使其更好地挥重要的生态服务功能和科学研究价值。
| [1] |
黄光宇. 山地城市学. 北京: 中国建筑工业出版社, 2002.
|
| [2] |
汤娜, 王志泰, 包玉, 陈信同, 马星宇, 韦光富. 城市基质对城市遗存自然山体植物群落物种多样性的影响——以贵阳为例. 生态学报, 2022, 42(15): 6320-6334. |
| [3] |
周寒冰, 王志泰, 王志杰, 包玉. 喀斯特山地城市生态系统服务价值地形梯度响应——以贵阳市中心城区为例. 水土保持研究, 2021, 28(6): 337-347. |
| [4] |
黄瑞建, 张岳恒, 陈波. 下沙新城区城市化过程中不同生境斑块荠菜种群数量特征及生活史变异. 杭州师范大学学报: 自然科学版, 2010, 9(6): 438-444. DOI:10.3969/j.issn.1674-232X.2010.06.009 |
| [5] |
Loiselle A, Pellerin S, Poulin M. Impacts of urbanization and agricultural legacy on taxonomic and functional diversity in isolated wetlands. Wetlands Ecology and Management, 2020, 28(1): 19-34. DOI:10.1007/s11273-019-09692-1 |
| [6] |
Salgado K, Martínez M L, Álvarez-Molina L L, Hesp P, Equihua M, Mariño-Tapia I. Impact of urbanization and landscape changes on the vegetation of coastal dunes along the Gulf of Mexico. Écoscience, 2022, 29(2): 103-116. DOI:10.1080/11956860.2021.1934299 |
| [7] |
McKinney M L. Urbanization, biodiversity, and conservation: the impacts of urbanization on native species are poorly studied, but educating a highly urbanized human population about these impacts can greatly improve species conservation in all ecosystems. Bioscience, 2002, 52(10): 883-890. DOI:10.1641/0006-3568(2002)052[0883:UBAC]2.0.CO;2 |
| [8] |
何荣晓, 杨帆, 闫蓬勃, 梁怀月. 城市土地利用对植物多样性的影响——以海口市为例. 中国城市林业, 2019, 17(4): 12-17. |
| [9] |
雷一东, 金宝玲. 同质化背景下城市植物多样性的保护. 城市问题, 2011(8): 28-32. |
| [10] |
MacArthur R H, Wilson E O. The Theory of Island Biogeography. Princeton: Princeton University Press, 1967.
|
| [11] |
Bender D J, Contreras T A, Fahrig L. Habitat loss and population decline: a meta-analysis of the patch size effect. Ecology, 1998, 79(2): 517-533. DOI:10.1890/0012-9658(1998)079[0517:HLAPDA]2.0.CO;2 |
| [12] |
Fahrig L. Effects of habitat fragmentation on biodiversity. Annual Review of Ecology, Evolution, and Systematics, 2003, 34(1): 487-515. DOI:10.1146/annurev.ecolsys.34.011802.132419 |
| [13] |
Liira J, Jürjendal I, Paal J. Do forest plants conform to the theory of island biogeography: the case study of bog islands. Biodiversity and Conservation, 2014, 23(4): 1019-1039. DOI:10.1007/s10531-014-0650-5 |
| [14] |
Munguía-Rosas M A, Jurado-Dzib S G, Mezeta-Cob C R, Montiel S, Rojas A, Pech-Canché J M. Continuous forest has greater taxonomic, functional and phylogenetic plant diversity than an adjacent naturally fragmented forest. Journal of Tropical Ecology, 2014, 30(4): 323-333. DOI:10.1017/S0266467414000194 |
| [15] |
Maestre F T, Escudero A. Is the patch size distribution of vegetation a suitable indicator of desertification processes?. Ecology, 2009, 90(7): 1729-1735. DOI:10.1890/08-2096.1 |
| [16] |
Du L T, Gong F, Zeng Y J, Ma L L, Qiao C L, Wu H Y. Carbon use efficiency of terrestrial ecosystems in desert/grassland biome transition zone: a case in Ningxia province, northwest China. Ecological Indicators, 2021, 120: 106971. DOI:10.1016/j.ecolind.2020.106971 |
| [17] |
Mohandass D, Campbell M J, Davidar P. Impact of patch size on woody tree species richness and abundance in a tropical montane evergreen forest patches of south India. Journal of Forestry Research, 2018, 29(6): 1675-1687. DOI:10.1007/s11676-018-0592-y |
| [18] |
Svejcar L N, Bestelmeyer B T, Duniway M C, James D K. Scale-dependent feedbacks between patch size and plant reproduction in desert grassland. Ecosystems, 2015, 18(1): 146-153. DOI:10.1007/s10021-014-9818-9 |
| [19] |
Mohandass D, Campbell M J, Hughes A C, Mammides C, Davidar P. The effect of altitude, patch size and disturbance on species richness and density of lianas in montane forest patches. Acta Oecologica, 2017, 83: 1-14. DOI:10.1016/j.actao.2017.06.004 |
| [20] |
Santana L D, Prado-Junior J A, Ribeiro J H C, Ribeiro M A S, Pereira K M G, Antunes K, Carvalho F A, Van Den Berg E. Edge effects in forest patches surrounded by native grassland are also dependent on patch size and shape. Forest Ecology and Management, 2021, 482: 118842. DOI:10.1016/j.foreco.2020.118842 |
| [21] |
Yamaura Y, Kawahara T, Iida S, Ozaki K. Relative importance of the area and shape of patches to the diversity of multiple taxa. Conservation Biology, 2008, 22(6): 1513-1522. DOI:10.1111/j.1523-1739.2008.01024.x |
| [22] |
Raus T, Karadimou E, Dimopoulos P. Taxonomic and functional plant diversity of the Santorini-Christiana island group (Aegean Sea, Greece). Willdenowia, 2019, 49(3): 363-381. DOI:10.3372/wi.49.49308 |
| [23] |
Loke L H L, Ladle R J, Bouma T J, Todd P A. Creating complex habitats for restoration and reconciliation. Ecological Engineering, 2015, 77: 307-313. DOI:10.1016/j.ecoleng.2015.01.037 |
| [24] |
Mörsdorf M A, Ravolainen V T, Yoccoz N G, Thórhallsdóttir T E, Jónsdóttir I S. Decades of recovery from sheep grazing reveal no effects on plant diversity patterns within Icelandic tundra landscapes. Frontiers in Ecology and Evolution, 2021, 8: 602538. DOI:10.3389/fevo.2020.602538 |
| [25] |
Gottfried M, Pauli H, Grabherr G. Prediction of vegetation patterns at the limits of plant life: a new view of the alpine-Nival ecotone. Arctic and Alpine Research, 1998, 30(3): 207-221. DOI:10.2307/1551968 |
| [26] |
Guisan A, Weiss S B, Weiss A D. GLM versus CCA spatial modeling of plant species distribution. Plant Ecology, 1999, 143(1): 107-122. DOI:10.1023/A:1009841519580 |
| [27] |
Deák B, Valkó O, Alexander C, Mücke W, Kania A, Tamás J, Heilmeier H. Fine-scale vertical position as an indicator of vegetation in alkali grasslands-case study based on remotely sensed data. Flora-Morphology, Distribution, Functional Ecology of Plants, 2014, 209(12): 693-697. DOI:10.1016/j.flora.2014.09.005 |
| [28] |
Subedi C K, Rokaya M B, Muzana M B, Z, Timsina B, Gurung J, Chettri N, Baniya C B, Ghimire S K, Chaudhary R P. Vascular plant diversity along an elevational gradient in the Central Himalayas, western Nepal. Folia Geobotanica, 2020, 55(2): 127-140. DOI:10.1007/s12224-020-09370-8 |
| [29] |
Liu C N, Y Huang Y, Wu F, Liu W J, Ning Y Q, Huang Z R, Tang S Q, Liang Y. Plant adaptability in Karst regions. Journal of Plant Research, 2021, 134(5): 889-906. DOI:10.1007/s10265-021-01330-3 |
| [30] |
张俞, 熊康宁, 喻阳华, 许敏, 程雯, 谭代军. 中国南方喀斯特石漠化地区3种经济林土壤呼吸日动态特征. 中南林业科技大学学报, 2019, 39(1): 92-99. |
| [31] |
Qiao Y N, Chen H, Jiang Y J. Quantifying the impacts of lithology on vegetation restoration using a random forest model in a karst trough valley, China. Ecological Engineering, 2020, 156: 105973. DOI:10.1016/j.ecoleng.2020.105973 |
| [32] |
Chen X T, Wang Z T, Bao Y. Cool island effects of urban remnant natural mountains for cooling communities: a case study of Guiyang, China. Sustainable Cities and Society, 2021, 71: 102983. DOI:10.1016/j.scs.2021.102983 |
| [33] |
范春苗, 王志泰, 汤娜, 邓国平. 基于形态学空间格局和空间主成分的贵阳市中心城区生态网络构建. 生态学报, 2022, 42(16): 6620-6632. |
| [34] |
向杏信, 黄宗胜, 王志泰. 喀斯特多山城市空间形态结构与植物群落物种多样性的耦合关系——以安顺市为例. 生态学报, 2021, 41(2): 575-587. |
| [35] |
邢龙, 王志泰, 涂燕茹. 黔中喀斯特城市遗存自然山体景观格局时空演变——以安顺市为例. 生态学报, 2021, 41(4): 1291-1302. |
| [36] |
金振洲. 植物社会学理论与方法. 北京: 科学出版社, 2009.
|
| [37] |
朱兆棋, 刘鲁光, 陈曦, 胡翠华. 银厂沟谢家店滑坡体植被物种多样性和群落稳定性研究. 西华师范大学学报: 自然科学版, 2021, 42(4): 348-354. |
| [38] |
刘守江, 张斌, 杨清伟, 胡翠华, 舒成强. 汶川地震非规范滑坡体上植被的自然恢复能力研究——以彭州银厂沟谢家店子滑坡体为例. 山地学报, 2010, 28(3): 373-378. |
| [39] |
张素梅, 王宗明, 张柏, 宋开山, 刘殿伟, 李方, 任春颖, 黄健, 张惠琳. 利用地形和遥感数据预测土壤养分空间分布. 农业工程学报, 2010, 26(5): 188-194. |
| [40] |
周启鸣, 刘学军. 数字地形分析. 北京: 科学出版社, 2006.
|
| [41] |
曾佩枫, 孙美丽, 常勇. DEM分辨率对地形因子提取影响——以安徽金寨县为例. 河北环境工程学院学报, 2020, 30(4): 90-93. |
| [42] |
隋刚, 郝兵元, 彭林. 利用高程标准差表达地形起伏程度的数据分析. 太原理工大学学报, 2010, 41(4): 381-384, 401-401. |
| [43] |
唐放, 蔡广鹏. 典型喀斯特地区土地利用转型的时空特征及其景观格局演变——以仁怀市为例. 科学技术与工程, 2020, 20(12): 4860-4870. |
| [44] |
郭雨桐, 韩震, 张志杰, 劳国栋. 杭州湾北岸上海段水域景观生态格局变化遥感研究. 海洋湖沼通报, 2021, 43(3): 55-61. |
| [45] |
李建平, 李栋, 周澎, 赵海溟. 华北油田二连分公司蒙古林作业区生态环境质量状况研究. 北方环境, 2010, 22(5): 98-101. |
| [46] |
Chun J H, Ali A, Lee C B. Topography and forest diversity facets regulate overstory and understory aboveground biomass in a temperate forest of South Korea. Science of the Total Environment, 2020, 744: 140783. |
| [47] |
Fu H, Wang X Y, Ge D B, Li W, Tan X Y, Yuan G X, Jeppesen E. Human activities uncouple the cascading effects of hydrological gradients on plant diversity and ecosystem functions in the lake Dongting wetland. Ecohydrology, 2022, 15(1): e2359. |
| [48] |
Coop J D, Massatti R T, Schoettle A W. Subalpine vegetation pattern three decades after stand-replacing fire: effects of landscape context and topography on plant community composition, tree regeneration, and diversity. Journal of Vegetation Science, 2010, 21(3): 472-487. |
| [49] |
曹裕松, 傅声雷, 周兵, 胡文杰, 李慧. 吉安市三种破碎生境中植物多样性研究. 井冈山大学学报: 自然科学版, 2011, 32(4): 117-123. |
| [50] |
Cheng X L, Nizamani M M, Jim C Y, Balfour K, Da L J, Qureshi S, Zhu Z X, Wang H F. Using SPOT data and FRAGSTAS to analyze the relationship between plant diversity and green space landscape patterns in the tropical coastal city of Zhanjiang, China. Remote Sensing, 2020, 12(21): 3477. |
| [51] |
Paciencia M L B, Prado J. Effects of forest fragmentation on Pteridophyte diversity in a tropical rain forest in brazil. Plant Ecology, 2005, 180(1): 87-104. |
| [52] |
王震洪, 段昌群, 侯永平, 杨建松. 植物多样性与生态系统土壤保持功能关系及其生态学意义. 植物生态学报, 2006, 30(3): 392-403. |
| [53] |
Zeng M L, Bao Y, Wang Z T, Huang Z S, Chen X T, Wei W F, Lin X, Li Q. Patch three-dimensional effect for plant species diversity of urban remnant mountain in multi-mountain city. Ecological Indicators, 2022, 140: 108996. |
| [54] |
沈泽昊, 张新时, 金义兴. 三峡大老岭森林物种多样性的空间格局分析及其地形解释. 植物学报, 2000, 42(6): 620-627. |
| [55] |
李芹, 容丽, 王敏. 地形对喀斯特山地植物物种多样性及分布格局的影响. 水土保持通报, 2019, 39(6): 27-34. |
| [56] |
陈利顶, 孙然好, 刘海莲. 城市景观格局演变的生态环境效应研究进展. 生态学报, 2013, 33(4): 1042-1050. |
| [57] |
邢龙. 岩溶地区城市山体自然遗存特征及其生态服务功能综合绩效评价——以贵阳市为例[D]. 贵阳: 贵州大学, 2020.
|
| [58] |
王志泰, 包玉, 王志杰. 城市遗存喀斯特自然山体资源及其公园化利用——以黔中多山城市为例. 北京: 科学出版社, 2022.
|
| [59] |
Kong N, Wang Z T. Response of plant diversity of urban remnant mountains to surrounding urban spatial morphology: a case study in Guiyang of Guizhou Province, China. Urban Ecosystems, 2022, 25(2): 437-452. |
| [60] |
汤娜. 城市遗存山体自然生境及植物群落特征对城市基质的响应——以贵阳市为例[D]. 贵阳: 贵州大学, 2021.
|
| [61] |
汤娜, 王志泰. 黔中城市喀斯特山体遗存植物群落公园化利用响应. 生态学报, 2021, 41(8): 3033-3052. |