 2022, Vol. 42
2022, Vol. 42文章信息
- 范慧珠, 金光泽
- FAN Huizhu, JIN Guangze
- 氮添加对红松人工林草本层植物多样性的影响
- Effects of nitrogen addition on species diversity of the understory plants in the Korean pine plantation
- 生态学报. 2022, 42(23): 9747-9760
- Acta Ecologica Sinica. 2022, 42(23): 9747-9760
- http://dx.doi.org/10.5846/stxb202110122873
-
文章历史
- 收稿日期: 2021-10-12
- 网络出版日期: 2022-07-27





2. 东北林业大学森林生态系统可持续经营教育部重点实验室, 哈尔滨 150040;
3. 东北林业大学东北亚生物多样性研究中心, 哈尔滨 150040
2. Key Laboratory of Sustainable Forest Ecosystem Management of Ministry of Education, Northeast Forestry University, Harbin 150040, China;
3. Northeast Asia Biodiversity Research Center, Northeast Forestry University, Harbin 150040, China
含氮(N)化合物从大气中降落到周围地表的过程称为大气氮沉降, 是氮素生物地球化学循环中的重要一环, 然而大量化石燃料的使用, 致使氮排放量增加, 干扰了地球正常的氮循环[1—4]。我国氮沉降速率在近几十年内急剧增加[5]。氮沉降所带来的诸多问题中, 植物多样性的丧失受到了广泛关注[6]。在森林生态系统中, 草本层植物的物种多样性最高, 具有乔木幼苗、灌木、草本、木质藤本以及蕨类等多种植物, 其对环境变化(如氮沉降)的响应比森林上层植物更为敏感[6—8]。
目前, 多数研究认为氮添加降低草本层植物的多样性[6, 9—12], 但关于氮沉降对生物多样性的影响机制仍存在争议, 有研究认为由于植物自身特性和对资源的竞争能力存在差异, 致使不同物种对氮沉降的响应不同, 进而导致生物多样性发生改变[13—14];部分研究认为慢性氮沉降可以通过增加土壤中的有效氮来改变草本层植物物种组成及多样性[1, 6, 15];也有研究认为长期添加氮会改变土壤化学性质, 损害植物根系, 进而影响草本层植物盖度和多样性[15]。目前相关研究多数在高寒草甸和荒漠草原等生态系统中进行[16], 主要是比较同一功能群的不同物种之间的竞争关系, 而针对不同功能群植物的响应研究较少, 而且草原和草甸等生态系统缺乏高大乔木的遮光作用, 有可能放大光照对植物的影响[17—18], 森林相对于草原生态系统生物多样性更为丰富, 在水平和垂直方向上具有更明显的分层, 因而研究森林不同功能群对氮添加的响应有助于理解生物多样性对氮沉降的响应机制。
氮和磷(P)是大多数陆地生态系统中植物生长的限制元素[19], N、P元素的相对平衡比单一元素的绝对含量更为重要。土壤-植被系统的碳(C) ∶ N ∶ P生态化学计量变化与物种多样性变化关系十分密切[16, 20—23]。目前, 关于生态化学计量对植物多样性产生影响的观点主要表现在两个方面:第一, 土壤中C ∶ N ∶ P生态化学计量的改变将影响根系生长, 进而降低植物多样性[15, 24—25], 同时, 长期氮沉降会加速土壤氮循环, 引起土壤C ∶ N ∶ P失衡, 导致生态系统服务功能降低[16, 26—28];第二, 植物C ∶ N ∶ P生态化学计量发生改变, 导致植物养分失衡, 影响植物正常生长, 进而改变植物多样性[21, 29]。在全球相关研究的整合分析(Meta-analysis)中发现, 土壤C ∶ N ∶ P化学计量比植物C ∶ N ∶ P化学计量对于氮添加有着更低的敏感度[27]。目前, 已有研究将化学计量理论应用于解释氮沉降增加而引起的物种多样性变化[20], 但从草本层植物多样性与群落尺度上, 植物、土壤C ∶ N ∶ P生态化学计量关系的研究还比较缺乏, 这将限制对氮沉降引起植物多样性改变机制的理解。
温带森林是生物多样性重要的资源库之一, 其中物种多样性最高的是草本层植物, 红松人工林作为我国温带森林的重要组成部分, 目前已进行氮添加对红松人工林针叶和土壤的生态化学计量、土壤酶活性以及氮转化和氮损失等的影响[30—32], 缺乏氮添加对红松人工林草本层植物多样性的相关研究, 因此本研究依托红松人工林模拟氮沉降实验样地, 通过6年的野外控制氮添加实验, 拟探究氮添加对红松人工林草本层植物多样性的影响, 试图回答以下问题:(1)氮添加对草本层植物的组成结构和多样性有何影响, 对草本层植物生态化学计量和土壤化学性质有何影响;(2)产生影响的原因是什么?以期为理解氮沉降对温带森林生态系统结构与功能的影响提供基础数据和科学依据。
1 材料和方法 1.1 研究区概况研究区位于黑龙江凉水国家级自然保护区(47° 10′ 50″ N, 128° 53′ 20″ E), 地处小兴安岭南坡达里带岭支脉东坡。该保护区气候类型为大陆性季风气候, 年平均气温、年平均降水量和年平均蒸发量分别为-0.3℃、676 mm和805 mm, 年积雪时长130—150 d。土壤类型为暗棕壤。自然氮沉降为12.93 kg hm-2 a-1[33]。本研究在1954年造林的红松人工林中进行, 总面积达41.6 hm, 平均胸径16.0 cm。主要灌木有光萼溲疏(Deutzia gladata)、东北山梅花(Philadephus schrenii)和瘤枝卫矛(Euonymus verrucosu)等, 常见的蕨类植物有粗茎鳞毛蕨(Dryopteris crassirhizoma)、东北蹄盖蕨(Athyrium brevifrons)和掌叶铁线蕨(Adiantum pedatum)等, 草本植物有酢浆草(Oxalis corniculata)、唢呐草(Mitella nuda)、水金凤(Impatiens noli-tangere)和中华金腰(Chrysosplenium sinicum)等。
1.2 实验设计与样品采集测定本研究依托于红松人工林模拟氮沉降实验样地, 样地从2014年开始至2019年进行了6年的氮添加实验。在红松人工林中选取立地条件基本一致的区域, 设置20个5 m×30 m的样方, 样方间设置10 m间隔带, 避免样方之间相互干扰。样地设置4个梯度, 随机化区组, 每个施氮处理组设置5个重复, 以20kg hm-2 a-1作为低氮处理标准, 按2倍系数递增, 将梯度从低到高依次设置为, N0(对照处理, 0 kg hm-2 a-1)、N20(低氮处理, 20 kg hm-2 a-1)、N40(中氮处理, 40 kg hm-2 a-1)和N80(高氮处理, 80 kg hm-2 a-1), 氮源为硝酸铵(NH4NO3)。自2014年开始对样方施氮, 分四次(每年6—9月中旬)等量施氮, 将NH4NO3溶于20 L水中, 然后用背包喷雾器将溶液均匀喷洒至土壤表面, 对照组施加相同体积的水, 避免样方间因施水量不同而产生的差异。
2014年和2019年7月中旬进行草本层植物多样性调查。在每个样方中随机设置3个1 m×1 m的小样方, 调查草本层植物(高度≤1 m的草本、蕨类和木本植物)的种类、物种个体数(以地上部分单株进行统计)、高度(每个物种随机测量5株植物高度, 不足5株按实际株数测量)和盖度, 盖度采用网格法测定, 将1 m×1 m的小样方分为100个10 cm×10 cm的小网格, 并用目测法估算盖度。使用Winscanopy2006a冠层分析仪采集(由数码相机Coolpix 45000和180°的鱼眼镜头Nikon FC-E8组成)叶面积指数, 确保设备固定在距离地面1.3 m处且保持水平, 为避免阳光直射所产生的误差, 选择在阴天近日出时进行采集。采集的图像利用DHP软件处理, 计算0°-60°天顶角范围内的有效叶面积指数。
2019年7月中旬采集大样方内土壤和植物地上部分样品, 在上述每个小样方的附近随机设置3个1 m×1 m的小样方, 将小样方内的草本层植物全部进行收割, 按物种分类。同时, 在小样方去除表面凋落物、粗根、石块等杂物后, 采集0—10 cm的土壤样品。将采集的土壤样品及植物样品封装于聚乙烯封口袋中带回实验室测定化学性质。植物样品在65℃下烘干至恒定重量, 用球磨机研磨, 过0.149 mm筛;鲜土样品保存在4℃的冰箱中;风干土样品在实验室中进行自然风干后过2 mm和0.149 mm的土壤筛后测定样品的化学性质。全碳使用碳氮分析仪(multi N/C 3000, Germany)采用灼烧法测定, 全磷和全氮经过(H2SO4+H2O2)硝煮后, 利用AQ2 discrete analyzer (SEAL Analytical, Inc., USA)测定;使用酸度计(HANNA pH211, Italy)测定土壤样品的pH值, 土液比为1 ∶ 2.5;土壤的硝态氮和铵态氮含量采用连续流动分析仪(BRAN+LUEBBE-AA3, Germany)测定, 浸提液为2mol/L KCl, 土液比为1 ∶ 5。其化学计量比按质量计算。
1.3 数据处理利用Excel整理数据, 并计算草本层植物的个体数、相对密度、相对高度、相对盖度、相对优势度和群落相似系数。应用R 3.4.4软件[34]的Vegan包[35]计算草本层植物群落的α多样性, 包括:物种丰富度、Shannon-Wiener物种多样性指数和Pielou均匀度指数。利用基尼(Gini)系数表达各功能群植物高度的不对称性, 系数越大, 代表各功能群的高度不对称性越大, 功能群之间对光的竞争能力差异越明显。具体公式如下:
物种丰富度:
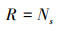
|
Shannon-Wiener多样性指数:
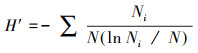
|
Pielou均匀度指数:
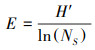
|
相对优势度:
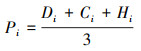
|
基尼系数:
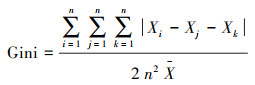
|
群落百分比相似系数:
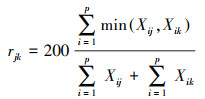
|
式中: Ns为群落中的总物种数;Ni为种i的个体数, N为所在群落的所有物种的个体数之和;Di是种i的相对密度, Ci是种i的相对盖度, Hi是种i的相对高度、Pi是种i的相对优势度;Xi、Xj和Xk分别表示功能群i、j和k植株的平均高度, n表示功能群数量, X表示所有功能群植株的平均高度;Xij和Xik分别表示两个不同群落中各物种的相对优势度。
利用多重比较和皮尔森相关分析对氮添加处理对草本层植物和土壤C ∶ N ∶ P生态化学计量进行分析, 先用Levene检验方差齐性, 若数据满足齐性检验, 则采用最小显著性差异法(least significant difference, LSD), 否则用Tamhane′s T2进行分析。使用层次分割分析(Hierarchical partioning analysis)和回归分析方法检验并解释氮添加导致植物和土壤的生物及非生物产生显著变化的因子对草本层植物多样性的影响, 层次分割分析可量化各因子对解释方差的独立贡献率, 使用R2作为模型的拟合优势度, 并根据95%置信区间确定随机独立贡献和统计的显著性[36—37]。层次分割分析通过R软件中的hier.part包[38]实现, 其他统计分析均在SPSS 22.0中完成, 绘图在Sigmaplot 10.0中完成。
2 结果 2.1 氮添加对草本层物种组成、群落结构和植物多样性的影响经过2014年与2019年两次草本层植物调查结果发现, 红松人工林草本层共出现34科、56种植物, 第1年N添加对各物种均未产生显著的影响, 然而6年的氮添加使其中6种植物在不同处理下的优势度存在显著差异(P < 0.05), 分别为酢浆草、白花野芝麻(Lamium barbatum)、黄芩(Scutellaria pekinensis)、宽叶荨麻(Urtica laetevirens)、狭叶荨麻(Urtica angustifolia)、唢呐草(附表 1)。
在第1年的氮添加实验中, 草本植物盖度在N40和N80处理下显著低于N20(P < 0.05), 其余草本层各功能群密度、高度和盖度均无显著变化(P>0.05)(图 1)。经过6年的氮添加实验, 草本植物在N40和N80处理显著低于N0和N20处理(P < 0.05), 蕨类和木本植物密度在施氮处理显著低于N0处理(P < 0.05)(图 1)。草本、蕨类和木本植物的高度在各处理间均无显著差异(P>0.05)(图 1)。草本植物的盖度在施氮处理显著低于N0处理(P < 0.05), 蕨类植物的盖度在N40和N80处理显著低于N0和N20(P < 0.05), 木本植物的盖度在N80处理均显著低于N0(P < 0.05)(图 1)。2014年和2019年, 各处理3个功能群的基尼系数均无显著差异(P>0.05)(图 2)。2014年不同氮添加条件下草本层的群落相似度无明显差异, 而在2019年随着施氮水平增加, 各施氮处理间群落相似度越小(表 1)。

|
| 图 1 氮添加对红松人工林草本层植物密度、高度和盖度的影响(平均值±标准误差, n=15) Fig. 1 The effect of nitrogen addition on the density, height and coverage of understory plants in the Korean pine plantation (mean±SE, n=15) N0:对照处理, 0 kg hm-2 a-1;N20:低氮处理, 20 kg hm-2 a-1;N40:中氮处理, 40 kg hm-2 a-1;N80:高氮处理, 80 kg hm-2 a-1;不同小写字母表示不同氮添加处理之间差异显著(P<0.05), 相同字母表示差异不显著(P>0.05) |

|
| 图 2 氮添加对红松人工林草本层植物基尼系数(不同功能群植物高度的不对称性)的影响(平均值±标准误差, n=15) Fig. 2 Change in Gini coefficient on the asymmetry of different functional groups plants height with nitrogen addition of understory plants in the Korean pine plantation (mean±SE, n=15) |
| 年份 Year |
施氮处理 Nitrogen treatment |
N0 | N20 | N40 | N80 |
| 2014 | N0 | — | |||
| N20 | 62.0 | — | |||
| N40 | 52.4 | 51.6 | — | ||
| N80 | 59.7 | 68.0 | 59.3 | — | |
| 2019 | N0 | — | |||
| N20 | 59.6 | — | |||
| N40 | 49.8 | 44.7 | — | ||
| N80 | 38.1 | 40.9 | 43.1 | — | |
| N0:对照处理, 0 kg hm-2 a-1;N20:低氮处理, 20 kg hm-2 a-1;N40:中氮处理, 40 kg hm-2 a-1;N80:高氮处理, 80 kg hm-2 a-1 | |||||
在第1年的氮添加实验中, 草本层植物多样性(R)、Shannon-Wiener多样性指数(H′)和Pielou均匀度指数(E)均无显著变化(P>0.05);经过6年的氮添加实验, 随着施氮浓度的增加草本植物的R和H′在N40和N80处理下均显著低于N0和N20(P < 0.05), 而蕨类和木本植物的R和H′在各处理间均无显著差异(P>0.05), 草本、蕨类和木本植物的E在各处理间均无显著差异(P>0.05)(图 3)。

|
| 图 3 氮添加对红松人工林草本层植物物种丰富度、Shannon-Wiener多样性指数和Pielou均匀度指数的影响(平均值±标准误差, n=15) Fig. 3 The effect of nitrogen addition on species richness, Shannon-Wiener diversity index and Pielou evenness index of understory plants in the Korean pine plantation (mean±SE, n=15) 不同小写字母表示不同氮添加处理之间差异显著(P<0.05), 相同字母表示差异不显著(P>0.05);在2019年调查发现, N40处理下仅出现一种蕨类植物, N80处理下仅出现一种木本植物, 因而其Shannon-Wiener多样性指数和Pielou均匀度指数为0 |
6年的氮添加实验后土壤的pH值在N20处理显著低于N0和N40(P < 0.05), 而土壤的C、N、P含量及其比值、硝态氮和铵态氮均无显著差异(P>0.05)(表 2)。氮添加对草本和木本植物的C ∶ N ∶ P生态化学计量产生不同的影响, 氮添加未对蕨类植物的C ∶ N ∶ P生态化学计量产生影响。氮添加使草本植物中, N0处理的C含量显著低于其余各处理(P < 0.05), N0和N20处理的N含量显著低于N40和N80处理(P < 0.05), N20处理的P含量显著低于其余各处理(P < 0.05), N0和N80处理的N ∶ P显著低于N20和N40处理(P < 0.05), N20处理的C ∶ P显著高于N0和N80处理(P < 0.05)。氮添加使木本植物中, N0和N20处理的P含量显著低于N80处理(P < 0.05), N20处理的N ∶ P显著高于其余各处理(P < 0.05), N20处理的C ∶ P显著高于N80处理(P < 0.05)(图 4)。
| 因子 Factor |
施氮处理 Nitrogen treatment | P | |||
| N0 | N20 | N40 | N80 | ||
| 土壤pH值 Soil pH | 5.32±0.18a | 5.06±0.25b | 5.30±0.37a | 5.17±0.32ab | < 0.05 |
| 土壤全碳 Soil C (g/kg) | 101.32±44.06 | 63.75±17.06 | 90.38±31.75 | 61.36±37.60 | 0.09 |
| 土壤全氮 Soil N (g/kg) | 11.84±3.40 | 12.56±5.31 | 11.83±5.62 | 12.76±5.09 | 0.97 |
| 土壤全磷 Soil P (g/kg) | 3.60±0.81 | 3.73±0.76 | 3.49±0.98 | 3.80±1.17 | 0.93 |
| 土壤硝态氮 Soil NO3-(mg/kg) | 9.75±6.28 | 4.30±4.16 | 10.90±8.47 | 6.35±4.43 | 0.09 |
| 土壤铵态氮 Soil NH4+(mg/kg) | 8.85±3.86 | 9.70±2.39 | 9.10±2.84 | 8.40±2.70 | 0.74 |
| 土壤氮磷比 Soil N∶P | 3.30±0.79 | 3.39±1.09 | 3.29±1.13 | 3.33±0.56 | 1 |
| 土壤碳氮比 Soil C∶N | 8.75±4.57 | 5.33±2.61 | 9.75±4.43 | 5.42±8.30 | 0.25 |
| 土壤碳磷比 Soil C∶P | 28.17±11.35 | 17.21±6.63 | 30.21±8.76 | 16.61±22.46 | 0.15 |
| 叶面积指数 Leaf area index | 4.61±0.12 | 4.67±0.10 | 4.60±0.14 | 4.57±0.18 | 0.966 |
| 单因素方差分析结果中P值小于0.05的部分, 再进行LSD检验, 不同小写字母表示不同氮添加处理之间差异显著(P<0.05), 相同字母表示差异不显著(P>0.05) | |||||

|
| 图 4 氮添加对红松人工林草本层植物C ∶ N ∶ P生态化学计量的影响(平均值±标准误差, n=15) Fig. 4 Effects of nitrogen addition on the ecological stoichiometry C ∶ N ∶ P of understory plants in the Korean pine plantation (mean±SE, n=15) |
氮添加仅对草本的丰富度和Shannon-Wiener多样性指数产生了显著影响, 因此利用层次分割分析量化了各因子对丰富度和Shannon-Wiener多样性指数的贡献率, 发现草本植物密度、盖度、生态化学计量和土壤化学性质共解释了物种丰富度和Shannon-Wiener多样性指数变异的60%和44%(表 3)。物种丰富度与植物盖度、密度、N含量均存在显著的相关关系(P < 0.01), Shannon-Wiener多样性指数与植物盖度、密度、N ∶ P均存在显著相关关系(P < 0.01), 盖度、密度、植物N含量解释了物种丰富度变异的18.83%、61.11%、5.74%, 盖度、密度、N ∶ P解释了Shannon-Wiener多样性指数变异的20.69%、43.72%、11.47%(表 3), 物种丰富度、Shannon-Wiener多样性指数与植物盖度、高度呈显著的正相关关系(P < 0.05), 而与植物N含量、N ∶ P呈显著的负相关关系(P < 0.01)(图 5)。
| 因子 Factor |
R2 | 草本植物 Herbs | 密度 Density |
盖度 Coverage |
土壤 Soil | ||||
| C | N | P | N∶P | C∶P | pH | ||||
| R | 0.6 | 1.37 | 5.74* | 3.27 | 5.07 | 3.69 | 61.11** | 18.83** | 0.92 |
| H′ | 0.44 | 2.43 | 9.48 | 8.04 | 11.47* | 3.15 | 43.72** | 20.69** | 1.02 |
| * P < 0.05, ** P < 0.01, *** P < 0.001 | |||||||||

|
| 图 5 红松人工林草本植物的N、N ∶ P、密度、盖度与物种多样性之间的相关关系 Fig. 5 The relationship between N, N ∶ P, density, coverage, and species diversity of herbs in the Korean pine plantation |
长期环境变化对群落的影响是分阶段进行的, 群落中物种消失, 群落结构发生变化, 从而影响群落物种多样性[12]。研究中发现第一年施氮, 各处理间群落相似度无明显变化, 而在施氮6年后, 随着施氮浓度的增加, 各施氮处理间群落相似度随之变小, 说明施氮导致物种组成和群落结构差异增大(表 1)。草本植物丰富度和Shannon-Wiener多样性指数显著减少, 而蕨类和木本植物无显著变化(图 3), 草本相对于其他功能群对氮添加反应更为敏感, 这可能与本地区的蕨类植物物种数和样本量少有关, 木本植物个体数少, 并且对氮添加反应相对迟钝有关。氮添加改变红松人工林草本层物种组成发生、密度和和盖度显著降低, 改变草本层群落结构, 从而降低植物多样性, 这一结果与多数北方温带森林的研究结果一致[10, 13—14, 39]。
植物经过长期的进化, 形成了独特的形态、生理生化和代谢特性, 植物体内元素的生态化学计量特征不仅体现了植物特有属性, 也反映了植物的生理代谢状况, 是物种对环境长期适应的结果[40]。植物叶片的养分含量, 尤其是N、P含量可以反映土壤养分的供给情况[41—42]。关于氮添加改变植物和土壤的生态化学计量, 从而影响草本植物多样性的原因, 目前有两种解释:
1) 研究认为氮添加改变土壤的化学性质, 增加土壤的毒性, 损害根系生长, 进而降低植物多样性[24—28]。氮添加会导致土壤养分异质化, 随着土壤中可利用养分的增加, 各物种生态位的重叠也随之增大, 物种间竞争加剧, 致使物种多样性降低[6, 43—44]。本研究结果发现, 6年的氮添加对土壤的C、N、P等含量及其比值均无显著差异(表 2), 尽管土壤pH值发生变化, 但随着施氮浓度的增加这种变化不显著, 可能因为土壤的总养分库远大于植物, 养分添加在总库中只占很小一部分, 因而响应不敏感。随着土壤中可用性氮的增加, 植物对氮的利用也随之增加, 进而导致土壤中可用性氮的改变不显著;也可能是由于采样时间处于夏季雨水较多的时间, 大量的雨水冲刷了土壤中可用性氮, 各处理的土壤可用性氮差异不显著。因而此观点不足以解释本研究中草本植物多样性降低的现象。
2) 研究认为氮添加使草本植物的C ∶ N ∶ P生态化学计量发生改变, 影响植物正常生长, 进而改变植物多样性[21, 29]。不同物种对于氮的利用特性不同, 对氮的耐受程度也存在差异[45—47]。本研究结果表明, 6年的氮添加使慢生型6种草本植物的相对优势度显著降低, 在不同施氮处理下各物种优势度均发生变化, 并且有4个物种消失(猪殃殃Galium spurium、露珠草Circaea cordata、林生茜草Rubia sylvatica、舞鹤草Maianthemum bifolium), 新增4个物种(白屈菜Chelidonium majus、北重楼Paris verticillate、狗尾草Setaria viridis、透茎冷水花Pilea pumila), 其中包括速生型物种(狗尾草)(附表 1), 说明氮添加降低红松人工林草本层植物多样性, 改变草本群落组成结构的原因主要与草本植物的氮利用特性有关, 过量的氮输入适合氮素营养转换速率较快的速生型物种的生存, 而氮素营养转换速率慢的慢生型物种的植物生长被限制和逐渐被淘汰[47], 从而导致草本群落的改变及生物多样性的降低。植物通过根系从土壤中吸收养分, 反过来土壤养分也受到植物的反馈调节[48—50]。但化学计量内稳态机制表明, 植物生态化学计量并非完全受控于土壤, 植物C ∶ N ∶ P化学计量对于土壤养分的响应会受到植物自身化学计量内稳态的调节[50—51]。本研究中, 草本植物物种丰富度、Shannon-Wiener多样性指数与植物盖度、密度呈显著的正相关关系, 草本植物丰富度与植物N含量呈显著的负相关关系, 草本植物Shannon-Wiener多样性指数与植物N ∶ P呈显著的负相关关系(表 3, 图 5), 氮添加引起草本植物的养分失衡, 尤其增加了植物对氮的吸收, N ∶ P存在显著差异说明各物种的养分阈值存在不同, 并且对氮的获取能力存在差异, 因而导致物种组成和群落结构发生改变, 从而导致红松人工林草本植物多样性降低。本实验结果支持这一观点。
氮添加显著降低草本植物的丰富度和Shannon-Wiener多样性指数, 而未对蕨类和木本植物产生显著影响, 导致该现象的原因除上文提到的氮添加引起草本层植物的养分失衡带来的影响外, 光照也可能是影响不同功能群植物生长的重要因素。冠层的叶面积指数决定了林内的光照[52], 叶面积指数越大, 光截获能力越强, 林下植被的光能利用率越低[53—56]。经过6年的氮添加红松人工林各施氮处理相对于对照的叶面积指数均无显著差异, 说明各样方光照条件无显著差异。高度是影响物种对光竞争能力的指标, 光竞争具有非对称性, 长得更高或者更快的植物要比长得更矮或者更慢的植物在单位时间和面积内获取的光能更多, 从而影响其对光的竞争能力[18, 57]。本研究中, 3个功能群在各施氮处理下高度均无显著差异(图 1), 并且各处理的基尼系数也无显著差异(图 2), 因而草本植物获取的光资源不变, 说明本地区草本植物丰富度和Shannon-Wiener多样性指数降低与光竞争无关, 说明6年的氮添加并未对草本植物的光竞争能力产生显著影响, 各功能群对光的竞争能力无明显变化。
由于本研究仅对施氮第一年与第六年夏季(生长旺季)的草本层植物进行分析, 而未考虑不同季节(如早春和秋季)和年际动态对氮添加的响应, 除上述原因外, 氮添加对植物的影响可能与土壤微生物类型及酶活性等存在相关性[31, 58—59], 因此关于氮沉降对草本层植物的影响机制仍需要进一步长期深入的研究。
4 结论6年氮添加改变植物群落中物种组成和群落结构、显著降低各功能群密度和盖度, 高度无显著差异。氮添加显著降低草本植物的丰富度和Shannon-Wiener多样性指数, 而未对蕨类和木本植物的丰富度和Shannon-Wiener多样性指数产生显著影响。本研究中, 氮添加显著降低草本植物物种丰富度和Shannon-Wiener多样性指数的主要因子为草本植物盖度、密度、N含量、N ∶ P, 其中草本物种丰富度、Shannon-Wiener多样性指数与植物盖度、密度呈显著的正相关关系, 与植物N含量、N ∶ P呈显著的负相关关系。本实验结果支持, 不同物种对于氮的利用特性和耐受程度存在差异, 氮添加引起草本植物的养分失衡, 增加了植物对氮的吸收, 引起物种组成和群落结构的改变, 从而影响草本植物多样性的观点。
附录| 物种 Species |
相对优势度 Relative advantages (2014) |
相对优势度 Relative advantages (2019) |
单因素方差分析 One-way ANOVA |
|||||||||
| N0 | N20 | N40 | N80 | N0 | N20 | N40 | N80 | P (2014) | P (2019) | |||
| 草本植物 | ||||||||||||
| 黄芩 Scutellaria pekinensis | 3.11±5.25 | 3.10±4.09 | 8.96±13.02 | 4.19±7.15 | 1.75±2.95a | 1.70±3.06a | 0.58±4.63b | 0.00b | 0.89 | < 0.05 | ||
| 宽叶荨麻 Urtica laetevirens | 2.45±6.15 | 3.17±17.09 | 0.50 | 1.26±0.99 | 3.77±8.25a | 2.59±5.09a | 0.00b | 0.00b | 0.65 | < 0.05 | ||
| 唢呐草 Mitella nuda | 5.73±8.11 | 6.81±7.83 | 2.53±6.01 | 2.40±3.64 | 5.66±11.14ab | 8.96±12.09a | 1.28±12.67b | 0.00b | 0.89 | < 0.05 | ||
| 狭叶荨麻 Urtica angustifolia | 1.06±3.11 | 1.99±3.08 | 1.51±0.57 | 1.62±5.27 | 3.66±10.09a | 1.67±4.86ab | 0.00b | 3.33±5.76a | 0.48 | < 0.05 | ||
| 白花野芝麻 Lamium barbatum | 2.06±3.76 | 2.25±3.23 | 0.00 | 2.42±4.14 | 1.30±2.83ab | 1.62±3.57ab | 0.90b | 2.51±7.00a | 0.35 | < 0.05 | ||
| 酢浆草 Oxalis corniculata | 8.79±14.81 | 5.84±18.65 | 4.52±10.73 | 5.66±19.74 | 6.99±11.69ab | 8.72±12.45a | 5.05±14.4b | 4.28±9.48b | 0.61 | < 0.01 | ||
| 猪殃殃 Galium spurium(-) | 1.65±4.11 | 0.00 | 0.00 | 0.00 | 0.70±1.20 | 1.37 | 0.00 | 0.00 | 0.40 | 0.57 | ||
| 露珠草 Circaea cordata(-) | 2.46±11.67 | 2.73±5.62 | 1.29±6.58 | 1.67±3.55 | 5.28±6.95 | 0.00 | 0.00 | 0.00 | 0.41 | 0.40 | ||
| 林生茜草 Rubia sylvatica(-) | 4.85±6.15 | 2.93±9.25 | 1.63±3.89 | 1.48±3.77 | 0.86±1.84 | 0.93±2.11 | 0.00 | 0.00 | 0.57 | 0.08 | ||
| 舞鹤草 Maianthemum bifolium(-) | 1.82±3.63 | 1.08±1.83 | 0.00 | 0.81 | 0.90±1.35 | 1.28±2.58 | 0.00 | 0.00 | 0.12 | 0.06 | ||
| 白屈菜 Chelidonium majus(+) | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 4.02 | 0.00 | - | 0.43 | ||
| 北重楼 Paris verticillate(+) | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 1.75 | 0.00 | - | 0.41 | ||
| 狗尾草 Setaria viridis(+) | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 2.22 | 0.00 | 0.00 | - | 0.41 | ||
| 透茎冷水花 Pilea pumila(+) | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 3.52 | 0.00 | 0.00 | - | 0.40 | ||
| 蔓孩儿参 Pseudostellaria davidii | 2.30±4.16 | 2.01±3.37 | 0.00 | 1.98±11.27 | 2.37±6.87 | 1.60±10.91 | 7.35±4.51 | 4.34±10.92 | 0.67 | 0.94 | ||
| 刺毛缘薹草 Carex pilosa var. auriculata | 1.18±2.57 | 0.00 | 1.56±2.46 | 4.31±16.9 | 0.80±2.02 | 1.62±3.57 | 1.78±5.87 | 1.48±7.19 | 0.28 | 0.80 | ||
| 类叶升麻 Actaea asiatica | 0.00 | 7.28±15.29 | 0.00 | 0.00 | 1.26±1.88 | 1.58±15.94 | 1.07 | 7.72 | 0.40 | 0.72 | ||
| 毒芹 Cicuta virosa | 0.00 | 0.00 | 0.00 | 0.00 | 0.78 | 0.00 | 1.82 | 0.00 | - | 0.58 | ||
| 东北羊角芹 Aegopodium alpestre | 2.22±4.44 | 2.54±7.14 | 2.15±6.25 | 3.00±8.93 | 4.13±6.70 | 3.67±6.52 | 4.14±15.21 | 14.64±24.7 | 0.29 | 0.50 | ||
| 荷青花 Hylomecon japonica | 0.00 | 0.00 | 0.00 | 0.00 | 2.95±10.38 | 4.35±15.48 | 6.23±5.30 | 1.46±2.50 | - | 0.50 | ||
| 白花碎米荠 Cardamine leucantha | 0.00 | 0.00 | 0.00 | 0.00 | 0.69 | 0.00 | 0.00 | 1.05 | - | 0.47 | ||
| 蔓乌头 Aconitum volubile | 1.87 | 0.00 | 0.00 | 0.00 | 1.42 | 0.00 | 0.00 | 0.00 | 0.40 | 0.40 | ||
| 球果堇菜 Viola collina | 1.00±1.30 | 0.76±2.41 | 0.00 | 0.00 | 0.00 | 0.33±1.58 | 0.00 | 0.00 | 0.15 | 0.40 | ||
| 山尖子 Parasenecio hastatus | 0.00 | 0.50 | 0.00 | 3.25 | 0.00 | 0.00 | 0.00 | 3.96 | 0.40 | 0.40 | ||
| 水金凤 Impatiens noli-tangere | 4.93±7.04 | 2.12±3.90 | 1.36±2.05 | 3.32±4.71 | 1.34±1.82 | 4.32±12.7 | 5.26±30.1 | 9.7±18.69 | 0.30 | 0.40 | ||
| 水杨梅 Geum chiloense | 0.00 | 1.21 | 0.00 | 1.88 | 0.00 | 0.00 | 4.2 | 0.00 | 0.31 | 0.40 | ||
| 问荆 Equisetum arvense | 0.00 | 0.00 | 0.00 | 0.00 | 0.31 | 0.00 | 0.00 | 0.00 | - | 0.40 | ||
| 拟扁果草 Enemion raddeanum | 1.20±1.70 | 2.08±2.29 | 1.52±2.58 | 1.95±6.58 | 0.75±2.65 | 0.97±2.14 | 1.7±5.93 | 3.36±42.87 | 0.13 | 0.36 | ||
| 唐松草 Thalictrum aquilegiifolium var. sibiricum | 0.00 | 5.25±11.77 | 0.00 | 0.00 | 0.00 | 1.73±3.95 | 0.77 | 0.00 | 0.30 | 0.35 | ||
| 木贼 Equisetum hyemale | 3.22 | 0.00 | 0.00 | 2.03±8.38 | 1.43 | 2.28 | 1.58 | 8.25±24.71 | 0.57 | 0.32 | ||
| 中华金腰 Chrysosplenium sinicum | 7.59±16.61 | 12.96±20.74 | 19.96±21.64 | 17.35±18.64 | 11.73±12.62 | 5.03±16.52 | 22.71±29.78 | 2.78±11.25 | 0.95 | 0.30 | ||
| 掌叶蚊子草 Filipendula palmata | 1.92±4.91 | 2.02±4.61 | 9.79±16.30 | 2.05±2.53 | 2.58±6.80 | 1.95±4.51 | 0.56±3.64 | 1.53±4.10 | 0.89 | 0.27 | ||
| 五福花 Adoxa moschatellina | 0.00 | 0.00 | 0.63±1.86 | 0.00 | 0.00 | 0.67±2.05 | 0.39±1.91 | 0.00 | 0.40 | 0.26 | ||
| 透骨草 Phryma leptostachya subsp. asiatica | 6.34±14.23 | 0.00 | 0.00 | 0.00 | 10.93±15.37 | 0.00 | 0.00 | 0.00 | 0.40 | 0.23 | ||
| 蕨类植物 | ||||||||||||
| 假冷蕨 Athyrium spinulosum(-) | 2.42±17.32 | 3.59±10.99 | 3.72±10.46 | 2.52±4.15 | 0.00 | 0.00 | 0.00 | 0.00 | 0.85 | - | ||
| 粗茎鳞毛蕨 Dryopteris crassirhizoma | 1.94±8.93 | 1.97±3.40 | 4.49±16.92 | 2.53±8.13 | 3.92±4.49 | 0.00 | 8.84±14.95 | 9.98±13.97 | 0.06 | 0.61 | ||
| 东北蹄盖蕨 Athyrium brevifrons | 0.00 | 0.00 | 0.00 | 4.17±5.93 | 4.62±9.55 | 4.55±25.31 | 3.69±35.64 | 4.94±16.03 | 0.10 | 0.27 | ||
| 掌叶铁线蕨 Athyrium pedatum | 0.00 | 1.95 | 0.40 | 3.99±6.77 | 0.00 | 4.09 | 0.00 | 4.27±24.48 | 0.61 | 0.22 | ||
| 木本植物 | ||||||||||||
| 白桦 Betula platyphylla(-) | 0.00 | 0.29±0.14 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.40 | - | ||
| 丁香 Syringa reticulata(-) | 0.00 | 1.21 | 4.14 | 1.59 | 0.00 | 0.00 | 0.00 | 0.00 | 0.57 | - | ||
| 毛榛 Corylus mandshurica(-) | 0.00 | 3.83 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.40 | - | ||
| 青楷槭 Acer tegmentosum(-) | 0.00 | 1.94±1.87 | 0.00 | 1.12 | 0.00 | 0.00 | 0.00 | 0.00 | 0.18 | - | ||
| 色木槭 Acer pictum subsp. mono(-) | 0.77±3.11 | 0.00 | 3.88±16.98 | 0.00 | 3.21±6.13 | 0.00 | 0.00 | 0.00 | 0.27 | 0.40 | ||
| 忍冬 Lonicera japonica(-) | 2.78±17.04 | 0.00 | 7.05±10.3 | 0.00 | 0.79±4.69 | 0.61 | 0.00 | 0.00 | 0.11 | 0.15 | ||
| 狗枣猕猴桃 Actinidia kolomikta(-) | 3.08±11.93 | 2.68±11.16 | 2.92±12.44 | 1.50±10.03 | 2.04±8.46 | 1.65±8.15 | 1.59±5.62 | 0.00 | 0.84 | 0.34 | ||
| 光萼溲疏 Deutzia gabrata(-) | 4.45±10.00 | 2.41±4.52 | 3.51±9.91 | 4.03±5.37 | 1.53±3.64 | 1.33±2.49 | 1.79±15.57 | 0.00 | 0.35 | 0.29 | ||
| 瘤枝卫矛 Euonymus verrucosus(-) | 0.50±2.06 | 0.00 | 1.21±1.51 | 1.97 | 1.08±2.85 | 0.91 | 0.00 | 0.00 | 0.49 | 0.21 | ||
| 绣线菊 Spiraea salicifolia(+) | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 3.42 | 0.00 | - | 0.40 | ||
| 红松 Pine koraiensis | 0.00 | 0.65 | 0.00 | 0.66±2.63 | 0.00 | 0.00 | 0.52 | 0.91±3.11 | 0.37 | 0.27 | ||
| 花楷槭 Acer ukurunduense | 2.40 | 0.00 | 1.71±4.79 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.50 | - | ||
| 山杨 Populus davidiana | 1.65 | 0.00 | 3.79±2.91 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.12 | - | ||
| 刺果茶藨子 Ribes burejense | 0.00 | 0.00 | 1.07±4.32 | 0.00 | 0.68 | 1.61 | 4.29±6.90 | 0.00 | 0.40 | 0.56 | ||
| 东北山梅花 Philadephus schrenii | 5.56±10.94 | 4.79±6.81 | 3.22±9.75 | 4.74±30.86 | 3.47±5.38 | 10.33±12.48 | 2.18±15.04 | 0.00 | 0.92 | 0.56 | ||
| 茶藨子 Ribes janczewskii | 0.00 | 2.74±6.00 | 0.00 | 4.93±2.01 | 0.00 | 2.05±17.37 | 0.00 | 3.74±9.87 | 0.10 | 0.55 | ||
| 山葡萄 Vitis amurensis | 1.6±5.67 | 1.88 | 0.00 | 0.00 | 2.23±21.46 | 7.20 | 0.00 | 0.00 | 0.58 | 0.53 | ||
| 刺五加 Eleutherococcus senticosus | 5.10±6.39 | 1.62±7.87 | 1.00±3.18 | 3.57±7.45 | 2.11±5.95 | 0.99±2.05 | 0.52±4.84 | 5.73±30.24 | 0.29 | 0.07 | ||
| N0:对照处理, 0 kg hm-2 a-1;N20:低氮处理, 20 kg hm-2 a-1;N40:中氮处理, 40 kg hm-2 a-1;N80:高氮处理, 80 kg hm-2 a-1;不同小写字母表示不同氮添加处理之间差异显著(P<0.05), 相同字母表示差异不显著(P>0.05);(+)新增物种(该物种2014年施氮处理的样方中未出现该物种, 2019施氮处理的样方中存在, 且对照组六年前后未消失或新增);(-)消失物种(该物种2014年施氮处理的样方中存在该物种, 2019施氮处理的样方中未出现, 且对照组在施氮六年前后未消失或新增) | ||||||||||||
| [1] |
Fenn M E, Poth M A, Aber J D, Baron J S, Bormann B T, Johnson D W, Lemly A D, McNulty S G, Ryan D F, Stottlemyer R. Nitrogen excess in north American ecosystems: predisposing factors, ecosystem responses, and management strategies. Ecological Applications, 1998, 8(3): 706-733. DOI:10.1890/1051-0761(1998)008[0706:NEINAE]2.0.CO;2 |
| [2] |
Liu X J, Duan L, Mo J M, Du E Z, Shen J L, Lu X K, Zhang Y, Zhou X B, He C N, Zhang F S. Nitrogen deposition and its ecological impact in China: an overview. Environmental Pollution, 2011, 159(10): 2251-2264. DOI:10.1016/j.envpol.2010.08.002 |
| [3] |
Galloway J N, Dentener F J, Capone D G, Boyer E W, Howarth R W, Seitzinger S P, Asner G P, Cleveland C C, Green P A, Holland E A, Karl D M, Michaels A F, Porter J H, Townsend A R, Vöosmarty C J. Nitrogen cycles: past, present, and future. Biogeochemistry, 2004, 70(2): 153-226. DOI:10.1007/s10533-004-0370-0 |
| [4] |
常运华, 刘学军, 李凯辉, 吕金岭, 宋韦. 大气氮沉降研究进展. 干旱区研究, 2012, 29(6): 972-979. DOI:10.13866/j.azr.2012.06.002 |
| [5] |
Yu G R, Jia Y L, He N P, Zhu J X, Chen Z, Wang Q F, Piao S L, Liu X J, He H L, Guo X B, Wen Z, Li P, Ding G A, Goulding K. Stabilization of atmospheric nitrogen deposition in China over the past decade. Nature Geoscience, 2019, 12(6): 424-429. DOI:10.1038/s41561-019-0352-4 |
| [6] |
Gilliam F S. Response of the herbaceous layer of forest ecosystems to excess nitrogen deposition. Journal of Ecology, 2006, 94(6): 1176-1191. DOI:10.1111/j.1365-2745.2006.01155.x |
| [7] |
Gilliam F S. Effects of Excess Nitrogen Deposition on the Herbaceous Layer of Eastern North American Forests. New York: Oxford University Press, 2014: 445-459.
|
| [8] |
Gilliam F S. The ecological significance of the herbaceous layer in temperate forest ecosystems. BioScience, 2007, 57(10): 845-858. DOI:10.1641/B571007 |
| [9] |
Lu X K, Mo J M, Gilliam F S, Zhou G Y, Fang Y T. Effects of experimental nitrogen additions on plant diversity in an old-growth tropical forest. Global Change Biology, 2010, 16(10): 2688-2700. DOI:10.1111/j.1365-2486.2010.02174.x |
| [10] |
Lai M Y, He S C, Yu S, Jin G Z. Effects of experimental N addition on plant diversity in an old-growth temperate forest. Ecology and Evolution, 2018, 8(11): 5900-5911. DOI:10.1002/ece3.4127 |
| [11] |
Kwak J H, Chang S X, Naeth M A. Eleven years of simulated deposition of nitrogen but not sulfur changed species composition and diversity in the herb stratum in a boreal forest in western Canada. Forest Ecology and Management, 2018, 412: 1-8. DOI:10.1016/j.foreco.2018.01.049 |
| [12] |
Smith M D, Knapp A K, Collins S L. A framework for assessing ecosystem dynamics in response to chronic resource alterations induced by global change. Ecology, 2009, 90(12): 3279-3289. DOI:10.1890/08-1815.1 |
| [13] |
Jung K, Kwak J H, Gilliam F S, Chang S X. Simulated N and S deposition affected soil chemistry and understory plant communities in a boreal forest in western Canada. Journal of Plant Ecology, 2018, 11(4): 511-523. DOI:10.1093/jpe/rtx030 |
| [14] |
Xing A J, Xu L C, Shen H H, Du E Z, Liu X Y, Fang J Y. Long term effect of nitrogen addition on understory community in a Chinese boreal forest. Science of the Total Environment, 2019, 646: 989-995. DOI:10.1016/j.scitotenv.2018.07.350 |
| [15] |
Lu X K, Mao Q G, Gilliam F S, Luo Y Q, Mo J M. Nitrogen deposition contributes to soil acidification in tropical ecosystems. Global Change Biology, 2014, 20(12): 3790-3801. DOI:10.1111/gcb.12665 |
| [16] |
牛玉斌, 余海龙, 王攀, 樊瑾, 王艳红, 黄菊莹. 氮磷添加对荒漠草原植物群落多样性和土壤C ∶ N ∶ P生态化学计量特征的影响. 生态学报, 2019, 39(22): 8462-8471. |
| [17] |
Bret-Harte M S, Mack M C, Goldsmith G R, Sloan D B, DeMarco J, Shaver G R, Ray P M, Biesinger Z, Chapin Ⅲ F S. Plant functional types do not predict biomass responses to removal and fertilization in Alaskan tussock tundra. Journal of Ecology, 2008, 96(4): 713-726. DOI:10.1111/j.1365-2745.2008.01378.x |
| [18] |
邹安龙, 马素辉, 倪晓凤, 蔡琼, 李修平, 吉成均. 模拟氮沉降对北京东灵山辽东栎群落林下植物物种多样性的影响. 生物多样性, 2019, 27(6): 607-618. |
| [19] |
Güsewell S, Koerselman W, Verhoeven J T A. Biomass N ∶ P ratios as indicators of nutrient limitation for plant populations in wetlands. Ecological Applications, 2003, 13(2): 372-384. DOI:10.1890/1051-0761(2003)013[0372:BNRAIO]2.0.CO;2 |
| [20] |
Tian D, Du E Z, Jiang L, Ma S H, Zeng W J, Zou A L, Feng C Y, Xu L C, Xing A J, Wang W, Zheng C Y, Ji C J, Shen H H, Fang J Y. Responses of forest ecosystems to increasing N deposition in China: a critical review. Environmental Pollution, 2018, 243: 75-86. DOI:10.1016/j.envpol.2018.08.010 |
| [21] |
Güsewell S. N ∶ P ratios in terrestrial plants: variation and functional significance. New Phytologist, 2004, 164(2): 243-266. DOI:10.1111/j.1469-8137.2004.01192.x |
| [22] |
陈军强, 张蕊, 侯尧宸, 马丽娜, 丁路明, 龙瑞军, 尚占环. 亚高山草甸植物群落物种多样性与群落C、N、P生态化学计量的关系. 植物生态学报, 2013, 37(11): 979-987. |
| [23] |
杨祥祥, 李梦琦, 何兴东, 王学增, 尤万学, 余殿, 张彩华, 陈娜. 沙地植被碳氮磷化学计量特征与物种多样性的关系. 应用生态学报, 2018, 29(9): 2819-2824. DOI:10.13287/j.1001-9332.201809.016 |
| [24] |
Bobbink R, Hicks K, Galloway J, Spranger T, Alkemade R, Ashmore M, Bustamante M, Cinderby S, Davidson E, Dentener F, Emmett B, Erisman J W, Fenn M, Gilliam F, Nordin A, Pardo L, De Vries W. Global assessment of nitrogen deposition effects on terrestrial plant diversity: a synthesis. Ecological Applications, 2010, 20(1): 30-59. DOI:10.1890/08-1140.1 |
| [25] |
Yue K, Jarvie S, Senior A M, Van Meerbeek K, Peng Y, Ni X Y, Wu F Z, Svenning J C. Changes in plant diversity and its relationship with productivity in response to nitrogen addition, warming and increased rainfall. Oikos, 2020, 129(7): 939-952. DOI:10.1111/oik.07006 |
| [26] |
Yang Y H, Fang J Y, Ji C J, Datta A, Li P, Ma W H, Mohammat A, Shen H H, Hu H F, Knapp B O, Smith P. Stoichiometric shifts in surface soils over broad geographical scales: evidence from China's grasslands. Global Ecology and Biogeography, 2014, 23(8): 947-955. DOI:10.1111/geb.12175 |
| [27] |
Yue K, Fornara D A, Yang W Q, Peng Y, Li Z J, Wu F Z, Peng C H. Effects of three global change drivers on terrestrial C ∶ N ∶ P stoichiometry: a global synthesis. Global Change Biology, 2017, 23(6): 2450-2463. DOI:10.1111/gcb.13569 |
| [28] |
Elser J J, Fagan W F, Kerkhoff A J, Swenson N G, Enquist B J. Biological stoichiometry of plant production: Metabolism, scaling and ecological response to global change. New Phytologist, 2010, 186(3): 593-608. DOI:10.1111/j.1469-8137.2010.03214.x |
| [29] |
Güsewell S, Bailey K M, Roem W J, Bedford B L. Nutrient limitation and botanical diversity in wetlands: Can fertilisation raise species richness?. Oikos, 2005, 109(1): 71-80. DOI:10.1111/j.0030-1299.2005.13587.x |
| [30] |
Yang D X, Song L, Jin G Z. The soil C ∶ N ∶ P stoichiometry is more sensitive than the leaf C ∶ N ∶ P stoichiometry to nitrogen addition: a four-year nitrogen addition experiment in a Pinus koraiensis plantation. Plant and Soil, 2019, 442(1/2): 183-198. |
| [31] |
吕来新, 宋蕾, 刘志理, 张金波, 金光泽. 红松人工林土壤酶活性与化学性质对氮添加的响应. 环境科学, 2020, 41(4): 1960-1967. DOI:10.13227/j.hjkx.201906168 |
| [32] |
Song L, Zhang J B, Müller C, Jin G Z. Responses of soil N transformations and N loss to three years of simulated N deposition in a temperate Korean pine plantation in northeast China. Applied Soil Ecology, 2019, 137: 49-56. DOI:10.1016/j.apsoil.2019.01.008 |
| [33] |
宋蕾, 田鹏, 张金波, 金光泽. 黑龙江凉水国家级自然保护区大气氮沉降特征. 环境科学, 2018, 39(10): 4490-4496. DOI:10.13227/j.hjkx.201801215 |
| [34] |
The R Core Team. R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing, 2018. |
| [35] |
Oksanen J, Blanchet F G, Friendly M, Kindt R, Legendre P, McGlinn D, Minchin P R, O'Hara R B, Simpson G L, Solymos P, Stevens M H H, Szoecs E, Wagner H. Vegan: Community Ecology Package, 2019.
|
| [36] |
Chevan A, Sutherland M. Hierarchical partitioning. The American Statistician, 1991, 45(2): 90-96. |
| [37] |
Fujita Y, Van Bodegom P M, Witte J P M. Relationships between nutrient-related plant traits and combinations of soil N and P fertility measures. PLoS One, 2013, 8(12): e83735. DOI:10.1371/journal.pone.0083735 |
| [38] |
Walsh C, Mac Nally R. hier. part: Hierarchical partitioning. R package version 1.0-4, 2013.
|
| [39] |
Hurd T M, Brach A R, Raynal D J. Response of understory vegetation of Adirondack forests to nitrogen additions. Canadian Journal of Forest Research, 1998, 28(6): 799-807. DOI:10.1139/x98-045 |
| [40] |
高宗宝, 王洪义, 吕晓涛, 王正文. 氮磷添加对呼伦贝尔草甸草原4种优势植物根系和叶片C ∶ N ∶ P化学计量特征的影响. 生态学杂志, 2017, 36(1): 80-88. |
| [41] |
Koerselman W, Meuleman A F M. The vegetation N ∶ P ratio: a new tool to detect the nature of nutrient limitation. Journal of Applied Ecology, 1996, 33(6): 1441-1450. DOI:10.2307/2404783 |
| [42] |
You C M, Wu F Z, Yang W Q, Xu Z F, Tan B, Yue K, Ni X Y. Nutrient-limited conditions determine the responses of foliar nitrogen and phosphorus stoichiometry to nitrogen addition: a global meta-analysis. Environmental Pollution, 2018, 241: 740-749. DOI:10.1016/j.envpol.2018.06.018 |
| [43] |
Wardle D A, Bardgett R D, Klironomos J N, Setälä H, Van Der Putten W H, Wall D H. Ecological linkages between aboveground and belowground biota. Science, 2004, 304(5677): 1629-1633. DOI:10.1126/science.1094875 |
| [44] |
Harpole W S, Tilman D. Grassland species loss resulting from reduced niche dimension. Nature, 2007, 446(7137): 791-793. DOI:10.1038/nature05684 |
| [45] |
Bobbink R, Hornung M, Roelofs J G M. The effects of air-borne nitrogen pollutants on species diversity in natural and semi-natural European vegetation. Journal of Ecology, 1998, 86(5): 717-738. DOI:10.1046/j.1365-2745.1998.8650717.x |
| [46] |
Aerts R, Van Der Peijl M J. A simple model to explain the dominance of low-productive perennials in nutrient-poor habitats. Oikos, 1993, 66(1): 144-147. DOI:10.2307/3545208 |
| [47] |
鲁显楷, 莫江明, 董少峰. 氮沉降对森林生物多样性的影响. 生态学报, 2008, 28(11): 5532-5548. DOI:10.3321/j.issn:1000-0933.2008.11.038 |
| [48] |
Jobbágy E G, Jackson R B. The distribution of soil nutrients with depth: global patterns and the imprint of plants. Biogeochemistry, 2001, 53(1): 51-77. DOI:10.1023/A:1010760720215 |
| [49] |
Cleveland C C, Liptzin D. C ∶ N ∶ P stoichiometry in soil: Is there a "Redfield ratio" for the microbial biomass?. Biogeochemistry, 2007, 85(3): 235-252. DOI:10.1007/s10533-007-9132-0 |
| [50] |
Garrish V, Cernusak L A, Winter K, Turner B L. Nitrogen to phosphorus ratio of plant biomass versus soil solution in a tropical pioneer tree, Ficus insipida. Journal of Experimental Botany, 2010, 61(13): 3735-3748. DOI:10.1093/jxb/erq183 |
| [51] |
庾强. 内蒙古草原植物化学计量生态学研究[D]. 北京: 中国科学院植物研究所, 2009.
|
| [52] |
苑振皓, 金光泽, 刘志理. 自动曝光对半球摄影法测量叶面积指数及其季节变化的影响. 应用生态学报, 2018, 29(12): 4004-4012. |
| [53] |
Matsuo T, Martínez-Ramos M, Bongers F, Van Der Sande M T, Poorter L. Forest structure drives changes in light heterogeneity during tropical secondary forest succession. Journal of Ecology, 2021, 109(8): 2871-2884. |
| [54] |
Kitajima K, Mulkey S S, Wright S J. Variation in crown light utilization characteristics among tropical canopy trees. Annals of Botany, 2005, 95(3): 535-547. |
| [55] |
Lebrija-Trejos E, Pérez-García E A, Meave J A, Bongers F, Poorter L. Functional traits and environmental filtering drive community assembly in a species-rich tropical system. Ecology, 2010, 91(2): 386-398. |
| [56] |
Whitmore T C, Brown N D, Swaine M D, Kennedy D, Goodwin-Bailey C I, Gong W K. Use of hemispherical photographs in forest ecology: measurement of gap size and radiation totals in a Bornean tropical rain forest. Journal of Tropical Ecology, 1993, 9(2): 131-151. |
| [57] |
Demalach N, Zaady E, Kadmon R. Light asymmetry explains the effect of nutrient enrichment on grassland diversity. Ecology Letters, 2017, 20(1): 60-69. |
| [58] |
王丽君, 程瑞梅, 肖文发, 沈雅飞, 曾立雄, 杨邵, 孙鹏飞, 陈天. 三峡库区马尾松人工林土壤酶活性和微生物生物量对氮添加的季节性响应. 生态学报, 2021, 41(24): 9857-9868. |
| [59] |
张海芳. 贝加尔针茅草原植物与土壤微生物群落对氮素和水分添加的响应[D]. 北京: 中国农业科学院, 2017.
|