 2022, Vol. 42
2022, Vol. 42文章信息
- 郭冬, 张同文, 吐尔逊·哈斯木, 张瑞波, 喻树龙, 刘可祥, 苟晓霞, 刘蕊, 石仁娜·加汗, 王兆鹏, 如先古丽·阿不都热合, 阿依姆古丽·赛麦提
- GUO Dong, ZHANG Tongwen, Tursun·Kasim, ZHANG Ruibo, YU Shulong, LIU Kexiang, GOU Xiaoxia, LIU Rui, Shirenna·Jiahan, WANG Zhaopeng, Ruxianguli·Abudureheman, Ayimuguli·Saimaiti
- 西昆仑山两种针叶树种径向生长及其对气候变化的响应
- Radial growth of two coniferous species in the western Kunlun Mountains and its response to climate change
- 生态学报. 2022, 42(24): 10226-10240
- Acta Ecologica Sinica. 2022, 42(24): 10226-10240
- http://dx.doi.org/10.5846/stxb202110112844
-
文章历史
- 收稿日期: 2021-10-11
- 网络出版日期: 2022-07-29





2. 中国气象局乌鲁木齐沙漠气象研究所, 新疆树轮生态重点实验室, 中国气象局树木年轮理化研究重点实验室, 乌鲁木齐 830002;
3. 新疆师范大学地理科学与旅游学院, 乌鲁木齐 830054
2. Key Laboratory of Tree-ring Physical and Chemical Research of China Meteorological Administration, Xinjiang Laboratory of Tree Ring Ecology, Institute of Desert Meteorology, China Meteorological Administration, Urumqi 830002, China;
3. School of geographical science and tourism, Xinjiang Normal University, Urumqi 830054, China
森林是陆地生态系统中重要的组成部分, 其生物量和初级生产能力占整个生态系统的近50%, 是生态圈重要的碳汇[1—4]。树木是森林的组成单位, 其生长除受自身生理特性、立地环境条件等因素的影响外, 还受到域内气候条件的密切影响。这使树木年轮能够真实地记录下每年有利或不利于树木生长的气候因素[5—6]。自工业革命以来, 全球气候持续变暖。在IPCC第五次评估报告指出, 1880—2012年期间, 全球地表气温较工业化前上升了0.65—1.06℃[7]。20世纪80年代以来, 北半球中高纬度干旱半干旱区对气候变化的响应尤其显著[8—9]。而且内陆干旱区的树木生长对气候要素响应敏感[10—11]。同时树木年轮还具有年分辨率高以及可靠性强等特点[12—15]。因此, 科研工作者利用树木年轮资料在这一区域开展了树轮宽度变化特征、树木径向生长气候响应、历史气候水文重建等系列研究工作[16—25]。
随着全球气候变暖, 树木的径向生长对气候因子的响应关系也呈现不稳定[26—27]。1995年, D′Arrigo等[28]提出了“分异现象”的概念, 指树轮宽度或树轮密度对气候变暖的敏感性不稳定的现象。国内外学者在树木径向生长与气候要素之间响应方面取得一系列研究进展[29—36]。目前, 围绕西昆仑山及附近区域开展了一系列树木径向生长与气候要素的响应关系等相关研究工作。尚华明等[20]通过对帕米尔高原东北部昆仑圆柏(Juniperus jarkendensis)气候响应研究发现年轮宽度年表与上年10月至当年7月降水量响应最好。叶尔羌河流域研究表明雪岭云杉(Picea schrenkiana)受上年6月到当年5月的降水量显著影响[21]。杨美琳等[22]在塔吉克斯坦北部发现土耳其斯坦圆柏(Juniperus turkestanica)树木径向生长与当年2—3月平均最低气温显著相关。西帕米尔高原地区的柏树的气候-径向生长响应分析表明, 7月气温和夏季降水量是影响其生长的主要气候因子[23], 并与当年2—4月月平均标准化降水蒸散指数SPEI和年6—7月自适应帕默尔干旱指数sc-PDSI较好的响应[24—25]。苟晓霞等[37]对塔吉克斯坦苦盏附近山区泽拉夫尚圆柏(Juniperus seravschanica)研究表明, 不同海拔和坡向所导致的圆柏径向生长对气候因子响应敏感性加强和减弱变化是不同的。该区域对同一生境不同针叶树种树木径向生长-气候要素响应关系的研究还有待深入。
本文利用研究区主要针叶树种雪岭云杉和昆仑圆柏树轮宽度资料建立标准化宽度年表, 分析其年表特征, 揭示混交林中树木径向生长的差异性及其对气候要素的响应机制, 并进一步探讨了两种树种对气候因子响应敏感性在随气候变暖背景下的变化情况。本研究对于西昆仑山不同针叶树种径向生长变化特征的认识, 以及今后在该区域开展树芯样本采集和基于树轮资料的历史气候重建研究提供参考。
1 材料与方法 1.1 研究区域及样本采集本研究的树芯样本采样点为阿克陶县的奥依塔克森林公园, 属于帕米尔高原东北部, 位于塔里木盆地西南缘, 昆仑山西侧(图 1)。研究区地处欧亚大陆腹地, 域内蒸发量大, 大陆性和干燥性较强, 降水量较少[38—39]。年内最高气温出现在7月, 最低气温则在1月。当地植被为山地高原旱生荒漠植被类型[40]。受地形影响, 森林带分布在海拔2900—3500 m。森林上限以上为亚高山草原带和冰川积雪, 森林下限以下为山地半荒漠带[29—31]。研究团队于2018年5月在山脉北坡海拔3000 m附近(38°57.6′E, 75°9.6′N)开展雪岭云杉树芯样本采集工作, 选择了26棵雪岭云杉, 共计采集53个树芯样本;2020年10月研究团队在相同区域再次进行采集, 选择了5棵雪岭云杉和23棵昆仑圆柏, 分别采集13和46个树芯样本。将采集的树芯样本封装和保存在纸管内, 标注明确采样相关信息后, 运送回实验室进行处理。

|
| 图 1 研究区域树木年轮采样点及气象站位置 Fig. 1 Location of the tree-ring sampling sites and the meteorological station in the study area |
按照树轮年轮学原理[41], 对采集的雪岭云杉和昆仑圆柏树芯样本进行干燥、固定、打磨、并在显微镜下目测定年, 使用宽度测量仪LINTAB测量树轮宽度(精确度为0.001 mm)。通过使用COFECHA程序进行交叉定年的质量控制, 分析树芯样本发生缺轮的年份与数量。雪岭云杉采样点树芯样本存在52个缺轮, 缺轮率为0.620%;昆仑圆柏采样点树芯样本存在12个缺轮, 缺轮率为0.317%。使用国际树木年轮数据库ARSTAN程序中的负指数函数, 去除树木生长趋势, 建立树木年轮标准化年表。
1.3 气象数据本研究选择了采样点附近喀什气象站(39°28′N, 75°59′E, 1290.7 m)的气象资料表征研究区域气候背景。该数据来源于中国气象科学数据共享服务网(http://cdc.cma.gov.cn)。使用1951—2020年逐月平均气温和降水数据;以及涵盖采样点位置Climatic Research Unit (CRU)格点数据(http://climexp.knmi.nl)中1982—2016年1 m深处土壤水分贮存量数据, 其数据分辨率为0.5°×0.5°(图 2)。此外, 还选取了喀什气象站1961—2020年气象干旱综合指数(MCI), 数据来源于新疆气候中心。以及田间持水量(AWC)数据使用了国际应用系统分析研究所(IIA-SA)、联合国粮农组织(FAO)共同建立世界土壤数据库(HWSD)土壤数据集(v1.2)(https://daac.ornl.gov/cgi-bin/dsviewer.pl?ds_id=1247), 数据空间分辨率为0.05°, 划分年份为2000年, 这些数据用于后续分析的气象干旱指数计算使用。从图 2中可以看出, 研究区年内逐月降水量和平均气温峰值出现时段不同。降水量为双峰型分布, 最大降水量峰值在5月, 降水量为12.26 mm;第二降水量峰值在8月, 降水量为9.32 mm;而12月降水量最少, 仅为1.94 mm。土壤水分贮存量最大在8月, 处于主要降水期;最小土壤水分贮存量出现在2月。1951—2020年期间, 年平均气温(y=-40.581+0.026505x, R2=0.401, F=45.481, P < 0.001)和年降水量(y=-1158.425+0.61995x, R2=0.110, F=8.39, P < 0.1)均呈显著上升趋势且气温的上升趋势更为显著, 而1982—2016年的年土壤水分贮存量则呈不显著下降趋势。

|
| 图 2 研究区气候与环境要素变化特征 Fig. 2 Characteristics of Climate and Environmental factors in the study area |
本研究使用SPSS 26[42]计算雪岭云杉和昆仑圆柏树木年轮宽度指数与气候因子的Pearson相关系数。使用u检验方法计算相关系数的差异性。|u|>1.96, 表示同一生境不同时段树轮年表与气候要素之间的相关系数有显著差异(P < 0.05), 反之, 则表示同一生境不同时段树轮年表与气候要素之间的相关系数差异不明显。树木年轮宽度年表和基部断面生长增量(BAI)常用来分析树木径向生长量变化[43—44]。本文选择无异常生长的雪岭云杉和昆仑圆柏的树木样芯。使用其树木年轮宽度测量值计算出单个树芯样本的BAI值。计算方法如下:
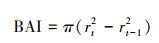
|
式中, rt表示树木年轮样本在t年的生长半径, rt-1表示树木年轮样本在t-1年的生长半径。平均BAI值是同一采样区内相同树种中所有树芯样本在对应年份内的平均值。使用非参数统计方法, 即Mann-Kendall(M-K)突变检验法对研究区域内的气候要素进行分析, 确定可能发生气候突变的年份。1989年世界气象组织提出了气候条件(如气温和降水)30年的平均值为平均气候状态[45]。本文使用了以30年为滑动窗口, 1年为滑动步长, 开展树轮宽度年表和气候要素的滑动相关分析。
2 结果与分析 2.1 气候变化趋势图 3表示1951—2020年喀什气象站气温和降水突变。使用M-K突变检验分析可得, 喀什气象站年平均气温1994年发生突变, 年降水量2008年发生突变。根据图 2可以看出, 气温上升趋势的显著性最好, 且降水量突变发生在1951—2020年的最后时段。因此, 选择年平均气温突变点1994年作为气候突变点, 将气温和降水量分割为两段, 即1968—1993年和1995—2020年时段。另外, 图 2也可以看出, 气温升高对土壤水分的影响超过降水增加的作用, 并使土壤水分贮存量缓慢下降。

|
| 图 3 气温(左)和降水量(右)的突变检验 Fig. 3 Abrupt test of mean annual temperature(left) and annual total precipitation(right) UFk: Mann-Kendall统计值组成的曲线; UBk: Mann-Kendall反序列统计值组成的曲线 |
树木年轮宽度年表特征参数见表 1。雪岭云杉和昆仑圆柏年表的标准方差别为0.241和0.266, 表明该采样点的树木年轮宽度年表中包含的气候变化信息可能更多。平均敏感度(Mean sensitivity)表示年轮时间序列对气候信息的敏感程度, 其中雪岭云杉和昆仑圆柏年表均符合树木年轮研究标准, 昆仑圆柏年表平均敏感度(0.205)较高。一阶自相关系数反映了上一年气候变化对树木年轮生长的持续性影响, 对比发现雪岭云杉年表的一阶自相关系数(0.874)大于昆仑圆柏年表, 说明上年气候特征变化对当年雪岭云杉径向生长影响比昆仑圆柏更大。雪岭云杉年表的样本间相关和树木间相关均大于昆仑圆柏年表, 表明雪岭云杉径向生长的一致性较昆仑圆柏更好。通常, 信噪比越高, 表明气候对树木生长影响越大[46], 通过对比发现雪岭云杉年表信噪比大于昆仑圆柏年表, 说明雪岭云杉比昆仑圆柏包含的气候信息含量更多。两个年表中的样本解释总量均大于0.85, 表明这两种树种的树木年轮宽度年表具有区域代表性。总体来说, 各项年表特征均表明, 雪岭云杉和昆仑圆柏年表均适用于树木年轮气候学分析。
| 树种 Tree species |
雪岭云杉 Picea schrenkiana |
昆仑圆柏 Juniperus jarkendensis |
| 标准方差Standard deviation | 0.241 | 0.266 |
| 平均敏感度Mean sensitivity | 0.175 | 0.205 |
| 一阶自相关First-order auto-correlation | 0.874 | 0.399 |
| 样本间相关Correlation coefficients between series | 0.303 | 0.197 |
| 树木间相关Correlation coefficients between trees | 0.292 | 0.183 |
| 信噪比Signal-to-noise ratio | 24.322 | 10.761 |
| 样本解释总量Expressed population signal | 0.961 | 0.915 |
| 信号强度SSS < 0.85起始年First year of SSS>0.85 | 1857 | 1936 |
| SSS: 子样本信号强度Subsample signal strength | ||
图 4为研究区域雪岭云杉和昆仑圆柏年轮宽度指数变化特征曲线。从图 4中可以看出, 雪岭云杉年表序列较长, 为282年(1739—2020年), 有效年表164年(1857—2020年);昆仑圆柏年表序列126年(1895—2020年), 有效年表85年(1936—2020年)。在气温突变前后, 雪岭云杉年表在1968—1993年时段内呈上升趋势, 1995—2020年时段内呈下降趋势。昆仑圆柏年表的变化趋势与雪岭云杉年表具有一致性, 但昆仑圆柏年表在1995—2020年下降趋势均不显著。使用高低通滤波器[47—48]对雪岭云杉和昆仑圆柏年表信息进行分解, 得到年表中的低频域和高频域。其中, 低频域为通过低通滤波器的低频(长波)信息;高频域为通过高通滤波器的高频(短波)信息。然后, 分别在不同时段的全频域、低频域和高频域做相关分析。在公共区间(1936—2020年)内, 雪岭云杉和昆仑圆柏宽度年表在全频域、低频域和高频域的相关系数分别为0.521(n=85, P < 0.001)、0.540(n=85, P < 0.001)、0.519(n=85, P < 0.001), 相关系数均超过99%的置信水平, 说明西昆仑山北坡森林的雪岭云杉和昆仑圆柏的树木年轮宽度年表存在较好的相关性, 说明雪岭云杉和昆仑圆柏的树木径向生长趋势存在相似性。

|
| 图 4 树木年轮宽度年表(细线)和样本量(粗线) Fig. 4 The tree-ring width chronology (filament line) and the amount of samples (thick line) 黑色虚线代表子样本信息强度SSS < 0.85的起始年份 |
图 5为1951—2020年雪岭云杉和昆仑圆柏的树木断面生长增量(BAI)进行分析。雪岭云杉和昆仑圆柏平均BAI在逐步增加, 且增长趋势非常显著(P<0.001)。雪岭云杉的平均BAI值明显小于昆仑圆柏的平均BAI值。另外, 雪岭云杉BAI和昆仑圆柏BAI在气温突变前(1968—1993年), 呈快速增加趋势(P<0.001);然而, 气温突变后(1995—2020年), 雪岭云杉和昆仑圆柏的BAI呈略微下降趋势, 但下降趋势变化不显著。这表明西昆仑山北坡森林中, 气温突变前后, 存在影响雪岭云杉和昆仑圆柏两种针叶树种的树木径向生长的要素超过了其生长能够承受的阈值范围。

|
| 图 5 基部断面生长增量(BAI)曲线变化 Fig. 5 The change curves of basal area increment(BAI) |
为了揭示同一区域的雪岭云杉和昆仑圆柏径向生长对气候因子的响应关系, 将雪岭云杉年表和昆仑圆柏年表与月平均气温和降水量两个气候因子做相关分析。根据山地气温垂直递减率和树木生理特征[49], 确认雪岭云杉和昆仑圆柏在西昆仑山的生长季为4—10月。并且研究表明, 树木生长对气候后变化的响应具有“滞后效应”[50—52], 因此本文选择上年4月至当年10月的气候因子与树木年轮年表做相关性分析。由图 6可知, 在1951—2020年时段内, 雪岭云杉与昆仑圆柏年表对气候因子的响应具有一定相似性和差异性。对比雪岭云杉与昆仑圆柏年表发现, 都与上年10月和当年4—6月气温显著负相关, 当年3月降水显著正相关。然而, 雪岭云杉年表与上年5—6月、8—9月和当年3月、10月的多个月份气温显著负相关, 昆仑圆柏年表还与当年4月、6月降水也显著正相关。

|
| 图 6 树轮宽度年表与气候因子的相关分析 Fig. 6 The correlation coefficients between the tree-ring width chronology and monthly climatic factors P表示上年, C表示当年;★代表P>0.05显著性, ★★代表P>0.01显著性 |
前文分析可知, 雪岭云杉和昆仑圆柏在1994年突变前后的径向生长趋势情况差异明显(图 4), 表明不同时段树木生长过程中对气候要素的响应程度有所不同。因此, 选取了1968—1993年和1995—2020年两个时段内分别分析气温和降水对两种树种树木生长的影响。如图 6所示, 1968—1993年时段内, 雪岭云杉和昆仑圆柏年表与气温因子的响应中, 雪岭云杉年表与上年5—7月、10月和当年5—7月、10月的气温因子显著负相关;昆仑圆柏年表与当年4月和6—7月的气温因子显著负相关。气温突变之后, 雪岭云杉年表与上年5月、9—10月和当年3—7月的气温因子显著负相关;昆仑圆柏年表与上年4月、9—10月和当年4—6月、10月的气温因子显著负相关。在降水方面(图 6), 1968—1993年时段内, 仅有昆仑圆柏年表与上年9月和当年9月降水因子显著负相关。气温突变后, 雪岭云杉年表与上年4月和当年3—4月、7月的降水温因子显著正相关;昆仑圆柏年表与当年4月的降水因子显著正相关。
雪岭云杉和昆仑圆柏的树木年轮宽度年表与不同时段(1968—1993年和1995—2020年)气候因子相关系数差异的u值检验结果如表 2所示, 发现雪岭云杉和昆仑圆柏年表均与气温因子和降水因子相关系数u值检验差异不明显。
| 月份 Month |
气温Temperature | 降水Precipitation | |||
| 雪岭云杉 | 昆仑圆柏 | 雪岭云杉 | 昆仑圆柏 | ||
| P4 | 0.231 | 0.746 | 1.148 | 1.016 | |
| P5 | 0.253 | 0.414 | 0.096 | 0.410 | |
| P6 | 0.927 | 1.003 | 0.878 | 0.515 | |
| P7 | 1.398 | 0.493 | 0.118 | 0.446 | |
| P8 | 0.229 | 0.908 | 0.026 | 0.111 | |
| P9 | 1.394 | 1.654 | 1.066 | 0.758 | |
| P10 | 0.045 | 1.449 | 1.489 | 0.802 | |
| P11 | 1.323 | 0.285 | 1.032 | 1.411 | |
| P12 | 0.159 | 0.741 | 0.295 | 0.329 | |
| C1 | 1.073 | 1.617 | 0.108 | 0.432 | |
| C2 | 1.827 | 1.705 | 0.557 | 0.426 | |
| C3 | 0.478 | 0.014 | 0.295 | 0.047 | |
| C4 | 1.185 | 0.718 | 1.829 | 1.695 | |
| C5 | 0.957 | 1.475 | 0.077 | 1.236 | |
| C6 | 0.073 | 0.736 | 0.144 | 0.042 | |
| C7 | 0.751 | 0.703 | 1.236 | 0.015 | |
| C8 | 0.128 | 0.967 | 1.310 | 1.611 | |
| C9 | 0.794 | 1.087 | 0.794 | 1.602 | |
| C10 | 0.791 | 0.969 | 0.651 | 1.448 | |
| P表示上年, C表示当年,数字代表月份 | |||||
雪岭云杉和昆仑圆柏年表与不同季节气温和降水的季节相关分析结果如图 7所示。结果显示:在1951—2020年时段内, 雪岭云杉和昆仑圆柏年表均与当年春季和夏季气温因子显著负相关, 雪岭云杉年表还与上年春季、夏季和秋季气温因子显著负相关。气温突变前后, 雪岭云杉和昆仑圆柏年表均对上年春季、当年春季和夏季气温的响应有负响应增强。在降水方面, 雪岭云杉和昆仑圆柏年表对当年春季降水从显著正相关, 雪岭云杉年表还与当年秋季降水呈显著负相关;并且在气温突变之后, 两种针叶树种树轮宽度与当年春季降水的正响应均有显著增强。

|
| 图 7 树木年轮年表与气候因子月份组合分析结果 Fig. 7 The correlation coefficients between the tree-ring chronology and seasonal climatic factors P表示上年, C表示当年;★代表P < 0.05显著性, ★★代表P < 0.05显著性 |
为了探究和揭示在气候变暖的情况下, 1951—2020年时段内雪岭云杉和昆仑圆柏径向生长对气候因子响应敏感程度的变化情况。因此, 对雪岭云杉和昆仑圆柏宽度年表与平均气温和降水之间做30年滑动相关。从图 8可以看出, 随着气候变暖, 树木年轮宽度年表对气候因子响应敏感程度总体在不断增强, 很大部分时段, 与部分气温因子的响应出现了相关系数正负转换的情况。

|
| 图 8 树木年轮宽度年表与气候因子滑动相关分析 Fig. 8 The moving correlation analysis between the tree-ring width chronology and monthly climatic factors ●表示P < 0.01显著性; P、C后面数据代表月份 |
从图 8可以看出, 1951—2020年时段内雪岭云杉和昆仑圆柏年表对气温变化的响应存在异同。雪岭云杉和昆仑圆柏年表与上年9—10月、当年4—6月和10月气温负响应逐渐增强。其中, 雪岭云杉年表还与上年4—6月、当年3月和7月气温具有负响应逐渐增强。图 7显示, 1972—2018年时段内雪岭云杉年表与上年4—6月气温显著负相关(P < 0.01);1976—2020年时段内与上年9—10月气温显著负相关(P < 0.01);1973—2020年时段内与3—7月(生长季)气温显著负相关(P < 0.01);1965—2020年时段内与当年10月气温显著负相关(P < 0.01)。图 8显示, 1980—2020年时段内昆仑圆柏年表与上年9—10月气温显著负相关(P < 0.01);1958(1978)—2020年时段内与当年4—6月(生长季)气温显著负相关(P < 0.01);1985—2020年时段内与当年10月气温显著负相关(P < 0.01)。
就降水而言, 雪岭云杉年表与昆仑圆柏年表对降水因子的响应明显不同。树木年轮宽度年表与降水因子的相关分析表明(图 8), 雪岭云杉年表仅在1953—1982年时段内与上年11月降水显著正相关(P < 0.01);与雪岭云杉不同, 昆仑圆柏年表在1985—2019年时段内与当年4月降水显著正相关(P < 0.01), 1972—2006年时段内与当年9月降水显著负相关(P < 0.01)。
3 讨论 3.1 不同树种树木径向生长特征在全球气候变暖的背景下, 1951—2020年期间研究区域附近喀什气象站在1994之后气温加速上升, 降水量快速增加, 气温和降水量均明显高于1968—1993年期间趋势变化, 说明此区域气温突变后呈明显增温增湿的趋势。在1968—1993年时段雪岭云杉和昆仑圆柏树木年轮宽度年表和BAI值均呈上升趋势;1994年后, 雪岭云杉和昆仑圆柏的树木年轮宽度年表和BAI值呈下降趋势(图 4和图 5)。雪岭云杉年表的有效长度明显高于昆仑圆柏年表(图 4)。在1951—2020年内, 两种针叶树种的年轮宽度年表在全频域、低频域和高频域之间的相关性较好, 说明同一区域不同树种树木径向生长趋势存在相似性。
对比图 4和图 5发现, 1968—1993年和1995—2020年时段内, 两树种年表及BAI变化趋势相近, 都表现1968—1993年内呈上升趋势, 1994年后呈下降趋势(图 4和图 5)。对比发现(图 5)昆仑圆柏BAI值增加速率快于雪岭云杉, 说明昆仑圆柏的树木径向生长速率高于雪岭云杉。在气候突变以后(1994年以后), 雪岭云杉和昆仑圆柏变化趋势均略微下降, 气温对雪岭云杉和昆仑圆柏径向生长的限制作用进一步加强。那么在未来气候持续增暖情况下, 西昆仑山的森林可能会出现衰退的现象。
3.2 雪岭云杉和昆仑圆柏树木径向生长的影响因子考虑到气候变化对树木径向生长存在累计作用, 因此使用了树轮宽度年表与月份和季节的气候要素的相关分析[20, 53]。将同一区域的树木年轮宽度年表与月份和季节气候要素做相关性分析发现(图 6和图 7), 不同树种对同一气候要素的响应存在异同。雪岭云杉树木年轮宽度年表与上年生长季(5—6月和8—10月)和当年生长季(3—6月和10月)气温因子显著负相关。而昆仑圆柏树木年轮宽度年表与上年生长季末期(10月)和当年生长季前期(4—6月)气温因子显著负相关。结合图 2, 气温的趋势变化与土壤水分贮存量趋势变化相反。这说明雪岭云杉和昆仑圆柏的树木径向生长主要受气温上升的水分胁迫影响, 尤其是在生长季前期, 其中雪岭云杉的树木径向生长受气温的限制作用的月份比昆仑圆柏更广泛。同时, 雪岭云杉的一阶自相关系数大于昆仑圆柏(表 1), 表明雪岭云杉受上年气温因子响应的“滞后效应”明显强于昆仑圆柏。相关性结果也表明上年气温对雪岭云杉树轮宽度生长的影响强于昆仑圆柏。有研究表明, 气温回升树木体内代谢加快, 其与大气、土壤间的物质和能量交换加快, 产生树干液流[54]。影响树干液流的主导因子为光合有效辐射强度[55], 并且液流通量和气温的关系较弱, 呈负相关, 与土壤因子也密切相关[56—58]。在气候变暖的情况下, 生长季前期气温偏高加大了土壤蒸发量和树木蒸腾作用[59], 从而使树木液流减少, 致使水分胁迫加重, 使得光合作用减弱, 影响树木径向生长, 导致当年形成窄轮。在生长季中期(7—8月), 伊朗副高东扩, 可能会在新疆西南部形成暴雨(7月下旬至8月上旬)[60], 从而缓解这段时间及之后月份的气温对径向生长的抑制作用。在7月之后, 气温对树木径向生长的限制作用影响不再显著。同时, 雪岭云杉属于浅根性树种, 具有耐阴, 耐寒、喜湿润的特点, 而昆仑圆柏具有喜光、耐寒、抗旱的特性[61], 这表明气温升高导致的水分胁迫对雪岭云杉的影响较昆仑圆柏更敏感。树木年轮的宽窄主要受到形成层细胞活性的影响, 树木生长初期形成层非结构性碳水化合物(NSC)越多细胞活性越强[62—63]。同时, 上年不良的气候条件会限制新芽、叶片和根系的形成[52], 从而影响当年生长季前期树木光合作用对水分季无机盐的吸收以及缺乏NSC形成层降低细胞活性, 进而导致当年年轮宽度偏窄。因此, 上年生长季气温对雪岭云杉和昆仑圆柏树木径向生长具有较强的影响。
在降水方面, 雪岭云杉树木年轮宽度年表与当年3月降水因子显著正相关;昆仑圆柏树木年轮宽度年表与在当年生长季开始的前后时段(3—4月、6月)降水因子显著正相关。树木生长季前期水分和养分的储存及生长季充足的水分有利于树木正常发育和宽轮的形成[52], 这表明当年生长季初期降水对雪岭云杉和昆仑圆柏的树木径向生长具有促进作用。从前文分析可知, 气温对雪岭云杉整个生长季的径向生长起限制作用, 而仅对昆仑圆柏生长季前期的生长有限制作用。因此, 雪岭云杉和昆仑圆柏树木径向生长对生长季前期的降水呈正相关关系。此外, 昆仑圆柏树木径向生长还与6月降水显著正相关。当年6月树木生长预备步入快生时期[64—65], 气温刚刚回升, 充足的降水(图 2)有利于树木年轮生长。对比气温和降水因子对雪岭云杉和昆仑圆柏树木径向生长响应情况可以发现, 当年生长季开始的前后时段气候变化对雪岭云杉和昆仑圆柏树木径向生长均影响较为显著, 表明生长季开始的前后时段是研究区影响针叶树种树木年轮宽度形成的主要时期。前文研究可知, 20世纪80年代以来, 土壤水分贮存量呈不断下降变化趋势(图 2)。这也表明了在新疆西南部气候暖湿化过程[66]中, 气温对树木生长的限制作用大于降水的促进作用。随着气候不断变暖, 可能将不利于研究区的雪岭云杉和昆仑圆柏的树木径向生长, 其中对云杉树木生长影响可能相对更大。
3.3 树木径向生长与气候因子关系的稳定性通过分析雪岭云杉和昆仑圆柏在1968—1993年和1995—2020年两个时期的树木年轮宽度年表和断面生长增量(BAI)变化趋势(图 3和图 4)发现, 雪岭云杉和昆仑圆柏树木径向生长具有相同的变化趋势, 1968—1993年时段内, 树木径向生长呈上升趋势, 1994年之后, 树木径向生长呈下降趋势。两种针叶树种与气温和降水均未出现差异现象(表 2)。从前文分析可知, 雪岭云杉和昆仑圆柏树木径向生长主要与当年生长季的气候因子相关, 因此选择使用了当年生长季平均气温、降水量和干旱指数与两种针叶树种宽度年表做31年滑动相关分析, 探索在气候变化背景下, 树木径向生长对气候因子总体响应趋势变化。从图 9中也可以看出, 随着气候变暖, 树木径向生长对生长季气温的响应从正相关转化为负相关, 且两种针叶树种树轮宽度对气温负响应相关显著增强。其中气温突变后, 雪岭云杉对气温响应的敏感性则高于昆仑圆柏。同时, 树轮宽度年表与生长季自适应帕默尔干旱指数(scPDSI)、标准化降水蒸散指数(SPEI)、气象干旱综合指数(MCI)的滑动相关分析结果表明(图 10), 树木径向生长与生长季干旱指数之间的相关性持续加强, 对树木径向生长的干旱胁迫不断加剧, 与上述对气温的响应相对应。

|
| 图 9 树木年轮宽度年表与生长季均温和降水滑动相关分析 Fig. 9 The moving correlation analysis between the tree-ring width chronology and mean temperature and precipitation during growing season 虚线表示0.05显著性 |

|
| 图 10 树木年轮宽度年表与生长季干旱指数滑动相关分析 Fig. 10 The moving correlation analysis between the tree-ring width chronology and the drought indices during growing season 虚线表示0.05显著性 |
图 8和图 9中总体仍表现出气温因子对树木生长的限制作用在加强以及降水因子对树木生长促进作用增强(雪岭云杉树木径向生长的降水促进作用未达到显著性水平), 但针叶树种在同一时段对气温和降水的响应是有差异的。通过单相关分析(图 6和图 7)和滑动相关分析(图 8和图 9)可以发现, 雪岭云杉树木径向生长对气温因子负响应增强的月份均多于昆仑圆柏;对降水因子正响应增强则昆仑圆柏强于雪岭云杉。这可能与雪岭云杉耐阴, 耐寒、喜湿润和昆仑圆柏具有耐寒、耐旱[61]有关, 也说明突变后雪岭云杉对气温的响应敏感性强于昆仑圆柏, 而对降水响应敏感性要弱于昆仑圆柏。在上年生长季前期(4—7月)和当年生长季末期(8—10月), 随着气候不断变暖, 雪岭云杉和昆仑圆柏树木径向生长与气温因子的相关系数都存在由正转负且不断加强(图 8)。研究表明, 蒸散发(ET)的增加趋势主要由地表温度升高驱动[67], 而一定阈值范围的温度对北半球针叶径向生长有促进作用[68], 而树木液流主要与有效光合作用和土壤水分有关[54—57]。气温上升逐步超出了树木径向生长温度阈值, 加快了土壤水分的散失(图 2), 减缓了树干液流的产生, 开始持续限制树木的径向生长。然而, 在当年3—7月(早材形成时期)降水因子对两种针叶树种树木径向生长的影响基本上保持促进作用(图 8)。在内蒙古东北部樟子松也与上年秋冬季和当年春夏季的降水的制约[69]。研究表明干旱和半干旱地区针叶树木年轮宽度与上年的秋冬季和当年春夏季的气候状况关系密切[41]。上年秋冬季降雨和降雪会在当年春末夏初树木早材生长时提供水分, 每年5—7月是树木早材生长的时期, 这一时期充沛的降水也有利于当年早材形成宽轮[69]。降水因子对树木径向生长的影响前后一致性比较好, 说明在西昆仑山区域两种针叶树种树木年轮宽度对降水响应的稳定性较好。
综上所述, 气候变暖对该研究区域的雪岭云杉和昆仑圆柏树木径向生长产生了一定影响。以上结论是否适用于西昆仑山其它区域, 则需要在更大范围和更多树木年轮样本的基础上继续开展分析研究。
4 结论全球变暖的背景下, 气候变化已经对世界森林生态系统产生深远影响。本研究以西昆仑山奥依塔克地区的雪岭云杉和昆仑圆柏为研究对象, 分析其树木生长特征以及在气候变化情况下对气候要素的响应。研究发现:近70年来气候变暖, 导致西昆仑山奥依塔克地区的混交林中雪岭云杉和昆仑圆柏树木径向生长趋势在气候突变前后变化不同, 即两种针叶树种在气温突变前树木宽度年表指数和BAI呈上升趋势, 然而气温突变后树木宽度年表指数和BAI呈下降趋势。并且, 昆仑圆柏的树木径向生长速率高于雪岭云杉。u值检验表明, 两种针叶树种树木径向生长对气温和降水均无明显差异。相关性分析结果表明, 雪岭云杉径向生长与上年生长季(5—6月和8—10月)和当年生长季(3—6月和10月)气温呈显著负相关, 而昆仑圆柏径向生长与上年10月和当年生长季前期(4—6月)气温呈显著负相关;两者均与当年春季降水呈显著正相关。从滑动相关分析中发现, 雪岭云杉和昆仑圆柏树木径向生长对气候的响应中, 上年和当年生长季气温对树木径向生长的限制作用在增强, 尤其在当年生长季前期更为显著;并且当年生长季前期降水对树木径向生长的促进作用也在增强。气候突变(1994年)后, 两种针叶树种对气温和降水响应的敏感性均有所增强, 并且雪岭云杉对气温的响应敏感性强于昆仑圆柏, 而对降水响应敏感性要弱于昆仑圆柏。随着气候不断变暖, 可能将不利于研究区两种针叶树种的树木径向生长, 其中对雪岭云杉树木生长影响可能相对更大。
| [1] |
秦莉, 尚华明, 张同文, 刘卫平, 张瑞波. 天山南北坡树轮稳定碳同位素对气候的响应差异. 生态学报, 2021, 41(14): 5713-5724. |
| [2] |
张辉, 张芸, 胡亚楠, 颜耀, 吴鹏飞, 曾爱聪, 马祥庆. 气候变暖背景下杉木年轮密度对气候因子的响应. 生态学报, 2021, 41(4): 1551-1563. |
| [3] |
Bonan G B. Forests and climate change: forcings, feedbacks, and the climate benefits of forests. Science, 2008, 320(5882): 1444-1449. DOI:10.1126/science.1155121 |
| [4] |
Pan Y D, Birdsey R A, Fang J Y, Houghton R, Kauppi P E, Kurz W A, Phillips O L, Shvidenko A, Lewis S L, Canadell J G, Ciais P, Jackson R B, Pacala S W, McGuire A D, Piao S L, Rautiainen A, Sitch S, Hayes D. A large and persistent carbon sink in the world' s forests. Science, 2011, 333(6045): 988-993. DOI:10.1126/science.1201609 |
| [5] |
高洪娜, 高瑞馨. 气象因子对树木生长量影响研究综述. 森林工程, 2014, 30(2): 6-9. DOI:10.3969/j.issn.1001-005X.2014.02.002 |
| [6] |
王晓春, 张小全, 朱建华, 侯振宏, 柴正礼. 树木年轮与全球变暖的关系研究进展. 世界林业研究, 2009, 22(6): 38-42. DOI:10.13348/j.cnki.sjlyyj.2009.06.005 |
| [7] |
IPCC. Climate change, the physical science basis. Cambridge: Cambridge University Press, 2013.
|
| [8] |
黄建平, 陈文, 温之平, 张广俊, 李肇新, 左志燕, 赵庆云. 新中国成立70年以来的中国大气科学研究: 气候与气候变化篇. 中国科学: 地球科学, 2019, 49(10): 1607-1640. |
| [9] |
Huang J P, Wang T H, Wang W C, Li Z Q, Yan H R. Climate effects of dust aerosols over East Asian arid and semiarid regions. Journal of Geophysical Research: Atmospheres, 2014, 119(19): 11398-11416. |
| [10] |
尚建勋, 时忠杰, 高吉喜, 徐丽宏, 吕世海, 冯朝阳, 王鲁秀. 呼伦贝尔沙地樟子松年轮生长对气候变化的响应. 生态学报, 2012, 32(4): 73-80. |
| [11] |
尚华明, 魏文寿, 袁玉江, 喻树龙, 张同文, 瓦合提·艾则买提, 李新建. 阿尔泰山南坡树轮宽度对气候变暖的响应. 生态学报, 2010, 30(9): 2246-2253. |
| [12] |
Franks S J, Weber J J, Aitken S N. Evolutionary and plastic responses to climate change in terrestrial plant populations. Evolutionary Applications, 2014, 7(1): 123-139. DOI:10.1111/eva.12112 |
| [13] |
杨柳, 李静茹, 彭剑峰, 霍嘉新, 陈亮. 1801年以来河南尧山地区油松高温变化及影响机制. 生态学报, 2021, 41(1): 79-91. DOI:10.3969/j.issn.1673-1182.2021.01.015 |
| [14] |
贾飞飞, 孙翠洋, 孙红月, 李鑫. 昌岭山两个优势树种径向生长对气候变化的响应. 生态学报, 2019, 39(17): 6332-6340. |
| [15] |
秦进, 白红英, 刘荣娟, 翟丹平, 苏凯, 王俊, 李书恒. 近144年来秦岭太白山林线区3-6月平均气温的重建. 生态学报, 2017, 37(22): 7585-7594. |
| [16] |
雷帅, 张劲松, 孟平, 何子兵, 程顺, 孙守家. 中国北部不同地点樟子松人工林径向生长对气候响应的差异. 生态学报, 2020, 40(13): 4479-4492. |
| [17] |
牛豪阁, 张芬, 于爱灵, 王放, 张军周, 勾晓华. 祁连山东部青杄年内径向生长动态对气候的响应. 生态学报, 2018, 38(20): 7412-7420. |
| [18] |
王延芳, 张永香, 勾晓华, 高琳琳, 王放. 祁连山中部低海拔地区青海云杉径向生长的气候响应机制. 生态学报, 2020, 40(1): 161-169. |
| [19] |
金敏艳, 李进军, 车宗玺, 王放, 张军周, 勾晓华. 祁连山中部祁连圆柏年内径向生长对气候因子的响应. 生态学报, 2020, 40(21): 7699-7708. |
| [20] |
尚华明, 魏文寿, 袁玉江, 木太力普·托乎提, 陆恒, 张同文, 范子昂, 陈峰, 秦莉. 帕米尔东北部昆仑圆柏850 a树轮宽度年表的建立及其气候意义. 沙漠与绿洲气象, 2015, 9(1): 6-11. |
| [21] |
尚华明, 麦麦提图尔荪·克比尔, 栗红, 喻树龙, 张瑞波, 张同文, 陈峰, 陆恒, 魏文寿, 袁玉江. 叶尔羌河流域雪岭云杉树轮宽度气候信息的探讨. 沙漠与绿洲气象, 2017, 11(3): 17-24. |
| [22] |
杨美琳, 他志杰, 吴燕良, 赵鹏, 于瑞德. 树轮宽度记录的塔吉克斯坦北部160 a平均最低气温变化. 干旱区研究, 2019, 36(2): 290-295. |
| [23] |
Opala M, Niedzwiedz T, Rahmonov O. The dendroclimatic potential of shrubs from western Pamir-Alay (Tajikistan). Trace 2014 Tree Rings in Archaeclogy, Climatology and Ecology, 2014. |
| [24] |
Yang M L, Yu Y, Zhang H Y, Wang Q, Gan M, Yu R D. Tree ring based drought variability in Northwest Tajikistan since 1895 AD. Journal of Arid Land, 2020, 12(3): 413-422. |
| [25] |
Chen F, Zhang T W, Andrea S, Yu S L, Zhang R B, Linderholm H W, Kobuliev Z V, Ahsan A, Anvar K. Juniper tree-ring data from the Kuramenian Mountains (Republic of Tajikistan), reveals changing summer drought signals in western Central Asia. Climate of the Past Discussions, 2018, 1-27. |
| [26] |
高琳琳, 勾晓华, 邓洋, 张芬, 方克艳. 树轮气候学中分异现象的研究进展. 冰川冻土, 2011, 33(2): 453-460. |
| [27] |
Gao L L, Gou X H, Deng Y, Zhang F, Fang K Y. An overview of the divergence phenomenon in dendroclimatology. Journal of Glaciology and Geocryology, 2011, 33(2): 453-460. |
| [28] |
D'Arrigo R, Wilson R, Liepert B, Cherubini P. On the 'Divergence Problem' in Northern Forests: a review of the tree-ring evidence and possible causes. Global and Planetary Change, 2008, 60(3/4): 289-305. |
| [29] |
Büntgen U, Frank D C, Schmidhalter M, Neuwirth B, Seifert M, Esper J. Growth/climate response shift in a long subalpine spruce chronology. Trees, 2006, 20(1): 99-110. |
| [30] |
loyd A H, Bunn A G. Responses of the circumpolar boreal forest to 20th century climate variability. Environmental Research Letters, 2007, 2(4): 045013. |
| [31] |
方克艳, 陈秋艳, 刘昶智, 曹春福, 陈亚君, 周非飞. 树木年代学的研究进展. 应用生态学报, 2014, 25(7): 1879-1888. |
| [32] |
郭明明, 张远东, 王晓春, 黄泉, 杨素香, 刘世荣. 升温突变对川西马尔康树木生长的影响. 生态学报, 2015, 35(22): 7464-7474. |
| [33] |
Zhang T W, Zhang R B, Jiang S X, Bagila M, Ainur U, Yu S L. On the 'divergence problem' in the alatau mountains, central Asia: a study of the responses of schrenk spruce tree-ring width to climate under the recent warming and wetting trend. Atmosphere, 2019, 10(8): 473. |
| [34] |
刘蕊, 王勇辉, 姜盛夏, 张瑞波, 秦莉, Bulkajyr T.Mambetov, Nurzhan Kelgenbayev, Daniyar Dosmanbetov, Bagila Maisupova, 张同文. 哈萨克斯坦阿尔泰山树木径向生长及其对气候要素的响应. 干旱区研究, 2019, 36(3): 723-733. |
| [35] |
康剑, 蒋少伟, 黄建国. 阿尔泰山萨彦岭4种优势树种径向生长对气候因子的响应. 生态学报, 2020, 40(17): 6135-6146. |
| [36] |
李宗善, 陈维梁, 韦景树, 买尔当·克依木, 张育新, 张霜, 王晓春. 北京东灵山辽东栎林树木生长对气候要素的响应特征. 生态学报, 2021, 41(1): 27-37. |
| [37] |
苟晓霞, 张同文, 喻树龙, 张瑞波, 姜盛夏, 郭玉琳. 不同生境下圆柏径向生长的气候响应. 生态学杂志, 2021, 40(6): 1574-1588. |
| [38] |
尚华明, 范煜婷, 张瑞波, 喻树龙, 张同文, 魏文寿, 刘卫平. 帕米尔高原东部径流量变化及其对气候变化的响应. 气候变化研究进展, 2021, 17(3): 352-360. |
| [39] |
郑度, 张百平. 喀喇昆仑山-西昆仑山地区的垂直自然带、环境和自然保护问题. 自然资源学报, 1989, 4(3): 254-266. |
| [40] |
徐海燕. 中国帕米尔高原合瓣花类(除菊科)植物研究[D]. 石河子: 石河子大学, 2007.
|
| [41] |
吴祥定. 树木年轮与气候变化. 北京: 气象出版社, 1990: 170-171.
|
| [42] |
周子建, 江源, 董满宇, 陶岩, 王明昌, 丁新原. 长白山北坡不同海拔红松径向生长-气候因子关系对气温突变的响应. 生态学报, 2018, 38(13): 4668-4676. |
| [43] |
Jiao L, Jiang Y, Wang M C, Kang X Y, Zhang W T, Zhang L N, Zhao S D. Responses to climate change in radial growth of Picea schrenkiana along elevations of the eastern Tianshan Mountains, northwest China. Dendrochronologia, 2016, 40: 117-127. |
| [44] |
Jiao L, Jiang Y, Zhang W T, Wang M C, Zhang L N, Zhao S D. Divergent responses to climate factors in the radial growth of Larix sibirica in the eastern Tianshan Mountains, northwest China. Trees, 2015, 29(6): 1673-1686. |
| [45] |
WMO. Calculation of monthly and annual 30-year standard normals, WCDP-No., 1989, 10, WMO-TD/No. 341. World Meteorological Organization, Geneva.
|
| [46] |
Wigley T M L, Briffa K R, Jones P D. On the average value of correlated time series, with applications in dendroclimatology and hydrometeorology. Journal of Climate and Applied Meteorology, 1984, 23(2): 201-213. |
| [47] |
Fritts H C. Tree Rings and Climate. London: Academic Press, 1976, 268-273, 295-300. |
| [48] |
张同文, 袁玉江, 魏文寿, 张瑞波, 喻树龙, 陈峰, 尚华明, 秦莉. 开都河流域天山桦树轮宽度年表的建立及其气候响应. 生态学报, 2015, 35(9): 3034-3042. |
| [49] |
王忠. 植物生理学. 北京: 中国农业出版社, 2000.
|
| [50] |
张同文, 袁玉江, 魏文寿, 尚华明, 喻树龙, 张瑞波, 陈峰, 范子昂, 秦莉. 浑善达克沙地白扦树轮早晚材宽度年表对比分析. 沙漠与绿洲气象, 2016, 10(1): 47-53. |
| [51] |
喻树龙, 袁玉江, 秦莉, 尚华明, 张同文, 陈峰, 张瑞波, 范子昂. 天山北坡中部不同海拔高度雪岭云杉树轮宽度气候响应对比分析. 沙漠与绿洲气象, 2016, 10(3): 30-38. |
| [52] |
秦莉, 袁玉江, 喻树龙, 范子昂, 尚华明, 陈峰, 张同文. 赛里木湖流域雪岭云杉(Picea schrenkiana)树木径向生长对气候变化的响应. 中国沙漠, 2015, 35(1): 113-119. |
| [53] |
董志鹏, 郑怀舟, 方克艳, 严容, 郑立伟, 杨玉盛. 福建三明马尾松树轮宽度对气候变化的响应. 亚热带资源与环境学报, 2014, 9(1): 1-7. |
| [54] |
白志强, 刘华, 佘春燕, 刘端, 韩燕梁. 西伯利亚落叶松树干液流的动态变化. 河北农业大学学报, 2016, 39(3): 49-54. |
| [55] |
吉春容, 邹陈, 范子昂, 李新建. 天山中段雪岭云杉树干液流变化及其与气象因子的关系. 干旱区资源与环境, 2013, 27(12): 119-123. |
| [56] |
王海珍, 韩路, 李志军. 胡杨、灰叶胡杨蒸腾耗水规律初步研究. 干旱区资源与环境, 2009, 23(8): 186-189. |
| [57] |
解婷婷, 张希明, 单立山, 梁少民, 杨小林. 灌溉量对多枝柽柳水分生理及生长的影响. 干旱区研究, 2008, 25(6): 802-807. |
| [58] |
张小由, 康尔泗, 司建华, 周茅先. 额济纳绿洲中柽柳耗水规律的研究. 干旱区资源与环境, 2006, 20(3): 159-162. |
| [59] |
张同文, 王丽丽, 袁玉江, 魏文寿, 喻树龙, 张瑞波, 陈峰, 尚华明, 范子昂. 利用树轮宽度资料重建天山中段南坡巴仑台地区过去645年来的降水变化. 地理科学, 2011, 31(2): 251-256. |
| [60] |
张俊兰, 罗继, 王荣梅. 近20年新疆升温融雪(冰)型洪水频次时空变化及大气环流型分析. 干旱区研究, 2021, 38(2): 339-350. |
| [61] |
杨昌友, 沈冠冕, 毛祖美. 新疆植物志. 乌鲁木齐: 新疆科局卫生出版, 1993.
|
| [62] |
Simard S, Giovannelli A, Treydte K, Traversi M L, King G M, Frank D, Fonti P. Intra-annual dynamics of non-structural carbohydrates in the cambium of mature conifer trees reflects radial growth demands. Tree Physiology, 2013, 33(9): 913-923. |
| [63] |
Kozlowski T T. Carbohydrate sources and sinks in woody plants. The Botanical Review, 1992, 58(2): 107-222. |
| [64] |
刘禹, 马利民. 树轮宽度对近376年呼和浩特季节降水的重建. 科学通报, 1999, 44(18): 1986-1992. |
| [65] |
朱海峰, 王丽丽, 邵雪梅, 方修琦. 雪岭云杉树轮宽度对气候变化的响应. 地理学报, 2004, 59(6): 863-870. |
| [66] |
胡汝骥, 姜逢清, 王亚俊, 樊自立. 新疆气候由暖干向暖湿转变的信号及影响. 干旱区地理, 2002, 25(3): 194-200. |
| [67] |
Pascolini-Campbell M, Reager J T, Chandanpurkar H A, Rodell M. A 10 per cent increase in global land evapotranspiration from 2003 to 2019. Nature, 2021, 593. |
| [68] |
Huang J G, Ma Q Q, Rossi S, Biondi F, Deslauriers A, Fonti P, Liang E Y, Mäkinen H, Oberhuber W, Rathgeber C B K, Tognetti R, Treml V, Yang B, Zhang J L, Antonucci S, Bergeron Y, Camarero J J, Campelo F, Čufar K, Cuny H E, de Luis M, Giovannelli A, Gričar J, Gruber A, Gryc V, Güney A, Guo X L, Huang W, Jyske T, Kašpar J, King G, Krause C, Lemay A, Liu F, Lombardi F, Martinez del Castillo E, Morin H, Nabais C, Nöjd P, Peters R L, Prislan P, Saracino A, Swidrak I, Vavrčík H, Vieira J, Yu B Y, Zhang S K, Zeng Q, Zhang Y L, Ziaco E. Photoperiod and temperature as dominant environmental drivers triggering secondary growth resumption in Northern Hemisphere conifers. PNAS, 2020, 117(34): 20645-20652. |
| [69] |
张同文, 袁玉江, 魏文寿, 喻树龙. 内蒙古东北部莫尔道嘎地区树轮年表的建立. 干旱区资源与环境, 2009, 23(12): 177-182. |