 2022, Vol. 42
2022, Vol. 42文章信息
- 朱时应, 王壮壮, 黄倩, 贺凯, 李天顺, 普布
- ZHU Shiying, WANG Zhuangzhuang, HUANG Qian, HE Kai, LI Tian, PU Bu
- 西藏年楚河流域湿地土壤纤毛虫群落特征
- Characteristics of soil ciliate community in wetland of the Nianchu River Basin, Tibet, China
- 生态学报. 2022, 42(22): 9005-9016
- Acta Ecologica Sinica. 2022, 42(22): 9005-9016
- http://dx.doi.org/10.5846/stxb202203290781
-
文章历史
- 收稿日期: 2022-03-29

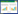




土壤是陆地生态系统中最为复杂成分之一, 为土壤动物提供了生存条件[1]。湿地生态系统是由陆地生态系统和水生态系统形成的综合生态系统, 有着“生物超市”和“地球之肾”的美称[2]。原生动物是自然界最原始、最简单的个体。土壤原生动物指的是生活在土壤中或者是土壤表面凋落物中的原生动物[3]。土壤纤毛虫一般泛指生活在土壤颗粒、土壤孔隙水中和覆盖在土壤表面凋落物水膜中的土壤原生动物, 土壤纤毛虫具有种类繁多, 体积微小, 形态多样, 纤薄细胞膜等特点, 并且能够对外界环境微小变化作出快速响应, 因此对土壤生态系统有着非常重要的指示作用[4]。国内目前对于土壤纤毛虫的研究集中于土壤纤毛虫群落特征和形态学描述, 如宁应之等对退牧还草、退耕还林生态恢复下土壤纤毛虫群落响应的研究, 结果显示已经退牧还草、退耕还林样地与尚未退牧还草、退耕还林的样地之间土壤纤毛虫物种分布存在差异[5—9];刘旻霞等对不同林地的土壤纤毛虫群落进行了研究, 表明不同林地中人工林土壤纤毛虫多样性指数最高[10—15];万玛央措等就甘肃农田四种土壤腹毛类纤毛虫进行了形态学描述, 发现在中国首次记录到柠檬管膜虫(Cytohymena citrina)、沙栖蠕形尖毛虫(Vermioxytericha arenicola)和背触毛拟腹柱虫(Gastrostylides dorsiciratus)[16]。
西藏地处青藏高原西南部, 因其独特的地理位置, 孕育了特殊的高原湿地生态系统, 蕴含着极为丰富的湿地生物多样性资源。对于土壤纤毛虫的研究在西藏地区就仅见普布、巴桑等人对双湖地区和拉鲁湿地的报道[17—18], 对于年楚河流域湿地土壤纤毛虫群落特征研究尚未见报道。年楚河流域湿地生态系统在保护生物多样性、调节小气候、改善水质、防止土壤沙化、减少温室效应等方面具有重要的作用, 但是该流域湿地生态系统十分脆弱, 一旦遭到外界不利因素的干扰, 很难恢复到原有状态, 研究年楚河流域湿地对于维持和保护青藏高原生态环境具有不可忽视的作用。土壤纤毛虫因其能够对于环境的微小变化做出积极的响应, 常被用作土壤变化的指示生物, 因此本文根据年楚河流域湿地分布特征, 选取了5个典型的湿地样地, 对不同季节土壤纤毛虫群落特征开展调查, 旨在深入研究年楚河流域湿地土壤纤毛虫物种组成及群落结构, 为该流域生物多样性提供基础研究资料, 从而为保护年楚河流域湿地生态系统, 促进流域湿地生态系统健康可持续发展提供科学依据。
1 研究区域概况和研究方法 1.1 研究区域概况年楚河是雅鲁藏布江右岸支流之首, 东起卡惹拉雪山, 西接夏布曲流域, 面积约为111 30km2, 整个流域位置在北纬28°10′—29°20′和东经88°35′—90°15′之间, 河流全长217km, 以河流湿地和沼泽湿地为主[19]。高原温带半干旱区覆盖整个流域, 形成了温度低, 太阳光辐射大等典型的高原气候特征, 该地区年平均降水量为365mm, 受地形和水汽的影响降水分布十分不均[20—22]。常见的植物有西藏嵩草(Kobresia tibetica)、长华筒马先蒿(Pedicularis longiflora)、青藏薹草(Carex moorcroftii)、蕨麻委陵菜(Potentilla anserina)等。湿地常见动物有赤麻鸭(Tadorna ferruginea)、斑头雁(Anser indicus)、普通秋沙鸭(Mergus merganser)等。
1.2 研究方法根据年楚河流域湿地分布特点, 以样地间距离为参考, 选取了五个典型未退化的高寒湿地作为样地[22], 即SD1、SD2、SD3、SD4和SD5(图 1)。分别在2020年5月(春)、8月(夏)和10月(秋)三个季节进行了三次采样, 在每个样地(20m×20m), 用GPS(型号: MAP 63lcsx厂家: 上海佳明航电企业管理有限公司)记录海拔及经纬度, 采用梅花五点采样法选取五个样方(20cmx20cm), 每个样方中用土壤温枪(型号: XTY0087)测定土壤表层温度(ST), 拣去土壤表面的凋落物后用土壤采集器采集0—10cm土壤土样(约4000g), 共获得75(5×5×3)个样品。将采集好的样品密封后带回实验室, 在牛皮纸上平整的铺开, 用草纸覆盖(防止空气中漂浮的原生动物影响后续实验结果), 在干燥阴凉处自然风干(30 d左右)。

|
| 图 1 年楚河流域湿地采样地示意图 Fig. 1 Schematic diagram of sampling area in wetland of the Nianchu River Basin SD1—SD5为样地 |
用干湿比重法测定土壤湿度(SWC), 其它土壤理化因子委托西藏博源环境检测有限公司进行检测, 用pHS-3C pH计测定土壤pH, 用酸式滴定管测定土壤全氮(TN)和土壤有机质(SOM), 火焰光度计测定速效钾(RAK), 有效磷(AP)用碳酸氢钠浸提-钼锑抗比色法测定。
1.3.2 土壤纤毛虫培养与鉴定取已风干土壤50g(约50/4000=1/80样品)于直径15cm的培养皿中, 恒温培养箱25℃下采用“非淹没培养皿法”培养土壤纤毛虫[23], 即在含有土壤样品的培养皿中加入土壤浸出液, 使土壤充分润湿而不被淹没。培养24 h后开始镜检(Leica DM500显微镜, 放大40—1000倍), 用“活体观察法”和“蛋白银染色法”[24], 根据形态特征和相应的参考资料[25—29]鉴定物种(Lynn(2008)分类系统[30]), 并记录所鉴定物种的个体数, 直至没有新的物种被发现为止。
1.4 数据处理和分析 1.4.1 多样性指数计算(1) 多样性指数(Shannon-Wiener)公式:

|
(2) 均匀度指数(Pieluo)公式:

|
(3) 优势度指数(Simpson)公式:
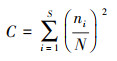
|
(4) 丰富度指数(Margalef)公式:
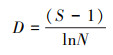
|
式中, H′表示物种多样性指数, ni为第i类群的个体数, N为总个体数, S为所有类群数。
1.4.2 C/P系数土壤纤毛虫r对策者(肾形纲)物种数和土壤纤毛虫k对策者(异毛纲、旋毛纲和瓶纤纲)物种数总和的比值。肾形纲的土壤纤毛虫一般生活较为恶劣的环境中, 因此C/P值的大小能够在一定程度上反映出该生境环境的优劣, 当C/P<1时, 表征该土壤生境中的环境相对较好;当C/P>1时, 表征该土壤生境中的环境较为恶劣[31]。
1.4.3 相似性指数计算用Jaccard相似性指数计算每个样地土壤纤毛虫群落特征的相似性。
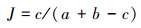
|
式中, c表示两个生境中共有的物种数, a和b表示两个不同样地中各自物种数。相似性指数在0—0.25之间表示极不相似;在0.25—0.5之间时表示中等不相似;在0.5—0.75之间时表示中等相似;在0.75—1之间时表示极相似。
1.4.4 数据统计与分析在Excel 2019中进行数据整理计算, 用SPSS 21.0对土壤纤毛虫与环境因子之间进行单因素方差分析(One-way ANOVA), 并进行Duncan多重比较, 运用R 4.0.5计算多样性指数和相似性系数[32—33], 用Origin 2019b作图;用Canoco 4.5 for Windows对物种和环境因子进行DCA和CCA分析。
2 结果与分析 2.1 年楚河流域湿地土壤纤毛虫群落结构年楚河流域5个湿地生境中共鉴定出土壤纤毛虫154种, 隶属于9纲、19目、43科、70属(表 1)。其中优势类群为前管目(Prorodontida), 共计18种, 占总物种数的11.69%;刺钩目(Haptorida)为次优势类群, 共计17种, 占总物种数的11.04%;罕见类群为游仆虫目(Euplotidae)、瓶纤目(Armophorida)、肾形目(Colpodida)、嗜污目(Philaslasterida)、齿管目(Chlamydodontida)和吸管目(Philaslasterida), 共计25种, 占总物种数的16.23%。SD1、SD2、SD3和SD4样地中旋毛纲(Spirotrichea)纤毛虫丰度最高, 其次为寡膜纲(Oligohymenophorea);SD5样地中肾形纲(Colpodea)纤毛虫丰度最高, 其次为旋毛纲(Spirotrichea), 图 2。
| 纲 Class |
目 Order |
科 Family |
属 Genus |
种 Species |
比例 Percentage/% |
| 核残基纲Karyorelictea | 原口目Protostomztida | 颈毛虫科Trachelocercea | 1 | 1 | 0.65% |
| 喙纤科Loxodidae | 1 | 2 | 1.30% | ||
| 异毛纲Heterotrichea | 异毛目Hererotrichida | 喇叭科Stentoridae | 1 | 1 | 0.65% |
| 赭虫科Blepharismidae | 1 | 2 | 1.30% | ||
| 突口科Condylostomidae | 1 | 1 | 0.65% | ||
| 旋毛虫科Spirostomidae | 1 | 2 | 1.30% | ||
| 旋毛纲Spirotrichea | 散毛目Sporadotrichia | 尖毛科Oxytrichidae | 6 | 13 | 8.44% |
| 急游科Strombidiidae | 1 | 1 | 0.65% | ||
| 弹跳科Halteriidae | 1 | 1 | 0.65% | ||
| 管柱科Trachelophyllidae | 1 | 1 | 0.65% | ||
| 尾柱虫目Urostylida | 尾柱虫科Urostylidae | 4 | 9 | 5.84% | |
| 侠盗科Strobilidiidae | 1 | 2 | 1.30% | ||
| 排毛目Sporadotrichida | 角毛科Keronidae | 1 | 1 | 0.65% | |
| 旋毛虫科Spirofilidae | 2 | 3 | 1.95% | ||
| 游仆虫目Euplotidae | 游仆虫科Euplotidae | 1 | 4 | 2.60% | |
| 盾纤目Scuticociliatidae | 楯纤科Aspidiscidae | 1 | 1 | 0.65% | |
| 瓶纤纲Armophorea | 瓶纤目Armophorida | 扭头科Metopidake | 1 | 2 | 1.30% |
| 裂口纲Litostomatea | 侧口目Pleurostomatida | 裂口科Amphileptidae | 4 | 9 | 5.84% |
| 漫游虫科Litonotidae | 1 | 3 | 1.95% | ||
| 刺钩目Haptorida | 刀口虫科Spathidiidae | 1 | 6 | 3.90% | |
| 圆口虫科Bilamellophrya | 1 | 5 | 3.25% | ||
| 管叶科Trachelophyllidae | 3 | 5 | 3.25% | ||
| 长吻虫科Lacrymariidae | 1 | 1 | 0.65% | ||
| 肾形纲Colpodea | 肾形目Colpodida | 肾形科Colpodidae | 1 | 7 | 4.55% |
| 篮环目Cyrtolophosidida | 篮环科Cyrtolophosidae | 1 | 4 | 2.60% | |
| 匙口科Platyophryidae | 1 | 1 | 0.65% | ||
| 前管纲Prostomatea | 前管目Prorodontida | 前管虫科Prorodontidae | 3 | 8 | 5.19% |
| 斜板科Plagiocampidae | 2 | 3 | 1.95% | ||
| 射纤科Actinoobolinidae | 1 | 1 | 0.65% | ||
| 裸口虫科Holophyryidae | 1 | 3 | 1.95% | ||
| 前口科Frontoniidae | 1 | 1 | 0.65% | ||
| 栉毛科Didiniidae | 2 | 2 | 1.30% | ||
| 寡膜纲Oligohymenophorea | 膜口目Hymenostomatida | 四膜科Tetrahymenidea | 4 | 5 | 3.25% |
| 瞬目科Glaucomidae | 2 | 6 | 3.90% | ||
| 固着目Sessilida | 钟虫科Vorticellidae | 1 | 12 | 7.79% | |
| 盖虫科Opercularia | 1 | 3 | 1.95% | ||
| 杯形科Scyphidiidae | 1 | 1 | 0.65% | ||
| 帆口目Pleuronematida | 帆口科Cyclidiidae | 3 | 8 | 5.19% | |
| 康纤科Cohnilemebidae | 1 | 1 | 0.65% | ||
| 斜头科Loxocephalidae | 1 | 1 | 0.65% | ||
| 嗜污目Philaslasterida | 映毛虫科Cinetochilidae | 1 | 2 | 1.30% | |
| 叶咽纲Phyllopharyngea | 齿管目Chlamydodontida | 斜管科Chilodonellidae | 2 | 5 | 3.25% |
| 吸管目Succorida | 足吸管科Podophryidae | 3 | 4 | 2.60% | |
| 优势类群是物种数最多的目, 次优势类群是物种数次多的目, 罕见类群是只有单一种的目 | |||||

|
| 图 2 年楚河流域湿地各样地土壤纤毛虫丰度 Fig. 2 Soil ciliate abundance of various wetland sites in NianChu River Basin, China |
对年楚河流域湿地土壤纤毛虫的多样性指数和物种数进行单因素方差分析, 结果表明多样性指数(H)(df=2, F=0.653, P=0.538), 均匀度指数(E)(df=2, F=1.878, P=0.195), 优势度指数(C)(df=2, F=1.263, P=0.318), 三个季节间均无显著性差异, 物种数(n)(df=2, F=4.264, P=0.023)在季节间具有显著性差异。SD4样地的多样性指数在5个样地中最高, SD1、SD4样地的各多样性指数(H、E、C)均是夏季>秋季>春季;而SD2、SD5样地的多样性指数(E、C)均为夏季>春季>秋季;SD3样地春季多样性指数最高;Margalef丰富度指数显示SD2、SD3、SD5样地均为秋季最高;优势度指数显示SD2和SD4样地间有差异性, 多样性指数显示SD4与SD2、SD3样地间有差异性, 均匀度指数和丰富度指数各样地间均没有差异。年楚河流域湿地土壤纤毛虫C/P(0.35) < 1, 说明年楚河流域湿地土壤环境未受污染。见图 3—6。

|
| 图 3 年楚河流域湿地土壤纤毛虫群落优势度指数 Fig. 3 Community dominance index of ciliates in wetland of the Nianchu River Basin ab: 不同的小写字母代表不同的显著水平 |

|
| 图 4 年楚河流域湿地土壤纤毛虫均匀度指数 Fig. 4 Evenness index of ciliates in wetland of the Nianchu River Basin |

|
| 图 5 年楚河流域湿地土壤纤毛虫多样性指数 Fig. 5 Diversity index of ciliates in wetland of the Nianchu River Basin |

|
| 图 6 年楚河流域湿地土壤纤毛虫多样性丰富度指数 Fig. 6 Margalef index of soil ciliates in wetland of the Nianchu River Basin |
春季不同湿地土壤纤毛虫群落相似性系数在0.13—0.21之间, 属于极不相似到中等不相似之间;夏季相似性系数范围为0.16—0.37, 属于极不相似到中等不相似之间;秋季相似性系数为0.13—0.26, 属于极不相似到中等不相似。
| 样地 Site |
春季Spring | 夏季Summer | 秋季Autumn | |||||||||||
| SD1 | SD2 | SD3 | SD4 | SD1 | SD2 | SD3 | SD4 | SD1 | SD2 | SD3 | SD4 | |||
| SD2 | 0.19 | 0.2 | 0.2 | |||||||||||
| SD3 | 0.21 | 0.2 | 0.3 | 0.37 | 0.2 | 0.2 | ||||||||
| SD4 | 0.17 | 0.1 | 0.16 | 0.3 | 0.29 | 0.28 | 0.2 | 0.3 | 0.26 | |||||
| SD5 | 0.17 | 0.2 | 0.16 | 0.17 | 0.2 | 0.16 | 0.22 | 0.2 | 0.2 | 0.1 | 0.23 | 0.25 | ||
对年楚河流域五个湿地样地进行聚类分析发现, SD2和SD3首先聚为一类, 然后和SD1聚为一类, 接着和SD4聚为一类, 最后和SD5聚为一类。说明SD5样地的纤毛虫群落组成空间异质性较大, 其次是SD4样地, 其中SD2和SD3两个样地的纤毛虫群落组成相似性程度最高。
2.4 年楚河流域湿地土壤理化性质湿地土壤理化性质变化会影响土壤纤毛虫的分布变化, 二者关系密切。单因素方差分析显示三个季节间, 土壤湿度(SWC)(df=2, F=0.150, P=0.863)、有效磷(AP)(df=2, F=2.768, P=0.103)、全氮(TN)(df=2, F=0, P=1)、土壤温度(ST)(df=2, F=0.647, P=0.541)、pH(df=2, F=0.438, P=0.656)和土壤有机质(SOM)(df=2, F=0.137, P=0.873)均没有显著性差异, 而速效钾(RAK)(df=2, F=4.916, P=0.028)具有显著性差异。多重比较(Duncan)结果显示, 五个样地间的土壤湿度(SWC)、有效磷(AP)和速效钾(RAK)没有显著差异(P>0.05), 其他理化因子均具有不同程度的差异。其中SD1的全氮(TN)、有效磷(AP)、速效钾(RAK)及有机质(SOM)含量明显高于其他样地;SD5样地全氮(TN)和有机质(SOM)含量最低;SD3有效磷(AP)、速效钾(RAK)含量最低。

|
| 图 7 年楚河流域湿地土壤纤毛虫聚类分析 Fig. 7 Cluster analysis of soil ciliates in wetland of the Nianchu River Basin |
| 样地 Site |
温度 ST/℃ |
土壤湿度 SWC/% |
pH | 全氮 TN(mg/kg) |
有效磷 AP(mg/kg) |
速效钾 RAK(mg/kg) |
有机质 SOM(g/kg) |
| SD1 | 8.48±2.52b | 46.18±1.68a | 6.97±0.061c | 8260.00±453.69a | 4.73±3.09a | 141.33±28.87a | 174.00±19.5a |
| SD2 | 14.1±1.90ab | 40.83±1.93a | 7.41±0.083b | 2460.00±106.93b | 1.4±0.41a | 76.67±5.67a | 35.13±9.11b |
| SD3 | 17.24±1.90ab | 38.8±0.66a | 7.41±0.079b | 1205.67±288.96c | 1.1±0.23a | 75.43±9.60a | 28.63±2.64b |
| SD4 | 11.65±2.80b | 37.45±0.53a | 7.36±0.017b | 1333.33±147.46c | 1.83±1.09a | 92.00±29.40a | 29.57±3.98b |
| SD5 | 21.99±3.61a | 25.22±7.17a | 7.67±0.053a | 546.67±101.94b | 1.17±0.20a | 110.67±44.02a | 11.29±4.10b |
| ST: 土壤温度soil temperature;SWC: 湿度soil water content;TN: 全氮total nitrogen;AP: 有效磷available phosphorus;RAK: 速效钾rapidly available potassium;SOM: 有机质soil organic matter;ab: 不同的小写字母代表不同的显著水平 | |||||||
春季年楚河流域土壤纤毛虫进行DCA前项选择, 发现排序轴轴长大于3, 可以使用单峰模型, 共筛选出三个环境因子土壤温度(ST), 湿度(SWC), 速效钾(RAK)进行CCA分析, 应用蒙特卡拟合方法对解释变量进行显著性检验。结果显示(图 8), 轴1和轴2共同解释了物种变异的60.15%, 说明轴1和轴2能够在较大程度上反映出年楚河流域湿地土壤纤毛虫群落与环境因子之间的关系。代表SWC的箭头长度最长, 表明SWC对于群落变化的影响最大, 样地SD1、SD2投影点与代表SWC的箭头距离最近, 说明SWC对该样地的影响最大;样地SD5投影点与代表ST的箭头距离最近, 说明ST对该样地的影响最强;样地SD3、SD4投影点与代表RAK的箭头距离最近, 说明RAK对该样地的影响最大。SWC与Cop、Kem、Coi等呈负相关关系, ST、RAK与Cop、Kem、Coi等呈正相关关系, 说明SWC、ST、RAK是影响春季年楚河流域湿地土壤原生动物群落分布的因素。

|
| 图 8 春季湿地土壤纤毛虫与环境因子的典范对应分析 Fig. 8 Canonical correspondence analysis between soil ciliates and environmental factors in spring wetland of the Nianchu River Basin Pln: 小匙口虫Plaiyophyrya nana;Tep: 梨形四膜虫Tetrahymena priformis;Stcr: 粗圆纤虫Strongylidium crassum;Sao: 椭圆嗜腐虫Sathrophilus ovatus;Coi: 膨胀肾形虫Colpoda inflata;Kem: 念珠角毛虫Keronopsis monilata;Saov: 卵形嗜腐虫Sathrophilus oviformis;Acr: 辐射射纤虫Actinobolina radians;Oca: 赫奕尖毛虫Oxytricha caudens;Cha: 非游斜管虫Chilodonella aplanata;Gof: 近亲殖口虫Gonostomum a ffina;Eup: 盘状游仆虫Euplotes patella;Coca: 弯豆形虫Colpidium campylum;Dic: 巨长颈虫Dileptus cygnus;Vco: 沟钟虫Vorticella convallaria;Lav: 蠕形长吻虫Lacrymaria vermicularis;Pll: 亮板纤虫Placus luciae;Stl: 矛形圆纤虫Strongylidium lanceolatum;Urf: 双叉尾毛虫Urotricha furcata;Lic: 天鹅漫游虫Litonotus cygnus;Acp: 乳头壳吸管虫Acineta papillifera;Stco: 天蓝喇叭虫Stentor coeruleus;Voc: 钟形钟虫Vorticella campanula;Sap: 污杇纤虫Saprodinium putrinium;Spc: 尾刀口虫Spathidium caudatum;Stg: 旋回侠盗虫Steobilidium gyrans;Cop: 盘状肾形虫Colpoda patella;Tap: 膜状急纤虫Tachysoma pellionella;Dia: 美洲长颈虫Dileptus americanus;Cht: 柱纤口虫Chaenea teres |
首先对夏季年楚河流域湿地土壤纤毛虫进行DCA前项选择, 发现排序轴轴长大于3, 可以使用单峰模型, 共筛选出三个环境因子有机质(SOM), 湿度(SWC), 速效钾(RAK)进行CCA分析, 应用蒙特卡拟合方法对环境因子显著性检验。结果显示(图 9), 排序轴1和排序轴2共同解释了纤毛虫群落方差的67.89%, 在一定程度上可以较好的反映出湿地土壤纤毛虫群落与环境因子的关系。速效钾(RAK)(F=0.012, P=0.036)是显著解释变量, 解释了纤毛虫群落变异的34.7%, 是影响年楚河流域湿地土壤纤毛虫群落分布的主要环境因子。代表RAK的箭头长度最长, 说明RAK对于群落变化的影响最大, 样地SD2、SD3投影点与其距离最近, 说明RAK对该样地的影响最大;样地SD1投影点与代表SOM的箭头, 说明SOM对该样地的影响最大;样地SD5与代表SWC的箭头距离最近, 说明SWC对该样地的影响最大。SWC、SOM、RAK与Eum、Cor和Dia等呈正相关关系, 与Hag、Asv和Stvi成呈负相关关系。说明SWC、SOM是影响湿地土壤纤毛虫分布的因素。

|
| 图 9 夏季湿地土壤纤毛虫与环境因子的典范对应分析 Fig. 9 Canonical correspondence analysis between soil ciliates and environmental factors in summer wetland of the Nianchu River Basin Hag: 大弹跳虫Halteria grandinella;Asv: 团睥睨虫Askenasia volvox;Stvi: 绿急游虫Strombidium viride;Stn: 背状棘尾虫Stylonychia notophora;Spu: 苔藓刀口虫Spathidium muscorum;Cor: 肾形肾形虫Colpoda reniformis;Eum: 黏游仆虫Euplotes muscicola;Stg: 旋回侠盗虫Steobilidium gyrans;Paw: 柔似尾枝虫Paraurostyla weisei;Vocu: 杯钟虫Vorticella cupifera;Voae: 领钟虫Voticella aequilata;Pom: 胶衣足吸管虫Podophrya maupasi;Dia: 美洲长颈虫Dileptus americanus;Gls: 闪瞬目虫Glaucoma scintillans;Oca: 赫奕尖毛虫Oxytricha caudens;Oxc: 叶绿尖毛虫Oxytricha chlorelligera;Glf: 前口瞬目虫Glaucoma frontata;Tap: 膜状急纤虫Tachysoma pellionella;Hosu: 沟裸口虫Holophrya sulcata;Spsp: 刀刀口虫Spathidium spathula;Hoa: 腔裸口虫Holophrya atra;Lou: 单核斜叶虫Loxophyllum uninucleatum;Hepu: 点滴半眉虫Hemiophrys punctata;Spc: 尾刀口虫Spathidium caudatum;Cop: 盘状肾形虫Colpoda patella;Urd: 差异瘦尾虫Uroleptus dispar;Urci: 契氏片尾虫Urosoma cienkowskii;Blsa: 半咸赭虫Blepharisma salinarum;Glm: 大口瞬目虫Glaucoma macrostoma;Cyma: 大蓝环虫Cyrtilophosis major |
首先对秋季年楚河流域湿地土壤纤毛虫进行DCA前项选择, 发现排序轴轴长大于3, 可以使用单峰模型, 共筛选出三个环境因子湿度(SWC), 土壤温度(ST), 速效钾(RAK)进行CCA分析, 应用蒙特卡拟合方法对环境因子显著性检验。结果如图 10所示, 轴1和轴2共同解释率物种变异的72.60%, 说明轴1和轴2能够较大程度上反映出秋季年楚河流域湿地土壤纤毛虫群落与环境因子之间的关系。代表ST的箭头长度最长, 表明ST对于群落变化的影响最大, 且样地SD1投影点与其距离最近, 说明ST对该样地的影响最大;样地SD2、SD3投影点与代表SWC的箭头距离最近, 说明SWC对该样地的影响最大;样地SD5与带包RAK的箭头距离最近, 说明RAK对该样地的影响最大。SWC与Cht、Urn、Lic等呈负相关关系, ST、RAK与Cyc、Sao、Urd等呈正相关关系, 说明SWC、ST、RAK是影响其分布的因素。

|
| 图 10 秋季湿地土壤纤毛虫与环境因子的典范对应分析 Fig. 10 Canonical correspondence analysis between soil ciliates and environmental factors in autumn wetland of the Nianchu River Basin Coc: 僧帽肾形虫Colpoda cucullus;Cyc: 居中膜袋虫Cyclidium centrale;Chm: 苔藓拟斜管虫Chilodontopsis muscorum;Mee: 如意扭头虫Metopus es;Cymu: 黏液蓝环虫Cyrtilophosis mucicola;Urd: 差异瘦尾虫Uroleptus dispar;Sao: 椭圆嗜腐虫Sathrophilus ovatus;His: 似织毛虫Histriculus similis;Din: 小单环栉毛虫Didinium balbianii nanum;Glf: 前口瞬目虫Glaucoma frontata;Dia: 美洲长颈虫Dileptus americanus;Cht: 柱纤口虫Chaenea teres;Epu: 瓶累枝虫Epistylis urceolata;Urn: 暗尾丝虫Uronema nigrican;Vopi: 点钟虫Vorticella picta;Pls: 游荡平体虫Platynematum solivagum;Lic: 天鹅漫游虫Litonotus cygnus;Sps: 浮雕刀口虫Spathidium scalpriforme;Fra: 尖锐前口虫Frontonia acuminata;Bls: 单核赭虫Blepharisma steini;Oxsa: 腐生尖毛虫Oxytricha saprobia;Lou: 单核斜叶虫Loxophyllum uninucleatum;Oxf: 伪尖毛虫Oxytricha fallax;Oxc: 叶绿尖毛虫Oxytricha chlorelligera;Psmo: 念珠伪钟虫Pseudovorticella moninata;Paw: 柔似尾枝虫Paraurostyla weisei;Stc: 弯棘尾虫Stylonychia curvata;Pom: 胶衣足吸管虫Podophrya maupasi;Trt: 细领颈毛虫Trachelocerca tenuicollis;Hea: 敏捷半眉虫Hemiophrys agilis |
湿地生态系统兼有陆地生态系统和水生态系统的某些特征, 是二氧化碳的“汇”, 在全球气候变化上具有重要的意义[2]。优势类群能够明显的控制土壤纤毛虫群落结构和群落环境, 是土壤纤毛虫群落基本特征的其中一部分[22]。年楚河湿地土壤纤毛虫优势类群为前管目, 次优势类群为刺钩目, 两者是年楚河流域湿地土壤纤毛虫最重要的组成部分, 对于湿地生态系统有着不可忽视的重要作用[34]。土壤纤毛虫是土壤中细菌和真菌的采食者, 是土壤中微生物循环系统的重要组成部分, 能够很好的促进微生物的生长, 促进C和N对较高营养水平的有效性[35], 并且其物种组成和种群密度会随着环境变化而变化, 是监测环境变化的有效手段。年楚河流域湿地土壤纤毛虫多样性指数(H、E、C)季节间均无显著性差异(P>0.05), SD4样地多样性指数在5个样地中均最高, 原因可能是SD4样地的海拔相对较低, 生存环境较为优渥, 物种丰富, 群落结构复杂[22, 36]。SD3多样性指数春季最高, 可能是因为春季温度开始升高, 土壤纤毛虫物种数增加, 使土壤纤毛虫的多样性指数升高。春季不同湿地土壤纤毛虫群落相似性系数在0.13—0.21之间, 属于极不相似到中等不相似之间;夏季相似性系数范围为0.16—0.37, 属于极不相似到中等不相似之间;秋季相似性系数为0.13—0.26, 属于极不相似到中等不相似, 其中夏季SD2和SD3样地的相似性程度最高为0.37。对年楚河流域五个湿地样地进行聚类分析发现, SD5样地和其他4个样地聚为一类, 说明SD5样地空间差异性较大, 原因可能是SD5样地的ST和RAK明显高于其他4个样地, 导致SD5样地土壤纤毛虫物种组成出现差异, 在空间上的相似性程度低。
3.2 年楚河流域湿地土壤纤毛虫群落影响因素SD1的全氮(TN)、有效磷(AP)、速效钾(RAK)及有机质(SOM)含量明显高于其他样地。该样地靠近牧区, 牛羊等畜牧的粪便, 可能导致该样地有机质等理化因子明显高于其他样地。年楚河流域湿地各样地SWC、RAK和SOM分布在轴1附近, 与轴1具有最大的相关性, ST分布在轴2附近, 与轴2具有最大的相关性, ST、SWC、RAK、SOM是影响年楚河流域湿地土壤纤毛虫群落分布的主要因素, 且SWC和RAK能够显著解释土壤纤毛虫物种分布。造成纤毛虫物种分布的主要原因是温度和降水, 最佳土壤温度适宜土壤动物生存, 土壤温度调节土壤纤毛虫的取食和代谢活动[37], Opperman等研究发现土壤温度升高会导致土壤纤毛虫数量明显增加, 群落结构发生改变[38], 土壤水分是限制土壤纤毛虫分布、生存和繁殖的主要环境因子[39], 冯伟松等研究南极菲尔德斯地区土壤原生动物时发现, 土壤原生动物密度与土壤含水量正相关[9];土壤动物能够通过生命活动在氮、磷元素循环中起着重要作用, 土壤动物的生物量与土壤中氮、磷的含量正相关[40], Patrick研究发现土壤原生动物在有机质丰富的土壤中丰度更高, 同等条件下添加粪浆的土壤中土壤原生动物总量明显更高[41], 且土壤中有机质的微小变化影响土壤纤毛虫物种分布, 土壤理化因子决定了土壤纤毛虫物种分布[42]。综上年楚河流域湿地土壤纤毛虫群落受多个环境因子的共同影响, 每种环境因子都对土壤纤毛虫群落特征产生不同的影响, 其共同作用影响土壤纤毛虫群落在不同样地间的分布。年楚河流域湿地土壤纤毛虫C/P(0.35) < 1, 说明年楚河流域湿地土壤环境未污染, 适宜土壤动物生存, 对于维持和保护中国生态屏障(青藏高原)生态环境良好发展具有重要作用, 因为肾形纲土壤纤毛虫为R-对策, 该对策土壤纤毛虫往往生活在土壤质量较差的环境中, 旋毛纲、异毛纲和瓶纤纲土壤纤毛虫为K-对策, 该对策土壤纤毛虫往往生活在土壤质量较好的环境中, C/P值越大, 即R-对策肾形纲土壤纤毛虫在土壤环境中较K-对策土壤纤毛虫更具优势, 生态环境质量较差。
4 结论年楚河流域5个湿地样地共鉴定出土壤纤毛虫154种, 隶属于9纲、19目、43科、70属;优势类群是前管目(Prorodontida), 共计18种, 占总物种数的11.69%;多样性指数(H、E、C)在季节间均无显著性差异(P>0.05), 且SD4样地多样性指数最高;年楚河流域湿地土壤纤毛虫C/P(0.35) < 1, 说明年楚河流域湿地土壤环境未污染;5个样地土壤纤毛虫相似程度不高, 物种空间分布差异性大;SWC、ST和RAK是年楚河流域湿地土壤纤毛虫的影响因子。
| [1] |
朱永官, 彭静静, 韦中, 沈其荣, 张福锁. 土壤微生物组与土壤健康. 中国科学: 生命科学, 2021, 51(1): 1-11. |
| [2] |
吕宪国. 湿地科学研究进展及研究方向. 中国科学院院刊, 2002, 17(3): 170-172. DOI:10.3969/j.issn.1000-3045.2002.03.004 |
| [3] |
陈素芳, 徐润林. 土壤原生动物的研究进展. 中山大学学报: 自然科学版, 2003, 42(S1): 187-194. |
| [4] |
宁应之, 万贯红, 杨元罡, 武维宁, 周小燕, 王一峰, 刘娜, 程晓, 陈凌云. 甘肃省徽县不同退耕还林模式下土壤纤毛虫群落特征. 生态学杂志, 2019, 38(6): 1697-1706. |
| [5] |
宁应之, 周小燕, 烟郑杰, 张世虎, 陈凌云, 万贯红. 土壤纤毛虫群落对退牧还草生态恢复的响应——以玛曲县为例. 生态学报, 2020, 40(7): 2386-2395. |
| [6] |
禹娟红, 包振国, 王秉忠, 刘玲玲, 车树理. 不同植被恢复模式对土壤纤毛虫群落特征的影响. 林业科技通讯, 2017(4): 3-7. |
| [7] |
宁应之, 杨永强, 董玟含, 张惠茹, 马继阳. 土壤纤毛虫群落对不同退还模式生态恢复的响应. 生态学报, 2018, 38(10): 3628-3638. |
| [8] |
宁应之, 武维宁, 杜海峰, 王红军. 土壤纤毛虫群落对退耕还林生态恢复的响应——以侧柏林为例. 生态学报, 2016, 36(2): 288-297. |
| [9] |
宁应之, 王婷婷, 武维宁, 陈凌云, 杜国祯. 甘南高寒草甸土壤纤毛虫对磷酸氢二铵添加的响应. 应用生态学报, 2017, 28(5): 1668-1678. |
| [10] |
刘旻霞, 李全弟, 蒋晓轩, 夏素娟, 南笑宁. 兰州市北山不同林地春夏季土壤纤毛虫群落特征. 生态学报, 2020, 40(12): 3955-3967. |
| [11] |
徐润林, 孙逸湘. 大鹏半岛土壤纤毛虫的群落特点. 应用生态学报, 2000, 11(3): 428-430. DOI:10.3321/j.issn:1001-9332.2000.03.025 |
| [12] |
冯伟松, 余育和. 南极菲尔德斯半岛地区土壤原生动物生态学研究. 水生生物学报, 2000, 24(6): 610-615. DOI:10.3321/j.issn:1000-3207.2000.06.006 |
| [13] |
王超, 徐润林. 鼎湖山不同森林类型土壤纤毛虫群落比较研究. 土壤, 2017, 49(4): 725-732. |
| [14] |
张灿, 刘旻霞, 李瑞, 赵瑞东, 邵鹏. 甘南亚高寒草甸冷暖季土壤纤毛虫群落对坡向的响应. 生态学杂志, 2017, 36(9): 2465-2472. |
| [15] |
刘旻霞, 张灿, 李瑞, 赵瑞东, 邵鹏. 不同坡向对高寒草甸秋冬季土壤纤毛虫群落的影响. 兰州大学学报: 自然科学版, 2019, 55(3): 380-387, 394-394. |
| [16] |
万玛央措, 徐富荣, 杨怀印, 沈小亚, 刘浩, 武维宁, 陈鹏, 刘娜, 陈凌云, 宁应之. 甘肃农田四种土壤腹毛类纤毛虫(纤毛门, 旋毛纲, 腹毛亚纲)的形态学研究. 动物学杂志, 2021, 56(4): 543-557. |
| [17] |
普布, 袁文军. 西藏双湖地区土壤纤毛虫群落特征. 高原科学研究, 2019(4): 12-20. |
| [18] |
巴桑, 黄香, 普布, 马正学. 拉鲁湿地夏秋季纤毛虫群落特征及其与水环境的关系. 生态学杂志, 2014, 33(10): 2740-2748. |
| [19] |
刘务林, 朱雪林. 中国西藏高原湿地. 北京: 中国林业出版社, 2013.
|
| [20] |
刘天仇, 陈爱兰. 年楚河流域降水特性. 成都科技大学学报, 1995(4): 12-18. |
| [21] |
王秀丽, 刘石臣. 西藏年楚河流域水资源调查. 东北水利水电, 2000, 18(9): 22-23. |
| [22] |
王壮壮, 贺凯, 朱时应, 普布. 西藏年楚河流域湿地大型土壤动物群落特征. 干旱区资源与环境, 2021, 35(11): 184-190. |
| [23] |
Foissner W. Estimating the species richness of soil protozoa using the "non-flooded petri dish method"//Lee J J, Soldo A T, eds. Protocols in Protozoology. Lawrence: Allen Press, 1922.
|
| [24] |
Wilbert N. Eine verbesserte technik der protargolimpragnation für ciliaten. Mikrokosmos, 1975, 64: 171-179. |
| [25] |
吕立平, 樊阳波, 胡晓钟. 十种旋唇纲纤毛虫的形态学描述. 水生生物学报, 2017, 41(5): 1126-1139. |
| [26] |
张琦. 十种土壤腹毛类纤毛虫的分类研究[D]. 保定: 河北大学, 2021.
|
| [27] |
宁应之, 马继阳, 吕昭. 六种土壤腹毛类纤毛虫的形态学研究. 动物学杂志, 2018, 53(3): 415-426. |
| [28] |
宁应之, 孙静, 王红军, 杜海峰, 刘汉成, 刘光龙. 甘肃甘南高原沼泽湿地6种土壤纤毛虫记述. 动物学杂志, 2014, 49(2): 261-271. |
| [29] |
宁应之, 沈韫芬. 中国典型地带29种土壤纤毛虫记述(纤毛门: 动基片纲). 西北师范大学学报: 自然科学版, 1999, 35(3): 75-82. |
| [30] |
Lynn D H. The Ciliated Protozoa: Characterization, Classification, and Guide to the Literature. 3rd ed. Dordrecht: Springer, 2008: 1-605.
|
| [31] |
Foissner W, Berger H, Xu K, Zechmeister-Boltenstern S. A huge, undescribed soil ciliate (Protozoa: Ciliophora) diversity in natural forest stands of Central Europe. Biodiversity & Conservation, 2005, 14(3): 617-701. |
| [32] |
马克平, 刘玉明. 生物群落多样性的测度方法: Ⅰα多样性的测度方法(下). 生物多样性, 1994, 2(4): 231-239. |
| [33] |
Pielou E C. Ecological Diversity. Hoboken: John Wiley & Sons, 1975.
|
| [34] |
王壮壮, 刘洋, 贺凯, 樊海德, 普布. 西藏年楚河流域大型土壤动物群落特征与生态位. 生态学杂志, 2021, 40(12): 3911-3921. |
| [35] |
Brussaard L, Behan-Pelletier V M, Bignell D E, Brown V K, Didden W, Folgarait P, Fragoso C, Freckman D W, Gupta V V S R, Hattori's T, Hawkworth D L, Klopatek C, Lavelle P, Malloch D W, Rusek J, Sderstrom B, Tiedje J S, Virginia R A, 闵庆文. 土壤中的生物多样性与生态系统功能. 人类环境杂志, 1997, 26(8): 555-562. |
| [36] |
王壮壮, 贺凯, 朱时应, 普布. 年楚河流域康马县高寒草地土壤节肢动物多样性研究. 湖南师范大学自然科学学报, 2021, 44(6): 76-84. |
| [37] |
邹涛, 申海香, 宁应之, 马正学. 甘肃小陇山麻沿林区土壤纤毛虫群落特征. 动物学杂志, 2009, 44(6): 64-73. |
| [38] |
Opperman M H, Wood M, Harris P J. Changes in microbial populations following the application of cattle slurry to soil at two temperatures. Soil Biology and Biochemistry, 1989, 21(2): 263-268. |
| [39] |
Schlesinger W H. Biogeochemistry: An Analysis of Global Change. 2nd ed. San Diego: Academic Press, 1997.
|
| [40] |
张雪萍, 李春艳, 殷秀琴, 陈鹏. 不同使用方式林地的土壤动物与土壤营养元素的关系. 应用与环境生物学报, 1999, 5(1): 26-31. |
| [41] |
Jjemba P K. The effect of chloroquine, quinacrine, and metronidazole on both soybean plants and soil microbiota. Chemosphere, 2002, 46(7): 1019-1025. |
| [42] |
王壮壮, 朱时应, 张秋涵, 何梦萨, 普布. 亏祖山秋季不同海拔土壤纤毛虫优势种生态位及其与环境因子的关系. 生态学报, 2022, 42(9): 3494-3503. |