 2022, Vol. 42
2022, Vol. 42文章信息
- 郑梦娜, 贾傲, 陈之光, 廣田充, 唐艳鸿, 杜明远, 古松
- ZHENG Mengna, JIA Ao, CHEN Zhiguang, HIROTA Mitsuru, TANG Yanhong, DU Mingyuan, GU Song
- 青藏高原矮火绒草(Leontopodium nanum)叶片性状对海拔高度变化的响应
- Responses of leaf traits of Leontopodium nanum to altitude changes on the Qinghai-Tibetan Plateau
- 生态学报. 2022, 42(24): 10305-10316
- Acta Ecologica Sinica. 2022, 42(24): 10305-10316
- http://dx.doi.org/10.5846/stxb202111143191
-
文章历史
- 收稿日期: 2021-11-14
- 网络出版日期: 2022-07-29





2. 筑波大学, 筑波 3050006;
3. 北京大学城市与环境学院, 北京 100871;
4. 日本农业环境技术研究所, 筑波 3050006
2. University of Tsukuba, Tsukuba 3050006, Japan;
3. College of Urban and Environmental Sciences, Peking University, Beijing 100871, China;
4. Japan Agricultural Environmental Technology Research Institute, Tsukuba 305006, Japan
叶片是植物地上部分与大气环境接触面积最大的器官, 也是植物与外界进行能量和物质交换的最主要场所, 直接影响植物的呼吸、光合和蒸腾过程, 被视为植物与环境之间相互联系的纽带[1]。然而, 叶片性状对环境变化十分敏感, 通常随环境的改变表现出外部形态、表皮特征及解剖结构的差异[2]。近几十年来, 在全球变化背景下, 植物如何适应环境要素变化已成为生态学研究的热点之一[3], 为此国内外学者开展了大量相关研究, 如利用温度、水分、养分、CO2浓度等控制实验研究植物对其变化的响应[4—5]。然而, 全球气候变暖不仅影响植物的生长发育过程, 还导致全球气候异常现象频发(如干旱、洪涝、极端温度事件等极端气象灾害), 在全球变化研究中最受关注[6]。因此, 国内外学者对生长在不同气候类型的植物叶片性状变化开展了诸多相关研究, 探讨温度变化对各种植物叶片性状的影响, 为预测植物对未来气候变暖的响应提供参考依据。例如, Wright等[3]的研究结果表明相较于小叶片, 面积较大的叶片更容易在寒冷地区遭受霜冻的破坏。He等[7]测量分析了分布在北半球热带至寒温带森林的多种植物叶片性状, 发现温度变化会使叶片厚度、栅栏组织和海绵组织厚度等解剖性状发生改变。Zhong等[8]推测与植物茎的解剖性状相比, 叶片解剖性状对于未来全球气候变暖更加敏感。杨继鸿等[9]的研究显示生长于高海拔地区的木本植物叶片会变得小而厚, 叶柄变短, 而气温是叶片性状变异的主导因子。但由于这些研究选取的植物对环境适应方式及所处的气候类型不同, 研究结果也会存在差异。因此, 研究植物叶片性状对温度变化的响应可以充分揭示植物对环境变化适应的方式。
随高山海拔的升高, 温度会发生显著的变化[10]。因此, 目前国内外关于各地区不同物种叶片性状随海拔高度变化的响应已有较多研究[11—14], 且取得了初步进展。但多数研究集中于平原地区, 而对高原地区极端环境中草本植物叶片性状响应的研究相对不足[15], 尤其是在由全球气候变化导致的高山生境气候变暖背景下的研究更为稀少。
青藏高原平均海拔高达4000 m以上, 是全球海拔最高、面积最大、纬度最低、最年轻的高原[16]。其独特的地理与气候条件孕育了许多特有物种, 是高寒生物的自然种质资源库[17]。青藏高原对全球气候变化十分敏感, 自1960年以来的平均升温速率是0.2—0.3℃/10a, 约为全球升温速率的3倍[18], 被称为全球气候变化的“敏感区”与“放大器”。在这样的地理位置及气候条件下, 高原植物常年处于低温环境中, 对于温度变化也非常敏感[19]。冷龙岭位于青藏高原东北部, 具有典型的高原大陆性气候特征, 在小区域范围内沿海拔高度上升垂直气候变化非常明显[20—21], 同时因其坡度较陡能够排除地理范围跨度改变导致的温度变化, 是研究植物对环境变化响应的理想场所。因此可以利用气候环境因子在冷龙岭不同海拔梯度上的剧烈改变, 以空间代替时间的方式, 来研究植物对全球气候变化的响应。
矮火绒草(Leontopodium nanum)是菊科(Compositae)火绒草属(Leontopodium)多年生草本植物, 在中国主要分布在西部及西南部地区。因其强大的生长繁殖和适应能力, 能够适应高海拔寒冷山区的气候条件, 在青藏高原冷龙岭不同海拔高度广泛生长, 对环境变化敏感且变异能力较强[22]。目前已有较多学者研究了矮火绒草的光合特性以及药用价值[23—24], 但关于其叶片性状的研究鲜为报道。
本研究以广泛分布于青藏高原冷龙岭不同海拔的矮火绒草为材料, 通过对其叶片的外部形态、表皮气孔特征和解剖结构进行测量分析, 旨在阐明:(1)矮火绒草叶片表型及解剖结构沿海拔梯度的变化;(2)矮火绒草对气候环境变化的响应与适应对策。通过本研究以期为预测高山植物对未来气候变暖的响应提供参考依据。
1 材料与方法 1.1 研究区概况冷龙岭是祁连山脉东段第一山, 位于青海省海北藏族自治州门源回族自治县北部和甘肃武威、金昌市的交界处, 地理位置为37°02′—38°5′N, 101°3′—102°52′E, 其顶峰——岗什卡峰海拔高达5254.5 m, 终年积雪。该区多年平均降水量为410 mm, 8月为最湿润月(多年平均降水量93 mm), 年均温度为0℃, 7月为最热月(年均温11.4℃), 属于大陆性高原季风气候区, 区域的气温和降水在水平和垂直方向上具有较大差异, 是典型的高寒草甸植被类型, 主要包括菊科(Compositae)、禾本科(Gramineae)、龙胆科(Gentianaceae)、毛茛科(Ranunculaceae)等植物[20, 25]。
1.2 实验方法 1.2.1 材料采集2019年8月, 在3400—4200 m海拔范围内的冷龙岭采集矮火绒草叶片, 沿垂直梯度每间隔200 m设置一个采样地, 即海拔高度分别为3400、3600、3800、4000 m和4200 m。每个海拔采样地随机设置5个样方, 在各样方随机选取生长健康、发育程度相近的成熟植株, 在植株相同的位置采集完全展开、无病虫害损坏的叶片9片, 每个采样地共采集45个叶片。将其中25片混匀装入自封袋中用于外部形态的测量;并将另外20片放入装有FAA固定液(70%乙醇∶甲醛∶冰乙酸=18∶1∶1)的封闭玻璃小瓶中, 带回实验室放入4℃的冰箱中保存, 用于叶片表皮特征和解剖结构的测量。样品都进行严格的编号。使用GPS定位, 记录各采样地的海拔高度、经纬度等信息(表 1)。
| 序号 Number |
海拔 Altitude/m |
经度 Longitude |
纬度 Latitude |
序号 Number |
海拔 Altitude/m |
经度 Longitude |
纬度 Latitude |
|
| 1 | 3400 | 101°19′52″E | 37°39′54″N | 4 | 4000 | 101°22′19″E | 37°42′27″N | |
| 2 | 3600 | 101°21′33″E | 37°41′46″N | 5 | 4200 | 101°22′33″E | 37°42′43″N | |
| 3 | 3800 | 101°22′06″E | 37°42′11″N |
用万分之一电子天平(BT 124S)称量叶片饱和鲜重(SFW, g), 然后将叶片平铺在扫描仪(CanoScan LiDE300)上进行扫描得到图片文件, 利用ImageJ图像处理软件测量叶长(LL, cm), 叶宽(LW, cm), 叶面积(LA, cm2)和叶周长(LP, cm)。把扫描后的叶片放入80℃恒温干燥箱(DG-202-2BS)中烘干72 h至恒重, 取出后称取叶片干重(DW, g)。结合以上数据计算叶片的长宽比(LL/LW)、比叶面积(SLA, cm2/g)、比叶重(SLW, mg/cm2)和叶干物质含量(LDMC, mg/g), 公式如下:
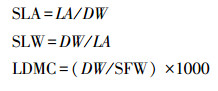
|
采用指甲油涂抹撕取法[26]。取出固定后的叶片, 将透明指甲油均匀涂抹在叶片的上、下表皮, 等待指甲油晾干后, 用镊子撕下已经干燥的指甲油涂抹层, 将其平铺在载玻片上, 制成临时装片。对于各海拔高度随机选取20个视野, 在OLYMPUS BX53光学显微镜的10倍目镜、40倍物镜条件下观察拍照, 利用CellSens软件测量叶片表皮气孔特征。测量指标包括气孔器长度(SL, μm)、气孔器宽度(SW, μm)、气孔器面积(SL, μm2)、气孔密度(SD, 个/mm2)和潜在气孔导度指数(PCI, %)。相关公式如下[27—28]:
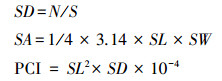
|
式中, N为单个视野内的气孔个数;S为单个视野面积。
1.2.4 叶片解剖结构测量采用常规石蜡切片制片法[29]。将固定好的叶片经过一系列酒精脱水、二甲苯透明、浸蜡并包埋, 再用旋转切片机切片(厚度8—10 μm), 用番红—固绿双重染色法进行染色, 中性树胶封片。在OLYMPUS CX21光学显微镜及图像分析软件(Image Measuring Software Version 6.0)中测量各项解剖学指标, 包括叶片厚度(LT, μm)、上角质层厚度(UCT, μm)、下角质层厚度(LCT, μm)、上表皮厚度(UET, μm)、下表皮厚度(LET, μm)、栅栏组织厚度(PTT, μm)、海绵组织厚度(STT, μm)、主脉厚度(MVT, μm)8项指标。结合以上测量结果计算叶片栅栏系数(CP)、叶片紧密度(CTR)、叶片疏松度(SR), 公式如下[30]:
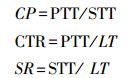
|
气象数据采集自各采样地附近的自动观测仪器, 观测要素主要包括空气温度(Ta)、光合有效辐射(PAR)、降水量(P)等, 每30 min输出一次平均数据。由于仪器故障和环境条件限制, 未能获取全部的连续观测数据(如PAR和P的观测数据不全), 但仍有部分可利用。本研究选取2018年生长季中8月的Ta、PAR和P这3个气象因子进行统计, 分析其随海拔高度的变化规律。发现气温随海拔高度的上升呈明显的直线降低趋势(0.52℃/100 m), 其中3400 m的温度为11.4℃, 而4200 m的气温已降至为7.4℃。而获取的部分数据显示, 随海拔的升高, 辐射稍有增加, 降水略有减少, 但各海拔高度之间的变化差异不显著。
1.3.2 数据分析本研究采用Excel软件计算数据及绘制图表。采用SPSS 22.0统计分析软件对数据进行单因素方差分析(one-way ANOVA)、Pearson相关性分析和线性回归分析。差异显著性分析采用Duncan′s检验法, 并设置P<0.05时为差异显著;相关性分析用双侧检验, 并设置P<0.01时为差异极显著。
1.3.3 可塑性指数(PI)计算根据Valladares等[31], 对于种群间差异显著的叶片解剖结构指标, 计算可塑性指数(PI), 公式如下:
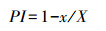
|
式中, x为各种群中的最小平均值;X为各种群中的最大平均值。
2 结果与分析 2.1 叶片外部形态沿海拔梯度的变化随海拔的升高, 矮火绒草叶长的变动较大, 叶宽、叶面积和叶周长均呈先升高后降低的趋势, 峰值均出现在海拔3600 m, 且显著高于其他海拔高度(P<0.05), 比叶面积整体呈显著降低趋势, 而叶长宽比、比叶重和叶干物质含量整体呈显著升高趋势(表 2)。
| 外部形态 External shape |
海拔Altitude/m | ||||
| 3400 | 3600 | 3800 | 4000 | 4200 | |
| LL/cm | 2.98±0.70ab | 4.08±0.58a | 1.85±0.16b | 3.88±0.09a | 2.62±0.20b |
| LW/cm | 0.36±0.02b | 0.83±0.09a | 0.26±0.03b | 0.35±0.02b | 0.33±0.04b |
| LA/cm2 | 0.64±0.06b | 2.37±0.56a | 0.39±0.07b | 0.81±0.05b | 0.59±0.10b |
| LP/cm | 7.14±1.33bc | 12.52±1.54a | 4.62±0.33c | 9.40±0.08b | 6.76±0.42bc |
| LL/LW | 8.26±2.46ab | 5.12±0.26b | 7.14±0.41b | 11.37±0.61a | 8.33±1.03ab |
| SLA/(cm2/g) | 283.48±14.95a | 159.73±12.57b | 259.41±50.65a | 174.42±17.14b | 158.82±16.95b |
| SLW/(mg/cm2) | 3.37±0.01c | 6.61±0.45ab | 4.46±0.69bc | 6.11±0.66ab | 6.85±0.78a |
| LDMC/(mg/g) | 257.27±23.12b | 305.28±17.00b | 376.07±75.10ab | 418.35±46.25ab | 485.92±49.34a |
| LL:叶长Leaf length;LW:叶宽Leaf width;LA:叶面积Leaf area;LP:叶周长Leaf perimeter;LL/LW:叶长宽比Ratio of leaf length to width;SLA:比叶面积Specific leaf area;SLW:比叶重Specific leaf weight;LDMC:叶干物质含量Leaf dry matter content; 每一行不同的小写字母表示在0.05水平下差异显著 | |||||
由表 3可知, 矮火绒草变异系数最大的3个外部形态性状分别是叶面积、叶宽和叶周长。其中叶面积平均值为1.01 cm2, 变化范围为0.18—4.68 cm2, 变异系数为0.90;叶宽的平均值为0.44 cm, 变化范围为0.16—1.12 cm, 变异系数为0.55;叶周长的平均值为8.28 cm, 变化范围为3.48—18.81 cm, 变异系数为0.41。而叶长、叶长宽比、比叶面积、比叶重和叶干物质含量的变异系数分别是0.35、0.36、0.37、0.34、0.31(表 3)。
| 性状 Traits |
平均值 Mean |
最小值 Minimum |
最大值 Maximum |
标准差 Standard deviation |
变异系数 Coefficient of variation |
| LL/cm | 3.09 | 1.25 | 6.49 | 1.07 | 0.35 |
| LW/cm | 0.44 | 0.16 | 1.12 | 0.24 | 0.55 |
| LA/cm2 | 1.01 | 0.18 | 4.68 | 0.91 | 0.90 |
| LP/cm | 8.28 | 3.48 | 18.81 | 3.39 | 0.41 |
| LL/LW | 7.90 | 3.72 | 14.13 | 2.83 | 0.36 |
| SLA/(cm2/g) | 190.18 | 89.86 | 483.93 | 71.01 | 0.37 |
| SLW/(mg/cm2) | 5.88 | 2.07 | 11.13 | 2.01 | 0.34 |
| LDMC/(mg/g) | 382.41 | 193.10 | 621.74 | 118.95 | 0.31 |
其中, 叶宽与海拔高度存在显著的负相关(r=-0.466, P<0.05), 叶长宽比和叶干物质含量分别与海拔高度存在显著(r=0.487, P<0.05)和极显著(r=0.664, P<0.01)的正相关。
2.2 叶片表皮特征沿海拔梯度的变化矮火绒草的表皮气孔特征随海拔梯度的变化趋势如图 1所示。叶片上表皮气孔密度随海拔升高呈增加趋势, 但各海拔之间差异并不显著, 而上表皮气孔器面积和潜在气孔导度指数均随海拔升高呈现先增加后降低的趋势, 且海拔3600 m的值显著高于其它各高度;对于下表皮来说, 气孔密度呈现先增加后降低的变化, 且海拔4000 m的值显著高于其它各高度, 而气孔器面积和潜在气孔导度指数均随海拔升高呈下降的趋势。除上表皮气孔密度、潜在气孔导度指数以及下表皮气孔密度以外, 其余气孔特征均与海拔高度存在极显著(P<0.01)的负相关关系(表 4)。

|
| 图 1 矮火绒草叶片气孔特征随海拔的变化 Fig. 1 Variations of stomatal characteristics of Leontopodium nanum with altitude 柱上不同的小写字母表示在0.05水平下差异显著 |
| 指标 Index |
上表皮Upper epidermis | 下表皮Lower epidermis | |||||||||
| SD | SL | SW | SA | PCI | SD | SL | SW | SA | PCI | ||
| 相关系数 Correlation coefficient |
0.207 | -0.370** | -0.362** | -0.407** | -0.087 | 0.088 | -0.532** | -0.609** | -0.623** | -0.372** | |
| SD:气孔密度Stomatal density;SL:气孔器长度Stomatal length;SW:气孔器宽度Stomatal width;SA:气孔器面积Stomatal apparatus area;PCI:潜在气孔导度指数Potential conductance index; **表示在0.01水平下显著相关 | |||||||||||
由图 2可知, 上表皮的气孔器面积和气孔密度之间没有显著的相关性, 但二者在下表皮存在显著的负相关关系(P<0.05)。而气孔器长度和气孔器宽度、潜在气孔导度指数和气孔密度、潜在气孔导度指数与气孔器面积无论是在上、下表皮之间都存在显著的正相关关系(P<0.05)。

|
| 图 2 矮火绒草叶片各气孔特征之间的关系 Fig. 2 Relationships among stomatal characteristics of Leontopodium nanum 只列出显著的回归关系(P<0.05) |
通过对叶片解剖结构的观察发现, 矮火绒草叶片表面有表皮毛结构且为异面叶, 叶肉组织有栅栏组织和海绵组织的分化。
随海拔的升高, 矮火绒草的叶片厚度、栅栏组织厚度和海绵组织厚度整体均呈增加趋势;栅栏系数呈上升趋势, 在3600 m显著低于其它4个海拔高度;叶片主脉厚度则呈显著下降趋势。叶片上、下表皮厚度的变化趋势相同, 均随海拔升高先下降, 又在4200 m时显著升高;上、下角质层厚度也具有同样的变化趋势, 均随海拔升高波动幅度较大, 呈现先下降后上升再下降的趋势。另外, 叶片紧密度也随海拔升高波动幅度较大, 而叶片疏松度呈先下降后上升的趋势, 在海拔3800、4000 m时显著低于其他海拔高度(图 3)。

|
| 图 3 矮火绒草叶片解剖结构随海拔梯度的变化趋势 Fig. 3 Variation trend of anatomical structures of Leontopodium nanum with altitude changing 柱上不同的小写字母表示在0.05水平下差异显著 |
其中, 矮火绒草的叶片厚度和栅栏组织厚度与海拔高度呈极显著(P<0.01)正相关, 海绵组织厚度和栅栏系数与海拔高度呈显著(P<0.05)正相关;而上、下角质层厚度与海拔高度呈极显著(P<0.01)负相关, 主脉厚度与海拔高度呈显著(P<0.05)负相关(表 5)。
| 指标Index | LT | MVT | UCT | LCT | UET | LET | PTT | STT | CP | CTR | SR |
| 相关系数Correlation coefficient | 0.251** | -0.361* | -0.312** | -0.239** | -0.093 | -0.137 | 0.321** | 0.172* | 0.196* | 0.007 | -0.045 |
| 可塑性指数Plasticity index | 0.37 | 0.37 | 0.48 | 0.48 | 0.30 | 0.44 | 0.28 | 0.36 | 0.32 | 0.25 | 0.21 |
| LT:叶片厚度Leaf thickness; MVT:主脉厚度Main Vein thickness; UCT:上角质层厚度Upper cuticle thickness; LCT:下角质层厚度Lower cuticle thickness; UET:上表皮厚度Upper epidermis thickness; LET:下表皮厚度Lower epidermis thickness; PTT:栅栏组织厚度Palisade tissue thickness; STT:海绵组织厚度Spongy tissue thickness; CP:栅栏系数Coefficient of palisade;CTR:叶片紧密度Cell tense ratio;SR:叶片疏松度Spongy ratio **和*分别表示在0.01和0.05水平下显著相关 | |||||||||||
由矮火绒草叶片的各项解剖结构指标的可塑性指数发现(表 5), 所有数值介于0.21—0.48之间。其中, 上、下角质层厚度的可塑性指数最大, 均为0.48;叶片疏松度的可塑性指数最小, 为0.21。
另外, 对矮火绒草叶片的各项解剖结构指标之间进行相关性分析, 发现除了主脉厚度、上、下角质层厚度和上、下表皮厚度与一些解剖结构指标没有显著相关性, 其余大部分解剖结构指标之间存在极显著(P<0.01)的相关性(表 6)。
| 解剖指标 Anatomical index |
LT | MVT | UCT | LCT | UET | LET | PTT | STT | CP | CTR | SR |
| LT | 1 | 0.434** | 0.002 | -0.038 | -0.095 | -0.273** | 0.482** | 0.668** | -0.219** | -0.429** | -0.200** |
| MVT | 1 | 0.153 | 0.262 | 0.133 | -0.125 | 0.035 | 0.309* | -0.315 | -0.342* | -0.075 | |
| UCT | 1 | 0.468** | 0.058 | -0.043 | 0.023 | -0.042 | -0.013 | 0.056 | -0.057 | ||
| LCT | 1 | -0.127 | -0.127 | -0.119 | -0.097 | -0.078 | -0.077 | -0.086 | |||
| UET | 1 | 0.558** | -0.028 | 0.135 | -0.077 | 0.088 | 0.281** | ||||
| LET | 1 | -0.213** | 0.021 | -0.078 | 0.093 | 0.323** | |||||
| PTT | 1 | 0.279** | 0.554** | 0.585** | -0.161* | ||||||
| STT | 1 | -0.645** | -0.355** | 0.595** | |||||||
| CP | 1 | 0.774** | -0.636** | ||||||||
| CTR | 1 | -0.003 | |||||||||
| SR | 1 | ||||||||||
| **和*分别表示在0.01和0.05水平下显著相关 | |||||||||||
本研究中, 矮火绒草的叶面积、叶宽、叶周长等外部形态性状的变异系数较大, 说明与叶片大小、形状相关的性状对于环境的变化比较敏感。随海拔的升高, 虽然叶面积呈先增加后下降的趋势, 但总体上叶片呈变小、变狭长的趋势, 这与许多研究结果一致[9, 11, 30]。随海拔的升高, 采样地温度呈明显的降低趋势, 较小的叶面积能够降低呼吸与蒸腾带来的消耗, 还能在环境恶劣的高海拔地区中有效减小与空气的接触面积, 避免意外损伤[32];同时在低温环境下狭长的叶形能够增加叶片边缘长度从而增加叶片边界层的阻力, 以降低热量损失[33]。
植物比叶面积、比叶重和叶干物质含量都与环境要素变化密切相关, 因为其代表植物利用环境资源及保存养分的能力, 反映植物所采取的生活史策略[34]。向琳等[35]研究发现鹿角杜鹃(Rhododendron latoucheae)的比叶面积随井冈山海拔升高而减小, 叶干物质含量则增加, Paridari等[36]发现高海拔低温环境会导致欧洲鹅耳枥(Carpinus betulus)有较大的比叶重, 这都与本研究结果相似。随海拔的升高, 矮火绒草的比叶面积减小、比叶重和叶干物质含量增加, 主要是为了适应低温环境:一方面, 植物为了抵御寒冷会减小叶面积[37];另一方面, 土壤温度较低会影响植物根系吸水, 使植物受到水分胁迫, 而较小的比叶面积可以减少水分的散失[38]。总而言之, 较小的比叶面积和较大的比叶重说明植物在资源匮乏、环境较恶劣的环境中将大部分物质用于构建保卫结构从而使自身有更长的生存寿命[39]。除此之外, 较大的比叶重和叶干物质含量说明植物叶片支撑与防御的功能越强、叶片更坚韧[40], 进而有利于植物在高海拔地区的存活。
3.2 叶片表皮特征对海拔变化的响应有学者探究了气孔性状对海拔高度变化的响应, 但所得到的结果有所不同[41—42], 这可能是因为植物自身特征有所差异且影响气孔性状的环境因素较为复杂。本研究中矮火绒草下表皮气孔密度多于上表皮, 能够保证矮火绒草在高原环境中与外界进行气体交换并减少水分蒸腾[43], 而王燕红等[44]对多种禾本科(Poaceae)梯牧草属(Phleum)及其近缘属植物的叶片表皮特征研究结果相反, 其原因可能是单子叶植物与双子叶植物的组织结构有所差异且各自所处的生境不同。随海拔的升高, 矮火绒草上表皮气孔密度没有发生显著变化, 而下表皮气孔密度呈先增加后下降的趋势。Xu和Zhou[45]认为适度的水分胁迫会使气孔密度增加, 而进一步严重的水分胁迫则会通过抑制保卫细胞的分裂降低气孔密度。本研究中随海拔升高, 采样地的温度逐渐降低, 因此矮火绒草会受到因降温引起的生理性干旱胁迫[46], 且程度随海拔升高不断加重, 导致不同海拔高度植物的气孔数量有所差异, 这对本文的实验结果有了合理的解释。
本研究中, 矮火绒草的上表皮气孔器面积随海拔升高呈先增加后降低的趋势;而下表皮气孔器面积随海拔升高呈显著下降趋势, 且与气孔密度有显著的负相关关系。这说明叶片气孔密度增加时会相应减小气孔面积, 小的气孔能更快对环境变化做出反应, 灵活调节气孔开度[47]。除此之外, 本研究还发现矮火绒草叶片上、下表皮的潜在气孔导度指数随海拔升高的变化趋势与其气孔器面积的变化趋势相似, 且与气孔密度和气孔器面积均有显著正相关关系, 意味着气孔导度是由气孔密度和气孔大小共同决定的[48]。上、下表皮的气孔器长度与宽度也存在显著正相关关系, 这在其它研究中也有报道[49], 说明植物在适应环境过程中各气孔特征之间联系紧密。
3.3 叶片解剖结构对海拔变化的响应本研究结果表明矮火绒草的叶厚、栅栏组织厚度和海绵组织厚度随海拔升高呈显著增厚的趋势, 这与郭文文等[50]的研究结果一致。海拔高度变化对上述解剖结构指标的主要影响可能表现在以下几方面:首先, 高寒植物在低温环境中生长, 且温度随海拔升高呈明显的降低趋势, 至4200 m采样地的8月平均气温仅为7.4℃, 远低于同纬度的其它平原地区, 而较厚的叶片有利于植物积累热量, 增强保温作用[50];其次, 通常随海拔的升高, 植物接受的辐射增强[10], 较厚的栅栏组织既能保护叶肉细胞不受强辐射的伤害, 又能保证较高的光合作用效率[51];同时, 增厚的叶片可以防止过度蒸腾, 提高水分利用效率[52];另外, 风速随海拔升高通常呈增大趋势, 而小而厚的叶片还可以降低高海拔地区的风力损伤[53]。综上所述, 矮火绒草叶片增厚是适应高海拔地区低温、强辐射、强风等环境的综合表现。除此之外, 矮火绒草的栅栏系数随海拔升高而显著增大, 对于植物来说, 栅栏系数是一个稳定的结构参数, 该值越大, 说明植物的净光合作用速率越大[54]。
高原草本植物的角质层和表皮细胞均较厚, 能起到隔热、保水、抗机械损伤等功能, 是植物对低温、强辐射以及生理干旱环境的适应[55]。本研究中, 随海拔的升高, 矮火绒草的上、下表皮厚度及上、下角质层厚度都发生显著变化, 但没有规律性。而段喜华等[56]和姜永雷等[57]的研究结果显示植物角质层和表皮厚度随海拔的升高会显著增厚。这一方面可能由于研究区域的环境与海拔梯度有所差异, 另一方面可能是所选植物的不同导致其适应环境变化的方式有所差异。
叶脉能够主导水分在叶片中的传输、散失和保持, 是维持叶片水分平衡的主要结构, 对于水分限制非常敏感。Wang等[41]就指出随海拔升高, 干旱胁迫会使叶脉密度降低, 寇萌等[58]也发现单子叶植物会通过增大叶脉直径来提高水分运输能力, 说明叶脉直径可以作为研究植物适应干旱环境的重要指标。本文研究结果显示, 矮火绒草主脉厚度随海拔的升高呈下降趋势, 这与王元元等[30]对玉龙雪山草血竭(Polygonum paleaceum)的研究结果相反, 其原因可能是不同植物对高海拔不同生境的适应方式不同。本研究中高海拔地区温度较低, 低温会通过降低植物的酶活性和吸水运动导致植物的生理干旱, 进而转化为植物的缺水胁迫问题, 因此矮火绒草会演化出一些能够阻止水分散失的结构来适应缺水胁迫, 如倾向于选择更小的主脉直径[59]。Wang等[41]还指出叶脉的发育与光合作用效率和激素水平有关, Scoffoni等[60]也发现植物叶脉密度还会受到光照强度的影响, 因此后期关于主脉厚度对海拔高度变化的响应还有待进一步研究。
表型可塑性是指植物根据外界环境的变化通过改变自身性状来调节性能的能力, 与其适应环境的能力密切相关[61]。Markesteijn等[62]指出表现出最大可塑性的叶片性状对生长在不同环境中的植物的叶片功能起到非常重要的作用。本文中矮火绒草叶片解剖结构中上、下角质层厚度的可塑性指数最大, 说明角质层厚度会影响植物对气候变化的适应性。冷龙岭矮火绒草可以通过改变角质层的厚度来适应高海拔地区低温、强辐射的环境, 使该物种具有更宽的生态幅, 能在高山环境下广泛分布。
另外, 本研究结果还表明, 虽然矮火绒草各解剖结构指标随海拔高度的变化有所不同, 但大部分解剖结构指标之间存在极显著的相关性。由于功能的相似, 任何植物对环境的适应并不是独立的一个性状发生改变, 而是通过各种各样的性状组合共同来适应不断变化的气候环境[63]。例如叶片厚度与叶片表皮厚度、叶脉之间会存在一定的关联, 反映了这些性状在维持植物水分平衡方面都起到了一定的作用[64]。由此可知, 矮火绒草叶片的部分解剖结构之间存在关联性, 当一种指标增大或减小时, 另外的指标也会随之发生改变。这种协同关联或者权衡关系能帮助植物适应高山恶劣的环境, 是一种生存策略的表现。
4 结论通过对青藏高原冷龙岭3400—4200 m之间5个不同海拔高度的矮火绒草叶片外部形态、表皮气孔特征和解剖结构进行测量分析。随海拔高度的升高, 矮火绒草的叶片整体表现出“小而厚”的趋势, 且整体上叶片性状之间存在相互联系和协同变化。说明矮火绒草主要采取了减小叶面积和增加叶厚度的对策, 帮助自身抵御寒冷、提高水分利用效率, 并将捕获的资源最大化地投入到植物生长发育中, 从而延长寿命, 有利于在寒冷的高山生境中存活。
| [1] |
张进友. 优良的草坪地被植物沿阶草. 草业科学, 2003, 20(2): 69-70. DOI:10.3969/j.issn.1001-0629.2003.02.019 |
| [2] |
李芳兰, 包维楷. 植物叶片形态解剖结构对环境变化的响应与适应. 植物学通报, 2005, 40(S1): 118-127. |
| [3] |
Wright I J, Dong N, Maire V, Prentice I C, Westoby M, Díaz S, Gallagher R V, Jacobs B F, Kooyman R, Law E A, Leishman M R, Niinemets, Reich P B, Sack L, Villar R, Wang H, Wilf P. Global climatic drivers of leaf size. Science, 2017, 357(6354): 917-921. DOI:10.1126/science.aal4760 |
| [4] |
Larbi A, Vázquez S, El-Jendoubi H, Msallem M, Abadía J, Abadía A, Morales F. Canopy light heterogeneity drives leaf anatomical, eco-physiological, and photosynthetic changes in olive trees grown in a high-density plantation. Photosynthesis Research, 2015, 123(2): 141-155. DOI:10.1007/s11120-014-0052-2 |
| [5] |
Guo Y J, Zhao X, Li Y, Li Z, Xiao Q L, Wang Y M, Zhang X F, Ni Y. Environment-driven adaptations of leaf cuticular waxes are inheritable for Medicago ruthenica. Frontiers in Plant Science, 2021, 12: 620245. DOI:10.3389/fpls.2021.620245 |
| [6] |
Luo Z H, Jiang Z G, Tang S H. Impacts of climate change on distributions and diversity of ungulates on the Tibetan Plateau. Ecological Applications: a Publication of the Ecological Society of America, 2015, 25(1): 24-38. DOI:10.1890/13-1499.1 |
| [7] |
He N P, Liu C C, Tian M, Li M L, Yang H, Yu G R, Guo D L, Smith M D, Yu Q, Hou J H. Variation in leaf anatomical traits from tropical to cold-temperate forests and linkage to ecosystem functions. Functional Ecology, 2018, 32(1): 10-19. DOI:10.1111/1365-2435.12934 |
| [8] |
Zhong M Y, Shao X Q, Wu R X, Wei X T, van Logtestijn R S P, Cornelissen J H C. Contrasting altitudinal trends in leaf anatomy between three dominant species in an alpine meadow. Australian Journal of Botany, 2018, 66(5): 448. DOI:10.1071/BT17247 |
| [9] |
杨继鸿, 李亚楠, 卜海燕, 张世挺, 齐威. 青藏高原东缘常见阔叶木本植物叶片性状对环境因子的响应. 植物生态学报, 2019, 43(10): 863-876. DOI:10.17521/cjpe.2019.0174 |
| [10] |
Körner C. The use of 'altitude' in ecological research. Trends in Ecology & Evolution, 2007, 22(11): 569-574. |
| [11] |
蒋琴, 龙文聪, 刘安萱, 肖析蒙, 刘远志, 李晨昊, 杨瑶君. 峨眉山不同海拔高度3种竹叶片结构特征变化研究. 竹子学报, 2018, 37(1): 64-72. DOI:10.3969/j.issn.1000-6567.2018.01.011 |
| [12] |
蒋艾平, 姜景民, 刘军. 檫木叶片性状沿海拔梯度的响应特征. 生态学杂志, 2016, 35(6): 1467-1474. DOI:10.13292/j.1000-4890.201606.017 |
| [13] |
Gratani L, Crescente M F, D'Amato V, Ricotta C, Frattaroli A R, Puglielli G. Leaf traits variation in Sesleria nitida growing at different altitudes in the Central Apennines. Photosynthetica, 2014, 52(3): 386-396. DOI:10.1007/s11099-014-0042-9 |
| [14] |
Krishna M, Winternitz J, Garkoti S C, Penuelas J. Functional leaf traits indicate phylogenetic signals in forests across an elevational gradient in the central Himalaya. Journal of Plant Research, 2021, 134(4): 753-764. DOI:10.1007/s10265-021-01289-1 |
| [15] |
Zhang L, Luo T X, Liu X S, Wang Y. Altitudinal variation in leaf construction cost and energy content of Bergenia purpurascens. Acta Oecologica, 2012, 43: 72-79. DOI:10.1016/j.actao.2012.05.011 |
| [16] |
张镱锂, 李炳元, 郑度. 论青藏高原范围与面积. 地理研究, 2002, 21(1): 1-8. |
| [17] |
孙鸿烈, 郑度, 姚檀栋, 张镱锂. 青藏高原国家生态安全屏障保护与建设. 地理学报, 2012, 67(1): 3-12. |
| [18] |
Li L, Yang S, Wang Z Y, Zhu X D, Tang H Y. Evidence of warming and wetting climate over the Qinghai-Tibet plateau. Arctic, Antarctic, and Alpine Research, 2010, 42(4): 449-457. DOI:10.1657/1938-4246-42.4.449 |
| [19] |
Kullman, L. The changing face of the alpine world. Global Change Newsletter, 2004, 57: 12-14. |
| [20] |
张富广, 曾彪, 杨太保. 近30年祁连山东段冷龙岭地区寒冻荒漠分布范围及其对气候变化的响应. 水土保持研究, 2017, 24(6): 305-312. DOI:10.13869/j.cnki.rswc.2017.06.046 |
| [21] |
Kang S C, Xu Y W, You Q L, Flügel W A, Pepin N, Yao T D. Review of climate and cryospheric change in the Tibetan Plateau. Environmental Research Letters, 2010, 5(1): 015101. DOI:10.1088/1748-9326/5/1/015101 |
| [22] |
许雪贇, 秦燕燕, 曹建军, 李梦天, 龚毅帆, 张小芳. 青藏高原火绒草叶片生态化学计量特征随海拔的变化. 应用生态学报, 2018, 29(12): 3934-3940. DOI:10.13287/j.1001-9332.201812.021 |
| [23] |
何涛, 吴学明, 王学仁, 张改娜, 贾敬芬. 不同海拔火绒草光合特性的研究. 西北植物学报, 2005, 25(12): 2519-2523. DOI:10.3321/j.issn:1000-4025.2005.12.026 |
| [24] |
武彦文, 高文远, 苏艳芳, 贾伟, 段宏泉, 肖培根. 火绒草属植物的化学成分和药理活性研究进展. 中国中药杂志, 2005, 30(4): 245-248. DOI:10.3321/j.issn:1001-5302.2005.04.001 |
| [25] |
杨林, 石莎, 冯金朝, 赵敏杰, 陶季, 蓝智钢. 青海海北高寒草甸植被多样性研究. 中央民族大学学报: 自然科学版, 2008, 17(S1): 126-132. |
| [26] |
刘明智, 努尔巴衣·阿布都沙力克, 潘晓玲. 指甲油涂抹撕取法制取植物叶气孔装片. 生物学通报, 2005, 40(10): 44, 63. |
| [27] |
Wise R R, Sassenrath-Cole G F, Percy R G. A comparison of leaf anatomy in field-grown Gossypium hirsutum and G. barbadense. Annals of Botany, 2000, 86(4): 731-738. DOI:10.1006/anbo.2000.1235 |
| [28] |
Holland N, Richardson A D. Stomatal length correlates with elevation of growth in four temperate species. Journal of Sustainable Forestry, 2009, 28(1/2): 63-73. |
| [29] |
李正理. 植物组织制片学. 北京: 北京大学出版社, 1996.
|
| [30] |
王元元, 齐丹卉, 刘文胜, 梁文斌. 玉龙雪山不同海拔草血竭叶片形态与解剖结构的比较研究. 西北植物学报, 2016, 36(1): 70-77. |
| [31] |
Valladares F, Wright S J, Lasso E, Kitajima K, Pearcy R W. Plastic phenotypic response to light of 16 congeneric shrubs from a Panamanian rainforest. Ecology, 2000, 81(7): 1925-1936. DOI:10.1890/0012-9658(2000)081[1925:PPRTLO]2.0.CO;2 |
| [32] |
Givnish T J, Burkhardt E L, Happel R E, Weintraub J D. Carnivory in the bromeliad Brocchinia reducta, with a cost/benefit model for the general restriction of carnivorous plants to sunny, moist, nutrient-poor habitats. The American Naturalist, 1984, 124(4): 479-497. DOI:10.1086/284289 |
| [33] |
韩威, 刘超, 樊艳文, 赵娜, 叶思阳, 尹伟伦, 王襄平. 长白山阔叶木本植物叶片形态性状沿海拔梯度的响应特征. 北京林业大学学报, 2014, 36(4): 47-53. |
| [34] |
宝乐, 刘艳红. 东灵山地区不同森林群落叶功能性状比较. 生态学报, 2009, 29(7): 3692-3703. DOI:10.3321/j.issn:1000-0933.2009.07.030 |
| [35] |
向琳, 陈芳清, 耿梦娅, 王玉兵, 吕坤, 杨世林. 井冈山鹿角杜鹃群落灌木层植物叶功能性状对海拔梯度的响应. 热带亚热带植物学报, 2019, 27(2): 129-138. |
| [36] |
Paridari I C, Jalali S G, Sonboli A, Zarafshar M, Bruschi P. Leaf macro- and micro-morphological altitudinal variability of Carpinus betulus in the Hyrcanian forest (Iran). Journal of Forestry Research, 2013, 24(2): 301-307. DOI:10.1007/s11676-013-0353-x |
| [37] |
Ch K. The nutritional status of plants from high altitudes. Oecologia, 1989, 81(3): 379-391. DOI:10.1007/BF00377088 |
| [38] |
Hultine K R, Marshall J D. Altitude trends in conifer leaf morphology and stable carbon isotope composition. Oecologia, 2000, 123(1): 32-40. DOI:10.1007/s004420050986 |
| [39] |
武瑞鑫, 邵新庆, 钟梦莹, 潘多, 李超群, 张德罡. 青藏高原东缘披针叶黄华叶片形态特征对海拔变化的响应. 草原与草坪, 2016, 36(2): 1-7. DOI:10.3969/j.issn.1009-5500.2016.02.001 |
| [40] |
Cornelissen J H C, Lavorel S, Garnier E, Díaz S, Buchmann N, Gurvich D E, Reich P B, Steege H T, Morgan H D, van der Heijden M G A, Pausas J G, Poorter H. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Australian Journal of Botany, 2003, 51(4): 335. DOI:10.1071/BT02124 |
| [41] |
Wang X F, Li R Y, Li X Z, Ma F J, Sun B N, Wu J Y, Wang Y K. Variations in leaf characteristics of three species of angiosperms with changing of altitude in Qilian Mountains and their inland high-altitude pattern. Science China Earth Sciences, 2014, 57(4): 662-670. DOI:10.1007/s11430-013-4766-3 |
| [42] |
Kouwenberg L L R, Kurschner W M, McElwain J C. Stomatal frequency change over altitudinal gradients: prospects for paleoaltimetry. Reviews in Mineralogy and Geochemistry, 2007, 66(1): 215-241. DOI:10.2138/rmg.2007.66.9 |
| [43] |
Sam O, Jeréz E, Dell'Amico J, Ruiz-Sanchez M C. Water stress induced changes in anatomy of tomato leaf epidermes. Biologia Plantarum, 2000, 43(2): 275-277. DOI:10.1023/A:1002716629802 |
| [44] |
王燕红, 张学杰, 樊守金. 梯牧草属(Phleum)叶表皮微形态特征研究. 植物研究, 2016, 36(6): 827-837. |
| [45] |
Xu Z Z, Zhou G S. Responses of leaf stomatal density to water status and its relationship with photosynthesis in a grass. Journal of Experimental Botany, 2008, 59(12): 3317-3325. DOI:10.1093/jxb/ern185 |
| [46] |
Xu Z Z, Zhou G S, Wang Y H. Combined effects of elevated CO2 and soil drought on carbon and nitrogen allocation of the desert shrub Caragana intermedia. Plant and Soil, 2007, 301(1/2): 87-97. |
| [47] |
Hetherington A M, Woodward F I. The role of stomata in sensing and driving environmental change. Nature, 2003, 424(6951): 901-908. |
| [48] |
Franks P J, Beerling D J. Maximum leaf conductance driven by CO2 effects on stomatal size and density over geologic time. Proceedings of the National Academy of Sciences of the United States of America, 2009, 106(25): 10343-10347. |
| [49] |
温婧雯, 陈昊轩, 滕一平, 张硕新, 王瑞丽. 太白山栎属树种气孔特征沿海拔梯度的变化规律. 生态学报, 2018, 38(18): 6712-6721. |
| [50] |
郭文文, 卓么草, 方江平, 卢杰, 权红, 任毅华. 藏东南色季拉山薄毛海绵杜鹃叶解剖结构特征与环境适应性. 西北植物学报, 2020, 40(5): 811-818. |
| [51] |
Mendes M M, Gazarini L C, Rodrigues M L. Acclimation of Myrtus communis to contrasting Mediterranean light environments-effects on structure and chemical composition of foliage and plant water relations. Environmental and Experimental Botany, 2001, 45(2): 165-178. |
| [52] |
蔡永立, 宋永昌. 浙江天童常绿阔叶林藤本植物的适应生态学I.叶片解剖特征的比较. 植物生态学报, 2001, 25(1): 90-98, 130. |
| [53] |
郭文文, 卓么草, 周尧治. 西藏高原硬叶柳叶片结构对寒旱环境的适应机制. 西北植物学报, 2019, 39(5): 784-790. |
| [54] |
Higuchi H, Sakuratani T, Utsunomiya N. Photosynthesis, leaf morphology, and shoot growth as affected by temperatures in cherimoya (Annona cherimola Mill.) trees. Scientia Horticulturae, 1999, 80(1/2): 91-104. |
| [55] |
Dongmei L, Liu Z H, Shao H B, Wu G. Improving the eco-environment in the western-China by applying local tree species: issues and implications for global arid areas. African Journal of Biotechnology, 2009, 8(20): 5430-5435. |
| [56] |
段喜华, 孙立夫, 马书荣, 祖元刚. 不同海拔高度泡沙参叶片形态研究. 植物研究, 2003, 23(3): 334-336, 388. |
| [57] |
姜永雷, 邓莉兰, 黄晓霞. 不同海拔川滇高山栎叶片的解剖结构特征. 江苏农业科学, 2015, 43(1): 195-198. |
| [58] |
寇萌, 尹秋龙, 焦菊英. 黄土丘陵沟壑区10种单子叶植物叶片解剖结构及环境适应性. 西北植物学报, 2019, 39(1): 102-109. |
| [59] |
Wang R Z, Huang W W, Chen L, Ma L N, Guo C Y, Liu X Q. Anatomical and physiological plasticity in Leymus chinensis (Poaceae) along large-scale longitudinal gradient in northeast China. PLoS One, 2011, 6(11): e26209. |
| [60] |
Scoffoni C, Kunkle J, Pasquet-Kok J, Vuong C, Patel A J, Montgomery R A, Givnish T J, Sack L. Light-induced plasticity in leaf hydraulics, venation, anatomy, and gas exchange in ecologically diverse Hawaiian lobeliads. The New Phytologist, 2015, 207(1): 43-58. |
| [61] |
Navas M L, Garnier E. Plasticity of whole plant and leaf traits in Rubia peregrina in response to light, nutrient and water availability. Acta Oecologica, 2002, 23(6): 375-383. |
| [62] |
Markesteijn L, Poorter L, Bongers F. Light-dependent leaf trait variation in 43 tropical dry forest tree species. American Journal of Botany, 2007, 94(4): 515-525. |
| [63] |
Wright I J, Groom P K, Lamont B B, Poot P, Prior L D, Reich P B, Schulze E D, Veneklaas E J, Westoby M. Short Communication: leaf trait relationships in Australian plant species. Functional Plant Biology: FPB, 2004, 31(5): 551-558. |
| [64] |
Brodribb T J, Jordan G J, Carpenter R J. Unified changes in cell size permit coordinated leaf evolution. The New Phytologist, 2013, 199(2): 559-570. |