 2022, Vol. 42
2022, Vol. 42文章信息
- 王亚如, 陈乐, 房玮, 张盼月, 吴彦, 张光明, 王洪杰, 付川
- WANG Yaru, CHEN Le, FANG Wei, ZHANG Panyue, WU Yan, ZHANG Guangming, WANG Hongjie, FU Chuan
- 典型冷暖季沉水植物凋落物分解特性及沉积物微生物群落变化
- Decomposition characteristics of typical warm season and cold season submerged plant litters and change of microbial community in sediment
- 生态学报. 2022, 42(24): 10214-10225
- Acta Ecologica Sinica. 2022, 42(24): 10214-10225
- http://dx.doi.org/10.5846/stxb202106281715
-
文章历史
- 收稿日期: 2021-06-28
- 网络出版日期: 2022-07-29





2. 北京林业大学环境科学与工程学院污染水体源控制与生态修复技术北京市高等学校工程研究中心, 北京 100083;
3. 重庆三峡学院三峡库区水环境演变与污染防治重庆市重点实验室, 万州 404020;
4. 河北工业大学能源与环境工程学院, 天津 300401;
5. 河北大学雄安生态研究院, 保定 071002
2. Engineering Research Center for Water Pollution Source Control & Eco-remediation, College of Environmental Science and Engineering, Beijing Forestry University, Beijing 100083, China;
3. Chongqing Key Laboratory of Water Environment Evolution and Pollution Control in Three Gorges Reservoir, Chongqing Three Gorges University, Wanzhou 404020, China;
4. School of Energy and Environmental Engineering, Hebei University of Technology, Tianjin 300401, China;
5. Xiong'an Institute of Eco-Environment, Hebei University, Baoding 071002, China
白洋淀是华北最大的淡水湖泊, 其水质对保障雄安新区生态安全具有重要意义。对于湖泊系统尤其是浅水湖泊, 沉水植物对水生生态系统的物质循环和能量流动发挥重要作用[1]。白洋淀冷季和暖季水生植物种类发生更替, 不仅维持冷季和暖季的生态系统稳定, 同时, 这些植物也分别在冷季和暖季发挥重要的水质净化作用[2]。然而, 每到季节更替期, 冷暖季沉水植物大量衰亡, 死亡的植物残体会堆积于湖泊水体中。在其衰老、死亡过程中, 植物腐烂分解所释放的营养物质导致水质季节性恶化[3]。因此, 沉水植物分解对水质的影响值得关注。
凋落物分解过程中的养分释放及其对水质的影响已有部分研究[4—6]。Balasubramanian等[4]研究了水葫芦叶、茎、根在不同营养条件下的分解速率和养分释放规律, 发现在水葫芦不同植物部位的碳(C)、氮(N)、磷(P)、木质素、半纤维素和纤维素的含量、分解速率存在显著差异。Wu等[7]研究了3种不同水生植物凋落物分解过程对水质的影响, 发现水葫芦的分解对水质的影响大于轮叶黑藻和香蒲, 这可能与植物凋落物的化学组成和结构相关。
溶解性有机物(DOM)是由不同分子特征的高度异质性混合物组成[8], 如腐殖质物质、蛋白质等, 是分解过程中的主要分解产物[9]。DOM中不同组分在水环境中发挥着不同的作用, 如DOM芳香度越高, 其可降解性越低。因此, DOM组分和特性对上覆水质有重要影响。然而, 少有研究关注植物凋落物分解过程中水中DOM的释放过程和结构特征的变化。微生物对植物凋落物的分解过程有很大贡献[10—11], 尤其在凋落物降解和破碎阶段发挥着重要作用[12], 但是参与植物凋落物分解各个阶段的主要微生物对凋落物分解的机制仍不清楚。
沉水植物的种类和生长发育期决定了植物吸收的营养成分不同, 也会导致其凋落物衰亡分解对水体的影响不同。金鱼藻和菹草分别属于白洋淀夏季和春季的强耐污种和强生长种[13], 广泛分布于白洋淀水生生态系统。因此, 本研究选择金鱼藻和菹草, 监测其凋落物分解释放的营养物质变化, 分析其分解释放特性以及参与分解的微生物群落变化, 揭示微生物群落与植物凋落物分解之间的关系、分解过程中主要分解产物DOM与微生物间的关系, 研究结果可为湿地植物管理提供技术指导。
1 材料与方法 1.1 植物和底泥植物与底泥均采自白洋淀。实验选取金鱼藻(Ceratophyllum demersum L.)、菹草(Potamogeton crispus L.)两种沉水植物作为研究材料。植物用自来水冲洗干净, 风干3 d, 在85 ℃下烘箱干燥至恒重。将干燥的样品切成约1 cm长的切片, 茎、叶均匀混合。将烘干后的植物分为两部分, 一部分用于分解实验, 另一部分测试初始凋落物C、N、P含量, 结果见表 1。
| 植物凋落物Plant litters | 碳C/% | 氮N/% | 磷P/% |
| 金鱼藻Ceratophyllum demersum L. | 37.53±0.59 | 3.19±0.28 | 0.40±0.001 |
| 菹草Potamogeton crispus L. | 39.33±1.03 | 3.83±0.34 | 0.42±0.002 |
将1 g烘干植物凋落物放入100目尼龙网袋中(孔径为0.2 mm, 10 cm×10 cm), 然后放入装有白洋淀底泥(约2 cm高)的2 L塑料烧杯中模拟凋落物分解。缓慢加入2 L自来水, 放置2 d, 与底泥进行充分传质模拟湿地上体, 初始水质见表 2。将不含植物凋落物(空白组)、含1 g植物凋落物(实验组)的尼龙网袋置于反应器中, 放置60 d。整个反应避光进行。所有的植物凋落物均浸于水下, 置于底泥上。于第0、2、4、6、8、10、15、20、30、40、50天和第60天采集上覆水水样进行检测, 并补充等量的自来水。
| 理化性质 Physicochemical properties |
温度 Temperature/℃ |
pH | 溶解氧 Dissolved oxygen/ (mg/L) |
总有机碳 Total organic carbon/(mg/L) |
总氮 Total nitrogen/ (mg/L) |
总磷 Total phosphorus/ (mg/L) |
| 数值Value | 11.0 | 7.86 | 7.81 | 23.82 | 0.90 | 0.008 |
使用元素分析仪测定植物凋落物中的碳和氮;浓硝酸—高氯酸消解后, 采用国家标准方法(国家环保总局《水和废水监测分析方法》第四版)测定最终溶液中的磷[14];植物凋落物失重率计算如式(1):
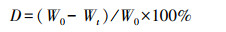
|
(1) |
式中, D为时间t时的失重率(%);Wt为时间t时的重量(g);W0为初始凋落物的重量(g)。
水样水质根据国家标准方法(国家环保总局《水和废水监测分析方法》第四版)测定氨态氮(NH4+-N)、硝态氮(NO3--N)、总氮(TN)和总磷(TP);溶解态的总有机碳(TOC)使用岛津TOC分析仪测定;利用紫外-可见光分光光度计(U-3900)分析水样, 波长250 nm和365 nm下吸收系数的比值为分子量大小参数值(M)[15], 可以反映DOM的分子量大小。所有测量重复3次, 取平均值。
1.3 DOM荧光分析利用荧光分光光度计(日立F-700)的三维荧光光谱(EEMs)分析水中的DOM, 水样过0.45 μm微孔滤膜, 扫描波长范围设置为:Ex=200—450 nm, Em=250—600 nm, 狭缝宽带为5 nm。采用EEMs结合平行因子法(EEMs-PARAFAC法)解析水体DOM的理化性质[1]。
1.4 DNA提取与微生物群落分析取初始沉积物作为对照(CK), 分别取金鱼藻和菹草分解4 d(释放营养物质浓度最高点, Cd-4和Pc-4)、15 d(分解缓慢期, Cd-15和Pc-15)和30 d(分解相对稳定期, Cd-30和Pc-30)时的沉积物作为实验组, 送至上海美吉生物医药科技有限公司进行Illumina高通量测序。
1.5 数据处理与分析用Origin 9.0绘制图表。采用Matlab 2015a软件进行EEMs-PARAFAC分析。通过Chao指数来表征微生物的丰度, 通过Shannon指数、Simpson指数反映微生物的多样性[16]。通过冗余分析(RDA)得到溶解性有机物各组分与微生物群落的相关性[17]。
2 结果与分析 2.1 植物凋落物分解特性两种植物凋落物质量损失率变化如图 1所示, 15 d内质量损失率迅速升高, 15 d后增加速度减缓, 45 d后基本不再变化。实验结束时(60 d), 金鱼藻和菹草凋落物质量损失率分别达到61.43%和66.72%。由图 1可以看出, TN、TP浓度变化呈先增加(0—15 d)后下降(15—30 d)直至稳定(30—60 d)的趋势, 可能由于凋落物的分解和微生物的活动, 导致了N、P营养物的快速释放和消耗。0—30 d, TN浓度高于1.5 mg/L(地表水环境质量标准IV类标准), 15—30 d, TN浓度基本低于1.5 mg/L(Ⅳ类标准);0—30 d, TP浓度高于0.1 mg/L(湖、库Ⅳ类标准);实验结束时(60 d), 金鱼藻和菹草凋落物分解体系TN(0.91 mg/L和1.19 mg/L)和TP(0.06 mg/L和0.09 mg/L)浓度均低于Ⅳ类标准, 说明植物凋落物分解对水质的影响呈阶段性, 分解初期(0—15 d)可暂时导致水体的恶化。0—4 d NH4+-N和NO3--N浓度较高, 占TN浓度的90%以上, 所以有机氮浓度较低, 4—60 d则相反, 表明植物凋落物分解过程中, 氮素主要以有机氮的形式释放到上覆水体中, 由于微生物分解有机氮导致NH4+-N和NO3--N浓度增加, 同时也存在NH4+-N和NO3--N的转化(图 1)。这与Wu等[7]和Lemley等[18]的研究不同, 说明不同植物在分解过程中氮释放形态有显著差异。

|
| 图 1 植物凋落物分解过程中质量损失率和上覆水中TN、NH4+-N、NO3--N和TP浓度变化 Fig. 1 Mass loss rate and concentration of TN, NH4+-N, NO3--N and TP during decomposition of plant litters |
两种植物凋落物分解过程中上覆水TOC浓度与TN、TP浓度变化趋势相似(图 1和图 2)。0—15 d, 凋落物分解使上覆水DOM浓度迅速增加, 与凋落物失重率变化一致(图 1), 说明分解初期凋落物中可溶性有机物(如多糖和羧酸)的快速溶出导致质量损失较快[19], 15—60 d下降直至稳定(低于初始值), 可能与植物凋落物有机物含量减少和微生物的降解有关。M值可以反映DOM的分子量大小, 即腐殖酸(M < 3.5)和富里酸(M>3.5)等在DOM中所占的比例[15]。由图 2可知, 不同分解期M值均大于3.5, 主要反映了富里酸的吸收特性, 这说明上覆水DOM分子量较小, 主要以富里酸为主, 而对照组M值小于3.5, 表明其DOM主要以腐殖酸为主。综上, 表明两种植物凋落物分解释放的DOM呈现低腐殖化特征。

|
| 图 2 植物凋落物分解过程中TOC浓度和M值的变化 Fig. 2 Changes of TOC concentrations and M value during decomposition of plant litters |
两种植物凋落物分解过程上覆水DOM的EEMs如图 3所示, 均检测出四个明显的荧光峰(T1、T2、A、C), 峰T1、T2分别为可见光区类、紫外光区类蛋白物质, 峰A、C分别为类富里酸、类腐殖酸物质。EEMs表明, 0—15 d上覆水DOM以类蛋白物质为主;15 d后类蛋白荧光强度减弱, 直到45 d未明显检测到, 而类富里酸和类腐殖酸物质荧光强度逐渐增强, 说明植物凋落物分解首先向水中释放易生物降解的物质如类色氨酸, 然后释放类富里酸和类腐殖酸等难降解有机化合物。因此, 植物凋落物分解后期(15—60 d)由于难降解有机物含量的增加, 可能会加剧水体甚至是沉积物的腐殖化程度, 影响湖泊水体颜色和嗅味等物理化学指标, 甚至污染物的迁移转化和生物效应。

|
| 图 3 金鱼藻和菹草凋落物分解过程上覆水DOM的EEMs图 Fig. 3 EEMs diagram of DOM in the overlying water during decomposition of Ceratophyllum demersum L. and Potamogeton crispus L. T1:Ex(270—290 nm)/Em(330—370 nm) 类蛋白荧光峰Fluorescence peak for protein-like substances;T2:Ex(230—240 nm)/Em(330—370 nm) 类蛋白荧光峰Fluorescence peak for protein-like substances;A:Ex(230—310 nm)/Em(380—460 nm) 类富里酸荧光峰Fluorescence peak for fulvic acid-like substances;C:Ex(345—450 nm)/Em(430—530 nm) 类腐殖酸荧光峰Fluorescence peak for humic acid-like substances[20—21] |
通过PARAFAC分析得出四种不同的荧光组分, 如图 4所示。两种植物凋落物分解释放的DOM发现荧光相似, 但荧光强度值不同。组分C1在235 nm/425 nm和325 nm/425 nm(Ex/Em)处出现峰值, 为类腐殖酸和类富里酸组分[22—23];组分C2在225 nm/340 nm和275 nm/340 nm(Ex/Em)处出现峰值, 代表类色氨酸组分[24—25];组分C3在270 nm/475 nm和360 nm/475 nm(Ex/Em)处出现峰值, 代表类腐殖酸组分[23, 26];组分C4在225 nm/405 nm和305 nm/405 nm(Ex/Em)处出现峰值, 代表类腐殖酸和类富里酸组分[20—21]。四种组分的最大荧光强度百分比如图 5所示。两种植物凋落物分解过程中各组分的百分比变化不同, 其中类色氨酸物质C2在4 d达到峰值, 分别为31.45%(金鱼藻分解体系)和22.31%(菹草分解体系), 难降解物质C1、C3和C4百分比缓慢增加, 与EEMs分析结果一致(图 3)。

|
| 图 4 平行因子组分荧光光谱图 Fig. 4 Fluorescence components identified by PARAFAC model C1:组分1(类腐殖酸和类富里酸) component 1 (Humic acid-and fulvic acid-like substances);C2:组分2(类色氨酸) component 2 (Tryptophan-like substannces);C3:组分3(类腐殖酸) component 3 (Humic acid-like substances);C4:组分4(类腐殖酸和类富里酸) component 4 (Humic acid-and fulvic acid-like substances) |

|
| 图 5 金鱼藻和菹草凋落物分解过程中PARAFAC组分的最大荧光强度百分比变化 Fig. 5 Change of the maximum fluorescence intensity percentages of PARAFAC components during decomposition of Ceratophyllum demersum L. and Potamogeton crispus L. |
如表 3所示, 随着凋落物分解过程的进行, Chao指数逐渐增加, Shannon指数低于原始沉积物、Simpson指数高于原始沉积物, 表明凋落物的分解, 增加了微生物群落丰富度, 降低了微生物多样性。两种植物凋落物分解过程微生物群落丰富度和多样性存在差异, 这可能导致了其不同的分解特性, 以及凋落物分解对水体水质不同的影响(图 1和图 2)。
| 样品Samples | CK | Cd-4 | Pc-4 | Cd-15 | Pc-15 | Cd-30 | Pc-30 |
| Chao指数Chao index | 3864.62 | 3723.61 | 3190.06 | 3794.89 | 4048.03 | 3917.22 | 4135.58 |
| Shannon指数Shannon index | 6.81 | 5.60 | 4.11 | 5.66 | 6.53 | 6.32 | 6.43 |
| Simpson指数Simpson index | 0.003 | 0.040 | 0.073 | 0.027 | 0.007 | 0.010 | 0.011 |
| CK:初始沉积物作为对照Control (Initial sediment);Cd-4、Cd-15、Cd-30:金鱼藻分别分解4、15、30 d时的沉积物Sediment after Ceratophyllum demersum L. decomposition for 4, 15, 30 d, respectively;Pc-4、Pc-15、Pc-30:菹草分别分解4、15、30 d时的沉积物Sediment after Potamogeton crispus L. decomposition for 4, 15, 30 d, respectively | |||||||
微生物群落相对丰度见图 6。从门水平看, 初始沉积物和分解过程沉积物中的主要微生物群落一致, 主要为变形菌门(Proteobacteria)、厚壁菌门(Firmicutes)、放线菌门(Actinobacteriota)、绿弯菌门(Chloroflexi)和拟杆菌门(Bacteroidota)。分解初期(4 d), 沉积物中变形菌门和厚壁菌门含量增加, 说明在植物凋落物分解快速期变形菌门、厚壁菌门这2类微生物起重要作用。分解缓慢期(15 d)和分解相对稳定期(30 d), 沉积物中拟杆菌门含量增加, 表明15 d后, 除了变形菌门、厚壁菌门这2类微生物, 降解难降解有机质的拟杆菌门发挥了重要作用[27]。从属水平看, 在植物凋落物分解过程中, 沉积物中主要微生物群落发生了明显变化。分解初期(4 d), 沉积物中主要微生物为假单胞菌属(Pseudomonas), 在金鱼藻和菹草凋落物分解体系分别占26.79%和34.65%。分解缓慢期(15 d)和分解相对稳定期(30 d), 假单胞菌属含量大幅降低, 而芽孢杆菌属(Bacillus)、Malikia属和黄杆菌属(Flavobacterium)含量增加, 分别占9.04%、9.62%和4.39%(金鱼藻分解体系), 2.61%、8.16%和1.63%(菹草分解体系)。假单胞菌属和Malikia属(变形菌门)、芽孢杆菌属(厚壁菌门)为植物凋落物分解过程的主要微生物, 且随着凋落物分解过程的进行, 降解难降解有机物的特定微生物黄杆菌属(拟杆菌门)含量增加[28—29]。

|
| 图 6 植物凋落物分解过程沉积物微生物群落相对丰度分布 Fig. 6 Distribution of relative abundance of microbial community in sediments during decomposition of plant litters 门水平(相对丰度>1%) Phylum level (relative abundance > 1%);属水平(前13的菌种) Genus level (top 13 strains);CK:初始沉积物作为对照Control (Initial sediment);Cd-4、Cd-15、Cd-30:金鱼藻分别分解4、15、30 d时的沉积物Sediment after Ceratophyllum demersum L. decomposition for 4, 15, 30 d, respectively;Pc-4、Pc-15、Pc-30:菹草分别分解4、15、30 d时的沉积物Sediment after Potamogeton crispus L. decomposition for 4, 15, 30 d, respectively |
沉积物中微生物与DOM荧光组分的RDA(图 7)揭示了两种植物凋落物分解过程中微生物群落与溶解性有机物的关系。前两个约束轴RDA1和RDA2最大程度解释了DOM荧光组分对微生物群落变化的影响, 贡献了微生物群落总体变化的66.35%, 其中RDA1贡献60.07%, 是决定微生物群落变化的主要因素;C3是主要影响因子, 与RDA1的相关性系数达到0.81, 表明分子量大、腐殖化程度较高的有机物与微生物群落变化关系密切, 所以类腐殖酸类有机物显著影响凋落物分解过程中沉积物微生物群落的变化。相比于分解初期(4 d)凋落物释放以易降解有机物(类色氨酸)为主, 分解缓慢期(15 d)和分解相对稳定期(30 d), 难降解有机物增加导致了微生物活性降低, 进而使植物凋落物分解缓慢, 营养物质释放量降低(图 1)。

|
| 图 7 植物凋落物分解过程沉积物微生物与DOM荧光组分的RDA Fig. 7 RDA of sediment microorganisms and DOM fluorescence components during decomposition of plant litters |
冷暖季沉水植物凋落物具有不同的化学组成和结构特征, 导致其分解过程具有差异性, 对水质的影响也具有差异性(图 1)。如表 4所示, 两种沉水植物凋落物分解的化学物质最大释放量不同。1 g(干重)凋落物分解对上覆水有机物浓度的影响为菹草显著大于金鱼藻;对上覆水N、P浓度的影响为金鱼藻大于菹草, 这可能与凋落物C、N、P含量、C/N值有关(表 1)。Balasubramanian等[4]和曹培培等[30]认为, 初始N含量和C/N值在植物凋落物分解过程中起关键作用。本研究发现, 冷暖季沉水植物凋落物分解过程中, 水体有机物、N和P浓度的变化与植物自身化学组成和结构特征相关, 其中N含量更高、C/N值更低的菹草凋落物分解更快, 凋落物损失量更大, 对水体水质的影响程度更大(图 1和图 2), 与蔡润发[28]对鄱阳湖湿地水生植物分解对水质的影响研究结果一致。
| 植物凋落物 Plant litters |
碳释放量 Carbon release/(mg/g) |
氮释放量 Nitrogen release/(mg/g) |
磷释放量 Phosphorus release/(mg/g) |
| 金鱼藻Ceratophyllum demersum L. | 37.74 | 3.08 | 1.24 |
| 菹草Potamogeton crispus L. | 55.94 | 2.90 | 1.06 |
EEMs-PARAFAC法将三维荧光光谱主要分为类蛋白物质和类腐殖质物质两大类, T1、T2两种荧光峰与类色氨酸有关, 通常由人为活动或者微生物活动产生[31];A、C两个荧光峰主要来源于植物分解和微生物代谢活动[32], 所代表的荧光物质分子量大、芳香化程度高, 是一类难降解的物质[33—34], 大量存在可导致水体的可生化性变差[35]。本研究上覆水中通过EEMs-PARAFAC法解析出的DOM荧光组分(类色氨酸)与Zhang等[5]对狐尾藻(Myriophyllum verticillatum)分解的研究结果(类络氨酸)不同, 说明不同种沉水植物凋落物分解释放的DOM浓度和组分存在差异。本研究发现凋落物分解释放的DOM呈现低腐殖化特征, 与洪志强等[36]对沉水植物分解的研究结果一致。胡晓明等[37]也发现白洋淀区域水中的DOM呈现较强的自生源、低腐殖化、低芳香化特征。相比于金鱼藻, 菹草凋落物分解释放的DOM浓度和难降解DOM组分均更大, 说明菹草凋落物腐败可能是白洋淀水体难降解DOM的一个重要内源。
3.2 冷暖季沉水植物凋落物分解与微生物的相互作用在水环境中, 早期植物凋落物分解的特点是快速浸析和微生物定殖[9]。0—15 d植物凋落物质量损失30%—40%, 约占总损失量(60%—70%)的一半以上, 营养物质(TN、TP和TOC)浓度显著增加(图 1和图 2)。0—15 d微生物对凋落物分解的影响较小[38], 但与原始沉积物中的微生物相比, 这些营养物质使沉积物中微生物快速生长(4 d和15 d), 同时降解植物凋落物的特定微生物群落逐渐增加, 如假单胞菌属、Malikia属和黄杆菌属(图 6)。15—60 d, 两种植物凋落物分解质量损失缓慢增加, 营养物质(TN、TP和TOC)缓慢释放(图 1和图 2), 说明微生物对植物凋落物中难降解有机物(如木质纤维素)的分解发挥了重要作用。沉积物中微生物与DOM荧光组分的RDA分析发现(图 7), 0—15 d物质释放以易降解有机物为主(图 3和图 5), 相对分子质量较小的有机物更适合微生物的生长, 而15—60 d物质释放以难降解有机物为主(图 3和图 5), 相对分子质量较大的难降解有机物影响微生物群落的变化[39]。与4 d和15 d相比, 30 d时两种植物凋落物体系中沉积物微生物丰富度增加、多样性降低(表 3), 可知凋落物分解过程中营养物质的释放促进了细菌群落的变化, 营养物质的利用导致了一系列特异性微生物成为优势微生物, 如Malikia属(变形菌门)、芽孢杆菌属(厚壁菌门)。研究表明, 变形菌门、厚壁菌门对有机污染物和N的去除均起主要作用[38, 40]。因此, 微生物对植物凋落物分解的贡献主要在后期。
3.3 冷暖季沉水植物管理措施及实践意义根据发育特征, 沉水植物可分为冷季型和暖季型。冷季型沉水植物秋季发芽, 冬春季生长, 5—6月逐渐衰亡腐败;暖季型沉水植物春季发芽, 夏秋季生长, 10—11月逐渐衰亡腐败。利用冷暖季沉水植物生理特性的显著差异, 通过冷暖季沉水植物的合理配置, 促进其生态位互补, 充分发挥不同发育期沉水植物的水质净化作用和生态效应。根据植物凋落物腐败分解的显著差异性, 有针对性地及时对冷暖季植物进行收割管理, 可以有效预防水生植物衰亡对湿地水质的负面影响。可以分别在冷季和暖季水生植物衰亡期前全面收割水生植物, 减少水生植物凋落物在衰亡过程中向水中释放污染物以及植物残体堆积可能造成的淤积问题[2]。同时, 在水生植物生长各个阶段, 也可对水生植物进行平衡收割, 促进植物的生长和减少植物凋落物的分解;适量植物凋落物的分解可能增加水体中的碳源, 提高系统中的C/N, 促进微生物群落的活动, 提高水质净化效果[41], 但这方面的研究仍需深入研究。
4 结论(1) 金鱼藻和菹草凋落物分解规律相似, 0—15 d快速分解, C、N和P释放量显著增加, 15—60 d缓慢分解导致释放量降低, 但也表现出各自的分解特性。菹草的有机物释放量明显高于金鱼藻, 而N和P释放量低于金鱼藻, 两种植物凋落物分解过程上覆水中的N主要以NH4+-N和有机氮的形式存在。
(2) EEMs-PARAFAC法解析出植物凋落物的分解产物包括一种类色氨酸物质C2, 3种类腐殖质物质C1、C3和C4, 易降解的类色氨酸有机物先增加后减少, 难降解的类富里酸和类腐殖酸有机物逐渐增加。EEMs和PARAFAC组分的最大荧光强度百分比表明, DOM在0—15 d以易降解有机物为主, 15—60 d以难降解有机物为主。金鱼藻和菹草凋落物分解过程释放的DOM含量及特性不同, 整体上呈低腐殖化特征, 可能是水中难降解DOM的一个重要内源。
(3) 沉积物中微生物群落随着分解过程中营养物质的动态变化发生显著改变。从门水平和属水平看, 参与植物凋落物分解的微生物含量增加, 如Proteobacteria门(Pseudomonas属和Malikia属)、Firmicutes门(Bacillus属)。同时, 降解难降解有机物类物质的微生物逐渐增加, 如Bacteroidota门(Flavobacterium属)。通过RDA发现类腐殖酸有机物可显著影响分解过程沉积物中微生物群落的变化。
(4) 金鱼藻和菹草凋落物分解对水质的影响具有阶段性, 在0—15 d向水体释放更多的N和P, 可暂时导致水体水质的恶化;15—60 d期间N和P释放量降低, 而难降解有机物含量逐渐增加, 可能会加剧水体甚至是沉积物的腐殖化程度, 尤其是菹草凋落物腐败分解对水质恶化的贡献较大。因此, 在植物衰亡期应及时打捞或者做好植物平衡收割管理, 避免因植物大量腐败导致水质恶化。
| [1] |
Li W T, Chen S Y, Xu Z X, Li Y, Shuang C D, Li A M. Characterization of dissolved organic matter in municipal wastewater using fluorescence PARAFAC analysis and chromatography multi-excitation/emission scan: a comparative study. Environmental Science & Technology, 2014, 48(5): 2603-2609. |
| [2] |
刘瑜. 藻苲淀近自然湿地冷暖季水生植物水质净化作用研究[D]. 北京: 北京林业大学, 2019.
|
| [3] |
Søndergaard M, Jeppesen E, Lauridsen T L, Skov C, van Nes E H, Roijackers R, Lammens E, Portielje R. Lake restoration: successes, failures and long-term effects. Journal of Applied Ecology, 2007, 44(6): 1095-1105. DOI:10.1111/j.1365-2664.2007.01363.x |
| [4] |
Balasubramanian D, Arunachalam K, Das A K, Arunachalam A. Decomposition and nutrient release of Eichhornia crassipes (Mart.) Solms. under different trophic conditions in wetlands of eastern Himalayan foothills. Ecological Engineering, 2012, 44: 111-122. DOI:10.1016/j.ecoleng.2012.03.002 |
| [5] |
Zhang L S, Zhang S H, Lv X Y, Qiu Z, Zhang Z Q, Yan L Y. Dissolved organic matter release in overlying water and bacterial community shifts in biofilm during the decomposition of Myriophyllum verticillatum. Science of the Total Environment, 2018, 633: 929-937. DOI:10.1016/j.scitotenv.2018.03.275 |
| [6] |
Yuan D H, Zhao Y X, Guo X J, Zhai L X, Wang X Y, Wang J Z, Cui Y Q, He L S, Yan C L, Kou Y Y. Impact of hydrophyte decomposition on the changes and characteristics of dissolved organic matter in lake water. Ecological Indicators, 2020, 116: 106482. DOI:10.1016/j.ecolind.2020.106482 |
| [7] |
Wu S Q, He S B, Zhou W L, Gu J Y, Huang J, Gao L, Zhang X. Decomposition characteristics of three different kinds of aquatic macrophytes and their potential application as carbon resource in constructed wetland. Environmental Pollution, 2017, 231: 1122-1133. DOI:10.1016/j.envpol.2017.07.049 |
| [8] |
Li C H, Wang B, Ye C, Ba Y X. The release of nitrogen and phosphorus during the decomposition process of submerged macrophyte (Hydrilla verticillate Royle) with different biomass levels. Ecological Engineering, 2014, 70: 268-274. DOI:10.1016/j.ecoleng.2014.04.011 |
| [9] |
Abril M, Muñoz I, Menéndez M. Heterogeneity in leaf litter decomposition in a temporary Mediterranean stream during flow fragmentation. Science of the Total Environment, 2016, 553: 330-339. DOI:10.1016/j.scitotenv.2016.02.082 |
| [10] |
Corstanje R, Reddy K R, Portier K M. Typha latifolia and Cladium jamaicense litter decay in response to exogenous nutrient enrichment. Aquatic Botany, 2006, 84(1): 70-78. DOI:10.1016/j.aquabot.2005.07.013 |
| [11] |
Leff J W, Nemergut D R, Grandy A S, O'Neill S P, Wickings K, Townsend A R, Cleveland C C. The effects of soil bacterial community structure on decomposition in a tropical rain forest. Ecosystems, 2012, 15(2): 284-298. DOI:10.1007/s10021-011-9510-2 |
| [12] |
Zhao B Y, Xing P, Wu Q L. Microbes participated in macrophyte leaf litters decomposition in freshwater habitat. FEMS Microbiology Ecology, 2017, 93(10): fix108. |
| [13] |
任俊龙. 白洋淀菹草生长特性、氮磷积累量和生态服务功能的研究[D]. 保定: 河北大学, 2012.
|
| [14] |
Chimney M J, Pietro K C. Decomposition of macrophyte litter in a subtropical constructed wetland in south Florida (USA). Ecological Engineering, 2006, 27(4): 301-321. DOI:10.1016/j.ecoleng.2006.05.016 |
| [15] |
Zhang Y L, Yin Y, Liu X H, Shi Z Q, Feng L Q, Liu M L, Zhu G W, Gong Z J, Qin B Q. Spatial-seasonal dynamics of chromophoric dissolved organic matter in Lake Taihu, a large eutrophic, shallow lake in China. Organic Geochemistry, 2011, 42(5): 510-519. DOI:10.1016/j.orggeochem.2011.03.007 |
| [16] |
Rogers M B, Firek B, Shi M, Yeh A, Brower-Sinning R, Aveson V, Kohl B L, Fabio A, Carcillo J A, Morowitz M J. Disruption of the microbiota across multiple body sites in critically ill children. Microbiome, 2016, 4(1): 66. DOI:10.1186/s40168-016-0211-0 |
| [17] |
Kernan M R, Helliwell R C. Partitioning the variation within the acid neutralizing capacity of surface waters in Scotland in relation to land cover, soil and atmospheric depositional factors. Science of the Total Environment, 2001, 265(1/2/3): 39-49. |
| [18] |
Lemley D A, Snow G C, Human L. The decomposition of estuarine macrophytes under different temperature regimes. Water SA, 2014, 40(1): 117. DOI:10.4314/wsa.v40i1.14 |
| [19] |
Liu S S, Zhu Y R, Meng W, He Z Q, Feng W Y, Zhang C, Giesy J P. Characteristics and degradation of carbon and phosphorus from aquatic macrophytes in lakes: insights from solid-state 13C NMR and solution 31P NMR spectroscopy. Science of the Total Environment, 2016, 543: 746-756. DOI:10.1016/j.scitotenv.2015.11.080 |
| [20] |
Stedmon C A, Markager S. Resolving the variability in dissolved organic matter fluorescence in a temperate estuary and its catchment using PARAFAC analysis. Limnology and Oceanography, 2005, 50(2): 686-697. DOI:10.4319/lo.2005.50.2.0686 |
| [21] |
Yamashita Y, Panton A, Mahaffey C, Jaffé R. Assessing the spatial and temporal variability of dissolved organic matter in Liverpool Bay using excitation-emission matrix fluorescence and parallel factor analysis. Ocean Dynamics, 2011, 61(5): 569-579. DOI:10.1007/s10236-010-0365-4 |
| [22] |
Coble P G. Characterization of marine and terrestrial DOM in seawater using excitation-emission matrix spectroscopy. Marine Chemistry, 1996, 51(4): 325-346. DOI:10.1016/0304-4203(95)00062-3 |
| [23] |
Murphy K R, Stedmon C A, Waite T D, Ruiz G M. Distinguishing between terrestrial and autochthonous organic matter sources in marine environments using fluorescence spectroscopy. Marine Chemistry, 2008, 108(1/2): 40-58. |
| [24] |
Guo W D, Xu J, Wang J P, Wen Y R, Zhuo J F, Yan Y C. Characterization of dissolved organic matter in urban sewage using excitation emission matrix fluorescence spectroscopy and parallel factor analysis. Journal of Environmental Sciences, 2010, 22(11): 1728-1734. DOI:10.1016/S1001-0742(09)60312-0 |
| [25] |
Baker A, Curry M. Fluorescence of leachates from three contrasting landfills. Water Research, 2004, 38(10): 2605-2613. DOI:10.1016/j.watres.2004.02.027 |
| [26] |
Stedmon C A, Markager S, Bro R. Tracing dissolved organic matter in aquatic environments using a new approach to fluorescence spectroscopy. Marine Chemistry, 2003, 82(3/4): 239-254. |
| [27] |
刘新, 刘浩, 江和龙, 宋娜. 不同水生植物腐解过程中有色可溶有机物(CDOM)的产生过程及微生物群落变化分析. 长江流域资源与环境, 2020, 29(5): 1140-1149. |
| [28] |
蔡润发. 鄱阳湖湿地3种植物凋落物的分解过程及细菌群落结构动态变化[D]. 南昌: 南昌大学, 2019.
|
| [29] |
Oliveira C S, Alcantara G B, Lião L M, Mesquita G M, Freitas S S, Petacci F. Decomposition dynamics of Typha angustifolia under aerobic conditions. Journal of the Brazilian Chemical Society, 2016, 27(9): 1687-1693. |
| [30] |
曹培培, 刘茂松, 唐金艳, 滕漱清, 徐驰. 几种水生植物腐解过程的比较研究. 生态学报, 2014, 34(14): 3848-3858. |
| [31] |
Yu H B, Song Y H, Tu X, Du E D, Liu R X, Peng J F. Assessing removal efficiency of dissolved organic matter in wastewater treatment using fluorescence excitation emission matrices with parallel factor analysis and second derivative synchronous fluorescence. Bioresource Technology, 2013, 144: 595-601. DOI:10.1016/j.biortech.2013.07.025 |
| [32] |
王迪, 张飞, 张兆永, 张海威, 朱世丹, 张贤龙. 新疆艾比湖流域枯、丰水期三维荧光光谱特性及其与水质的关系. 湖泊科学, 2020, 32(2): 483-495. |
| [33] |
Lee S, Hur J. Heterogeneous adsorption behavior of landfill leachate on granular activated carbon revealed by fluorescence excitation emission matrix (EEM)-parallel factor analysis (PARAFAC). Chemosphere, 2016, 149: 41-48. DOI:10.1016/j.chemosphere.2016.01.081 |
| [34] |
贾陈忠, 王焰新, 张彩香, 秦巧燕. 垃圾渗滤液中溶解性有机物组分的三维荧光特性. 光谱学与光谱分析, 2012, 32(6): 1575-1579. DOI:10.3964/j.issn.1000-0593(2012)06-1575-05 |
| [35] |
杜伊, 胡玮璇, 王晓燕, 陈永娟, 刘洋. 北京市北运河水体中化学需氧量组分含量及其可生化性研究. 湿地科学, 2017, 15(3): 470-477. |
| [36] |
洪志强, 熊瑛, 李艳, 崔骏, 刘钰钦, 何江伟, 王京刚, 袁冬海. 白洋淀沉水植物腐解释放溶解性有机物光谱特性. 生态学报, 2016, 36(19): 6308-6317. |
| [37] |
胡晓明, 崔骏, 裴元生. 秋冬交替季节白洋淀沉积物有机质特性研究. 环境科学研究, 2019, 32(4): 636-646. |
| [38] |
Blanchet M, Pringault O, Panagiotopoulos C, Lefèvre D, Charrière B, Ghiglione J F, Fernandez C, Aparicio F L, Marrasé C, Catala P, Oriol L, Caparros J, Joux F. When riverine dissolved organic matter (DOM) meets labile DOM in coastal waters: changes in bacterial community activity and composition. Aquatic Sciences, 2017, 79(1): 27-43. |
| [39] |
王彤, 文刚, 黄廷林, 司凡, 王晨旭, 路林超, 林薇. 水库热分层期沉积物中有机物特性及对好氧反硝化细菌脱氮的影响. 环境科学, 2020, 41(11): 5027-5036. |
| [40] |
Tang V T, Rene E R, da Fang Fu, Singh R P, Behera S K, Pugazhendhi A. Effect of mixed microbial culture addition on enhanced river water quality: Pollutant removal and microbial community characteristics. Environmental Technology & Innovation, 2020, 18: 100707. |
| [41] |
Zhao D H, Chen C, Lu Q Q, Zhang M, Zhang H, An S Q. Combined use of cold-season and warm-season macrophytes in floating constructed wetlands to increase nitrogen removal in the early cold season. Journal of Cleaner Production, 2020, 254: 120054. |