 2022, Vol. 42
2022, Vol. 42文章信息
- 段俊鹏, 王峰, 张卫军, 戴文红, 宋垚彬, 董鸣
- DUAN Junpeng, WANG Feng, ZHANG Weijun, DAI Wenhong, SONG Yaobin, DONG Ming
- 极小种群野生植物密叶红豆杉(Taxus fuana)径向生长对气候的响应
- Responses of radial growth to climate factors in a wild plant with extremely small populations, Taxus fuana
- 生态学报. 2022, 42(24): 10276-10287
- Acta Ecologica Sinica. 2022, 42(24): 10276-10287
- http://dx.doi.org/10.5846/stxb202111023073
-
文章历史
- 收稿日期: 2021-11-02
- 网络出版日期: 2022-07-28

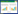




树木的径向生长, 可用树木年轮宽度来度量[1], 是由树种本身的遗传特性, 以及如土壤湿度和矿质元素等立地条件、气候条件(如温度、降水、日照)等的外部环境条件共同作用的结果[2—3]。外部环境因子中气候要素对树轮变化的影响尤为重要[4]。通常在降水量充足、相对湿润的区域, 如我国南方大部分区域, 植物不受干旱胁迫, 植被的生长与温度成正相关。例如, 对位于陕西省南郑县的北亚热带马尾松(Pinus massoniana)树轮宽度的研究发现, 生长季内的温度和降水对马尾松的径向生长以促进作用为主, 树木生长受温度影响最大[5]。在相对干旱的区域, 降水不足是植被生长的限制因子, 植被生长通常与降水量成正相关。因此, 可以通过树轮宽度来研究树木对气候的响应。
气候变暖是一个毋庸置疑的事实[6], 且高海拔地区升温速度最快[7]。由于不同的气候驱动因素、环境异质性、物种差异等, 导致气候变暖对树木生长的影响具有不确定性[8—9]。温度是影响树木生长重要的气候因素, 但目前尚不清楚制约树木生长的关键气候因素[10]。树木在生长过程中对温度敏感性可能是由水分条件介导和影响的[11—12]。对萨加玛塔(喜马拉雅中部珠穆朗玛峰)地区西藏冷杉(Abies spectabilis)和糙皮桦(Betula utilis)的研究发现, 两者对气候变化的响应有一定的相似性, 生长季的温度和降水是影响树木径向生长的主要气候因子[13]。但是温度和降水对径向生长的影响具有复杂性, 同一区域的不同生物气候区对气候的响应也不同。在干旱的西喜马拉雅地区, 树木生长与冬、春两季的降水呈正相关关系, 而在湿润的藏东南地区, 树木生长与冬季温度和春季降水都呈正相关关系[14]。对尼泊尔东部科西河流域干旱河谷松林的乔松(Pinus wallichiana)和西藏长叶松(Pinus roxburghii)研究发现, 松属的生长主要受冬季水分平衡的控制[15];也有研究者通过研究对喜马拉雅中部乔松的生长发现, 生长季早期和全生长季的水分有效性是乔松生长的主要限制因素[16]。目前大多数关于树轮的研究都更多地关注常见种对气候的响应[17], 尤其是在恶劣的外界环境(干旱等)植株生长的响应[18], 而相对较少地关注到珍稀濒危树种, 还不曾关注过极小种群野生树种。
Schickhoff等[7]研究认为气候变暖会导致树木分布到更高海拔。高海拔地区的树木对气候变暖也更敏感, 主要受到低温限制, 导致此地区树木生长对气候响应幅度更大[18—19]。因此, 对高海拔地区树木生长的气候敏感性研究成为全球气候变暖背景下树轮生态学的研究热点[20—21]。喜马拉雅山脉是世界上最长、最高的山地系统, 气候多样, 森林资源丰富[22]。目前, 对于喜马拉雅地区的树轮研究主要涉及三个方面:其一是对单一物种的研究, 如对落叶松(Larix griffithiana)[23]、乔松[24]、喜马拉雅雪松(Cedrus deodara)[25]、糙皮桦[22]、扫帚岩须(Cassiope fastigiata)[26]、秀丽水柏枝(Myricaria elegans)[27]、喜马拉雅冷杉(A. spectabilis)[28—30]等;其二是多个物种对气候的响应进行比较, 如兴都库什-喜马拉雅高原的刺柏(Juniperus)、云杉(Picea)和冷杉(Abies)[19]、尼泊尔中部喜马拉雅地区的西藏红杉(L. griffithii)和喜马拉雅红杉(L. himalaica)[31]等;其三是对气候的重建[32—35]。尽管研究在如像青藏高原那样的高海拔地区的极小种群野生植物径向生长对气候的响应有助于揭示极小种群野生植物生长的限制因子, 但对于同一个物种不同种群的极小种群野生植物径向生长对气候响应的比较研究还相对较少。
密叶红豆杉(Taxus fuana)是喜马拉雅山脉西部特有的濒危植物[36], 是红豆杉科红豆杉属的常绿裸子植物, 为喜马拉雅山脉中仅有的3种红豆杉之一[37], 主要分布于阿富汗东部沿西喜马拉雅地区至中国西藏西南部吉隆的亚热带至温带地区, 印度北部与尼泊尔也均有分布, 我国主要分布在西藏吉隆地区[38—39], 被列入极小种群野生植物物种名录。喜马拉雅山区域海拔较高, 对全球气候变化敏感, 其环境变化信息被良好地记录在生于斯的密叶红豆杉的树轮中。密叶红豆杉年轮清晰, 易于树轮分析。密叶红豆杉的研究对于研究吉隆地区的环境变迁、植被演化等过程具有极高的科研价值[40—41]。密叶红豆杉是喜马拉雅山地区生物多样性的重要组成[42—43], 对其野生种群的认识和保护刻不容缓。因此, 本文以西藏吉隆地区为研究区域, 运用树木年代学的方法, 选取两个不同海拔的密叶红豆杉种群, 研究密叶红豆杉径向生长对气候变化的响应及其在不同海拔上的差异。本文拟回答以下科学问题:(1)西藏吉隆地区密叶红豆杉径向生长的主要限制因子是什么?(2)不同海拔上密叶红豆杉径向生长对气候因子的响应是否一致?旨在探讨密叶红豆杉的生长特性及其对环境因子、气候条件的要求, 为密叶红豆杉保护及其野生种群的繁育和扩展提供理论依据。
1 材料与方法 1.1 研究物种与研究区域 1.1.1 研究物种密叶红豆杉, 又称喜马拉雅密叶红豆杉。植物体为常绿乔木或大灌木, 一年生枝绿色, 干后呈淡褐黄色、金黄色或淡褐色, 二、三年生枝淡褐色或红褐色。叶条形, 排列成彼此重叠的不规则2列, 通常狭直, 密集, 上下等宽, 先端急尖, 基部两侧对称。种子长圆形, 上部两侧微有钝脊, 顶端有突尖。密叶红豆杉产于中国西藏吉隆, 在印度、巴基斯坦、尼泊尔也有分布。该树材质较为坚硬、韧性强度大、颜色绚丽, 常作为工艺品和家具的良好材料, 植株树干挺拔, 树叶碧绿, 观赏价值明显, 具有极为重要的文化价值[44—45]。
1.1.2 研究区域研究样地位于中国西藏自治区日喀则市吉隆县南部吉隆藏布河畔, 在28°21′—28°29′N, 85°13′—85°21′E范围内, 处于亚热带和温带之间的过渡区域, 降水丰富, 太阳辐射强, 干湿季明显。研究区域海拔2600—3200 m, 年平均气温为8—11 ℃, 年均降水量800 mm左右, 年均湿度大于60%;土壤类型常为棕壤以及黄棕壤等[46—47]。密叶红豆杉所在群落中常见的其他组成物种有:高山栎(Quercus semecarpifolia)、密叶红豆杉、华榛(Corylus chinensis)、华山松(Pinus armandii)、朴树(Celtis sinensis)、乔松、云南铁杉(Tsuga dumosa)、西藏冷杉(Abies spectabilis)等[48]。
1.2 树轮采集与分析 1.2.1 树轮采集当地的密叶红豆杉种群曾遭到大量盗伐, 本文将被盗伐后残留的密叶红豆杉树桩作为研究材料。本研究于2017年对6个主要分布的野生种群[41]进行调查时, 在野外用工具将残留树桩截取树盘标本, 做好标记, 进行编号, 带回实验室风干保存。由于盗伐所残留的密叶红豆杉伐桩数量有限以及适合取样的伐桩数量较少, 仅开热和吉普种群(图 1)的样本数为≥15个(表 1), 且两个种群分别处于该地区密叶红豆杉分布的海拔上限和下限。根据之前的调查发现, 密叶红豆杉砍伐植株的径级范围主要为6—30 cm, 具有较好的径级范围代表性, 在空间上表现为一定的聚集性、远离道路[47], 相比于树芯采样的随机性, 树盘的取样依赖于被砍伐树桩的分布, 可能不能兼顾取样随机性原则。保护植物是不能被采伐和扰动, 而从砍伐树桩上取样, 可以避免对密叶红豆杉植株的直接伤害和干扰。因此, 本研究主要针对开热和吉普这两个种群的树盘标本进行研究。

|
| 图 1 两个密叶红豆杉种群取样点分布图 Fig. 1 Geographic distribution of sampling plots for populations of Taxus fuana |
| 种群Population | 开热Kaire | 吉普Jipu |
| 海拔Altitude/m | 3140—3170(高) | 2650—2780(低) |
| 经度Longitude | 85°13′34.12″ | 85°20′15.87″ |
| 纬度Latitude | 28°28′43.35″ | 28°21′51.04″ |
| 种群面积Population area(m×m) | 800×160 | 1, 600×300 |
| 种群大小Population size/株 | 1766 | 约3000 |
| 样本量Samples size | 19 | 15 |
| 平均胸径Average diameter at breast height/cm | 10.5 | 15.8 |
| 序列长度Length of series/a | 1946—2016 | 1918—2017 |
| 共同区间Common interval/a | 1976—2009, 34 | 1971—2011, 41 |
| 平均敏感度Mean sensitivity | 0.1482 | 0.2294 |
| 标准差Standard deviation | 0.3332 | 0.4321 |
| 信噪比Signal to noise ratio | 8.116 | 2.119 |
| 共同区间的样本总体代表性Express population signal | 0.890 | 0.852 |
| 树间相关系数Correlation coefficient among-trees | 0.323 | 0.191 |
将从研究样地带回的密叶红豆杉树轮树盘放置在恒温(22 ℃)通风干燥的实验室内风干90 d, 随后将风干后的密叶红豆杉树盘进行切割, 使其更为规整。运用角磨机将其识别读取面进行粗打磨, 使表面平整。然后, 运用博世Bosch GSS1400平板砂纸机进行精细打磨, 直至树轮树盘表面光滑清晰达到树轮信息识别和读取要求。
使用ESPON V700扫描仪对打磨好的密叶红豆杉树轮树盘进行扫描。利用万深LA-S植物图像分析仪系统(年轮分析测量系统)测量仪进行密叶红豆杉树轮宽度和数量的测量[49—50], 获得树轮宽度和树龄的信息, 树轮宽度测量精度为0.001 mm。运用COFECHA程序[51]检查测量的树轮宽度值的准确性并进行交叉定年, 由此获得每个树轮起止年份的准确信息。交叉定年完成后通过ARSTAN程序[52]建立密叶红豆杉树轮宽度指数标准年表(对订正后的去趋势序列、去除树木生长中与年龄增长相关联的生长趋势得到标准化年表)、差值年表(在标准化年表基础上去掉树木个体由于局部小环境变化而造成的对于后期生长的持续性影响得到差值年表)和自回归年表(估计采样点树木群体所共有的持续性造成的生长量, 再将其加到差值年表上得到的)。结合考虑年表统计特征值, 如样本量(参与构成年表的树轮数量)、序列长度(参与构成年表的树轮年份总长度)、共同区间(构成年表的树轮相同年份)、平均敏感度(反映了年轮宽度序列对气候的敏感度以及气候的短周期变化或高频变化)、标准差(用来体现各年表所指示的树木生长变异程度)、信噪比(非气候因素与气候信号造成的噪音之比)、共同区间的样本总体代表性(指树轮年表能够代表整个群体生长趋势的程度大小)、树间相关系数(不同树轮之间年轮宽度数据变化的同步性和相似性水平), 本研究在不同海拔处分别选择标准年表(图 1)与气候因子进行分析。
1.3 气候数据获取由于吉隆镇周边没有相关气象站, 没有相关仪器监测数据, 所以本研究从Climate China(http://climateap.net/china.aspx)[53]获取了开热和吉普的气候数据, 包括降水量(水分亏缺也会造成叶片气孔关闭或者部分关闭, 从而抑制植物对CO2的吸收)、平均温度(光照强度适宜, 则会促进树木叶片中与光合作用有关的酶)、最高温度(对打破树木的休眠期起重要重要, 温度太低, 植株无法按照正常时期生长)、最低温度(树木为维护正常生命活动, 可能会减少根系、叶片及芽的生理活动);同时, 在对气候数据与树轮年表数据进行相关性分析前, 将错误的和异常的气候数据去除(即, 替换为缺失值)[19]。随后继续检验, 取样点的气候数据(如降水量、温度等)可靠, 没有明显突变值, 检验的结果表明, 该站气候数据可用来代表自然气候的变化[54], 可用于后续的相关性分析等。
如图 1, 两个密叶红豆杉种群样地的各项气象因子变化趋势有明显的一致性, 每年的6月下旬至9月上旬温度最高, 一年中最高温度出现在7月和8月。该时期也是植物的最佳生长期。降水集中在每年的6月下旬至9月下旬, 7月、8月和9月是降水高峰期, 并且在3月有个相对降水小高峰。根据温度变化将植物生长时期进行划分, 3月至5月为生长季前期, 6月至9月为生长季, 10月和11月为生长季后期, 12月至次年2月为休眠期。

|
| 图 2 两个种群1981—2010年气象因子变化 Fig. 2 Changes of meteorological factors in two sites from 1981 to 2010 |
对两个种群的密叶红豆杉年轮宽度指数和对应生境的温度、降水等气候因子进行相关分析。由于气候对树木生长的影响具有明显的“滞后效应”, 尤其是上一生长季末期的气候对下一年树木的生长有很大影响, 所以本研究选取了某年9月至次年11月的气候因子与密叶红豆杉径向生长进行相关性分析。利用主成分分析将气候因子间自相关去除, 再与树轮宽度指数(年表)做逐步回归分析, 并将各主分量的回归系数转换成对应原始气候数据的回归系数, 能够更加准确的反映出样本数据受环境因子的影响程度, 分析软件选用了DendroClim2002[55]。树木生长对气候因子的响应程度用回归系数的大小和正负来表示[25], 通过DendroClim2002软件1000次Bootstrap抽样过程计算相关系数, 在0.05水平下检验相关显著性[55]。部分数据使用SPSS 20进行Pearson相关分析, 分析结果图形绘制使用了SigmaPlot 10.0。
2 结果与分析 2.1 不同种群密叶红豆杉年表基本统计特征的分析对开热种群和吉普种群的树轮宽度序列进行交叉定年并检验通过后, 利用ARSTAN程序建立标准年表。两个密叶红豆杉种群标准年表的特征值如表 1。
两个密叶红豆杉种群的标准年表序列如图 3所示, 最初几年和最后几年的年轮序列波动剧烈, 可能与树轮重复数量较少有关。开热种群和吉普种群密叶红豆杉的年轮生长序列径向增长量在共同年份区间总体变化趋势有相对较高的一致性, 但也相对地存在一定差异, 这种差异反映了两个密叶红豆杉种群的生长环境具有一定的差异性。为了更好地反映两个种群密叶红豆杉的实际生长情况, 本研究选取复本量≥ 10, 且EPS > 0.85的年表区间进行密叶红豆杉径向生长与温度、降水等气候因子的相关关系分析。

|
| 图 3 密叶红豆杉树轮标准年表序列 Fig. 3 Standard chronological sequence of tree-ring of T. fuana |
如图 4所示, 两个种群的密叶红豆杉年轮指数序列与历年月平均温度的相关分析表明, 不同海拔的密叶红豆杉径向生长与历年月平均温度的相关性存在明显差异。开热种群密叶红豆杉的径向生长与某年10月、11月、12月及其次年1月、2月、3月、4月、和8月的平均温度显著正相关, 而吉普种群密叶红豆杉的径向生长与某年9月及其次年3月的平均温度显著负相关。

|
| 图 4 树轮宽度与某年及其次年月平均温度的响应 Fig. 4 Response of tree-ring width to mean monthly temperature in a certain year and the coming year * P < 0.05水平的显著性 |
如图 5所示, 开热种群和吉普种群的密叶红豆杉树轮径向生长与历年月最高温度的相关性也存在明显差异。开热种群密叶红豆杉的径向生长与某年10月、11月、12月及其次年2月、3月、4月、和8月的最高温度显著正相关, 相关系数高于同期吉普种群月平均温度的相关系数。吉普种群密叶红豆杉的径向生长与某年12月及其次年3月的最高温度显著负相关。

|
| 图 5 树轮宽度与某年及其次年月最高温度的响应 Fig. 5 Response of tree-ring width to maximum monthly temperature in a certain year and the coming year * P < 0.05水平的显著性 |
如图 6所示, 开热种群和吉普种群密叶红豆杉的径向生长与历年月最低温度的相关性也存在明显差异。开热种群树轮的径向生长与除了10月的其余月份的最低温度显著正相关, 相关系数高于和同期月平均温度的相关性。吉普种群密叶红豆杉的径向生长与某年9月及其次年3月的最低温度显著负相关。

|
| 图 6 树轮宽度与某年及其次年月最低温度的响应 Fig. 6 Response of tree-ring width to minimum monthly temperature in a certain year and the coming year * P < 0.05水平的显著性 |
如图 7所示, 开热种群密叶红豆杉的径向生长与某年9月及其次年5月、7月、9月的降水量显著正相关, 与某年12月的降水量显著负相关。吉普种群密叶红豆杉的径向生长与各月份的降水量均无显著相关关系。

|
| 图 7 树轮宽度与某年及其次年月降水量的响应 Fig. 7 Response of tree-ring width to monthly precipitation in a certain year and the coming year * P < 0.05水平的显著性 |
吉隆地区的降水集中在每年的6月至9月, 正好覆盖了整个生长季, 且两个海拔生长区域的降水量相差微弱, 以及该地区相对湿润, 因此生长季的降水不是密叶红豆杉生长的主要限制因子。休眠期(12月和1月)的降水对高海拔的密叶红豆杉径向生长具有抑制作用。
3 讨论 3.1 径向生长对历年月平均温度的响应开热种群径向生长与月平均温度的相关分析表明, 上个生长季后期, 休眠期和生长季前期以及生长季8月较高的温度明显促进了开热种群的径向生长。上个生长季后期至当前生长季前期, 较高的温度可使密叶红豆杉免受低温胁迫的伤害, 以较好的状态度过冬季, 为生长季植株恢复快速生长打下生理基础。同时, 也可能是由于开热种群所在区域地势相对平坦, 坡向为西南, 可接收到较为充足的光照, 从而升高局域温度并相应地提高植物细胞的生理活动[56], 进而提高光合作用效率, 储存更多的能量, 促进密叶红豆杉在生长季的径向生长。然而, 坡向、海拔及其相互作用对这些种群间差异影响的相对贡献还需做进一步的研究。这与Gaire等[29]对喜马拉雅中部马纳斯鲁山脉海拔的喜马拉雅山冷杉的径向生长及其与气候的关系研究结果一致。
吉普种群的径向生长与某年9月及其次年3月的平均温度呈显著负相关, 这与Zheng等[19]对兴都库什喜马拉雅地区高海拔的3个优势属的树种(冷杉属、刺柏属和云杉属)生长对气候变化的响应研究结果一致。吉普种群的生长区域海拔相对低, 温度较高, 温度可能不是密叶红豆杉径向生长的主要限制因子, 较高的温度会使密叶红豆杉的生理活动保持在较高水平。生长季末9月的高温会导致密叶红豆杉消耗大量营养物质进行各项生理活动。然而, 该地区9月的阴雨天气较多, 云层较厚, 会导致光照时间不足, 光合作用效率低, 以致于营养物质积累不足, 造成后续生长速度减慢, 同时下一年生长季的生长也受到限制。次年3月温度升高也会加快植株的生理活动, 例如呼吸作用等, 导致营养物质大量消耗, 而同期吉隆地区恰好处于3月降水小高峰期, 阴雨天气及坡向等原因导致日照时间过短, 密叶红豆杉不能充分进行光合作用, 生理活动受到严重影响, 生长缓慢。这也不利于生长季前期的营养物质积累, 对后续生长季植株的径向生长产生负面影响[57]。
3.2 径向生长对历年月最高温度的响应开热种群的径向生长与月最高温度的相关性表明, 上个生长季后期、休眠期(1月除外)和生长季前期以及生长季8月的较高的温度明显促进了开热种群的径向生长。其原因与上述密叶红豆杉径向生长与月平均温度的相关性分析一致。1月的最高温度与密叶红豆杉径向生长也有一定的正相关性, 但是不显著。这可能是1月的最高温度超出了该时期密叶红豆杉生长所需的最适温度范围。
吉普种群的径向生长与某年12月及其次年3月的最高温度呈显著负相关。与开热种群相反。同平均温度的响应关系一样, 吉普种群的生存区域较高的温度会使密叶红豆杉的内部生理活动保持在较高水平。径向生长与某年12月的最高温度呈显著负相关, 可能是因为12月的最高温度较高, 导致密叶红豆杉保持相对较高水平的生理活动, 而未能及时调整进入休眠状态应对1月的降温和严寒, 导致产生冻害, 进而对下一生长季植株的整个生长过程产生负面影响, 生长速度减慢, 降低当年的径向生长[58—59]。径向生长与次年3月的最高温度呈显著负相关的原因同平均温度的原因一致:即3月最高温度较高加快密叶红豆杉的生理活动, 营养物质消耗增大, 而同期阴雨天气较多及坡向原因导致日照时间过短, 不能充分进行光合作用, 不利于生长季前期的能量积累, 降低生长季植株的径向生长速度[60—61]。
3.3 径向生长对历年月最低温度的响应开热种群的径向生长与月最低温度的相关性表明, 开热种群的整个生活史中较高的最低温度明显促进了开热种群的径向生长。其原因与上述密叶红豆杉径向生长与月平均温度的相关性分析一致。10月的最低温度与密叶红豆杉径向生长也有一定的正相关性, 但不显著, 可能是10月的降水量相比于某年7月、8月和9月骤降有关。
吉普种群的径向生长与某年9月和当年3月的最低温度呈显著负相关。同与平均温度和最高温度的相关性一样, 吉普种群密叶红豆杉的生存区域, 温度不是低温限制因子, 最低温度较高会使密叶红豆杉的生理活动增强。某年9月的高温, 导致密叶红豆杉细胞活动消耗大量营养物质, 而9月的降水多、云量大, 以及吉普种群生长区域地形的原因导致入射辐射量小, 光合作用效率低, 而使植株在生长季的生长速度减慢, 径向生长相应降低。3月温度升高也会使植株的生理活动加强, 导致营养物质消耗。同样因为阴雨天气和坡向, 密叶红豆杉光照时间不足, 不能充分进行光合作用, 能量积累少, 不利于后续时期植株的生长, 径向生长降低[59, 62—63]。
3.4 径向生长对历年月温度和降水的综合响应处于吉隆地区密叶红豆杉海拔分布上限的开热种群的径向生长与年平均温度呈极显著正相关, 与各月份的温度也表现出较高的正相关性, 尤其与各月份的最低温度相关性最高, 可能由于海拔高, 环境温度相应降低, 成为影响密叶红豆杉径向生长的限制因子。密叶红豆杉径向生长与雨季各月份的平均温度和最高温度的相关性较低, 可能是雨季云量较多, 光照条件较弱, 密叶红豆杉光合作用速率降低, 营养物质的产生减慢, 而同期温度较高又会使各种生理代谢强度提高, 营养物质的消耗加快, 影响了营养物质的积累, 进而使径向生长受限[64—65]。径向生长与年降水量呈显著正相关, 较高的年际降水有利于密叶红豆杉的径向生长。径向生长与冬季休眠期的降水量呈负相关, Gaire等[29]对喜马拉雅中部马纳斯鲁山脉海拔的喜马拉雅山冷杉的径向生长及其与气候的关系研究中也发现冬季休眠期的降水量抑制了树木的径向生长, 冬季休眠期的降水可能导致温度进一步降低, 使密叶红豆杉产生冷害或冻害, 进而影响到下一生长季的生长过程。这也可能是冬季(某年12月)大量降水使土壤水分环境改变, 导致树木过早萌动, 生长初期用于生长的营养物质被大量消耗, 从而降低树木的径向生长[66]。径向生长与生长季初期(5月)和末期(9月)的降水量呈正相关, 生长季初期的降水有利于早材的形成, 生长季末期的潮湿条件有利于晚材的形成[67], 从而促进密叶红豆杉的径向生长。径向生长在生长高峰期(6月、7月和8月), 与7月的降水量呈正相关, 与6月和8月的降水量无显著相关性, 这与Rai等[28]对喜马拉雅山脉高海拔地区的喜马拉雅山冷杉的径向生长及其与气候的关系研究结果相似。这可能是因为7月的降水多集中在夜晚, 夜晚的降水为植株白天进行光合作用提供了充足的水分, 对密叶红豆杉的生长有促进作用。径向生长与年无霜期呈极显著正相关, 和与温度的相关性一致, 与年际气候水分亏缺不相关, 说明开热地区气候湿润, 干旱程度未达到对密叶红豆杉生长产生胁迫的水平。
处于低海拔地区的吉普种群的径向生长与年平均温度呈负相关, 但不显著。吉普种群生长区域海拔较低, 温度条件较好, 径向生长与温度的响应关系较弱。温度幅度可能超过了密叶红豆杉生长的最适温度范围, 对其生长产生了胁迫, 不利于其生长。径向生长与年降水量呈不显著正相关, 与各月份降水的相关性也较弱。本研究调查时发现, 该区域腐殖质层较厚, 土壤含水率高, 高山积雪融水和腐殖质层涵水使得区域内水流密布、水分充足, 密叶红豆杉的生长几乎不受干旱胁迫, 因此与降水的响应关系较弱。径向生长与年无霜期、气候水分亏缺均成一定程度的负相关, 但不显著。这说明吉普村区域气候相对温暖湿润, 低温和干旱程度通常未达到对密叶红豆杉生长产生胁迫的水平。
通过对比两个海拔密叶红豆杉径向生长与各项气候因子的相关关系, 可以发现开热种群的径向生长与温度、降水的相关性均更高, 受到温度和降水的影响显著大于低海拔的吉普种群, 尤其与月最低温度的相关性最高, 最低温度的升高对开热种群密叶红豆杉的生长有促进作用。而吉普种群的生长区域温度较高, 水分充足, 热量和水分条件较好, 因此与水分无显著相关性, 与次年3月温度呈显著负相关, 与其他月份的温度多成正相关, 但相关性较低。
4 总结与展望温度和降水是分布于高海拔区域密叶红豆杉径向生长的最主要影响因子, 且温度的影响明显大于降水。分布于低海拔的区域的密叶红豆杉对温度和降水的变化不敏感, 据此推测, 影响其生长的最主要因子可能是日照时数。随着全球气候变暖, 高海拔区域密叶红豆杉受到的低温胁迫可能会减弱, 从而表现出更好的生长态势, 适合其生存的海拔区间可能会逐步地扩大, 对其种群扩大有一定的促进作用。
本研究所利用的树轮材料取自于密叶红豆杉遭到盗伐后残留的伐桩, 取样范围受限, 不可能在种群中随机取样, 未能进行径向生长与群落特征、海拔特征相关性的研究。建议今后利用树木生长锥钻取树芯进行更为深入的研究。例如, 将用生长锥获取的树芯进行逐年剥离后, 再测定其碳等稳定同位素, 就可以基于其记录的更多和更精准的信息, 来更深入地研究密叶红豆杉径向生长与气候的响应关系, 更好地探究影响密叶红豆杉生长的各种环境因子, 以及进行吉隆地区的气候重建。
| [1] |
张芬. 青藏高原东北部树轮生态学研究[D]. 兰州: 兰州大学, 2012.
|
| [2] |
Fritts H C. Tree Rings and Climate. New York: Academic Press, 1976.
|
| [3] |
路伟伟, 余新晓, 贾国栋, 李瀚之, 刘自强. 密云山区油松树轮δ13C对气温和降水量变化的响应. 林业科学, 2018, 54(3): 1-7. |
| [4] |
彭剑峰, 勾晓华, 陈发虎, 刘普幸, 张永, 方克艳. 阿尼玛卿山地不同海拔青海云杉(Picea crassifolia)树轮生长特性及其对气候的响应. 生态学报, 2007, 27(8): 3268-3276. DOI:10.3321/j.issn:1000-0933.2007.08.021 |
| [5] |
王瑞丽, 程瑞梅, 肖文发, 封晓辉, 刘泽彬, 王晓荣. 北亚热带马尾松年轮宽度与NDVI的关系. 生态学报, 2011, 31(19): 5762-5770. |
| [6] |
Core Writing Team, Pachauri R K, Meyer L A. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Geneva, Switzerland: IPCC, 2014.
|
| [7] |
Schickhoff U, Bobrowski M, Böhner J, Bürzle B, Chaudhary R P, Gerlitz L, Heyken H, Lange J, Müller M, Scholten T, Schwab N, Wedegärtner R. Do Himalayan treelines respond to recent climate change? An evaluation of sensitivity indicators. Earth System Dynamics, 2015, 6(1): 245-265. DOI:10.5194/esd-6-245-2015 |
| [8] |
Girardin M P, Bouriaud O, Hogg E H, Kurz W, Zimmermann N E, Metsaranta J M, De Jong R, Frank D C, Esper J, Büntgen U, Guo X J, Bhatti J. No growth stimulation of Canada's boreal forest under half-century of combined warming and CO2 fertilization. Proceedings of the National Academy of Sciences of the United States of America, 2016, 113(52): E8406-E8414. |
| [9] |
Babst F, Bouriaud O, Poulter B, Trouet V, Girardin M P, Frank D C. Twentieth century redistribution in climatic drivers of global tree growth. Science Advances, 2019, 5(1): eaat4313. DOI:10.1126/sciadv.aat4313 |
| [10] |
Panthi S, Fan Z X, Bräuning A. Ring widths of rhododendron shrubs reveal a persistent winter warming in the central Himalaya. Dendrochronologia, 2021, 65: 125799. DOI:10.1016/j.dendro.2020.125799 |
| [11] |
Tiwari A, Thapa N, Aryal S, Rana P, Adhikari S. Growth performance of planted population of Pinus roxburghii in central Nepal. Journal of Ecology and Environment, 2020, 44: 31. DOI:10.1186/s41610-020-00171-w |
| [12] |
Jevšenak J, Tychkov I, Gričar J, Levanič T, Tumajer J, Prislan P, Arnič D, Popkova M, Shishov V V. Growth-limiting factors and climate response variability in Norway spruce (Picea abies L.) along an elevation and precipitation gradients in Slovenia. International Journal of Biometeorology, 2021, 65(2): 311-324. DOI:10.1007/s00484-020-02033-5 |
| [13] |
Pandey S, Cherubini P, Saurer M, Carrer M, Petit G. Effects of climate change on treeline trees in Sagarmatha (Mt. Everest, Central Himalaya). Journal of Vegetation Science, 2020, 31(6): 1144-1153. DOI:10.1111/jvs.12921 |
| [14] |
Zheng L L, Gaire N P, Shi P L. High-altitude tree growth responses to climate change across the Hindu Kush Himalaya. Journal of Plant Ecology, 2021, 14(5): 829-842. DOI:10.1093/jpe/rtab035 |
| [15] |
Thapa U K, George S S. Detecting the influence of climate and humans on pine forests across the dry valleys of eastern Nepal's Koshi River basin. Forest Ecology and Management, 2019, 440: 12-22. DOI:10.1016/j.foreco.2019.03.013 |
| [16] |
Gaire N P, Dhakal Y R, Shah S K, Fan Z X, Bräuning A, Thapa U K, Bhandari S, Aryal S, Bhuju D R. Drought (scPDSI) reconstruction of trans-Himalayan region of central Himalaya using Pinus wallichiana tree-rings. Palaeogeography, Palaeoclimatology, Palaeoecology, 2019, 514: 251-264. DOI:10.1016/j.palaeo.2018.10.026 |
| [17] |
王丽丽, 邵雪梅, 黄磊, 梁尔源. 黑龙江漠河兴安落叶松与樟子松树轮生长特性及其对气候的响应. 植物生态学报, 2005, 29(3): 380-385. DOI:10.3321/j.issn:1005-264X.2005.03.006 |
| [18] |
Ettinger A K, Ford K R, HilleRisLambers J. Climate determines upper, but not lower, altitudinal range limits of Pacific northwest conifers. Ecology, 2011, 92(6): 1323-1331. DOI:10.1890/10-1639.1 |
| [19] |
Zheng L L, Shi P L, Song M H, Zhou T C, Zong N, Zhang X Z. Climate sensitivity of high altitude tree growth across the Hindu Kush Himalaya. Forest Ecology and Management, 2021, 486: 118963. DOI:10.1016/j.foreco.2021.118963 |
| [20] |
Seddon A W R, Macias-Fauria M, Long P R, Benz D, Willis K J. Sensitivity of global terrestrial ecosystems to climate variability. Nature, 2016, 531(7593): 229-232. DOI:10.1038/nature16986 |
| [21] |
Hébert R, Lovejoy S. Regional climate sensitivity- and historical-based projections to 2100. Geophysical Research Letters, 2018, 45(9): 4248-4254. DOI:10.1002/2017GL076649 |
| [22] |
Dawadi B, Liang E Y, Tian L D, Devkota L P, Yao T D. Pre-monsoon precipitation signal in tree rings of timberline Betula utilis in the central Himalayas. Quaternary International, 2013, 283: 72-77. DOI:10.1016/j.quaint.2012.05.039 |
| [23] |
Chaudhary V, Bhattacharyya A. Tree ring analysis of Larix griffithiana from the eastern Himalayas in the reconstruction of past temperature. Current Science, 2000, 79(12): 1712-1715. |
| [24] |
Shah S K, Bhattacharyya A, Chaudhary V. Climatic influence on radial growth of Pinus wallichiana in Ziro Valley, northeast Himalaya. Current Science, 2009, 96(5): 697-702. |
| [25] |
Yadav R R. Long-term hydroclimatic variability in monsoon shadow zone of western Himalaya, India. Climate Dynamics, 2011, 36(7): 1453-1462. |
| [26] |
Liang E Y, Liu W W, Ren P, Dawadi B, Eckstein D. The alpine dwarf shrub Cassiope fastigiata in the Himalayas: does it reflect site-specific climatic signals in its annual growth rings?. Trees, 2015, 29(1): 79-86. DOI:10.1007/s00468-014-1128-5 |
| [27] |
Dolezal J, Leheckova E, Sohar K, Dvorsky M, Kopecky M, Chlumska Z, Wild J, Altman J. Annual and intra-annual growth dynamics of Myricaria elegans shrubs in arid Himalaya. Trees, 2016, 30(3): 761-773. DOI:10.1007/s00468-015-1318-9 |
| [28] |
Rai S, Dawadi B, Wang Y F, Lu X M, Ru H, Sigdel S R. Growth response of Abies spectabilis to climate along an elevation gradient of the Manang Valley in the central Himalayas. Journal of Forestry Research, 2020, 31(6): 2245-2254. DOI:10.1007/s11676-019-01011-x |
| [29] |
Gaire N P, Fan Z X, Bräuning A, Panthi S, Rana P, Shrestha A, Bhuju D R. Abies spectabilis shows stable growth relations to temperature, but changing response to moisture conditions along an elevation gradient in the central Himalaya. Dendrochronologia, 2020, 60: 125675. DOI:10.1016/j.dendro.2020.125675 |
| [30] |
Panthi S, Fan Z X, van der Sleen P, Zuidema P A. Long-term physiological and growth responses of Himalayan fir to environmental change are mediated by mean climate. Global Change Biology, 2020, 26(3): 1778-1794. DOI:10.1111/gcb.14910 |
| [31] |
Aryal P C, Dhamala M K, Gaire N P, Bhatta S, Suwal M K, Bhuju D R, Chhetri P K. Tree-ring climate response of two Larix species from the central Nepal Himalaya. Tropical Ecology, 2020, 61(2): 215-225. DOI:10.1007/s42965-020-00082-w |
| [32] |
Cook E R, Krusic P J, Jones P D. Dendroclimatic signals in long tree-ring chronologies from the Himalayas of Nepal. International Journal of Climatology, 2003, 23(7): 707-732. DOI:10.1002/joc.911 |
| [33] |
Yang B, Bräuning A, Liu J, Davis M E, Shao Y. Temperature changes on the Tibetan Plateau during the past 600 years inferred from ice cores and tree rings. Global and Planetary Change, 2009, 69(1/2): 71-78. |
| [34] |
Chen F, Yuan Y J, Wei W S, Yu S L, Shang H M, Zhang T W, Zhang R B, Wang H Q, Qin L. Tree-ring based temperature reconstruction for the west Qinling Mountains (China): linkages to the High Asia, solar activity and Pacific-Atlantic Ocean. Geochronometria, 2014, 41(3): 234-244. DOI:10.2478/s13386-013-0159-9 |
| [35] |
Cánovas J A B, Trappmann D, Shekhar M, Bhattacharyya A, Stoffel M. Regional flood-frequency reconstruction for Kullu district, Western Indian Himalayas. Journal of Hydrology, 2017, 546: 140-149. DOI:10.1016/j.jhydrol.2016.12.059 |
| [36] |
Poudel R C, Möller M, Gao L M, Ahrends A, Baral S R, Liu J, Thomas P, Li D Z. Using morphological, molecular and climatic data to delimitate yews along the Hindu Kush-Himalaya and adjacent regions. PLoS One, 2012, 7(10): e46873. DOI:10.1371/journal.pone.0046873 |
| [37] |
Möller M, Gao L M, Mill R R, Li D Z, Hollingsworth M L, Gibby M. Morphometric analysis of the Taxus wallichiana complex (Taxaceae) based on herbarium material. Botanical Journal of the Linnean Society, 2007, 155(3): 307-335. DOI:10.1111/j.1095-8339.2007.00697.x |
| [38] |
Shah A, Li D Z, Gao L M, Li H T, Möller M. Genetic diversity within and among populations of the endangered species Taxus fuana (Taxaceae) from Pakistan and implications for its conservation. Biochemical Systematics and Ecology, 2008, 36(3): 183-193. DOI:10.1016/j.bse.2007.09.012 |
| [39] |
Poudel R C, Gao L M, Möller M, Baral S R, Uprety Y, Liu J, Li D Z. Yews (Taxus) along the Hindu Kush-Himalayan region: exploring the ethnopharmacological relevance among communities of Mongol and Caucasian origins. Journal of Ethnopharmacology, 2013, 147(1): 190-203. DOI:10.1016/j.jep.2013.02.031 |
| [40] |
徐阿生. 西藏吉隆林区珍稀树种资源评价. 山地研究, 1995(3): 181-186. |
| [41] |
宋垚彬, 徐力, 段俊鹏, 张卫军, 申屠晓露, 李天翔, 臧润国, 董鸣. 西藏极小种群野生植物密叶红豆杉种群的性比及雌雄空间格局. 生物多样性, 2020, 28(3): 269-276. |
| [42] |
Siddiqui M F, Shaukat S S, Ahmed M, Khan N, Khan I A. Vegetation-environment relationship of conifer dominating forests of moist temperate belt of Himalayan and Hindukush regions of Pakistan. Pakistan Journal of Botany, 2013, 45(2): 577-592. |
| [43] |
臧润国, 董鸣, 李俊清, 陈小勇, 曾宋君, 江明喜, 李镇清, 黄继红. 典型极小种群野生植物保护与恢复技术研究. 生态学报, 2016, 36(22): 7130-7135. |
| [44] |
徐耀文, 王亮, 胡蝶. 红豆杉属植物在荆州市园林景观设计中的应用. 绿色科技, 2019(19): 6-9. DOI:10.16663/j.cnki.lskj.2019.19.003 |
| [45] |
张浩, 刘全儒. 密叶红豆杉. 生物学通报, 2017, 52(10): 14-14. DOI:10.3969/j.issn.0006-3193.2017.10.005 |
| [46] |
Shi J F, Cook E R, Lu H Y, Li J B, Wright W E, Li S F. Tree-ring based winter temperature reconstruction for the lower reaches of the Yangtze River in Southeast China. Climate Research, 2010, 41(2): 169-175. |
| [47] |
杨人凡. 西藏吉隆盆地冲锥堆积体的成因研究[D]. 成都: 成都理工大学, 2011.
|
| [48] |
旦增, 李炳章, 尹志坚. 珠穆朗玛峰国家级自然保护区密叶红豆杉所在群落的特征及区系分析. 植物科学学报, 2020, 38(1): 58-67. |
| [49] |
张勇强, 李智超, 宋立国, 厚凌宇, 孙启武. 密叶红豆杉自然分布及群落生态学特征. 生态学报, 2020, 40(6): 1999-2009. |
| [50] |
许振文, 王晓东, 范卫红, 王桂君, 刘惠清. 长白山北坡林线长白落叶松与岳桦对环境适应的对比分析. 地理科学, 2014, 34(6): 748-756. DOI:10.13249/j.cnki.sgs.2014.06.010 |
| [51] |
罗春旺, 于健, 刘琪璟, 徐倩倩, 张国春. 应用照片测量技术提取和自动校正年轮宽度. 生态学杂志, 2016, 35(10): 2845-2851. DOI:10.13292/j.1000-4890.201610.023 |
| [52] |
Grissino-Mayer H D. Evaluating crossdating accuracy: a manual and tutorial for the computer program COFECHA. Tree-Ring Research, 2001, 57(2): 205-221. |
| [53] |
Cook E R, Holmes R L. Users Manual for Program ARSTAN: Laboratory of Tree-Ring Research. Tucson: University of Arizona, 1986.
|
| [54] |
Wang T L, Hamann A, Spittlehouse D L, Murdock T Q. ClimateWNA-high-resolution spatial climate data for western north America. Journal of Applied Meteorology & Climatology, 2012, 51(1): 16-29. |
| [55] |
靳翔. 川西亚高山岷江冷杉树木年轮对气候要素的响应[D]. 北京: 中国林业科学研究院, 2013.
|
| [56] |
Biondi F, Waikul K. DENDROCLIM2002:a C++ program for statistical calibration of climate signals in tree-ring chronologies. Computers & Geosciences, 2004, 30(3): 303-311. |
| [57] |
Deslauriers A, Morin H, Begin Y. Cellular phenology of annual ring formation of Abies balsamea in the Quebec boreal forest (Canada). Canadian Journal of Forest Research, 2003, 33(2): 190-200. DOI:10.1139/x02-178 |
| [58] |
Nagavciuc V, Kern Z, Ionita M, Hartl C, Konter O, Esper J, Popa I. Climate signals in carbon and oxygen isotope ratios of Pinus cembra tree-ring cellulose from the Cǎlimani Mountains, Romania. International Journal of Climatology, 2020, 40(5): 2539-2556. DOI:10.1002/joc.6349 |
| [59] |
Zhu H F, Fang X Q, Shao X M, Yin Z Y. Tree ring-based February-April temperature reconstruction for Changbai Mountain in Northeast China and its implication for east Asian winter monsoon. Climate of the Past, 2009, 5(4): 661-666. DOI:10.5194/cp-5-661-2009 |
| [60] |
Karlsen S R, Solheim I, Beck P S A, Høgda K A, Wielgolaski F E, Tømmervik H. Variability of the start of the growing season in Fennoscandia, 1982-2002. International Journal of Biometeorology, 2007, 51(6): 513-524. DOI:10.1007/s00484-007-0091-x |
| [61] |
Shutova E, Wielgolaski F E, Karlsen S R, Makarova O, Berlina N, Filimonova T, Haraldsson E, Aspholm P E, Flø L, Høgda K A. Growing seasons of Nordic mountain birch in northernmost Europe as indicated by long-term field studies and analyses of satellite images. International Journal of Biometeorology, 2006, 51(2): 155-166. DOI:10.1007/s00484-006-0042-y |
| [62] |
Zhu H F, Zheng Y H, Shao X M, Liu X H, Xu Y, Liang E Y. Millennial temperature reconstruction based on tree-ring widths of Qilian juniper from Wulan, Qinghai Province, China. Chinese Science Bulletin, 2008, 53(24): 3914-3920. |
| [63] |
Duan J P, Zhang Q B, Lv L X, Zhang C. Regional-scale winter-spring temperature variability and chilling damage dynamics over the past two centuries in southeastern China. Climate Dynamics, 2012, 39(3/4): 919-928. |
| [64] |
Fan Z X, Bräuning A, Cao K F, Zhu S D. Growth-climate responses of high-elevation conifers in the central Hengduan Mountains, Southwestern China. Forest Ecology and Management, 2009, 258(3): 306-313. DOI:10.1016/j.foreco.2009.04.017 |
| [65] |
李玲玲, 史江峰, 侯鑫源, 叶金水, 毛海波, 赵欣慰, 鹿化煜. 中国东南高海拔黄山松生长对气候的响应——以浙江省九龙山和安徽省牯牛降为例. 应用生态学报, 2014, 25(7): 1849-1856. DOI:10.13287/j.1001-9332.20140429.008 |
| [66] |
张春雨, 高露双, 赵亚洲, 贾玉珍, 李金鑫, 赵秀海. 东北红豆杉雌雄植株径向生长对邻体竞争和气候因子的响应. 植物生态学报, 2009, 33(6): 1177-1183. |
| [67] |
Ward J K, Dawson T E, Ehleringer J R. Responses of Acer negundo genders to interannual differences in water availability determined from carbon isotope ratios of tree ring cellulose. Tree Physiology, 2002, 22(5): 339-346. DOI:10.1093/treephys/22.5.339 |