 2022, Vol. 42
2022, Vol. 42文章信息
- 张钦弟, 刘剑荣, 杨磊, 卫伟
- ZHANG Qindi, LIU Jianrong, YANG Lei, WEI Wei
- 半干旱黄土区植被恢复对土壤团聚体稳定性及抗侵蚀能力的影响
- Effect of vegetation restoration on stability and erosion resistance of soil aggregates in semi-arid loess region
- 生态学报. 2022, 42(22): 9057-9068
- Acta Ecologica Sinica. 2022, 42(22): 9057-9068
- http://dx.doi.org/10.5846/stxb202202090324
-
文章历史
- 收稿日期: 2022-02-09
- 网络出版日期: 2022-07-12





2. 中国科学院生态环境研究中心 城市与区域生态国家重点实验室, 北京 100085
2. State Key Laboratory of Urban and Regional Ecology, Research Center for Eco-Environmental Sciences, Chinese Academy of Sciences, Beijing 100085, China
黄土高原因土壤的易侵蚀性、典型的季风性气候以及人类活动, 还有其特有的侵蚀沟壑地貌等因素造成其生态脆弱, 土壤侵蚀严重, 成为中国乃至全球土壤受到侵蚀最严重的地区之一[1]。解决水土流失问题是全球人类生存与发展的环境问题, 经受国家政策引导, 通过在黄土高原进行大规模的退耕还林(草)活动, 有效的改善黄土高原生态失衡、侵蚀剧烈的局面[2]。黄土区生态得到改善, 逐渐增加的植被覆盖度, 使其生态环境得以提升, 同时流域径流量和输沙量也逐年减少[3]。
土壤团聚体是土壤各级粒径通过各种物理、化学以及生物作用形成的直径为10—0.25 mm的结构性单位[4], 在提高土壤抗蚀能力方面具有重要作用[5]。土壤团聚体稳定性越好就能充分协调土壤水、气和养分等因素的循环, 为植被生长创造条件[6]。土壤的水稳定性团聚体可以更好的反映土壤的结构及稳定性[7—8], 利用土壤团聚体稳定性及粒径组成能间接性的比较土壤的抗侵蚀能力[9]。关于土壤团聚体稳定性的影响因素方面已经有大量研究, 土壤各级粒径的团聚度以及稳定性是决定和影响土壤可侵蚀性的重要物理性质之一[10];土壤有机碳能将直径较小的土壤颗粒粘结成较大的土壤颗粒, 而其含量能促进土壤团聚过程及稳定性[11—12]。土壤容重增加会对土壤团聚体稳定性产生不利影响, 从而降低土壤的抗蚀能力[13]。土壤团聚体的稳定性还会受到其他因素比如成土母质的类型以及立地因子等因素的影响, 因而研究结果不尽相同[14]。同样, 植被恢复也会对土壤团聚体稳定性产生一定的影响, 随着退耕地恢复年限的延长, 0—60 cm土层粒径直径大于0.25 mm土壤团聚体的含量而不断增加[15];在植被自然恢复过程中, 土壤团聚体结构会随着植被群落演替逐渐高级而趋于稳定[16—17]。
目前, 关于半干旱黄土区不同植被恢复方式之间土壤团聚体的组成及稳定性与土壤可蚀性之间的关系研究相对较少, 无法满足提升该地区植被恢复和土壤保持服务功能的需要。研究不同植被恢复方式土壤团聚体组成和土壤团聚体稳定性与土壤抗侵蚀能力的关系可以调节该地区土壤抗侵蚀能力, 对提升土壤服务功能具有重要价值。本研究选取甘肃定西龙滩流域3种恢复方式(天然荒草、自然恢复、人工恢复)7种植被类型(大针茅草地、长芒草草地、赖草草地、苜蓿草地、柠条灌丛、山杏林、油松林)为研究对象, 探究植被恢复对土壤团聚体稳定性及抗侵蚀能力影响, 有助于掌握该地区土壤团聚体稳定性和土壤可蚀性的规律, 为当地选择合适的植被恢复策略进行生态恢复工作提供参考依据。
1 研究区概况龙滩流域(104°27′—104°32′ E, 35°43′—35°46′ N)位于甘肃省定西市安定区巉口镇(图 1), 海拔1929—2211 m, 年均气温6.8℃, 年均降水量386 mm, 属于典型黄土丘陵沟壑区[18]。该区土质以结构疏松、含沙量高、肥力低的黄绵土为主, 属于典型草原地带[19]。天然植被多以长芒草草地(Stipa bungeana)、大针茅草地(Stipa grandis)为主, 经大规模退耕还林(草)的实施, 现有人工植被多以苜蓿草地(Medicago sativa)、柠条灌丛(Caragana korshinskii)、山杏林(Armeniaca sibirica)、油松林(Pinus tabulaeformis)等为主。近年来, 由于农村人口流失严重, 部分农田撂荒闲置, 自然恢复植被以赖草草地(Leymus secalinus)为主。

|
| 图 1 研究区地理位置及实验样点 Fig. 1 Location of the study area and experimental sites |
本研究以代表性和典型性原则, 选取大针茅草地、长芒草草地、赖草草地、苜蓿草地、柠条灌丛、山杏林及油松林7种典型植被类型共39个样地进行研究。其中, 大针茅草地和长芒草草地作为该研究区代表性植被, 未经种植、耕种等人为干扰活动, 即天然荒草地;赖草草地为农地休耕后自然恢复植被;苜蓿草地、柠条灌丛、山杏林及油松林均为人工恢复植被。样地数量及基本特征见表 1。
| 恢复方式 Restoration methods |
植被类型 Vegetation type |
样本数 Sample size |
恢复年限 Restoration age |
海拔 Elevation/m |
坡向 Aspect/(°) |
坡度 Slope/(°) |
盖度 Coverage/% |
| 天然荒草 Natural grassland |
大针茅草地 Stipa grandis grassland |
7 | — | 2053.31±36.42 | 245±86.31 | 18±11.42 | 0.53±0.11 |
| 长芒草草地 Stipa bungeana grassland |
7 | — | 2076.31±52.65 | 203±128.43 | 17±10.45 | 0.45±0.03 | |
| 自然恢复 Natural restoration |
赖草草地 Leymus secalinus grassland |
5 | 15 | 2076.26±83.16 | 266±65.11 | 13±6.88 | 0.75±0.09 |
| 人工恢复 Artificial restoration |
苜蓿草地 Medicago sativa grassland |
10 | 15 | 2033.38±46.27 | 151±134.90 | 16±7.80 | 0.86±0.21 |
| 柠条灌丛 Caragana korshinskii shrub |
4 | 30 | 2087.53±34.82 | 80±8.33 | 29±6.38 | 0.47±0.13 | |
| 山杏林 Armeniaca sibirica forest |
3 | 47 | 2072.50±75.50 | 236±73.15 | 21±5.03 | 0.49±0.05 | |
| 油松林 Pinus tabulaeformis forest |
3 | 47 | 1977.77±17.90 | 342±16.92 | 14±0.58 | 0.44±0.10 | |
| 坡向计量标准是以正北方向为起点, 按顺时针方向计量 | |||||||
本研究于2021年5月进行土壤样本采集, 每个样地选两处相近的位置分别取0—60 cm土层, 以20 cm为间隔分层取样。其中一处位置采用环刀法进行取土样用于土壤理化性质的测定, 环刀体积为100 cm3, 共117份土样;另一处位置进行土壤水稳性团聚体的测定, 选用规格为12 cm×10 cm×8 cm的铝盒采集原状土, 共117份土样, 先除去土壤表层的枯枝落叶、动物尸体以及石块等干扰物质, 再分层取样后带回实验室, 依据土壤的自然结构轻轻剥剥成直径为1 cm左右的小土块, 剔除其中植物的粗根与小石块, 平摊在通风处自然风干待用。
2.2.2 土壤理化性质测定土壤有机碳采用重铬酸钾外加热氧化法测定[20];依据国际制土壤质地分级标准, 采用Mastersizer 2000激光颗粒测试仪测定土壤的粒径组成[21]。
2.2.3 土壤水稳性团聚体测定将自然风干好的各层土样(200 g)分别过孔径依此为5、2、1、0.5、0.25 mm的套筛, 将筛分后的土样按照径级分别装入塑封带中, 分别称取重量后备用。将干筛后得到的各级土壤粒径按质量比例配成50 g土样, 采用湿筛法[22]获得>5 mm、5—2 mm、2—1 mm、1—0.5 mm、0.5—0.25 mm、< 0.25 mm6个级别的水稳性土壤团聚体含量, 并通过计算获取各粒级土壤水稳性质量百分比[23]。
2.3 土壤水稳性团聚体平均重量直径土壤水稳性团聚体平均重量直径的计算公式如下[23]:
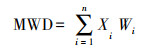
|
式中, WMD为平均重量直径;Xi为每粒级团聚体的平均直径(mm);Wi为第Xi级的团聚体的重量百分比。土壤水稳性团聚体稳定性的划分[24]: 土壤水稳性团聚体平均重量直径>2.0 mm代表团聚体非常稳定;2.0—1.3 mm代表团聚体稳定;1.3—0.8 mm代表团聚体中度稳定;0.8—0.4 mm代表团聚体不稳定;0—0.4 mm代表团聚体非常不稳定。
2.4 土壤团聚体破坏率土壤团聚体破坏率的计算公式如下[25]:
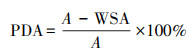
|
式中, PAD为土壤团聚体破坏率;WSA为>0.25 mm水稳性团聚体的含量;A为>0.25 mm干筛稳定性团聚体的含量。
2.5 土壤可蚀性因子先依据Williams等[26]建立了EPIC模型计算估算土壤可蚀性KEPIC值, 计算公式为:

|
式中, KEPIC为土壤可蚀性因子;CLA为粘粒含量;SIL为粉粒含量;SAN为砂粒含量;C为有机碳含量;SN1=1-SAN/100。
再基于张科利等[27]建立了适合用于我国土壤侵蚀预测模型, 计算土壤可蚀性因子:
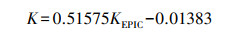
|
式中, K为土壤可蚀性因子, 单位是美国制, 计算后将K值乘以0.1317, 转化为国际制单位t hm2 h MJ-1mm-1hm-2。土壤可蚀性因子K值可以反映土壤抗水蚀的能力大小, K值越大, 土壤抗侵蚀能力越强, 反之, 土壤抗侵蚀能力越强。
2.6 数据分析在SPSS Statistics 17.0采用单因素方差分析法(one-way ANOVA)对同一因素不同水平间进行显著性差异分析, 相关数据采用平均值±标准差(Mean±SD)表示;用Pearson法(α=0.05)对土壤水稳性大团聚体、土壤水稳性平均重量直径、土壤团聚体破坏率及土壤可蚀性因子K值间进行相关性分析;采用Origin 2016进行数据绘图。
3 结果与分析 3.1 不同植被类型土壤水稳性团聚体粒径组成及稳定性分析0—20 cm土层赖草、长茅草和大针茅的土壤水稳性团聚体粒径>5 mm的质量百分比最大, 分别为22.80%、19.23%、15.22%, 山杏和油松最低, 分别为12.65%和9.01%。苜蓿的土壤水稳性团聚体粒径在5—2 mm、2—1 mm、1—0.5 mm、0.5—0.25 mm的质量百分比均显著高于其他植被类型(P < 0.05), 大针茅、长芒草、赖草的团聚体质量百分比随粒径减小总体变化趋势为先增加后减少, 而人工恢复植被苜蓿、柠条、山杏以及油松表现为随粒径减小团聚体质量百分比逐渐增大。20—40 cm土层各植被类型团聚体质量百分比随粒径减小总体变化趋势与0—20 cm土层基本相同。40—60 cm土层各植被类型团聚体质量百分比随粒径减小总体变化趋势均为逐渐增大, 其中, 大针茅和赖草粒径>5 mm时, 团聚体质量百分比显著高于其他植被(P < 0.05), 人工恢复植被苜蓿、柠条、山杏以及油松粒径 < 0.25 mm时, 团聚体重量百分比显著高于其他植被(P < 0.05)(图 2)。

|
| 图 2 不同植被类型不同土层各粒径团聚体质量百分比 Fig. 2 Mass percentage of aggregates with different grain sizes under different soil layers of different vegetation types 不同小写字母表示同一土层和粒级不同植被类型间差异显著(P < 0.05) |
所研究的0—60 cm土层中, 大团聚体(粒径直径>0.25 mm)质量百分比基本在67.97%—90.12%之间(表 2)。0—20 cm土层各植被类型的平均重量直径表现为赖草最大, 油松最小, 表明赖草草地团聚体非常稳定, 油松林团聚体非常不稳定。20—40 cm土层苜蓿、柠条、山杏以及油松的团聚体稳定性均表现为不稳定。40—60 cm土层赖草平均重量直径最大, 团聚体稳定性为稳定, 大针茅、长茅草、山杏的团聚体稳定性为不稳定, 苜蓿、柠条、油松三者之间差异不显著, 团聚体均表现为非常不稳定。从恢复方式整体来看(表 3), 0—60 cm土层平均重量直径均表现为自然恢复方式显著高于天然荒草和人工恢复方式(P < 0.05);人工恢复方式的大团聚体质量百分比在40—60 cm土层显著低于其他方式, 在0—20 cm和40—60 cm与天然荒草无显著差别, 但都显著低于自然恢复方式(P < 0.05)。
| 植被类型 Vegetation type |
土壤深度 Soil depth/cm | |||||||
| 0—20 | 20—40 | 40—60 | ||||||
| 平均重量直径 WMD/mm |
大团聚体 质量百分比WSA/% |
平均重量直径 WMD/mm |
大团聚体 质量百分比WSA/% |
平均重量直径 WMD/mm |
大团聚体 质量百分比WSA/% |
|||
| 大针茅草地 S.grandis grassland | 1.95±0.64a | 80.22±5.64ab | 1.11±0.26a | 82.33±4.68a | 0.71±0.09a | 76.69±6.49a | ||
| 长芒草草地 S.bungeana grassland | 2.04±0.57a | 83.61±3.57a | 0.96±0.03a | 84.26±6.32a | 0.78±0.04a | 75.58±1.69ab | ||
| 赖草草地 L.secalinus grassland | 2.66±0.89a | 90.12±7.36a | 1.92±0.73a | 89.61±3.65a | 1.39±0.26a | 81.64±8.36a | ||
| 苜蓿草地 M.sativa grassland | 0.84±0.08b | 85.66±4.27a | 0.50±0.17b | 82.64±2.68ab | 0.38±0.09b | 74.13±5.19ab | ||
| 柠条灌丛 C.korshinskii shrub | 1.28±0.18a | 81.69±7.77a | 0.60±0.23a | 80.45±5.32ab | 0.30±0.63b | 73.98±3.68ab | ||
| 山杏林 A.sibirica forest | 0.67±0.13b | 82.89±7.64a | 0.56±0.66b | 79.45±6.58ab | 0.46±0.09b | 74.59±7.69ab | ||
| 油松林 P.tabulaeformis forest | 0.30±0.02b | 79.31±7.98b | 0.44±0.83b | 77.89±4.69b | 0.39±0.07b | 67.97±8.87ab | ||
| 不同小写字母表示同一土层不同植被类型间的差异显著(P < 0.05)。WMD: 平均重量直径, Mean weight diameter;WSA: 大团聚体质量百分比, Water stable aggregate | ||||||||
| 恢复方式 Restoration methods |
土壤深度 Soil depth/cm | |||||||
| 0—20 | 20—40 | 40—60 | ||||||
| 平均重量直径 WMD/mm |
大团聚体质量百分比 WSA/% |
平均重量直径 WMD/mm |
大团聚体质量百分比 WSA/% |
平均重量直径 WMD/mm |
大团聚体质量百分比 WSA/% |
|||
| 天然荒草 Natural grassland | 2.00±0.36b | 81.92±6.95b | 1.04±0.11b | 83.30±6.22b | 0.75±0.06b | 76.14±5.32b | ||
| 自然恢复 Natural restoration | 2.66±0.89a | 90.12±7.36a | 1.92±0.73a | 89.61±3.65a | 1.39±0.26a | 81.64±8.36a | ||
| 人工恢复 Artificial restoration | 0.77±0.64c | 82.39±9.33b | 0.53±0.02b | 80.11±4.69b | 0.38±0.13b | 70.67±6.32c | ||
| 不同小写字母表示同一土层不同恢复方式间的差异显著(P < 0.05)。WMD: 平均重量直径, Mean weight diameter;WSA: 大团聚体质量百分比, Water stable aggregate | ||||||||
随土壤深度增加, 各植被类型土壤团聚体破坏率基本呈现上升趋势(图 3)。相同土壤深度下, 赖草土壤团聚体破坏率显著低于其他植被类型(P < 0.05)。0—20 cm土层, 大针茅、长芒草的土壤团聚体破坏率为7.27%和7.98%。人工恢复植被土壤团聚体破坏率为8.58%—11.9%, 其中柠条土壤团聚体破毁坏率最低, 但比大针茅和长芒草分别增加6.99%、15.39%, 赖草土壤团聚体破坏率最低为6.28%, 比大针茅、长芒草分别降低13.68%、21.68%。20—40 cm和40—60 cm土层, 赖草土壤团聚体破坏率均低于其他恢复类型植被(P < 0.05), 油松和苜蓿最高, 相较于大针茅、长芒草分别增加15.18%—27.19%、40.93%—69.06%。总体来看, 人工恢复方式土壤团聚体破坏率在各土层均显著高于天然荒草和自然恢复方式的植被(P < 0.05), 0—20 cm、20—40 cm天然荒草与自然恢复方式植被差异不显著, 40—60 cm土层自然恢复方式显著低于其他两种恢复方式(表 4)。

|
| 图 3 不同植被类型土壤团聚体破坏率PAD分布特征 Fig. 3 PAD distribution characteristics of failure rate of soil aggregates under different vegetation types 不同小写字母表示同一植被类型不同土层间差异显著(P < 0.05), 不同大写字母表示同一土层不同植被类型间差异显著(P < 0.05); 图中数据为平均值±标准差; PAD: 土壤团聚体破坏率 The soil of percentage of aggregate destruction |
| 恢复方式 Restoration methods |
土壤深度 Soil depth/cm | |||||||
| 0—20 | 20—40 | 40—60 | ||||||
| 土壤团聚体破坏率 PAD |
土壤可蚀性因子 K |
土壤团聚体破坏率 PAD |
土壤可蚀性因子 K |
土壤团聚体破坏率 PAD |
土壤可蚀性因子 K |
|||
| 天然荒草 Natural grassland |
7.63±2.47Ba | 0.027±0.001Bb | 6.97±2.01Ba | 0.030±0.005Bb | 6.12±0.99Bb | 0.033±0.008Ba | ||
| 自然恢复 Natural restoration |
6.28±1.93Ba | 0.021±0.002Bb | 5.25±0.33Bb | 0.022±0.002Cb | 4.26±0.54Cc | 0.028±0.004Ca | ||
| 人工恢复 Artificial restoration |
10.07±1.49Aa | 0.037±0.004Ab | 8.52±0.89Ab | 0.044±0.004Ab | 8.81±0.70Ab | 0.048±0.005Aa | ||
| 不同小写字母表示同一恢复方式不同土层间差异显著(P < 0.05), 不同大写字母表示同一土层不同恢复方式间差异显著(P < 0.05); PAD: 土壤团聚体破坏率, The soil of percentage of aggregate destruction;K: 土壤可蚀性因子, Soil erodibility factor K | ||||||||
从土壤可蚀性因子K值来看, 各植被类型各土层抗侵蚀能力基本相同, 表现为0—20 cm土层土壤可蚀性因子最小, 40—60 cm土层土壤可蚀性因子最大(图 4)。其中, 赖草的土壤可蚀性因子显著低于其他植被类型(P < 0.05)。0—20 cm土层, 大针茅、长芒草土壤可蚀性因子K值为0.025和0.028, 油松K值为0.048, 相较于大针茅、长芒草土壤抗侵蚀能力降低约48%, 而赖草K值为0.021, 抗侵蚀能力相较于大针茅、长芒草约增加25%;20—40 cm和40—60 cm土层土壤抗侵蚀能力表现与0—20 cm土层相同, 均为油松土壤抗侵蚀能力最弱, 赖草土壤抗侵蚀能力最强。综合3种植被恢复方式, 0—60 cm土层人工恢复方式土壤可蚀性因子K值高于其他恢复方式, 而自然恢复方式K值小于其他恢复方式(P < 0.05)(表 4)。其中, 40—60 cm土层人工恢复方式K值最大为0.048, 抗侵蚀能力最弱, 相较于天然荒草植被土壤抗侵蚀能力降低约45%;0—20 cm土层自然恢复方式抗侵蚀能力最强, K值最小为0.021, 相较于天然荒草植被土壤抗侵蚀能力增加约25%。

|
| 图 4 不同植被类型土壤可蚀性因子K变化特征 Fig. 4 Variation characteristics of soil erodibility factor K under different vegetation types 不同小写字母表示同一植被类型不同土层间差异显著(P < 0.05), 不同大写字母表示同一土层不同植被类型间差异显著(P < 0.05); 图中数据为平均值±标准差 |
水稳性大团聚体与土壤平均重量直径呈显著性正相关, 与土壤团聚体破坏率呈极显著性负相关, 与土壤可蚀性因子K值之间呈负相关但不显著;平均重量直径与土壤可蚀性因子K值呈极显著性负相关;土壤团聚体破坏率与平均重量直径呈极显著性正相关, 与土壤可蚀性因子K值之间呈正相关但不显著(图 5)。

|
| 图 5 土壤团聚体与土壤稳定性之间的关系 Fig. 5 Relationship between soil aggregates and soil stability |
土壤水稳性团聚体含量越高, 土壤结构稳定性越好[28], 本研究自然恢复植被赖草水稳性大团聚体质量百分比含量最高, 人工恢复植被油松最低, 说明大面积种植油松不利于水稳性大团聚体的形成。土壤水稳性大团聚体的含量可以反映土壤抗侵蚀能力的强弱[29], 本文所研究的3种恢复方式7种植被类型大团聚体的质量百分比均在67.96%以上, 表明土壤结构较好, 其土壤抗侵蚀能力较强。土壤水稳性微团聚体(粒径 < 0.25 mm)是构成团聚体的基础, 其质量百分比以及分布对土壤物理性质有很大影响。本研究中人工恢复植被油松、山杏二者的土壤水稳性微团聚体(粒径 < 0.25 mm)含量均高于其他植被, 原因可能是研究区定西属于典型草原地带, 且年降雨量较少[30], 水分不足以供山杏和油松这类乔木的生长, 细根生物量少, 不利于大团聚体的形成;还可能是由于研究区油松林物种单一, 有大量的“小老树”, 掉落的松针外部裹有蜡质, 也会影响到水稳性大团聚体的形成。此外, 不同的降水分配以及林下植被等也可能影响植被分布, 造成土壤抗侵蚀能力的不同[31]。本文所研究的0—60 cm土层中人工恢复方式油松林土壤微团聚体的质量百分比最大, 可能因为本文所研究的油松林郁闭度高、冠层截留多造成林下光照弱, 净降雨量(穿透雨和树干径流)少, 林下植被稀疏, 造成因林下植被分布的土壤理化性质如有机质含量的差异, 进而导致土壤团聚体情况的异质性。
4.2 不同植被类型土壤团聚体稳定性分析土壤团聚体平均重量直径与土壤团聚体的团聚度、稳定性及土壤结构和质量呈正相关关系[32], 本文通过土壤团聚体平均重量直径分级结果可以得到, 自然恢复方式赖草草地土壤团聚体相较于其他恢复方式植被最稳定, 土壤抗侵蚀能力也较强。前期研究表明[33], 自然恢复方式更有利于增加植物物种多样性, 因而赖草草地比人工植被具有更高的物种多样性, 易于分解的草本凋落物归还相应增多。冯嘉仪等[34]发现土壤有机质可以增加土壤水稳性团聚体含量及其稳定性, 而凋落物是有机质的重要来源之一。其中, 凋落物分解后可以供给微生物养分使其得以生存和繁殖, 微生物通过分泌分泌物或分解作用形成多糖等胶结物质对土壤起到胶结作用, 将土壤小颗粒黏聚成较大的土壤团聚体, 有效提高土壤团聚体稳定性[35]。此外, 本研究发现表层土壤的土壤团聚体稳定性和抗蚀性最强, 可能表层土壤的凋落物含量、温度、水分以及含氧量等相对较深层土壤更充足, 为微生物群落的繁殖提供足够的条件, 微生物通过对土壤的胶结作用促进将微小团聚体聚成大粒径团聚体, 增加土壤水稳性团聚体的稳定性, 提高抗侵蚀能力[36]。除此之外, 土壤团聚体破坏率可以表明土壤团聚体在水蚀作用下的分散程度, 也是表明土壤团聚体稳定性的一个重要指标之一[37]。本研究通过与天然荒草进行对比发现, 自然恢复方式的赖草能显著降低土壤团聚体破坏率, 有研究表明[38—39]水稳性大团聚体含量越高, 团聚体破坏率就越小, 土壤结构与土壤抗侵蚀能力就越强, 赖草的水稳性大团聚体含量显著高于其他植被类型, 因此赖草的土壤团聚体破坏率最低, 土壤团聚体的稳定性最强。
4.3 土壤团聚体组成、稳定性及抗侵蚀能力关系分析土壤水稳性大团聚体(粒径直径>0.25 mm)含量可表明土壤团聚体稳定性[40], 本文研究的0—60 cm土层, 人工恢复方式的水稳性大团聚体含量均低于自然恢复方式(表 3), 水稳性大团聚体含量越高, 说明土壤团聚体越稳定, 所以自然恢复方式相较于人工恢复方式更有利于提高土壤团聚体稳定性。土壤水稳性团聚体平均重量直径和土壤可蚀性是评价土壤团聚体稳定性的两个重要指标[41]。土壤可蚀性因子K值与土壤团聚体关系密切, 有研究认为, 土壤水稳性团聚体平均重量直径、土壤水稳性大团聚体与侵蚀量和径流强度之间均呈现负相关, 可以用该指标衡量土壤抗侵蚀能力[25, 42—43], 土壤水稳性团聚体平均重量直径也能更好的反映土壤团聚体稳定性与侵蚀产沙和产流间的关系[44]。在本研究中, 土壤的水稳性大团聚体含量越高, 土壤平均重量直径越大、土壤团聚体破坏率越低, 土壤结构的稳定性就越好, 这与李程程等[30]对黄土高原土壤稳定性及抗侵蚀能力研究的结果一致, 大团聚体含量越高, 土壤团聚体越稳定;土壤可蚀性因子K值与土壤平均重量直径之间相关性更强, 且呈极显著负相关, 这与张钦等[25]和杨振奇[45]等研究的结果基本一致。
5 结论通过对比分析3种恢复方式, 7种植被类型对土壤水稳性团聚体稳定性及抗侵蚀能力的影响, 可以初步得出以下结论: 不同恢复方式不同植被类型间土壤水稳性团聚体稳定性差异性显著, 自然恢复方式的土壤水稳性团聚体平均重量直径显著高于天然荒草和人工恢复方式, 且7种植被类型土壤大团聚体质量百分比均在67.97%—90.12%之间;人工恢复方式土壤水稳性团聚体稳定性较差, 其中油松林稳定性最差。土壤团聚体破坏率和土壤抗侵蚀能力均表现出自然恢复方式更有利于土壤结构稳定性的提高。土壤水稳性大团聚体含量越高, 土壤团聚体越稳定, 土壤结构稳定性就越好;平均重量直径与土壤可蚀性因子K值呈极显著性负相关。综合所述, 自然恢复方式土壤团聚体的稳定性最高, 其土壤抗侵蚀能力最强, 建议该区采用自然恢复方式更有利于区生态恢复的有效进行。
| [1] |
刘国彬, 王国梁, 上官周平, 张文辉, 穆兴民, 谢永生, 李敏. 黄土高原地区水土保持科学研究的重点领域. 中国水土保持, 2008(12): 37-39, 72. DOI:10.3969/j.issn.1000-0941.2008.12.012 |
| [2] |
Chen N, Ma T Y, Zhang X P. Responses of soil erosion processes to land cover changes in the Loess Plateau of China: a case study on the Beiluo River Basin. CATENA, 2016, 136: 118-127. DOI:10.1016/j.catena.2015.02.022 |
| [3] |
Wang Z J, Jiao J Y, Rayburg S, Wang Q L, Su Y. Soil erosion resistance of "Grain for Green" vegetation types under extreme rainfall conditions on the Loess Plateau, China. CATENA, 2016, 141: 109-116. DOI:10.1016/j.catena.2016.02.025 |
| [4] |
周金星, 孙启祥, 崔明. 洞庭湖退田还湖区生态修复研究. 北京: 中国林业出版社, 2014.
|
| [5] |
Wang Q, Wang D, Wen X F, Yu G R, He N P, Wang R F. Differences in SOM decomposition and temperature sensitivity among soil aggregate size classes in a temperate grasslands. PLoS One, 2015, 10(2): e0117033. DOI:10.1371/journal.pone.0117033 |
| [6] |
黄晓强, 信忠保, 赵云杰, 马凤原. 北京山区典型人工林土壤团聚体组成及其有机碳分布特征. 水土保持学报, 2016, 30(1): 236-243. DOI:10.13870/j.cnki.stbcxb.2016.01.043 |
| [7] |
Das B, Chakraborty D, Singh V K, Aggarwal P, Singh R, Dwivedi B S, Mishra R P. Effect of integrated nutrient management practice on soil aggregate properties, its stability and aggregate-associated carbon content in an intensive rice-wheat system. Soil and Tillage Research, 2014, 136: 9-18. DOI:10.1016/j.still.2013.09.009 |
| [8] |
李嘉, 刘楠, 韩进斌, 赵建儒, 刘敏. 冻融对五台山典型林线植被土壤水稳性大团聚体的影响. 干旱区资源与环境, 2020, 34(5): 140-146. |
| [9] |
Zuo F, Li X, Yang X, Wang Y, Ma Y, Huang Y, Wei C. Soil particle-size distribution and aggregate stability of new reconstructed purple soil affected by soil erosion in overland flow. Journal of Soils and Sediments, 2020, 20(5): 272-283. |
| [10] |
Bryan R B. Soil erodibility and processes of water erosion on hillslope. Geomorphology, 2000, 32(3/4): 385-415. |
| [11] |
Krause L, Klumpp E, Nofz I, Missong A, Amelung W, Siebers N. Colloidal iron and organic carbon control soil aggregate formation and stability in arable Luvisols. Geoderma, 2020, 374: 114421. DOI:10.1016/j.geoderma.2020.114421 |
| [12] |
Okolo C C, Gebresamuel G, Zenebe A, Haile M, Eze P N. Accumulation of organic carbon in various soil aggregate sizes under different land use systems in a semi-arid environment. Agriculture, Ecosystems & Environment, 2020, 297: 106924. |
| [13] |
谢贤健, 张彬. 基于耦合关联分析的护岸植被恢复土壤抗蚀性综合评价. 土壤, 2019, 51(3): 609-616. DOI:10.13758/j.cnki.tr.2019.03.026 |
| [14] |
陈涛, 周利军, 齐实, 孙保平, 聂泽旭. 华蓥市山区典型林分土壤团聚体稳定性及抗蚀能力. 浙江农林大学学报, 2021, 38(6): 1161-1169. |
| [15] |
马祥华, 焦菊英, 白文娟. 黄土丘陵沟壑区退耕植被恢复地土壤水稳性团聚体的变化特征. 干旱地区农业研究, 2005, 23(3): 69-74. DOI:10.3321/j.issn:1000-7601.2005.03.015 |
| [16] |
郭曼, 郑粉莉, 安韶山, 刘雨, 王彬, Frederic Darboux. 应用Le Bissonnais法研究黄土丘陵区土壤团聚体稳定性. 中国水土保持科学, 2010, 8(2): 68-73. DOI:10.3969/j.issn.1672-3007.2010.02.012 |
| [17] |
由政, 姚旭, 景航, 谷利茶, 王国梁. 不同演替阶段群落根系分布与土壤团聚体特征的协同变化. 水土保持研究, 2016, 23(6): 20-25, 31. |
| [18] |
莫保儒, 王子婷, 蔡国军, 杨磊, 党宏忠, 王多锋, 薛睿. 半干旱黄土区成熟柠条林地剖面土壤水分环境及影响因子研究. 干旱区地理, 2014, 37(6): 1207-1215. |
| [19] |
殷地迟, 王立, 蔡国军, 莫保儒, 柴春山, 马驰. 半干旱黄土丘陵区不同植被类型的土壤水分特征及其稳定性. 水土保持通报, 2020, 40(1): 65-71. |
| [20] |
史瑞和, 鲍士旦, 秦怀英. 土壤农化分析. 北京: 中国农业出版社, 1998: 25-97.
|
| [21] |
翟婷婷, 谌芸, 李铁, 向明辉, 王润泽, 唐菡, 周涛, 刘枭宏. 植物篱篱前淤积带与篱下土坎土壤水库和抗剪性能对比研究. 生态学报, 2020, 40(2): 599-607. |
| [22] |
付鑫, 王俊, 刘全全, 李蓉蓉. 不同覆盖材料及旱作方式土壤团聚体和有机碳含量的变化. 植物营养与肥料学报, 2015, 21(6): 1423-1430. |
| [23] |
李柏桥, 付玉, 李光录, 张腾, 郑腾辉. 退耕年限与方式对土壤团聚体稳定性及有机碳分布的影响. 干旱地区农业研究, 2017, 35(3): 238-244. |
| [24] |
Le bissonnais Y. Aggregate stability and assessment of soil crustability and erodibility: I. Theory and methodology. European Journal of Soil Science, 1996, 47(4): 425-437. |
| [25] |
张钦, 于恩江, 林海波, 张爱华, 陈正刚, 朱青, 曹卫东, 姚单君. 连续种植不同绿肥作物的土壤团聚体稳定性及可蚀性特征. 水土保持研究, 2019, 26(2): 9-16. |
| [26] |
JR W, KG R, PT D. EPIC: A new method for assessing erosion's effect on soil productivity Journal of Soil and Water Conservation, 1983, 38(5): 381-383.
|
| [27] |
张科利, 彭文英, 杨红丽. 中国土壤可蚀性值及其估算. 土壤学报, 2007, 44(1): 7-13. |
| [28] |
Chaplot V, Cooper M. Soil aggregate stability to predict organic carbon outputs from soils. Geoderma, 2015, 243. |
| [29] |
严方晨, 焦菊英, 曹斌挺, 于卫洁, 魏艳红, 寇萌, 胡澍. 黄土丘陵沟壑区撂荒地不同演替阶段植物群落的土壤抗蚀性——以坊塌流域为例. 应用生态学报, 2016, 27(1): 64-72. |
| [30] |
李程程, 曾全超, 贾培龙, 安韶山. 黄土高原土壤团聚体稳定性及抗蚀性能力经度变化特征. 生态学报, 2020, 40(6): 2039-2048. |
| [31] |
吕宸, 宫渊波, 车明轩, 许蔓菁, 康成芳, 龚伟, 刘韩. 川西高寒山地灌丛草甸土壤抗蚀性研究. 水土保持学报, 2020, 34(2): 9-17. |
| [32] |
郑子成, 李廷轩, 张锡洲, 王永东, 杨玉梅, 汪曦微. 不同土地利用方式下土壤团聚体的组成及稳定性研究. 水土保持学报, 2009, 23(5): 228-231, 236. |
| [33] |
杨智姣, 温晨, 杨磊, 李宗善, 卫伟, 张钦弟. 半干旱黄土小流域不同恢复方式对生态系统多功能性的影响. 生态学报, 2020, 40(23): 8606-8617. |
| [34] |
冯嘉仪, 储双双, 王婧, 吴道铭, 莫其锋, 高婕, 林佳慧, 曾曙才. 华南地区几种典型人工林土壤有机碳密度及其与土壤物理性质的关系. 华南农业大学学报, 2018, 39(1): 83-90. |
| [35] |
龙健, 赵畅, 张明江, 吴劲楠, 吴求生, 黄博聪, 张菊梅. 不同坡向凋落物分解对土壤微生物群落的影响. 生态学报, 2019, 39(8): 2696-2704. |
| [36] |
宋日, 刘利, 马丽艳, 吴春胜. 作物根系分泌物对土壤团聚体大小及其稳定性的影响. 南京农业大学学报, 2009, 32(3): 93-97. |
| [37] |
宫阿都, 何毓蓉. 金沙江干热河谷区退化土壤结构的分形特征研究. 水土保持学报, 2001, 15(3): 112-115. |
| [38] |
史晓梅, 史东梅, 文卓立. 紫色土丘陵区不同土地利用类型土壤抗蚀性特征研究. 水土保持学报, 2007, 21(4): 63-66. |
| [39] |
吕文星, 张洪江, 王伟, 杜士才, 吴煜禾, 何萍, 肖莉. 重庆四面山不同林地土壤团聚体特征. 水土保持学报, 2010, 24(4): 192-197, 202. |
| [40] |
赵红, 袁培民, 吕贻忠, 李季. 施用有机肥对土壤团聚体稳定性的影响. 土壤, 2011, 43(2): 306-311. |
| [41] |
季波, 时龙, 徐金鹏, 何建龙, 王占军, 吴旭东, 蒋齐. 宁夏典型天然草地土壤团聚体稳定性及其有机碳分布特征. 生态学报, 2021, 41(19): 7669-7678. |
| [42] |
Barthès B, Roose E. Aggregate stability as an indicator of soil susceptibility to runoff and erosion; validation at several levels. CATENA, 2002, 47(2): 133-149. |
| [43] |
闫峰陵, 史志华, 蔡崇法, 李朝霞. 红壤表土团聚体稳定性对坡面侵蚀的影响. 土壤学报, 2007, 44(4): 577-583. |
| [44] |
陈正发, 史东梅, 谢均强, 张兵. 紫色土旱坡地土壤团聚体稳定性特征对侵蚀过程的影响. 中国农业科学, 2011, 44(13): 2721-2729. |
| [45] |
杨振奇, 郭建英, 秦富仓, 刘铁军, 刘艳萍, 邢恩德. 裸露砒砂岩区不同植被类型土壤团聚体稳定性与抗蚀性能. 水土保持通报, 2021, 41(3): 8-14. |