 2022, Vol. 42
2022, Vol. 42文章信息
- 黄林娟, 于燕妹, 安小菲, 余林兰, 薛跃规
- HUANG Linjuan, YU Yanmei, AN Xiaofei, YU Linlan, XUE Yuegui
- 天坑森林植物群落叶功能性状、物种多样性和功能多样性特征
- Leaf functional traits, species diversity and functional diversity of plant community in Tiankeng forests
- 生态学报. 2022, 42(24): 10264-10275
- Acta Ecologica Sinica. 2022, 42(24): 10264-10275
- http://dx.doi.org/10.5846/stxb202110273025
-
文章历史
- 收稿日期: 2021-10-27
- 网络出版日期: 2022-07-29





2. 广西师范大学 广西漓江流域景观资源保育与可持续利用重点实验室, 桂林 541006;
3. 广西师范大学 生命科学学院, 桂林 541006
2. Guangxi Key Laboratory of Landscape Resources Conservation and Sustainable Utilization in Lijiang River Basin Guilin, Guangxi Normal University, Guilin 541006, China;
3. College of Life Sciences, Guangxi Normal University, Guilin 541006, China
叶片是植物光合作用和制造有机养分的场所, 也是对环境变化最为敏感的器官[1—2]。叶功能性状作为植物的重要特性之一, 能更为精确的反映植物对特定环境生理生态功能的适应和调控机制, 表征养分循环、物种生产力水平和生态系统服务功能等, 是维持生态系统功能的基本要素[3—4]。叶片的形态、结构和功能性状与生态系统的结构和功能密切相关, 叶的功能性状对于认识物种进化、植被分布、生物多样性保护有重要意义[5]。物种多样性和功能多样性是研究生态系统功能的重要内容[6]。物种多样性能够反映森林植物群落组成的丰富程度[7], 物种多样性的增加会提高群落的稳定性和生态系统功能[8]。功能多样性是理解生态系统过程和功能及其对环境差异或干扰响应的关键[9], 反映了植物功能性状空间变化的范围, 提供群落性状结构、功能及对资源的利用效率[10]。物种多样性和功能多样性是预测生态系统功能的关键因子, 而叶功能性状在生态系统功能与权衡中起着重要作用[11]。因此, 了解叶功能性状是如何影响生态系统功能, 应阐明叶功能性状、物种多样性和功能多样性之间的关系。由于群落组成差异[12]、环境条件差异[13]等, 目前关于叶功能性状、物种多样性、功能多样性之间的联系尚未达成一致[9—13]。所以, 结合不同生境条件下叶功能性状、物种多样性、功能多样性的研究, 既可以探究植物为适应环境所形成的生存策略, 反映植物对气候和生境的(趋同/趋异)适应, 也可以增加对植物叶功能性状与生物多样性的理解。
天坑森林, 为喀斯特负地形生境下形成的亚热带森林生态系统, 其地质结构[14]、物种分布[15]、群落演替[16]等方面均与一般的喀斯特森林存在差异。天坑森林是喀斯特地区天坑负地形生态系统的主体, 是喀斯特石山地区植被类型最为完整而原始的森林之一[17], 在中国西南地区生物多样性的保护和保育中发挥着重要作用[18]。近年来, 随着气候的变暖、人为干扰等因素的影响, 该区域的森林环境也遭受到一定的破坏, 因此, 亟待加强对该区域生态系统和生物多样性的保护。目前的相关研究主要针对单一的天坑森林内部生物(动物、植物、微生物)群落结构和物种多样性特征[15—21]、天坑地质特征及景观价值[22—23]、垂直梯度上物种分布的差异与环境因子的关系[24—25]、优势种生态位特征[26]、种间联结性[27]和功能性状[28]等的研究, 而对于天坑内外森林叶功能性状、物种多样性和功能多样性的研究较少, 忽视了天坑外部喀斯特森林对其物种组成的影响, 进而引起天坑内部森林生物多样性锐减问题[29]。基于这一背景下, 本文选取大石围天坑群天坑内外森林为研究对象, 沿天坑内部-边缘-外部(天坑森林内部-天坑森林边缘-地表喀斯特森林)3种生境开展了植物群落学调查及叶功能性状测定, 试图探究以下2个科学问题:(1)植物的生存策略是否随天坑内外生境差异而变化;(2)天坑内外森林叶功能性状、物种多样性和功能多样性的内在联系。旨在为天坑森林物种多样性维持与保护, 以及生态脆弱地区植被的恢复与优化提供参考。
1 材料与方法 1.1 地理位置研究区位于广西百色市乐业县大石围天坑群(106°21′49″—106°41′37″E, 24°51′50″—24°56′06″N), 生境较为独特。北隔红水河, 西隔南盘江, 处贵州高原向广西盆地过渡的斜坡地带, 地势西南高, 并向东西北三面降低;气候为中亚热带山地气候, 干湿季节分明, 雨热同期, 年平均气温16.4℃, 最热月(7月)平均气温23.4℃, 最冷月(1月)平均气温7.5℃。研究区常见耐阴、喜阳物种, 如香木莲(Manglietia aromatica)、山胡椒(Lindera glauca)、建润楠(Machilus oreophila)、南酸枣(Choerospondias axillaris)、掌叶木(Handeliodendron bodinieri)和化香树(Platycarya strobilacea)等, 植被类型多样, 主要为常绿落叶阔叶混交林[15—20]。根据天坑独特负地形, 将天坑分为天坑内部、天坑边缘和天坑外部3种生境。天坑内部由天坑森林内部组成, 位于天坑负地形内部下坡坡位, 土壤堆积, 土壤成片分布, 岩石裸露较高, 岩石裂隙发育, 水分充足, 光照适中, 环境封闭。天坑边缘由天坑森林边缘组成, 位于天坑负地形顶部, 坡体的上部, 土壤浅薄且多分布于岩石裂缝处, 水分条件较差, 光照充足, 环境开放[21—25]。天坑外部由地表喀斯特森林组成, 位于天坑周边喀斯特石山下坡位, 与天坑森林边缘相连, 土壤浅薄呈不连续分布, 土壤养分易于流失, 水分充足, 光照适中, 环境开放, 受到人类干扰较大。
1.2 样地设置2019年6月—10月, 在充分踏查了大石围天坑群及其周边喀斯特森林的基础上, 在距大石围天坑6 km范围内选取5个分布较为集中的天坑(由近到远依次为罗家天坑、苏家天坑、神木天坑、流星天坑和大曹天坑)和2个天坑外部的喀斯特森林为研究区, 并依据天坑内部、边缘和外部3种生境各设置10个20 m×20 m的典型的植物群落样地(表 1)。大曹天坑、苏家天坑和罗家天坑均属中型天坑, 口部面积介于1.02×104—2.37×104 m之间, 天坑形态为井筒型;神木天坑和流星天坑属大型天坑, 神木天坑口部面积为7.09×104 m, 天坑形态为井筒型, 流星天坑的口部面积为14.88×104 m, 天坑形态为漏斗型。天坑森林内部生境样地包括:大曹天坑4个、苏家天坑2个和流星天坑4个;天坑森林边缘样地包括:苏家天坑2个、罗家天坑4个和神木天坑4个;天坑外部样地包括:罗家天坑附近4个、苏家天坑附近6个。避开放羊、砍柴、种植等人为严重干扰区, 选择受到干扰较轻或中度的地段, 设置样地进行植物群落调查, 并在此基础上, 将每个样地划分为4个10 m×10 m的小样方进行灌木调查。对样方内所有胸径≥2 cm的木本植物进行全面调查, 对乔木层植株进行每木检尺, 记录并测量种名、树高、胸径、冠幅、基径等, 灌木层植株记录种名、树高、盖度等。同时记录每个样方的海拔、经纬度、坡度、坡向等地形因子。所调查的样地共有木本植物120种95属86科, 其中, 天坑森林内部有58种42属35科, 天坑森林边缘有81种52属41科, 周边喀斯特森林有64种43属38科。各生境的优势物种及其重要值见附录1。
| 生境 Habitats |
经纬度 Latitude and longitude |
位置 Location |
海拔 Altitude/m |
坡度 Slope/(°) |
干扰程度 Interference level |
林分密度 Stand density/ (株/hm2) |
| 天坑内部 The inside of Tiankeng |
106°27′22.11″—106°30′50.89″E, 24°47′20.17″—24°48′31.55″N |
天坑负地形内部 | 1072—1274 | 31—56 | 轻度 | 3088 |
| 天坑边缘 The fringe of Tiankeng |
106°27′07.87″—106°27′59.97″E, 24°48′17.71″—24°48′42.92″ |
天坑负地形顶部 | 1276—1368 | 22—46 | 轻度 | 2458 |
| 天坑外部 The outside of Tiankeng |
106°27′05.32″—106°27′23.18″E, 24°48′05.43″—24°48′32.23″N |
喀斯特石山下坡位 | 1321—1390 | 25—46 | 中度 | 5178 |
本研究选取了8个常用的植物叶功能性状作为主要指标, 分别为:叶组织密度(LTD)、比叶面积(SLA)、叶干物质含量(LDMC)、叶面积(LA)、叶碳含量(LCC)、叶氮含量(LNC)、叶磷含量(LPC)和叶钾含量(LKC)。于2020年7月—8月, 在每个样方内采集所有物种的成熟叶片带回实验室进行叶功能性状的测量。叶面积使用CanoScan LiDE 400扫描仪扫描, 然后用Image J软件计算得出[30];叶厚度用游标卡尺(精确度为0.01 mm)在叶片中间非叶脉处进行叶厚度测量;叶鲜重的测量方法:将新鲜叶片用水湿润后, 置于5℃保鲜箱中放置12 h后用1/1000的电子天平称量[31]。叶干重的测量为:将叶片放入烘箱, 然后在60—70℃下烘干48—72 h, 使用1/1000的电子天平称其叶干重[31]。比叶面积=(叶面积/叶干重)×100%, 叶干物质含量=(叶干重/叶鲜重)×100%, 叶组织密度=[叶干重/(叶面积×叶厚度)]×100%。叶碳含量、叶氮含量的测量使用全自动元素分析仪进行测量[32];叶磷含量采用浓硫酸消煮-钼锑抗比色-紫外分光光度计测量[33];叶钾含量采用浓硫酸消煮-火焰原子吸收光谱法测量[34]。
1.4 多样性指数的计算方法本研究选取了4种常用的物种多样性指数, Margalef指数(M)、Shannon-Wiener指数(W)、Simpson指数(S)和Pielou′s均匀度指数(E), 计算公式如下[35—36]:
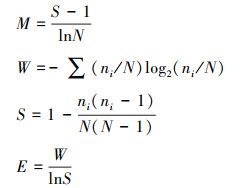
|
式中, S为样方内的总物种数, N为样方中观察到的个体数, ni为第i个物种的个体数。
本研究选取了4种常用的功能多样性指数, 功能丰富度(FRic)、功能均匀度(FEve)、功能离散度(FDis)和Rao′s二次熵(RaoQ), 基于8个叶功能性状为基础的多度加权进行功能多样性的计算。
功能丰富度(FRic)指物种在群落中占据的功能空间大小, 其值越高表示潜在的有效资源被利用的也越多, 计算公式如下:
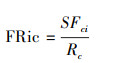
|
式中, SFci为群落i内物种占据的生态位空间, Rc为性状c的绝对范围。
功能均匀度(FEve)指群落内物种功能特征在生态位空间分布的均匀度, 体现物种对占有的有效资源的利用效率, 计算公式如下[37—38]:
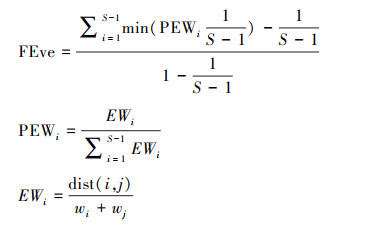
|
式中, EWi为均匀度权重, dist(i, j)为物种i和物种j的欧式距离, Wi为物种i的相对多度, i为分支长, PEWi为分支长的权重。
功能离散度(FDis)指群落性状的多度在性状空间的最大离散度, 其值越大表示物种间的竞争越强, 种间的生态位互补能力越强, 计算公式如下[39]:

|
式中, aj为物种j的多度, zj为物种j到加权质心的距离。
Rao′s二次熵(RaoQ)指群落中性状值的一致性, 其值越大表示物种间的竞争越弱, 种间的生态位重叠越弱, 计算公式如下[39]:
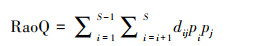
|
式中, S为群落物种丰富度, dij为物种i和物种j的性状值概率密度函数的重叠, pi和pj分别为物种i和物种j的个体数占群落内总物种个体数的比例。
1.5 数据处理群落水平的叶功能性状值通过物种水平的功能性状值按物种多度进行加权平均, 得到的各性状在群落水平的平均功能性状值[40], 多样性指数均采用总体多样性水平来表征整体状况。数据的初步处理及物种多样性的计算均在Excel 2020中进行, 功能性状加权平均值及功能多样性由R 4.0.0中的“FD”包[39]完成。在SPSS 19.0中, 采用单因素分析结合最小显著性差异法(LSD)分析植物群落水平叶功能性状、物种多样性、功能多样性在3种生境中的差异, 采用Pearson相关性分析结合多元逐步回归分析揭示群落水平叶功能性状、物种多样性、功能多样性之间的内在联系。
2 结果与分析 2.1 植物群落叶功能性状总体变化特征从表 2可知, LTD在天坑外生境最高(0.32 g/cm3), 与天坑边缘差异显著(P < 0.05), 但与天坑内部差异不显著(P>0.05);SLA(198.75 cm2/g)在天坑森林内部最高, 与天坑边缘、天坑外部差异显著;LDMC在天坑外部最高(0.41 g/g), 与天坑内部差异显著, 但与天坑边缘差异不显著;LCC在天坑边缘最高(460.34 g/kg), 与天坑内部、天坑外部差异显著, 表明沿天坑生境由内至外, 物种的防御策略增强。LA在天坑内部最高(42.70 cm2), 与天坑边缘、外部差异显著, 但在边缘与外部差异不显著;LPC(1.70 g/kg)在天坑森林内部最高, 与天坑边缘、天坑外部差异显著;LKC在天坑内部最高(10.27 g/kg), 与天坑外部差异显著, 但与天坑边缘差异不显著, 表明沿天坑生境由内至外变化, 物种的生长投入策略减弱。LNC的值为23.8—25.4 g/kg, 在天坑内部-边缘-外部未见明显差异, 表明叶片同化CO2的能力未随生境发生显著变化。综上表明, 沿天坑内部-边缘-外部生境变化, 植物对环境的适应机制和生存策略产生了部分调整, 物种的防御策略增强, 生长投入策略减弱。
| 性状 Traits |
天坑内部 The inside of Tiankeng |
天坑边缘 The fringe of Tiankeng |
天坑外部 The outside of Tiankeng |
| 叶组织密度Leaf tissue density(LTD)/(g/cm3) | 0.31±0.03ab | 0.27±0.07b | 0.32±0.02a |
| 叶面积Leaf area(LA)/(cm2) | 42.70±14.77a | 27.11±7.37b | 28.72±3.53b |
| 叶干物质含量Leaf dry matter content(LDMC)/(g/g) | 0.38±0.03b | 0.40±0.02ab | 0.41±0.02a |
| 比叶面积Specific leaf area(SLA)/(cm2/g) | 198.75±19.30a | 124.48±18.88c | 158.42±9.05b |
| 叶碳含量Leaf total carbon content(LCC)/(g/kg) | 446.56±13.56b | 460.34±11.50a | 443.33±5.68b |
| 叶氮含量Leaf total nitrogen content(LNC)/(g/kg) | 25.40±1.21a | 24.78±4.14a | 23.80±0.86a |
| 叶磷含量Leaf total phosphorus content(LPC)/(g/kg) | 1.70±0.39a | 0.82±0.10c | 1.10±0.06b |
| 叶钾含量Leaf total potassium content(LKC)/(g/kg) | 10.27±3.15a | 8.06±2.60ab | 7.71±0.30b |
| 不同小写字母表示不同生境叶功能性状差异显著(P < 0.05) | |||
从表 3可知, Margalef指数以天坑边缘最高(5.17), 与天坑外部差异不显著, 但与天坑内部差异显著;Shannon-Wiener指数(2.82)和Simpson指数(0.92)以天坑外部最高, 与天坑边缘差异不显著, 但与天坑内部差异显著;功能均匀度以天坑边缘最高(0.63), 与天坑外部差异不显著, 但与天坑内部差异显著, 表明随天坑生境由内至外, 物种分布及其功能性状分布总体上更为均匀、物种数量增多。Pielou′s均匀度指数和功能离散度的值分别为0.86—0.87、1.60—1.83, 但在3种生境中差异不显著, 表明天坑内外生境森林种间对资源竞争较为激烈。功能丰富度以天坑内部最高(1.05), 与天坑外部差异不显著, 但与天坑边缘差异显著;Rao′s二次熵以天坑内部最高(4.52), 与天坑边缘、外部差异显著, 表明随天坑生境由内至外, 物种的功能性状差异逐渐减少。综上表明, 沿天坑内部-边缘-外部生境的变化, 植物功能性状的差异减少, 物种分布及其功能性状分布总体上更为均匀、物种数量增多。
| 多样性指数 Diversity index |
天坑内部 The inside of Tiankeng |
天坑边缘 The fringe of Tiankeng |
天坑外部 The outside of Tiankeng |
| Margalef指数Margalef index (M) | 3.36±0.41b | 5.17±1.27a | 4.76±0.72a |
| Shannon-Wiener指数Shannon-Wiener index(W) | 2.43±0.33b | 2.75±0.25a | 2.82±0.08a |
| Simpson指数Simpson index(S) | 0.83±0.09b | 0.90±0.02a | 0.92±0.01a |
| Pielou′s均匀度指数Pielou′s evenness index (E) | 0.86±0.13a | 0.86±0.03a | 0.87±0.02a |
| 功能丰富度Functional richness index(FRic) | 1.05±1.29a | 0.21±0.34b | 0.60±0.42ab |
| 功能均匀度Functional evenness index(FEve) | 0.58±0.04b | 0.63±0.06a | 0.59±0.06a |
| 功能离散度Functional dispersion index(FDis) | 1.88±0.45a | 1.60±0.31a | 1.63±0.10a |
| Rao′s二次熵Rao′s quadratic entropy index(RaoQ) | 4.52±1.78a | 3.25±1.24b | 3.04±0.53b |
| 不同小写字母表示不同生境多样性指数差异显著(P < 0.05) | |||
由Pearson相关性分析可知(图 1), 物种多样性指数之间、功能多样性指数之间和叶功能性状之间相关性强, 物种多样性和功能多样性指数之间无显著相关性, 各叶功能性状与物种多样性、功能多样性的相关性强。根据多元逐步分析可知(表 4), LTD主要影响M和RaoQ, LA主要影响S、E和FRic, LDMC主要影响FDis, SLA主要影响M和FDis, LCC主要影响FEve, LNC主要影响M、W、E、FRic、FEve和RaoQ, LPC主要影响S、M、FRic和FDis, LKC主要影响E、FRic和RaoQ。综上表明植物群落物种多样性与功能多样性存在不同的制约关系, 叶性状与物种多样性、功能多样性的耦合效应较强, 叶性状对生态学过程的变化较为敏感。

|
| 图 1 植物叶功能性状、物种多样性、功能多样性之间的Pearson相关性分析(P < 0.05*, P < 0.01 **) Fig. 1 The analysis of Pearson correlation between plant leaf functional traits, species diversity and functional diversity (P < 0.05*, P < 0.01 **). M: Margalef指数Margalef index; W: Shannon-Wiener指数Shannon-Wiener index; s: Simpson指数Simpson index; E: Pielou's均匀度指数Pielou' s evenness index; FRic: 功能丰富度Functional richness index; FEve: 功能均匀度Functional evenness index; FDis: 功能离散度Functional dispersion index; RaoQ: Rao's次熵Rao's quadratic entropy index; LTD: 叶组织密度Leaf tsse density; LA: 叶面积Leaf area; LDMC: 叶干物质含量Leaf dry matter content; SLA: 比叶面积Specific leaf area; LCC: 叶碳含量Leaf total carbon content; LNC: 叶氮含量Leaf total nitrogen content; LPC: 叶磷含量Leaf total phosphorus content; LKC: 叶钾含量leaf total potassium content。 |
| 逐步回归方程 Stepwise regression equation |
R2 | F | 显著性 Significant |
| LTD=0.50-0.03M-0.02RaoQ | 0.67 | 31.02 | P < 0.001 |
| LA=46.38-76.41S+7.82FRic+57.13E | 0.69 | 22.29 | P < 0.001 |
| LDMC=0.469-0.04FDis | 0.27 | 11.48 | P < 0.001 |
| SLA=180.99-21.63M+44.24FDis | 0.62 | 13.56 | P < 0.001 |
| LCC=400.61+82.90FEve | 0.11 | 4.42 | P < 0.001 |
| LNC=27.17+1.02RaoQ+3.35M-0.83FRic-18.07FEve-12.29W+26.66E | 0.74 | 4.96 | P < 0.001 |
| LPC=2.36+0.22FRic-1.93S+0.54FDis+0.12M | 0.87 | 47.63 | P < 0.001 |
| LKC=3.61+0.95RaoQ+1.36FRic+9.31E | 0.67 | 20.26 | P < 0.001 |
从全球尺度来看, 气候差异会引起温度、降水、湿度、光照和土壤等环境因子的变化, 造成水分和热量在空间分配上的不均匀, 进而引起植物功能性状的差异[9]。伴随着天坑生境由内至外的变化, SLA和LDMC均发生显著变化。SLA和LDMC与植物的生长速率及对资源的利用效率有关, SLA高LDMC低的植物通常具有较高的养分获取能力, 以资源获取型策略为主, 而SLA低LDMC高的植物通常体内资源的存储能力强, 以资源保守型策略为主[4, 41]。本研究中天坑内部的植物叶片SLA较大而LDMC较小, 天坑边缘及天坑外部的SLA较小而LDMC较大, 表明了天坑内外森林对生境的适应机制和生存策略存在差异。由于天坑内部养分资源丰富、环境湿润[15], 林分内物种主要通过提高生长率、增强养分循环等生长高投入策略来适应脆弱的天坑生境与物种之间的竞争。天坑边缘和天坑外部受到人类活动影响较大[16], 林分内物种则主要通过构建自身机械抗性、降低养分循环等防御高投入策略(如减少LA、增强植物光合作用速率、增加LTD等)来适应胁迫干扰下的环境。LA和LPC是表征植物生长发育状况的综合参数, 其值低有利于防止水分的散失、提高光合作用速率, 指示植物受到水分、温度、养分等胁迫更大[13, 28, 42], 本研究区天坑内部的LA、LPC显著高于天坑边缘、天坑外部, 说明天坑内部受到的环境胁迫最小。叶片N∶P比和C∶P比反映了植物生长所受的养分限制元素, 天坑森林内部、边缘和周边喀斯特森林叶片N∶P比分别为17.50、29.02、21.17, 大于全球16的平均水平[43], C∶P比分别为276.16、552.32、366.67, 大于全球232的平均水平[43], 表明研究区森林均易受到P限制。LNC与叶片同化CO2的能力有关, 是影响植物光合物质代谢和生长的关键因子[44], 本研究中随着天坑由内至外的变化, CO2的同化能力略有减弱, 是因为随着天坑生境由内至外的变化土壤养分的汇集效应逐渐减弱, 可获得的N逐渐减少[27], 但并未达到显著水平。综上可知, 该区域植物对环境的适应机制和生存策略产生了部分调整, 由天坑内部至外部, 物种的防御策略增强, 生长投入策略减弱, 但是单一的指标仅仅只能反映部分特性, 未来需注重综合性状指标的权衡。
中度干扰假说表明, 局部物种多样性在中等强度干扰下最高[45], 本研究结果表明, 沿天坑内部-边缘-外部生境变化, Margalef指数、Shannon-Wiener指数和Simpson指数在人为干扰下的天坑边缘和天坑外部森林显著大于未受人为干扰的天坑内部, 符合中度干扰假说。因为天坑内部森林位于负地形内部, 四周险峻的崖壁基本隔绝了人为干扰, 天坑边缘位于天坑坑口位置, 与外部喀斯特森林相连, 存在少量的人类活动, 而外部地表森林经常伴随着人类耕种、放羊等中度干扰[14—16], 从而导致物种数量的增多。功能多样性是环境筛选和种间相互作用的结果, 反映了植物对环境的适应[37]。本研究结果显示, 天坑森林内部的功能丰富度和Rao′s二次熵显著高于其边缘及周边喀斯特森林, 这是因为天坑森林内部环境多样, 具有多个微生境, 而这大大增加了植物功能性状的变化范围, 促进物种对有效资源的利用[46]。边缘和周边喀斯特森林均为胁迫环境生境, 土壤贫瘠、保水能力差、土壤易于流失, 植物的更新慢, 使得功能均匀度要显著高于天坑森林内部。均匀度指数在天坑内外生境的变化不显著, 表明天坑内外群落存在不同的功能群落, 因为不同的功能群落在天坑森林上的圈层分布增强了群落的稳定[27]。功能离散度是表征植物在性状空间的分布状况[39], 本研究中随着天坑内部-边缘-外部森林的变化呈现先减少后增加的趋势, 但并未达到显著水平, 这是由于天坑森林及其周边喀斯特森林在西南喀斯特地区具有相似的生态系统服务功能, 对喀斯特石山地区植被的恢复和保护具有重要作用[14]。综合可得, 随天坑内部-边缘-外部生境变化, 植物功能性状的差异减少, 物种分布及其功能性状分布总体上更为均匀、物种数量增多。
3.2 植物叶功能性状、物种多样性和功能多样性的内在联系植物叶功能性状之间的关系代表了植物叶片对其生境差异的适应和调整[3], 群落的物种多样性之间的关系代表了生态系统中生物种类的丰富程度[7]、物种分布格局[8], 而功能多样性揭示了植物群落对环境变化的响应和预测生态系统功能的变化, 有利于跨环境梯度解释群落的构建机制[46]。本研究中, 植物物种多样性指数之间、功能多样性指数之间和叶功能性状之间均存在较强的相关性, 这是因为植物群落是生物与环境相互适应的结果[9], 不同的物种之间、群落之间、叶功能性状之间存在不同的制约关系, 而这种制约关系对于物种的选择、功能性状的筛选、群落的组成及调控起着重要作用, 决定了群落的发展与演替速度[16, 47]。吴昊等[46]和贺文君等[42]的研究也表明, 群落内物种多样性指数之间、功能多样性指数之间和叶功能性状之间存在不同耦合关系, 并决定着群落物种的组成及功能的冗余, 与群落构建过程和生态系统功能维持机制有关。
物种多样性与功能多样性之间的关系有正相关[46]、负相关[6]和不相关[48]等, 本研究中物种多样性与功能多样性之间的关系呈不显著相关, 总体上功能多样性随物种多样性的增大而减小, 这与臧岳铭等[49]的结果一致, 其原因在于天坑是由多变的生境资源镶嵌、重叠组合而成的生态系统, 生境异质性强[14, 27], 各种抑制和促进因子综合作用, 共同影响群落结构和功能特性。天坑内部共存物种之间生态位分化强[27], 多样化的生境增大了植物功能性状的分布范围[46], 使得天坑森林内部保持较高的功能丰富度, 而天坑边缘和外部物种数量增多, 胁迫环境下物种的生态位重叠增强, 进而减少了植物功能性状的差异, 引起功能丰富度的降低。物种多样性和功能多样性的关系十分复杂, 因此, 在探讨物种多样性和功能多样性的关系时, 还应该结合地形、土壤、干扰等[9, 39]多个因子进行综合分析。
本研究中叶功能性状与物种多样性、功能多样性之间的相关性强, LDMC、LCC、LTD、LA、SLA、LNC、LPC和LKC均显著影响物种多样性指数、功能多样性指数, 且贡献率均最高可达87%, 这反映了该研究区木本植物叶功能性状与物种多样性、功能多样性存在较强的耦合效应。可能原因为功能性状是指示生态系统功能变化的敏感属性, 能反映出整个群落或生态系统之间的生态关系[40], 土壤性质和土壤质量可以直接影响植物的叶功能性状[47], 而物种多样性和功能多样性是研究生态系统功能的基础[48—49]。该研究区地形地貌复杂、多变, 生境异质性强, 加上地表碳酸岩层的坍塌下降、地下河流水的不断侵蚀, 使得地形特征、岩石特性影响土壤分布状况, 进而影响物种的分布及其功能特性[20, 27—28]。未来需要深入结合物种特性、地形地貌状况和土壤养分对三者之间的影响机制, 探究不同生态系统功能的权衡, 并通过适度的干扰提高生态系统的自我调控能力和平衡能力。
4 结论(1) 沿天坑内部-边缘-外部生境变化, 植物对环境的适应机制和生存策略产生了部分调整, 物种的防御策略增强, 生长投入策略减弱。
(2) 沿天坑内部-边缘-外部生境变化, 植物功能性状的差异减少, 物种分布及其功能性状分布总体上更为均匀、物种数量增多。
(3) 物种多样性指数之间、功能多样性指数之间和叶功能性状之间相关性强, 叶功能性状与物种多样性、功能多样性的相关性强, 但物种多样性和功能多样性之间相关性较弱。
| 生境 Habitats |
优势种 Dominant species |
多度 Abundance |
频度 Frequency |
相对多度 Relative density/% |
相对频度 Relative frequency/% |
相对优势度/ 相对盖度 Relative dominance/ Relative coverage/% |
重要值 Important value/% |
| 天坑内部 | 山胡椒 | 280 | 22 | 22.17 | 10.44 | 2.86 | 11.82 |
| The inside of | 南酸枣 | 56 | 15 | 4.43 | 7.04 | 16.08 | 9.18 |
| Tiankeng | 掌叶木 | 51 | 12 | 4.04 | 5.83 | 14.17 | 8.01 |
| 贵州鹅掌柴 | 89 | 14 | 7.05 | 6.80 | 2.14 | 5.33 | |
| 川钓樟 | 52 | 9 | 4.12 | 4.37 | 3.51 | 4.00 | |
| 中华野独活 | 66 | 9 | 5.23 | 4.37 | 1.73 | 3.78 | |
| 朴树 | 45 | 7 | 3.56 | 3.16 | 3.54 | 3.42 | |
| 近轮叶木姜子 | 58 | 9 | 4.59 | 4.13 | 1.32 | 3.35 | |
| 白楠 | 43 | 8 | 3.40 | 3.88 | 1.86 | 3.05 | |
| 紫弹树 | 37 | 9 | 2.93 | 4.13 | 1.68 | 2.91 | |
| 粗糠柴 | 39 | 9 | 3.09 | 4.13 | 1.46 | 2.89 | |
| 香木莲 | 8 | 3 | 0.63 | 1.21 | 6.08 | 2.64 | |
| 细叶黄皮 | 30 | 6 | 2.38 | 2.91 | 1.74 | 2.34 | |
| 粗叶榕 | 22 | 5 | 1.74 | 2.18 | 2.86 | 2.26 | |
| 华润楠 | 21 | 6 | 1.66 | 2.67 | 2.32 | 2.22 | |
| 裂果卫矛 | 21 | 6 | 1.66 | 2.67 | 2.13 | 2.15 | |
| 滇鼠刺 | 25 | 4 | 1.98 | 1.94 | 2.50 | 2.14 | |
| 豺皮樟 | 19 | 4 | 1.50 | 1.70 | 1.46 | 1.55 | |
| 狗骨柴 | 1 | 1 | 0.08 | 0.49 | 2.80 | 1.12 | |
| 天坑森林边缘 | 南酸枣 | 48 | 10 | 4.68 | 4.83 | 17.35 | 8.95 |
| The fringe of | 川钓樟 | 80 | 27 | 7.80 | 13.04 | 2.31 | 7.72 |
| Tiankeng | 豺皮樟 | 138 | 9 | 13.45 | 4.35 | 1.79 | 6.53 |
| 掌叶木 | 46 | 12 | 4.48 | 5.56 | 5.73 | 5.26 | |
| 长梗冬青 | 33 | 7 | 3.22 | 3.14 | 7.39 | 4.58 | |
| 滇鼠刺 | 44 | 9 | 4.29 | 4.11 | 4.47 | 4.29 | |
| 狗牙花 | 9 | 4 | 0.88 | 1.93 | 8.75 | 3.85 | |
| 猴樟 | 22 | 5 | 2.14 | 2.42 | 4.09 | 2.88 | |
| 小果朴 | 28 | 7 | 2.73 | 3.14 | 2.69 | 2.85 | |
| 朴树 | 13 | 4 | 1.27 | 1.69 | 5.10 | 2.69 | |
| 粉叶润楠 | 43 | 4 | 4.19 | 1.93 | 0.11 | 2.08 | |
| 化香树 | 20 | 4 | 1.95 | 1.93 | 1.97 | 1.95 | |
| 鼠刺 | 16 | 4 | 1.56 | 1.69 | 1.73 | 1.66 | |
| 多毛茜草树 | 1 | 1 | 0.10 | 0.24 | 4.33 | 1.56 | |
| 日本杜英 | 6 | 3 | 0.58 | 1.21 | 2.14 | 1.31 | |
| 针齿铁仔 | 8 | 4 | 0.78 | 1.93 | 0.96 | 1.22 | |
| 青皮木 | 9 | 3 | 0.88 | 1.21 | 1.40 | 1.16 | |
| 细叶黄皮 | 12 | 3 | 1.17 | 1.21 | 0.99 | 1.12 | |
| 天坑外部 | 朴树 | 172 | 27 | 8.17 | 8.92 | 4.03 | 7.04 |
| The outside of | 石斑木 | 228 | 18 | 10.83 | 6.06 | 1.99 | 6.29 |
| Tiankeng | 掌叶木 | 12 | 5 | 0.57 | 1.52 | 15.72 | 5.94 |
| 滇鼠刺 | 72 | 14 | 3.42 | 4.55 | 6.32 | 4.76 | |
| 青榨槭 | 95 | 18 | 4.51 | 6.06 | 3.26 | 4.61 | |
| 长毛籽远志 | 173 | 14 | 8.21 | 4.55 | 0.89 | 4.55 | |
| 猴樟 | 107 | 15 | 5.08 | 4.88 | 1.80 | 3.92 | |
| 短萼海桐 | 60 | 12 | 2.85 | 3.87 | 3.37 | 3.36 | |
| 细叶黄皮 | 99 | 12 | 4.70 | 4.04 | 1.14 | 3.30 | |
| 狗牙花 | 98 | 12 | 4.65 | 3.87 | 1.06 | 3.20 | |
| 球核荚蒾 | 98 | 12 | 4.65 | 3.87 | 0.89 | 3.14 | |
| 长梗冬青 | 18 | 4 | 0.85 | 1.35 | 7.21 | 3.14 | |
| 南酸枣 | 14 | 5 | 0.66 | 1.52 | 7.02 | 3.07 | |
| 化香树 | 30 | 6 | 1.42 | 1.85 | 3.05 | 2.11 | |
| 野八角 | 32 | 6 | 1.52 | 1.85 | 2.65 | 2.01 | |
| 檵木 | 53 | 8 | 2.52 | 2.53 | 0.93 | 1.99 | |
| 鼠刺 | 22 | 6 | 1.04 | 1.85 | 1.67 | 1.52 | |
| 青冈 | 17 | 5 | 0.81 | 1.68 | 1.94 | 1.48 | |
| 川钓樟 | 20 | 5 | 0.95 | 1.68 | 1.34 | 1.33 | |
| 日本杜英 | 14 | 5 | 0.66 | 1.68 | 1.58 | 1.31 | |
| 星毛鸭脚木 | 9 | 4 | 0.43 | 1.18 | 2.18 | 1.26 | |
| 雀梅藤 | 14 | 4 | 0.66 | 1.35 | 1.20 | 1.07 | |
| 赤楠 | 31 | 3 | 1.47 | 0.84 | 0.78 | 1.03 | |
| 重要值的计算:乔木:重要值=100%×(相对多度+相对频度+相对优势度)/3, 灌木:重要值=100%×(相对多度+相对频度+相对盖度)/3 | |||||||
| [1] |
Qin J, Shangguan Z P. Effects of forest types on leaf functional traits and their interrelationships of Pinus massoniana coniferous and broad-leaved mixed forests in the subtropical mountain, Southeastern China. Ecology and Evolution, 2019, 9(12): 6922-6932. DOI:10.1002/ece3.5259 |
| [2] |
季子敬, 全先奎, 王传宽. 兴安落叶松针叶解剖结构变化及其光合能力对气候变化的适应性. 生态学报, 2013, 33(21): 6967-6974. |
| [3] |
刘旻霞, 李俐蓉, 车应弟, 焦骄. 高寒草甸不同演替阶段植物叶片功能性状研究. 植物研究, 2019, 39(5): 760-769. |
| [4] |
岳喜元, 左小安, 庾强, 徐翀, 吕朋, 张晶. 降水量和短期极端干旱对典型草原植物群落及优势种羊草(Leymus chinensis)叶性状的影响. 中国沙漠, 2018, 38(5): 1009-1016. |
| [5] |
李耀琪, 王志恒. 植物叶片形态的生态功能、地理分布与成因. 植物生态学报, 2021, 45(10): 1154-1172. |
| [6] |
张中华, 周华坤, 赵新全, 姚步青, 马真, 董全民, 张振华, 王文颖, 杨元武. 青藏高原高寒草地生物多样性与生态系统功能的关系. 生物多样性, 2018, 26(2): 111-129. |
| [7] |
Petchey O L, Gaston K J. Functional diversity: back to basics and looking forward. Ecology Letters, 2006, 9(6): 741-758. DOI:10.1111/j.1461-0248.2006.00924.x |
| [8] |
Xu W, Ma Z Y, Jing X, He J S. Biodiversity and ecosystem multifunctionality: advances and perspectives. Biodiversity Science, 2016, 24(1): 55-71. DOI:10.17520/biods.2015091 |
| [9] |
韩涛涛, 唐玄, 任海, 王俊, 刘楠, 郭勤峰. 群落/生态系统功能多样性研究方法及展望. 生态学报, 2021, 41(8): 3286-3295. |
| [10] |
Tolonen K E, Leinonen K, Marttila H, Erkinaro J, Heino J. Environmental predictability of taxonomic and functional community composition in high-latitude streams. Freshwater Biology, 2017, 62(1): 1-16. DOI:10.1111/fwb.12832 |
| [11] |
潘权, 郑华, 王志恒, 文志, 杨延征. 植物功能性状对生态系统服务影响研究进展. 植物生态学报, 2021, 45(10): 1140-1153. |
| [12] |
付瑞玉. 弄岗北热带喀斯特植被不同演替阶段的群落构建机制研究[D]. 南宁: 南宁师范大学, 2020.
|
| [13] |
郭雯, 张建, 漆良华, 商泽安, 王锐, 杨畅. 毛竹及其变种叶功能性状与影响因素. 森林与环境学报, 2020, 40(3): 260-268. |
| [14] |
朱学稳. 乐业天坑. 南宁: 广西科学技术出版社, 2018.
|
| [15] |
冯慧喆. 广西大石围天坑群植物区系的起源和演化研究[D]. 桂林: 广西师范大学, 2015.
|
| [16] |
范蓓蓓. 广西大石围天坑群天坑植物群落特征及演替研究[D]. 桂林: 广西师范大学, 2014.
|
| [17] |
林宇. 广西大石围天坑群天坑森林物种多样性研究[D]. 桂林: 广西师范大学, 2005.
|
| [18] |
Pu G Z, Lv Y N, Xu G P, Zeng D J, Huang Y Q. Research progress on Karst Tiankeng ecosystems. The Botanical Review, 2017, 83(1): 5-37. DOI:10.1007/s12229-017-9179-0 |
| [19] |
Su Y Q, Tang Q M, Mo F Y, Xue Y G. Karst tiankengs as refugia for indigenous tree flora amidst a degraded landscape in southwestern China. Scientific Reports, 2017, 7: 4249. DOI:10.1038/s41598-017-04592-x |
| [20] |
Huang L J, Yang H, An X F, Yu Y M, Yu L L, Huang G, Liu X Y, Chen M, Xue Y G. Species abundance distributions patterns between Tiankeng forests and nearby non-Tiankeng forests in southwest China. Diversity, 2022, 14(2): 64-64. DOI:10.3390/d14020064 |
| [21] |
任娟刚, 洪增林, 郭垚鑫, 崔娜, 唐力, 张瑜, 张静. 汉中天坑群喀斯特生境生物多样性特征及规律. 陕西理工大学学报: 自然科学版, 2021, 37(4): 63-69. DOI:10.3969/j.issn.1673-2944.2021.04.009 |
| [22] |
任娟刚, 洪增林, 张静, 唐力, 刘晓峰, 秦臻, 王鹏, 王研. 陕西宁强禅家岩天坑群喀斯特地质遗迹特征及成因. 中国岩溶, 2021, 40(3): 539-547. |
| [23] |
韦跃龙, 李成展, 陈伟海, 吴忠, 罗劬侃, 翟秀敏, 白冰. 重庆奉节天坑地缝景区喀斯特景观特征及其形成演化分析. 地球学报, 2019, 40(5): 747-766. |
| [24] |
朱粟锋, 江聪, 税伟, 郭平平, 张永永, 冯洁, 高宸, 鲍玉珍. 退化天坑倒石堆阴坡植物群落垂直分布特征——以云南沾益深陷塘为例. 应用生态学报, 2020, 31(5): 1496-1504. |
| [25] |
吴金, 李承义, 李小芳, 张朝晖. 喀斯特天坑内孔雀藓科植物垂直分布特征及其与环境因子关系的研究. 植物研究, 2019, 39(1): 45-53. |
| [26] |
郭平平, 税伟, 江聪, 简小枚, 朱粟锋, 张永永, 冯洁, 陈毅萍. 退化天坑倒石坡林下优势物种生态位特征. 应用生态学报, 2019, 30(11): 3635-3645. |
| [27] |
黄林娟, 于燕妹, 安小菲, 余林兰, 薛跃规. 广西大石围天坑群天坑森林主要木本植物种间关联性研究. 广西植物, 2021, 41(5): 695-706. |
| [28] |
冯洁, 江聪, 税伟, 朱粟锋, 郭平平, 孙祥, 张永永, 刘橼锰. 喀斯特退化天坑阴坡阳坡壳斗科植物的功能性状特征. 应用生态学报, 2021, 32(7): 2301-2308. |
| [29] |
蒲高忠, 王柯懿, 莫凌, 曾丹娟, 陈霞霞. 中国喀斯特天坑演化及植被生态系统研究进展. 广西植物, 2021, 41(10): 1632-1643. |
| [30] |
戴志聪, 杜道林, 司春灿, 林英, 郝建良, 孙凤. 用扫描仪及Image J软件精确测量叶片形态数量特征的方法. 广西植物, 2009, 29(3): 342-347. |
| [31] |
Pérez-Harguindeguy N, Díaz S, Garnier E, Lavorel S, Poorter H, Jaureguiberry P, Bret-Harte M S, Cornwell W K, Craine J M, Gurvich D E, Urcelay C, Veneklaas E J, Reich P B, Poorter L, Wright I J, Ray P, Enrico L, Pausas J G, de Vos A C, Buchmann N, Funes G, Quétier F, Hodgson J G, Thompson K, Morgan H D, ter Steege H, Sack L, Blonder B, Poschlod P, Vaieretti M V, Conti G, Staver A C, Aquino S, Cornelissen J H C. New handbook for standardised measurement of plant functional traits worldwide. Australian Journal of Botany, 2013, 61(3): 167. |
| [32] |
王巧环, 任玉芬, 孟龄, 李虹, 傅慧敏, 王华锋. 元素分析仪同时测定土壤中全氮和有机碳. 分析试验室, 2013, 32(10): 41-45. |
| [33] |
鲍士旦. 土壤农化分析 (3版). 北京: 中国农业出版社, 2000.
|
| [34] |
刘生涛. 皇甫川流域不同水土治理方式下土壤钾含量的变化. 北方环境, 2011, 23(3): 58-61. |
| [35] |
Keylock C J. Simpson diversity and the Shannon-Wiener index as special cases of a generalized entropy. Oikos, 2005, 109(1): 203-207. |
| [36] |
刘晓红, 李校, 彭志杰. 生物多样性计算方法的探讨. 河北林果研究, 2008, 23(2): 166-168. |
| [37] |
Mason N W H, Mouillot D, Lee W G, Wilson J B. Functional richness, functional evenness and functional divergence: the primary components of functional diversity. Oikos, 2005, 111(1): 112-118. |
| [38] |
Villéger S, Mason N W H, Mouillot D. New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology, 2008, 89(8): 2290-2301. |
| [39] |
Laliberté E, Legendre P. A distance-based framework for measuring functional diversity from multiple traits. Ecology, 2010, 91(1): 299-305. |
| [40] |
Garnier E, Cortez J, Billès G, Navas M L, Roumet C, Debussche M, Laurent G, Blanchard A, Aubry D, Bellmann A, Neill C, Toussaint J P. Plant functional markers capture ecosystem properties during secondary succession. Ecology, 2004, 85(9): 2630-2637. |
| [41] |
Wright I J, Reich P B, Westoby M, Ackerly D D, Baruch Z, Bongers F, Cavender-Bares J, Chapin T, Cornelissen J H C, Diemer M, Flexas J, Garnier E, Groom P K, Gulias J, Hikosaka K, Lamont B B, Lee T L, Lee W, Lusk C, Midgley J J, Navas M L, Niinemets V, Oleksyn J, Osada N, Poorter H, Poot P, Prior L, Pyankov V I, Roumet C, Thomas S C, Tjoelker M G, Veneklaas E J, Villar R. The worldwide leaf economics spectrum. Nature, 2004, 428(6985): 821-827. |
| [42] |
贺文君, 韩广轩, 颜坤, 管博, 王光美, 路峰, 周英锋, 张乐乐, 周莉. 微地形对滨海盐碱地土壤水盐分布和植物生物量的影响. 生态学杂志, 2021, 40(11): 3585-3597. |
| [43] |
Koerselman W, Meuleman A F M. The vegetation N: P ratio: a new tool to detect the nature of nutrient limitation. The Journal of Applied Ecology, 1996, 33(6): 1441. |
| [44] |
Dier M, Sickora J, Erbs M, Weigel H J, Zörb C, Manderscheid R. Decreased wheat grain yield stimulation by free air CO2 enrichment under N deficiency is strongly related to decreased radiation use efficiency enhancement. European Journal of Agronomy, 2018, 101: 38-48. |
| [45] |
Meyer P, Schmidt M, Feldmann E, Willig J, Larkin R. Long-term development of species richness in a central European beech (Fagus sylvatica) forest affected by windthrow-Support for the intermediate disturbance hypothesis?. Ecology and Evolution, 2021, 11(18): 12801-12815. |
| [46] |
吴昊, 肖楠楠, 林婷婷. 秦岭松栎林功能多样性与物种多样性和环境异质性的耦合关系. 生态环境学报, 2020, 29(6): 1090-1100. |
| [47] |
Wen Z, Zheng H, Smith J R, Ouyang Z Y. Plant functional diversity mediates indirect effects of land-use intensity on soil water conservation in the dry season of tropical areas. Forest Ecology and Management, 2021, 480: 118646. |
| [48] |
Zhang Z H, Hou J H, He N P. Predictability of functional diversity depends on the number of traits. Journal of Resources and Ecology, 2021, 12(3): 332-345. |
| [49] |
臧岳铭, 朱志红, 李英年, 王文娟, 席博. 高寒矮嵩草草甸物种多样性与功能多样性对初级生产力的影响. 生态学杂志, 2009, 28(6): 999-1005. |