 2022, Vol. 42
2022, Vol. 42文章信息
- 高苏日固嘎, 斯琴朝克图, 乌兰图雅, 赛西雅拉图, 张卫青, 美丽
- GAO Suriguga, SIQIN Chaoketu, WULAN Tuya, SAI Xiyalatu, ZHANG Weiqing, MEI Li
- 克氏针茅草原群落物种多样性与生物量关系对放牧强度的响应
- Relationship between species diversity and biomass of Stipa krylovii steppe community and its responses to grazing intensity
- 生态学报. 2022, 42(23): 9736-9746
- Acta Ecologica Sinica. 2022, 42(23): 9736-9746
- http://dx.doi.org/10.5846/stxb202110162921
-
文章历史
- 收稿日期: 2021-10-16
- 网络出版日期: 2022-07-27





2. 内蒙古自治区蒙古高原灾害与生态安全重点实验室, 呼和浩特 010020;
3. 蒙古高原气候变化与区域响应自治区高等学校重点实验室, 呼和浩特 010020
2. Inner Mongolia Key Laboratory of Disaster and Ecological Security on the Mongolian Plateau, Hohhot 010020, China;
3. Provincial Key Laboratory of Mongolian Plateau′s Climate System, Inner Mongolia Normal University, Hohhot 010020, China
克氏针茅草原是内蒙古典型草原的主要组成部分, 分布于锡林郭勒草原的中部和西部, 面积约为1639.10万hm2, 约占18.35%[1]。克氏针茅草原生态环境类型独特具有草原生物群落的基本特征, 并能全面反映内蒙古高原典型草原生态系统的结构和生态过程[2]。目前, 锡林郭勒克氏针茅草原形成不同程度的退化, 不合理的放牧方式是影响克氏针茅草原退化的主要原因之一[3], 长期的超载过牧引起的草地生态系统受损问题越来越受到国内外广大学者门的关注。为此, 如何确定合理的放牧强度, 恢复退化的牧场, 维持草地生态系统稳定, 兼顾草原畜牧业发展, 是当前克氏针茅草原所面临的现实问题。
放牧是锡林郭勒克氏针茅草原最主要的利用形式, 在过去很长一段时间里, 过度放牧使锡林郭勒克氏针茅草原遭到了严重的破坏, 牧草平均年产量下降, 物种多样性减少[4—5]。2002年内蒙古政府实施“退牧还草”政策以来, 对锡林郭勒克氏针茅草原采取围栏封育、禁牧、休牧等措施, 部分退化的草原得到了一定的修复[6]。目前, 国内外诸多学者对放牧方式与草地植被关系进行了大量研究。一些研究认为, 牲畜的选择性采食、践踏和排泄, 直接或间接地影响某些植物的种群结构, 甚至影响整个草地生态系统[7—8];部分研究认为, 合理的放牧强度可以促进草地植物生长, 提高植物群落的物种多样性, 使草地群落达到相对稳定平衡, 保证草地生态系统的可持续发展[9—10];另有研究认为, 不合理的放牧对草地生态系统产生负面影响, 一种是禁牧, 禁牧能增加优良牧草在群落中的优势度, 促进草地植被恢复, 但长期禁牧也可能造成单个或少数优势种在群落的优势地位过度增加, 降低草地物种丰富度和多样性[11—12];另一种是过度放牧, 使草地种群生境恶化, 原有群落主要植物种功能群优势地位发生明显的替代变化, 植物群落高度和地上生物量显著下降, 群落物种多样性降低, 结构简单化[13—14]。但这些研究主要集中在高寒草甸、高山草地、草甸草原以及荒漠草原, 在典型草原区域的研究相对较少, 尤其在克氏针茅草原植被特征如何响应不同放牧强度的相关研究有待深入。
综上所述, 放牧强度对克氏针茅草原植物群落物种多样性和地上生物量关系的研究尤为必要。本研究以典型草原优势种克氏针茅为实验样地, 通过设置禁牧、轻度放牧、中度放牧、重度放牧4种方式的草场进行采样。拟解决以下问题:①分析不同放牧强度对克氏针茅草原植物群落结构特征的影响;②分析不同放牧强度对克氏针茅草原植物群落多样性的影响及其关系;③探究不同放牧强度下植物群落物种多样性及地上生物量的相关性。为此, 确定合理的放牧强度, 实现克氏针茅草原可持续利用, 为草原生态系统的恢复与稳定提供科学参考。
1 材料与方法 1.1 研究区概况研究区位于内蒙古锡林郭勒盟阿巴嘎旗那仁宝拉格苏木阿拉坦陶高图嘎查(44°36′—44°54′N, 114°55′—114°14′E)(图 1), 地处锡林郭勒波状高平原北部, 地形以低山丘陵为主, 地势由东北向西南倾斜, 海拔1240—1483 m。气候为温带大陆性干旱半干旱气候, 年平均气温0.5℃, 年最大温差50℃, 年平均降水量220 mm, 年无霜期100—110 d, 降水集中于6—8月份。土壤类型为暗栗钙土和淡栗钙土。植物群落的优势种为克氏针茅(Stipa krylovii)、羊草(Leymus chinensis)等, 伴生种为冷蒿、糙隐子草、寸草苔等[15]。自1996年实行“畜草双承包”制度以来, 研究区放牧方式由游牧转向定牧, 牧户以网围栏定居放牧为主[16], 由于牲畜数量增加、畜群结构变化、草场管理不当等多重原因, 牧户草场发生不同程度退化。

|
| 图 1 研究区及采样点分布 Fig. 1 Distribution of study area and sampling points |
本研究选取放牧强度不同的3个牧户, 牧户草场的地形、土壤、植物组成以及养殖牲畜结构基本相同。自2003年实施禁牧政策以来, 各牧户的畜群数量和结构变化不大。放牧试验采用随机区组设计, 每年在植被生长季为6月上旬—9月中旬, 期间为最佳放牧时期, 放牧一般在每天7:00左右开始, 18:00左右结束。如表 1所示, 本文借鉴前人有关放牧强度研究[17—19], 并根据研究区植物群落、载畜量以及放牧方式等情况, 把放牧强度划分为4个等级, 分别为禁牧(NG, No grazing)、轻度放牧(LG, Light grazing)、中度放牧(MG, Middle grazing)、重度放牧(HG, Heavy grazing), 标准载畜量分别为0、0.34、0.69、1.78羊单位/hm2。各类草场分布如图 1所示, 自北向南依次为轻牧、禁牧、中牧和重牧草场。
| 样地类型 Sample type |
草场面积 Pasture area/hm2 |
海拔高度 Altitude/m |
样点数 Sample Points |
主要优势植物 Major dominant plants |
| NG | 310 | 1330 | 6 | 克氏针茅(Stipa krylovii)、羊草(Leymus chinensis)、糙隐子草(Cleistogenes squarrosa) |
| LG | 1258 | 1337 | 6 | 克氏针茅(Stipa krylovii)、羊草(Leymus chinensis)、寸草苔(Carex duriuscula) |
| MG | 835 | 1319 | 6 | 猪毛菜(Salsola collina)、羊草(Leymus chinensis)、多根葱(Allium polyrhizum) |
| HG | 510 | 1297 | 6 | 猪毛菜(Salsola collina)、糙隐子草(Cleistogenes squarrosa)、多根葱(Allium polyrhizum) |
| NG:禁牧 No grazing;LG:轻度放牧 Light grazing;MG:中度放牧 Middle grazing;HG:重度放牧 Heavy grazing | ||||
2019年8月初我们在研究区进行植物群落调查, 以牧户草场为采样区, 根据4类不同放牧强度, 每类样区随机选取6个采样点, 为1 m × 1 m的植被样方, 邻近两个样方距离为200 m, 共计24个采样点。测定所有样方的植物高度、密度、盖度、地上生物量以及植物种类组成。其中高度是用卷尺测量从植株底部到植物冠层的自然高度, 每种植物测量3—5株, 并取平均高度;密度是样方内所有植物的株数;盖度用相机垂直于样方拍摄, 运用Photoshop 6软件, 根据植物与土壤的颜色差异得出样方内植物的像素, 植物像素除以样方总像素即为样方的盖度值;地上生物量是对每个样方采用收割法, 分物种获取每种植物的鲜重, 并带回实验室, 在65℃下烘干至恒重并测量其干重[20]。
1.3 数据处理与方法本研究按物种生活型差异, 将植物群落中出现的物种划分为4种功能群[21—22]:多年生禾草、多年生杂类草、一、二年生草本和灌木、半灌木。本研究使用a多样性指标来表示植物群落多样性[23], 即Shannon-Wiener指数(多样性指数, H′)、Margalef指数(丰富度指数, MA)、Pielou指数(均匀度指数, E)和Simpson指数(优势度指数, P)。植物群落中物种的优势度由重要值(Pi)确定[24]。计算公式如下:
Shannon-Wiener指数:
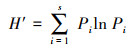
|
(1) |
Margalef指数:
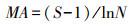
|
(2) |
Pielou指数:
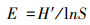
|
(3) |
Simpson指数:
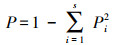
|
(4) |
重要值:

|
(5) |
式中, S表示植物群落中物种数目;N表示所有物种个体数总和;Pi表示物种重要值。运用Excel软件预处理原数据, 利用Origin 9.1作图。运用SPSS 22.0软件对不同放牧强度下植物群落特征和地上生物量进行单因素方差分析(One-way ANVOA)和差异显著性检验(α=0.05);不同放牧强度对物种多样性和地上生物量的关系采用简单线性回归分析。
2 结果与分析 2.1 放牧强度对植物群落特征的影响 2.1.1 植物群落物种组成与结构草原植物群落组成决定群落结构、外貌、功能以及群落的演替方向, 是植被最直观的特征之一, 放牧强度可以改变植物群落组成和结构特征, 同样植物群落特征的变化也可以反映草地对不同放牧强度的响应[24]。如表 2所示, 放牧强度对植物群落组成具有一定影响, NG、LG、MG和HG区物种数分别为19个、24个、20个和16个。从物种的重要值来看, NG区的优势种为克氏针茅、羊草、糙隐子草, 上述物种重要值占比为46.83%;LG区的优势种为克氏针茅、羊草、寸草苔, 上述物种重要值占比为48.27%;MG区的优势种为猪毛菜、羊草、多根葱, 上述物种重要值占比为43.29%;HG区的优势种为猪毛菜、糙隐子草、多根葱, 上述物种重要值占比为58.09%。由此可见, NG和LG区的优势种变化不大较为稳定, 而MG和HG区的优势种变化较大, 尤其是猪毛菜、多根葱、寸草苔等物种的出现表明克氏针茅草原退化严重。
| 功能群 Functional groups |
植物名称 Species name |
植物重要值 Important value of vegetation/% |
|||
| NG | LG | MG | HG | ||
| 多年生禾草 | 冰草 Agropyron cristatum | — | 0.33 | 2.27 | — |
| Perennial grasses | 羊草 Leymus chinensis | 12.45 | 15.58 | 12.99 | 6.42 |
| 糙隐子草 Cleistogenes squarrosa | 12.19 | 8.83 | 3.23 | 15.07 | |
| 克氏针茅 Stipa krylovii | 22.19 | 21.37 | 3.73 | 7.85 | |
| 多年生杂类草 | 柴胡 Bupleurum chinense | 0.62 | 0.88 | — | — |
| Perennial forbs | 寸草苔 Carex duriuscula | 3.41 | 11.32 | 2.17 | 3.89 |
| 多根葱 Allium Polyrhizum | 1.86 | 1.91 | 12.05 | 13.55 | |
| 二裂委陵菜 Potentilla bifurca | 1.03 | — | — | — | |
| 山韭 Allium senescens | — | — | 4.78 | — | |
| 细叶葱 Allium tenuissimum | — | 1.33 | 5.85 | 2.78 | |
| 阿尔泰狗娃花 Heteropappus altaicus | 2.66 | 0.43 | 0.61 | — | |
| 阿氏旋花 Convolvulus ammannii | 0.95 | 0.43 | 3.88 | 5.79 | |
| 北芸香 Haplophyllum dauricum | 1.47 | 0.66 | — | 0.77 | |
| 草地麻花头 Serratula komarovii | — | 1.39 | — | — | |
| 毛棘豆 Oxytropis pagobia | 3.28 | 3.63 | 3.26 | 2.83 | |
| 沙葱 Allium mongolicum regel | 5.53 | — | — | — | |
| 风毛菊 Saussurea aponica | — | 0.45 | — | — | |
| 星毛委陵菜 Potentilla acaulis | — | 1.21 | 1.40 | — | |
| 一、二年生草本 | 猪毛菜 Salsola collina | 8.67 | 2.64 | 18.25 | 29.47 |
| Annuals and biennials | 刺藜 Chenopodium aristatum | 0.10 | 1.26 | 1.14 | 1.43 |
| 钝叶瓦松 Orostachys malacophylla | 2.58 | — | — | — | |
| 黄花蒿 Artemisia annua | — | 3.61 | 4.97 | — | |
| 灰绿藜 Chenopodium glaucum | — | 3.39 | — | 0.68 | |
| 灌木、半灌木 | 燥原荠 Ptilotricum canescens | 3.56 | 1.89 | 3.62 | — |
| Shrubs and semi-shrubs | 冷蒿 Artemisia frigida | 9.73 | 5.39 | 8.33 | 2.49 |
| 栉叶蒿 Neopallasia pectinata | — | 0.48 | 3.03 | — | |
| 狭叶锦鸡儿 Caragana stenophylla | 6.98 | 9.45 | 3.33 | 1.77 | |
| 独行菜 Lepidium apetalum | — | 0.33 | — | 2.39 | |
| 东北丝裂蒿 Artemisia adamsii bess | 0.74 | 1.81 | 1.11 | 2.82 | |
| 物种总数 Total number of species | 19 | 24 | 20 | 16 | |
| “—”表示植物群落中该物种没有出现 | |||||
如图 2所示, 从物种的功能群来看, NG和LG区以多年生禾草和多年生杂草为主, MG和HG区中一、二年生植物居多。其中, NG、LG、MG、HG区多年生禾草占比均值分别为46.83%、46.20%、29.34%、20.69%;多年生杂草占比均值分别为22.23%、19.81%、30.12%、35.73%;一、二年生植物占比均值分别为23.27%、22.40%、34.90%、43.31%;灌木、半灌木占比均值分别为7.71%、11.59%、5.63%、4.43%。此外, NG、LG区草原群落不同功能群的类群占比排序为:多年生禾草>一、二年生草本植物>多年生杂草>灌木、半灌木;MG、HG区排序为:一、二年生草本植物>多年生杂草>多年生禾草>灌木、半灌木。由此可见, 随着放牧强度的增加, 植物群落结构逐步向退化方向演替。

|
| 图 2 不同放牧强度各采样点植物生活型重要值百分比 Fig. 2 Percentage of important values of plant life forms at each sampling point with different grazing intensities |
群落特征是生态恢复和重建的基础, 更是植被对生态系统适应的综合体现[25]。放牧是影响克氏针茅草原植物群落高度、密度、盖度、地上生物量的主要原因之一, 不同放牧强度对植物群落数量特征产生一定的影响[26]。如图 3所示, NG、LG、MG、HG区植物群落平均高度分别为9.31、8.41、6.55、5.19 cm, 其中NG、LG、MG区之间无显著差异, 而与HG区具有显著差异(P < 0.05), 表明随着放牧强度增加群落高度逐渐降低, 尤其在HG条件下群落高度显著降低;如图 3所示, NG、LG、MG、HG区植物群落密度分别为11.99、16.96、26.43、39.71株/m2, 其中NG、LG区与MG区无显著差异, 而与HG区具有显著差异(P < 0.05), 表明随着放牧强度增加群落密度逐渐增加, 尤其在HG条件下群落密度显著增加;如图 3所示, NG、LG、MG、HG区群落盖度分别为62.83%、72.83%、54.66%、53.66%, 其中NG与MG区之间无显著差异, LG与HG区具有显著差异(P < 0.05), 表明随着放牧强度增加群落盖度总体呈下降趋势。

|
| 图 3 各样区植物群落数量特征 Fig. 3 Quantitative characteristics of plant communities in each plot 每个盒子的小立方体代表平均值; 不同小写字母表示不同放牧强度之间差异显著(P < 0.05) |
植物群落地上生物量是反映草原生态系统和放牧系统稳定性的一个重要指标[9], 放牧强度直接影响植物群落地上生物量。如图 4所示, NG、LG、MG、HG区群落总地上生物量分别为193.58、218.65、122.36、115.33 g/m2, 其中NG、LG区与MG、HG区具有显著差异(P < 0.05), 表明随着放牧强度增加, 地上生物量总体呈下降趋势, MG和HG区显著低于LG区;研究区原有群落优势种主要包括克氏针茅和羊草, 如图 4所示, NG、LG、MG、HG区原有群落优势种地上生物量分别为80.65、92.62、38.2、26.7 g/m2, 其中NG、LG区与MG、HG区具有显著差异(P < 0.05), 表明随着放牧强度增加, 原有群落优势种地上生物量总体呈下降趋势, MG和HG区显著低于LG区;研究区退化指示物种主要包括冷蒿、星毛委陵菜、二裂委陵菜、糙隐子草、阿尔泰狗娃花、多根葱、阿氏旋花、寸草苔等[27], 如图 4所示, NG、LG、MG、HG区退化指示物种地上生物量分别为42.6、58.41、80.28、193.13 g/m2, NG、LG、MG区与HG区具有显著差异(P < 0.05), 表明随着放牧强度增加, 退化指示物种的地上生物量总体呈增加趋势, 尤其在HG区显著增加(P < 0.05)。

|
| 图 4 各样区地上生物量分布 Fig. 4 Aboveground biomass distribution in each sample area 每个盒子的小立方体代表平均值; 不同小写字母表示不同放牧强度之间差异显著(P < 0.05) |
植物群落的Shannon Wiener指数、Margalef指数、Pielou指数和Simpson指数4个群落a多样性指数表述群落多样性变化特征[28]。如表 3所示, 随着放牧强度增加, 植物群落的Shannon Wiener指数、Margalef指数、Pielou指数和Simpson指数均呈现先增后减的趋势。MG区的Shannon Wiener指数、Pielou指数和Simpson指数最高, 分别为2.25、0.93、0.88, LG区的Margalef指数最高, 为2.03。其中, Shannon Wiener指数在不同放牧强度下无显著差异;Margalef指数在LG区显著高于HG区(P < 0.05);Pielou指数在LG、MG区显著高于NG、HG区(P < 0.05);Simpson指数在MG区显著高于NG、HG区(P < 0.05)。
| 放牧强度 Grazing intensity |
多样性指数 Shannon-Wiener index |
丰富度指数 Margalef index |
均匀度指数 Pielou index |
优势度指数 Simpson index |
| NG | 2.13±0.09a | 1.87±0.10ab | 0.87±0.01b | 0.84±0.01b |
| LG | 2.06±0.05a | 2.03±0.10a | 0.92±0.02a | 0.85±0.01ab |
| MG | 2.25±0.11a | 1.88±0.09ab | 0.93±0.02a | 0.88±0.02a |
| HG | 2.14±0.04a | 1.70±0.12b | 0.86±0.04b | 0.83±0.02b |
| 不同小写字母表示不同放牧强度之间差异显著(P < 0.05) | ||||
植物群落物种多样性与地上生物量及其相互关系是生态系统发生与发展的基础[29], 不同放牧强度下, 植物群落物种多样性和地上生物量之间的关系存在差异。如图 5所示, 在所有样区地上生物量与Shannon-Wiener指数均呈显著正相关关系(P < 0.05);在MG、HG区地上生物量与Margalef指数均呈显著正相关关系(P < 0.05), 而在NG、LG区无相关关系;在NG、LG和MG区地上生物量与Pielou指数均呈显著正相关关系(P < 0.05), 而在HG区无相关关系;在所有样区地上生物量与Simpson指数均呈显著负相关关系(P < 0.05)。说明克氏针茅草原物种多样性与生物量的关系比较密切, 不同的放牧强度影响地上生物量与物种多样性的关系。

|
| 图 5 各样区地上生物量与物种多样性关系 Fig. 5 The relationship between aboveground biomass and species diversity in each sample area |
群落结构是草原植物群落是否健康的重要指标, 尤其是优势种的变化, 对草原生态系统演替的方向起着重要的作用[30]。不同放牧强度改变草地群落组成和结构特征。克氏针茅草原是以克氏针茅、羊草为优势种的典型草原, 包括多种牲畜喜食的、营养价值高的多年生禾草。本研究结果, 在轻度放牧条件下, 适度放牧促进多年生禾草再生长, 使其重要值在植物群落中增加[14];而中度和重度放牧条件下, 牲畜过度采食多年生禾草, 使其重要值在植物群落中减少, 这与徐粒等[31]的研究结果一致。另外, 中度和重度放牧条件下, 一、二年生草本和灌木、半灌木等植物的重要值增加, 牲畜过量采食使得适口性较好的多年生禾草的枝叶减少, 有效光合面积减少, 导致多年生禾草被多年生杂草、一、二年生草本及灌木取代, 植物群落发生退化演替, 这与张小红等[32]的研究结果一致。此外, 刘佳慧等[33]的研究表明, 随着放牧强度增加, 牲畜过量采食多年生禾草以外, 过度踩踏也可导致牧草的数量和种类减少, 使多年生禾草重要值减少。
3.2 放牧强度与植物群落数量放牧强度的大小影响草地植被的利用程度, 引起植物群落数量特征变化[27]。本研究结果, 与禁牧、轻度放牧相比, 中度和重度放牧条件下植物群落的盖度、高度和地上生物量显著降低, 这与孙宗玖等[34]的研究结果相一致。轻度放牧条件下植物群落的盖度最高, 高度次之、低于禁牧, 因为牲畜喜食植株顶端的鲜嫩枝叶, 群落高度受到影响, 而且植物的避牧生长策略, 使植物横向生长[35], 增加了植被盖度;重度条件下植物群落密度最高, 因为过度放牧造成牲畜喜食的多年生禾草大幅减少, 同时牲畜不喜食的一、二年生杂类草大幅增加, 使群落密度增高。另外, 本研究结果随着放牧强度增加, 植物群落总地上生物量、优势种生物量总体呈下降趋势(P < 0.05), 这与黄琛[36]的研究结果相同, 因为放牧使植物减少, 植物总光合面积减少, 造成植物有机物积累效率降低;随着放牧强度增加, 退化指示物种的地上生物量总体呈增加趋势(P < 0.05), 这与赵丽娅的研究结果相同[37], 中度和重度放牧条件下, 因为牲畜的选择性采食, 牲畜不喜食的退化指示植物如猪毛菜、灰绿藜等物种大量生长, 其地上生物量大幅增加。
3.3 放牧强度与植物群落物种多样性植物群落物种多样性是反映群落结构的可测定性指标, 与生态系统的功能密切相关, 不同的放牧强度使群落多样性发生变化[38]。放牧影响植物群落的物种多样性, 而且影响程度因放牧强度的大小而有所不同。其中适度放牧可增加物种多样性, 而过度放牧则降低物种多样性。多数研究认同“中度干扰假说”, 认为在中度放牧条件下草地多样性较高[39]。本研究结果, Simpson指数和Shannon-Wiener指数在中度放牧条件下最高, 而Margalef指数和Pielou指数在轻度放牧下最高。表明在轻度和中度放牧时物种多样性高于禁牧和重牧, 因为适度的干扰会抑制原有优势种的生长竞争, 为竞争力差的物种提供了机会, 使其能在适度干扰下更好生存, 致使此环境下植物群落保持了较高的多样性[40]。因此, 适度放牧可以使草地生态系统功能持续维持, 而过度放牧可引起草地植被退化, 所以要合理的、适度的把握放牧强度。
3.4 放牧强度与植物群落生物量和物种多样性的关系研究植物群落地上生物量和物种多样性的关系, 对于正确理解草地生态系统结构和功能具有重要的意义。赵生龙等[41]研究认为, 植物群落物种多样性和地上生物量的关系因能量、资源和环境因素存在较大的时空差异而表现为多种形式, 如线性正相关[42]、线性负相关[43]、单峰形[44]和不相关等[45]。本研究结果显示, 在不同放牧强度下植物群落地上生物量与Margalef指数、Shannon-Wiener指数均呈显著正线性关系(P < 0.05), 这与刘万弟等[46]的研究结果相同。因为同一植物群落中物种间存在着生态位差异, 有较多物种数的植物群落所占据的“功能空间”范围更广, 物种多样性较大的生态系统可以更有效地利用各种资源, 获得更高的生产力[47]。Pielou指数在禁牧、轻度、中度放牧条件下与地上生物量呈显著正相关, 而在重度放牧条件下与地上生物量无相关关系, 因为植物群落的空间异质性随放牧强度增加而增加。生存环境受到胁迫时, 植物群落中竞争能力弱的植物逐渐减少或者消失, 群落中的物种向退化方向演替, 多样性指数降低[18]。本研究结果中植物群落物种多样性与地上生物量的关系未呈现单峰形式, 因为该4种放牧强度下植物群落所构成的环境范围还未达到单峰形式的下降部分, 或者该模式就是克氏针茅草原在不同放牧强度下的植物群落物种多样性与地上生物量关系的基本形式。有关不同放牧强度下物种多样性与地上生物量的相互关系及其影响机制有待进一步深入研究。
4 结论(1) 放牧强度影响植物群落组成和功能, 随着放牧强度的增加植物群落结构逐步向退化方向演替, 其中多年生禾草的重要值逐渐减少, 多年生杂草和一、二年生草本植物的重要值逐渐增加。
(2) 随着放牧强度增加植物群落高度逐渐降低(P < 0.05), 尤其在重度放牧条件下群落高度显著降低;群落密度逐渐增加(P < 0.05), 尤其在重度放牧条件下群落密度显著增加;群落盖度总体呈下降趋势(P < 0.05)。
(3) 随着放牧强度增加, 植物群落和原有群落优势种地上生物量总体呈下降趋势(P < 0.05), 其中轻度放牧条件下最大, 重度放牧条件下最小;而退化指示物种的地上生物量逐渐增加(P < 0.05), 尤其在重度放牧条件下最大。
(4) 随着放牧强度增加, 植物群落的Shannon Wiener指数、Margalef指数、Pielou指数和Simpson指数均呈现先增加后减少的趋势, 其中, 轻度、中度放牧下群落物种Margalef指数、Pielou指数、Simpson指数均显著高于重度放牧(P < 0.05), 表明适度放牧可促进草原物种多样性和地上生物量。
(5) 在所有放牧强度下, 地上生物量与Shannon-Wienner指数、Margalef指数和Pielou指数呈正相关(P < 0.05), 而与Simpson指数呈负相关(P < 0.05)。克氏针茅草原物种多样性与生物量的关系比较密切, 不同的放牧强度影响地上生物量与物种多样性的关系。
| [1] |
候伟峰, 乌恩, 敖特根, 王成杰, 金净. 放牧强度对克氏针茅典型草原地上生物量的影响. 中国草地学报, 2016, 38(6): 71-77. |
| [2] |
郑阳. 内蒙古克氏针茅典型草原生态系统放牧管理优化模式研究[D]. 兰州: 甘肃农业大学, 2010.
|
| [3] |
邵蕊, 刘颖慧, 李悦, 牛磊, 武山梅. 施氮对典型草原克氏针茅光合特性的影响. 北京师范大学学报: 自然科学版, 2016, 52(5): 651-657. |
| [4] |
马梅, 张圣微, 魏宝成. 锡林郭勒草原近30年草地退化的变化特征及其驱动因素分析. 中国草地学报, 2017, 39(4): 86-93. |
| [5] |
徐松鹤, 陈宇浩. 休牧对锡林郭勒典型草原土壤养分和植物群落特征的影响. 水土保持研究, 2018, 25(4): 15-21. |
| [6] |
杨晨晨, 陈宽, 周延林, 潮洛濛, 呼格吉勒图, 陈瑜. 放牧对锡林郭勒草甸草原群落特征及生产力的影响. 中国草地学报, 2021, 43(5): 58-66. |
| [7] |
高露, 张圣微, 朱仲元, 徐冉, 张鹏. 放牧对干旱半干旱草原植物群落结构和生态功能的影响. 水土保持研究, 2019, 26(6): 205-211. |
| [8] |
郭彩贇, 赵东升, 郑度, 朱瑜. 放牧对内蒙古草原植被群落特征的影响. Journal of Resources and Ecology, 2021, 12(3): 319-331. |
| [9] |
张璐璐, 王孝安, 朱志红, 李英年. 模拟放牧强度与施肥对青藏高原高寒草甸群落特征和物种多样性的影响. 生态环境学报, 2018, 27(3): 406-415. |
| [10] |
杨晶晶, 吐尔逊娜依·热依木, 张青青, 阿马努拉·依明尼亚孜, 雪热提江·麦提努日. 放牧强度对天山北坡中段山地草甸植被群落特征的影响. 草业科学, 2019, 36(8): 1953-1961. |
| [11] |
丁海君, 韩国栋, 王忠武, 王春霞, 张睿洋, 胡吉亚. 短花针茅荒漠草原不同载畜率对植物群落特征的影响. 中国草地学报, 2014, 36(2): 55-60. DOI:10.3969/j.issn.1673-5021.2014.02.010 |
| [12] |
张娜, 秦艳, 金轲, 纪磊, 崔志强. 放牧对典型草原群落特征及土壤物理性状的影响. 中国草地学报, 2020, 42(4): 91-100. |
| [13] |
吴婷, 宋乃平, 陈晓莹, 李敏岚, 陈娟. 围栏封育和放牧对盐池荒漠草原植物群落特征的影响. 草地学报, 2019, 27(3): 651-660. |
| [14] |
毛绍娟, 吴启华, 李红琴, 张法伟, 李英年. 放牧强度对高寒杂草类草甸群落结构及生物量的影响. 冰川冻土, 2015, 37(5): 1372-1380. |
| [15] |
萨茹拉, 赛西雅拉图, 春风, 张卫青. 典型草原定居放牧草场群落特征与土壤有机质含量关系分析. 干旱区资源与环境, 2018, 32(9): 145-149. |
| [16] |
张旭冉, 张卫青, 王海茹, 卢晓霞, 春风, 赛西雅拉图. 克氏针茅草原土壤生态化学计量特征对放牧强度的响应. 生态学报, 2021, 41(13): 5309-5316. |
| [17] |
冯秀, 李元恒, 李芳, 丛日慧, 刘燕丹, 丁勇, 王育青. 典型草原合理放牧强度阈值研究. 中国草地学报, 2019, 41(5): 120-127. |
| [18] |
李颖, 龚吉蕊, 刘敏, 侯向阳, 丁勇, 杨波, 张子荷, 王彪, 朱趁趁. 不同放牧强度下内蒙古温带典型草原优势种植物防御策略. 植物生态学报, 2020, 44(6): 642-653. |
| [19] |
宋彦涛, 乌云娜, 张靖, 霍光伟, 张凤杰. 放牧强度对克氏针茅(Stipa krylovii)草原植被景观格局的影响. 中国沙漠, 2016, 36(3): 674-680. |
| [20] |
詹天宇, 孙建, 张振超, 刘某承. 基于meta分析的放牧压力对内蒙古高原草地生态系统的影响. 中国生态农业学报: 中英文, 2020, 28(12): 1847-1858. |
| [21] |
韩涛涛, 唐玄, 任海, 王俊, 刘楠, 郭勤峰. 群落/生态系统功能多样性研究方法及展望. 生态学报, 2021, 41(8): 3286-3295. |
| [22] |
张峰, 郑佳华, 赵萌莉, 陈大岭, 杨阳, 乔荠瑢. 刈割留茬高度对大针茅草原群落结构及稳定性的影响. 应用生态学报, 2020, 31(5): 1551-1559. |
| [23] |
张振, 李元恒, 丁勇, 李芳, 孙娟, 李西良. 典型草原群落及功能群物种多样性对放牧因素分解的响应. 中国草地学报, 2020, 42(5): 48-54. |
| [24] |
高露, 张圣微, 朱仲元, 徐冉, 张鹏. 放牧对干旱半干旱草原植物群落结构和生态功能的影响. 水土保持研究, 2019, 26(6): 205-211. |
| [25] |
刘燕丹, 乌日力嘎, 李元恒, 杨婷婷, 纪磊, 萨茹拉, 王云锋, 张慧, 丁勇. 不同放牧制度下典型草原生产效益与生态效应. 内蒙古大学学报: 自然科学版, 2021, 52(4): 425-436. |
| [26] |
王凯, 王聪, 冯晓明, 伍星, 傅伯杰. 生物多样性与生态系统多功能性关系研究进展. 生态学报, 2022(0): 11-23. |
| [27] |
张宇, 侯路路, 闫瑞瑞, 辛晓平. 放牧强度对草甸草原植物群落特征及营养品质的影响. 中国农业科学, 2020, 53(13): 2550-2561. |
| [28] |
向明学, 郭应杰, 古桑群宗, 张晓庆, 潘影, 武俊喜, 拉多. 不同放牧强度对拉萨河谷温性草原植物群落和物种多样性的影响. 草地学报, 2019, 27(3): 668-674. |
| [29] |
陈加际, 常生华, 王召锋, 彭泽晨, 程云湘, 陈先江, 侯扶江. 阿巴嘎旗典型草原植物物种多样性与地上生物量的关系. 草业科学, 2018, 35(9): 2068-2078. |
| [30] |
常虹, 孙海莲, 刘亚红, 邱晓, 石磊, 温超. 东乌珠穆沁草甸草原不同退化程度草地植物群落结构与多样性研究. 草地学报, 2020, 28(1): 184-192. |
| [31] |
徐粒, 高琼, 王亚林. 围封6年对温带典型草原坡地物种多样性及其与地上生物量的关系的影响. 生态环境学报, 2014, 23(3): 398-405. |
| [32] |
张小红, 宋彦涛, 乌云娜, 霍光伟, 张凤杰, 王晓光, 邸择雷. 放牧强度对克氏针茅草原植物功能群的影响. 草业科学, 2017, 34(10): 2033-2041. |
| [33] |
刘佳慧, 张韬. 放牧扰动对锡林郭勒典型草原植被特征及土壤养分的影响. 生态环境学报, 2017, 26(12): 2016-2023. |
| [34] |
孙宗玖, 朱进忠, 张鲜花. 短期放牧强度对昭苏草甸草原植被特征及多样性影响. 新疆农业大学学报, 2014, 37(1): 35-39. |
| [35] |
董乙强, 孙宗玖, 安沙舟, 荀其蕾. 放牧强度对伊犁绢蒿种群特征及其群落多样性的影响. 草地学报, 2016, 24(1): 22-27, 46. |
| [36] |
黄琛, 张宇, 赵萌莉, 韩国栋. 放牧强度对荒漠草原植被特征的影响. 草业科学, 2013, 30(11): 1814-1818. |
| [37] |
赵丽娅, 钟韩珊, 赵美玉, 张劲. 围封和放牧对科尔沁沙地群落物种多样性与地上生物量的影响. 生态环境学报, 2018, 27(10): 1783-1790. |
| [38] |
蒯晓妍, 邢鹏飞, 张晓琳, 梁艳, 王常慧, 董宽虎. 短期放牧强度对半干旱草地植物群落多样性和生产力的影响. 草地学报, 2018, 26(6): 1283-1289. |
| [39] |
宋洁, 王凤歌, 温璐, 王立新, 李金雷, 武胜男, 徐智超. 放牧对温带典型草原植物物种多样性及土壤养分的影响. 草地学报, 2019, 27(6): 1694-1701. |
| [40] |
董世魁, 汤琳, 张相锋, 刘世梁, 刘全儒, 苏旭坤, 张勇, 武晓宇, 赵珍珍, 李钰, 沙威. 高寒草地植物物种多样性与功能多样性的关系. 生态学报, 2017, 37(5): 1472-1483. |
| [41] |
赵生龙, 左小安, 张铜会, 吕朋, 岳平, 张晶. 乌拉特荒漠草原群落物种多样性和生物量关系对放牧强度的响应. 干旱区研究, 2020, 37(1): 168-177. |
| [42] |
Liu J, Feng C, Wang D L, Wang L, Wilsey B J, Zhong Z W. Impacts of grazing by different large herbivores in grassland depend on plant species diversity. Journal of Applied Ecology, 2015, 52(4): 1053-1062. |
| [43] |
Winck B R, Rigotti V M, Saccol de Sá E L. Effects of different grazing intensities on the composition and diversity of Collembola communities in southern Brazilian grassland. Applied Soil Ecology, 2019, 144: 98-106. |
| [44] |
Xiong D P, Shi P L, Zhang X Z, Zou C B. Effects of grazing exclusion on carbon sequestration and plant diversity in grasslands of China—A meta-analysis. Ecological Engineering, 2016, 94: 647-655. |
| [45] |
马文静, 张庆, 牛建明, 康萨如拉, 刘朋涛, 何欣, 杨艳, 张艳楠, 邬建国. 物种多样性和功能群多样性与生态系统生产力的关系——以内蒙古短花针茅草原为例. 植物生态学报, 2013, 37(7): 620-630. |
| [46] |
刘万弟, 李小伟, 黄文广, 马惠成, 马红英, 王文晓. 宁夏草原针茅属植物群落物种多样性和生产力格局及影响因素研究. 草业学报, 2021, 30(1): 12-23. |
| [47] |
牛钰杰, 杨思维, 王贵珍, 刘丽, 杜国祯, 花立民. 放牧干扰下高寒草甸物种和生活型丰富度与地上及地下生物量的关系. 生态学报, 2018, 38(8): 2791-2801. |