 2022, Vol. 42
2022, Vol. 42文章信息
- 郑昊哲, 张岩, 张涛, 樊庆山, 侯扶江
- ZHENG Haozhe, ZHANG Yan, ZHANG Tao, Fan Qingshan, HOU Fujiang
- 高寒草甸物种多样性对藏系牧羊放牧行为的影响
- Effects of species diversity on grazing behavior of Tibetan sheep in an alpine meadow
- 生态学报. 2022, 42(22): 8994-9004
- Acta Ecologica Sinica. 2022, 42(22): 8994-9004
- http://dx.doi.org/10.5846/stxb202112313728
-
文章历史
- 收稿日期: 2021-12-31
- 网络出版日期: 2022-07-13





2. 农业农村部草牧业创新重点实验室, 兰州 730020;
3. 贵州大学新农村发展研究院, 贵阳 550025
2. Key Laboratory of Grassland Livestock Industry Innovation Ministry of Agriculture and Rural Affairs, Lanzhou University, Lanzhou 730020, China;
3. Institute of New Rural Development, Guizhou University, Guiyang 550025, China
草原是全球面积最大的陆地生态系统, 放牧是草原最经济、最普遍的管理方式[1]。草畜互作是维持草原生物多样性的基础[2], 物种多样性与产草量有较为明确的数量关系[3]。牧草产量和品质是制约家畜放牧行为的重要因素, 草原植物物种多样性也影响放牧家畜的选择性采食和牧草品质。在一定范围内, 草原产草量与牧草品质负相关, 因此,植物物种多样性直接或间接地作用于家畜的放牧行为[4](图 1)。揭示草原植物物种多样性对家畜放牧行为的作用, 是阐明草畜互作机制、生态与生产兼顾实现可持续放牧管理的关键基础之一。

|
| 图 1 物种多样性与家畜放牧行为的关系概念图 Fig. 1 Sketch diagram of relationship between species diversity of grassland and grazing behavior of livestock |
家畜的放牧行为受气候和地形等自然因素、牧草和家畜等生物因素、补饲和放牧制度等管理因素的综合影响[5]。在高寒草甸, 高放牧率增加藏系牧羊的采食时间和采食速率, 减少其反刍时间;暖季放牧提高藏系牧羊的采食速率, 冷季降低其采食速率、觅食速率和反刍时间[6];灌丛盖度的增加会提高牦牛的采食速率和觅食时间, 减少牦牛反刍行为[7]。夏季甘肃马鹿在祁连山高寒灌丛放牧, 采食速率从出牧到午时逐渐升高;在高寒典型草原放牧, 采食速率变化规律呈“驼峰”曲线[8]。家畜采食降低高山牧场优势植物的生物量, 有利于不喜食牧草入侵, 增加植物物种丰富度[9]。不喜食牧草聚集分布时, 家畜日采食量增加, 而均匀分布时,家畜日采食量降低[10]。草原群落物种组成随放牧强度和放牧季节的变化各异[11]:高寒草甸家畜的放牧活动提高植物物种丰富度、Shannon-Wiener指数和Pilelou均匀度指数[12], 但随放牧率增加, 植物物种丰富度和牧草品质降低[11, 13], 由于家畜粪便分解速率缓慢, 抑制植物种子的萌发, 对群落植物Pilelou均匀度指数产生影响[14—16]。另一方面, 季节性轮牧改变了高寒草甸植物群落的结构和功能, 暖季放牧提高植物Shannon-Wiener指数和Pilelou均匀度指数, 冷季放牧促进植物地上生物量的积累[17]。可见, 选择性采食既影响家畜的放牧行为, 也改变草原群落多样性, 其中对放牧行为与产草量、牧草品质关系的作用可能是重要的途径(图 1), 虽然放牧对草原群落结构与功能的影响有大量研究报道, 然而较少涉及草原物种多样性与家畜放牧行为的关系。
青藏高原拥有全球连片放牧面积最大的草原之一, 是物种多样性对放牧管理响应最敏感的草原区之一[18]。藏系牧羊(Oura-type)作为青藏高原主要的家畜品种之一, 为当地和周边地区提供肉、奶和主要收入[19], 但冬季藏系牧羊生产力降低, 牧民通过增加饲养规模牺牲生态环境来维持收入水平, 草原退化日益严重[20—21]。为此, 我们在青藏高原高寒草甸开展藏系牧羊轮牧试验, 探索物种多样性与家畜放牧行为之间的关系, 力图揭示青藏高原放牧生态系统的草畜互作机制, 改进高寒草甸放牧管理提供依据。
1 材料与方法 1.1 研究区概况研究在兰州大学玛曲草地农业试验站(Maqu Grassland Agriculture Research Station of Lanzhou University, MGAS)(33°40′ N, 101°52′ E, 海拔3700 m)进行, 位于甘肃省玛曲县阿孜畜牧科技示范园区, 地处川、甘、青交汇处(图 2)。气候类型为高原大陆性高寒湿润气候, 低温多雨, 年均温1.2 ℃, 年均降水量610.06 mm(图 3)。土壤类型为亚高山草甸土壤[14]。根据草原综合顺序分类法, 草原类型为寒冷潮湿高山草甸草原类[22]。主要植物建群种和优势种有垂穗披碱草(Elymus nutans)和高山嵩草(Kobresia pygmaea);亚优势物种有赖草(Leymus secalinus);主要伴生种有紫花针茅(Stipa purpurea)、矮嵩草(Kobresia humilis)、草地早熟禾(Poa pratensis)等[23]。植物4月中下旬开始返青, 八月中下旬陆续枯黄。

|
| 图 2 研究区域的地理位置 Fig. 2 Location of the study site |

|
| 图 3 研究区月均降水量与气温变化 Fig. 3 Precipitation and Average temperature date from 2010—2011 for the Maqu County |
2010年开始划区轮牧试验, 7月1日—9月30日暖季(WG)轮牧, 放牧期10 d, 放牧周期30 d;10月1日—12月30日冷季(CG)轮牧, 放牧期15 d, 放牧周期30 d。每个放牧季节拥有6个100 m×100 m和6个50 m×100 m的放牧小区。每个放牧小区选取8只体况健康、较为一致的1岁羯羊。放牧率分别为8羊/hm2和16羊/hm2, 每个放牧率6个重复。每年有四个放牧处理分别是:暖季8羊/hm2(WG8);暖季16羊/hm2(WG16);冷季8羊/hm2(CG8);冷季16羊/hm2(CG16)。
1.2.2 植被测定2010年和2011年, 在牧草生长旺盛期进行植被测定和取样, 每个放牧小区布设4个0.5 m×0.5 m样方, 分别记录每个物种的密度以及株高。牧草齐地面刈割, 并将样品置入65 ℃恒温箱48 h, 测定每个物种的地上生物量。
样方中物种数目为物种丰富度, Shannon-Wiener指数和Pielou均匀度指数按以下公式计算。
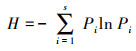
|
(1) |
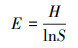
|
(2) |
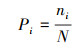
|
(3) |
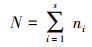
|
(4) |
式中:S为0.25 m2样方内的物种数;Pi为第ni个种的重要性;ni为第i个种的重要值;E为物种均匀度;N为所有种的重要值之和[24]。
1.2.3 藏系牧羊放牧行为观测每个放牧小区随机选取3只体型相近的藏系牧羊, 观测藏系牧羊的采食速率、觅食速率和采食时间等。
采食速率是在固定时间内观察到藏系牧羊对牧草采食的口数;觅食速率是藏系牧羊在采食过程中单位时间的行走速度。采食时间是藏系牧羊采食间隔小于15 s的时间的累加之和[6]。日采食量是藏系牧羊每天采食的牧草总量, 采用扣笼法测定, 家畜个体的日采食量根据采食口数和单口日采食量校正[25—26]。
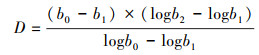
|
(5) |
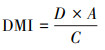
|
(6) |
式中:D为牧草消耗量;b0为牧前笼内和笼外的生物量; b1为牧后笼外的生物量; b2为牧后笼内的生物量;DMI为藏系牧羊的干物质日采食量;A为放牧小区的面积;C为藏系牧羊的数量[11]。
1.3 统计分析物种多样性与藏系牧羊放牧行为的拟合曲线通过Origin Pro 2021绘制, 通过SPSS 26.0分析物种多样性与藏系牧羊放牧行为的关系, 并进行一元线性回归模型的预测, 显著性水平为P=0.05。
通过R 4.1.0软件对数据进行统计分析, 拟合结构方程模型(Structural Equation Model, SEM)确定放牧管理与物种多样性-放牧行为间的关系。对植物物种丰富度、Shannon-Wiener多样性指数和物种均匀度指数进行主成分分析(Principal Component Analysis, PCA), 提取第一主成分并命名为物种多样性;对藏系牧羊采食速率、觅食速率和采食时间进行主成分分析, 提取第一主成分命名为放牧行为(图 7)。
2 结果与分析 2.1 物种丰富度与藏系牧羊放牧行为的关系放牧地物种丰富度与藏系牧羊的采食速率显著正相关(P < 0.05)(图 4), 物种丰富度每增加1个单位(1种/0.25m2), 暖季藏系牧羊采食速率平均提高0.40口/min, 冷季平均上升0.51口/min。可见, 物种丰富度越高, 高品质的优良牧草物种数越多, 藏系牧羊的采食速率越快。2010年放牧地物种丰富度与藏系牧羊的觅食速率显著正相关(P < 0.05)(图 4), 物种丰富度每增加1种, 藏系牧羊的觅食速率平均提高0.22步/min。但2011年物种丰富度与藏系牧羊觅食速率的关系对放牧季节的响应更加敏感(图 4), 两者关系暖季显著正相关(P < 0.05), 冷季负相关。同时, 放牧地物种丰富度与藏系牧羊采食时间的关系存在年际差异, 2010年显著负相关, 平均减少3.53 min(P < 0.05)(图 4);2011年两者关系对放牧率更加敏感, 8羊/hm2放牧率下正相关, 16羊/hm2放牧率下负相关(图 4)。

|
| 图 4 物种丰富度对藏系牧羊放牧行为的影响 Fig. 4 Effect of species richness on grazing behavior of Tibetan sheep 显著性水平:P < 0.05 |
放牧率显著影响物种丰富度与藏系牧羊日采食量的关系(P < 0.05)(图 4)。在8羊/hm2放牧率下, 两年间物种丰富度与藏系牧羊的日采食量显著正相关(P < 0.05), 物种丰富度每增加1种, 藏绵羊的日采食量平均增加7.30 g。16羊/hm2放牧率下, 除2010年冷季外, 物种丰富度与藏系牧羊日采食量显著负相关(P < 0.05)。
2.2 Shannon-Wiener多样性指数与藏系牧羊放牧行为的关系通过对两年的数据进行比较分析后发现(图 5), Shannon-Wiener指数增加后, 2011年藏系牧羊采食速率在16羊/hm2放牧率下有上升趋势(P < 0.05);2010年藏系牧羊觅食速率无显著变化, 但2011年暖季与Shannon-Wiener指数正相关, 冷季负相关(P < 0.05)(图 5);两年间在8羊/hm2放牧率下藏系牧羊采食时间呈夏减冬增的趋势(P < 0.05)(图 5); Shannon-Wiener指数与藏系牧羊日采食量的关系受放牧率和放牧季节的双重影响, 2010年藏系牧羊日采食量在8羊/hm2放牧率下增加, 16羊/hm2放牧率下降低(P < 0.05)(图 5); 2011年藏系牧羊日采食量对放牧季节敏感(P < 0.05), 暖季日采食量增加, 冷季日采食量降低(图 5)。

|
| 图 5 Shannon-Wiener指数与藏系牧羊放牧行为的关系 Fig. 5 The response of Tibetan sheep grazing behavior to Shannon-winner index |
除2011年16羊/hm2放牧率外, Pilelou均匀度指数与藏系牧羊采食速率显著负相关(P < 0.05)(图 6)。2010年Pilelou均匀度指数与藏系牧羊觅食速率的关系呈季节性变化, 暖季正相关, 冷季负相关(P < 0.05), 而2011年藏系牧羊觅食速率不受放牧管理的影响, 与Pilelou均匀度指数呈负相关(图 6)。两年间藏系牧羊采食时间与Pilelou均匀度指数呈负相关(P < 0.05)(图 6)。2010年放牧率影响了Pilelou均匀度指数与藏系牧羊日采食量的关系, 8羊/hm2放牧率下, 两者关系负相关(P < 0.05);2011年受气温的影响, 放牧季节成为影响两者关系的主要因素, 暖季藏系牧羊日采食量降低(P < 0.05)(图 6)。

|
| 图 6 Pielou均匀度指数与藏系牧羊放牧行为的关系 Fig. 6 The response of Tibetan sheep grazing behavior to Pielou index |
放牧季节对物种多样性和放牧行为有负面的影响, 而放牧率对放牧行为有直接积极的影响, 对物种多样性无显著影响(图 7)。放牧季节是影响物种多样性-放牧行为关系的重要因子, 与物种多样性和藏系牧羊放牧行为显著负相关, 与藏系牧羊日采食量显著正相关。放牧率与藏系牧羊放牧行为显著正相关, 与藏系牧羊日采食量显著负相关。物种多样性显著降低了藏系牧羊的日采食量, 但放牧行为对日采食量显著正相关。

|
| 图 7 放牧管理对物种多样性和藏系牧羊放牧行为的关系 Fig. 7 The relationship of grazing management to species diversity and grazing behavior of Tibetan sheep 箭头反映因果关系, 实线代表正相关, 虚线代表负相关, 箭头粗细与标准化路径系数呈正比; 显著性水平:***P < 0.001; **0.001≤P < 0.01; *0.01≤P < 0.05 |
物种多样性可预测藏系牧羊放牧行为, 植物物种多样性和藏系牧羊放牧行为可预测藏系牧羊日采食量(表 1)。
| 放牧行为 Grazing behavior |
方程 Equation |
R2 | P |
| BPM | = 0.743 SR+0.770 SW+21.481 | 0.649 | < 0.001 |
| T | = 6.937 SR+62.080 SW+125.366 | 0.322 | < 0.001 |
| DMI | = 16.014 SR+591.95 E+994.188 | 0.341 | < 0.001 |
| DMI | = 1.415 BPM+7.687 SPM+252.611 | 0.622 | < 0.001 |
| DMI | = -2.637 SR-419.758 SW+3.826 BPM+76.988 SPM+601.900 | 0.667 | < 0.001 |
| SR代表物种丰富度;SW代表Shannon-Wiener多样性指数;E代表Pielou均匀度指数;BPM、SPM、T和DMI分别代表藏系牧羊采食速率、觅食速率、采食时间和日采食量 | |||
藏系牧羊的放牧行为对物种丰富度的响应不同, 物种丰富度通过改变牧草营养品质、毒素和味道影响羊的放牧行为[27]。放牧显著增加高寒草甸植物物种丰富度, 为家畜提供了更为优质的牧草资源[28], 藏系牧羊选择牧草的时间逐渐减少, 选择性采食能力显著降低, 觅食时间缩短[5, 27]。面对复杂的觅食环境, 羊通过减少采食时间, 增加觅食速率, 选择更多的植物组合来满足自身的营养需求。同时, 植物物种丰富度的增加, 植物纤维含量降低, 进一步刺激羊的觅食行为, 增加羊的采食速率[28]。在松嫩平原, 物种丰富度的增加为绵羊提供了更多的食物资源, 从而增加绵羊的日采食量[29]。在高强度的放牧下, 土壤水分降低, 容重增加, 禾本科植物功能群比例下降, 杂草类植物功能群比例上升[30], 植物物种丰富度增加, 但高放牧率使藏系牧羊剔除了喜食物种的生物量[31], 物种丰富度增加后, 群落中低矮且有毒的植物物种数量增加, 营养品质下降, 藏系牧羊日采食量降低。
3.2 Shannon-Wiener指数对藏系牧羊放牧行为的影响放牧对物种多样性指数有较显著的影响[32], 放牧增加Shannon-Wiener指数, 丰富草原的物种多样性[33]。2010年藏系牧羊的放牧活动抑制了优势牧草的竞争优势, 促进一些弱势植物种的入侵和定居[34], 牧草适口性下降, 为竞争更为优质的牧草, 藏系牧羊增强选择性采食能力, 在短时间内增加采食速率和觅食速率, 从而竞争更为优质的牧草。季节性放牧不影响草地物种多样性, 但显著影响草地植物的营养品质[11, 13]。2011年夏季降水少, 冬季降水多, 藏系牧羊放牧行为受季节影响较大。暖季干旱条件下, 放牧减少植被地上生物量, 土壤腐殖质含量下降, 土壤养分含量降低, 使得牧草生长受到抑制, 提前进入枯黄期, 营养品质降低, 藏系牧羊加快觅食速率、降低采食时间, 以便寻找品质更佳的牧草;冷季雨水增加, 放牧的践踏使得土壤腐殖质增加, 加之雨水的淋溶和雪层的保温, 加快牧草返青, 藏系牧羊通过降低觅食速率、延长采食时间, 从而减少自身能量的消耗, 保证采食效率。研究表明,高寒草甸草原放牧强度与植物地上生物量和牧草营养品质均负相关, 但与Shannon-Wiener指数无显著关系[13, 32, 35]。因此, 放牧率与牧草地上生物量和营养品质呈现负相关。随植物物种多样性增加, 藏系牧羊提高自身日采食量, 获取更多的营养维持生长代谢。高放牧率下, 植物密度和高度降低[11], 藏系牧羊的采食难度增加, 对牧草的选择性降低[36], 低矮不易采食的植物种类和数量偏多, 地上生物量趋于下降, 藏系牧羊的日采食量降低。
3.3 物种均匀度对藏系牧羊放牧行为的影响物种均匀度反应了不同地区植物物种的分配程度, 与物种丰富度正相关[37]。研究表明, 放牧条件下, Pilelou均匀度指数年际差异显著, 与放牧年份显著负相关[32]。随Pilelou均匀度指数的增加, 植物种类增多, 可食性牧草充足, 藏系牧羊的采食速率下降。由于放牧增加了植物种间竞争作用, 加之气候因素的影响, 次年在高放牧率下, 可食性牧草减少, 藏系牧羊的采食速率增加。在牧草生长旺期, 较低的放牧率促进草地向优势物种均匀化方向发展, 对植物的影响主要体现在叶片营养品质的增加[38], 降低了藏系牧羊寻找食物的难度。因此, 藏系牧羊觅食速率与放牧率负相关, 日采食量与其正相关。在冷季, 藏系牧羊觅食速率和日采食量下降, 可能是由于恶劣的气候加之藏系牧羊的践踏作用使得优势牧草高度下降, 加剧土壤水分的蒸发和土壤C、N养分流失, 牧草营养品质降低。禾本科等优势牧草随着藏系牧羊的采食削弱了自身的竞争优势, 使得群落整体向低矮化植物的方向转变[39], 导致Pilelou均匀度指数与藏系牧羊觅食时间显著正相关, 与藏系牧羊采食时间显著负相关。
3.4 放牧管理对物种多样性-藏系牧羊采食行为关系的影响家畜放牧行为的变化主要受环境因素影响, 是由家畜和草地管理方法所塑造的草地植物群落特征变化的结果[40—41]。研究发现, 植物的群落特征、生物量和营养品质随季节的变化而变化, 从而影响家畜的放牧行为[11]。从暖季到冷季, 一年生植物枯萎凋落, 优势物种的分配更为均匀, Pilelou均匀度指数增加, 但降低了植物的高度和密度, 导致植物物种丰富度和Shannon-Wiener指数呈现下降的趋势。随植物物种丰富度的降低, 地上生物量的骤减, 藏系牧羊的日采食量下降, 同时藏系牧羊为抵御寒冷减少能量消耗维持自身代谢, 降低了采食速率、觅食速率和采食时间[11]。
放牧强度的变化影响家畜的放牧时间[42], 但对植物物种多样性无显著影响[13, 36]。本研究中, 放牧率直接影响藏系牧羊的放牧行为, 高强度的放牧下, 藏系牧羊的采食和践踏降低了牧草高度和密度, 在受限的环境内, 随着牧草的地上生物量和营养品质下降, 藏系牧羊必须增加觅食时间和采食时间去选择和采食更加优质牧草, 因此藏系牧羊的采食速率、日采食量和觅食速率下降。
4 结论放牧季节和放牧率导致藏系牧羊的放牧行为对不同植物多样性表现出多种响应。物种丰富度的变化直观地反映藏系牧羊选择性采食, Shannon-Wiener多样性指数和Pilelou均匀度指数易受气候的影响, 藏系牧羊放牧行为呈现季节性动态。物种丰富度的变化可以预测藏系牧羊的放牧行为和日采食量, 为提高藏系牧羊生产力, 维护草原生态稳定提供科学依据。
| [1] |
侯扶江, 杨中艺. 放牧对草地的作用. 生态学报, 2006, 26(1): 244-264. DOI:10.3321/j.issn:1000-0933.2006.01.031 |
| [2] |
王德利, 王岭. 草地管理概念的新释义. 科学通报, 2019, 64(11): 1106-1113. |
| [3] |
包乌云, 邢旗, 张健, 刘亚玲, 程磊, 陈翔, 杜宇凡, 阿拉腾苏和. 乌拉盖草原植物群落多样性现状. 草原与草业, 2018, 30(3): 13-20. DOI:10.3969/j.issn.2095-5952.2018.03.006 |
| [4] |
石岳, 马殷雷, 马文红, 梁存柱, 赵新全, 方精云, 贺金生. 中国草地的产草量和牧草品质: 格局及其与环境因子之间的关系. 科学通报, 2013, 58(3): 226-239. |
| [5] |
Hilario M C, Wrage-Mönnig N, Isselstein J. Behavioral patterns of (co-)grazing cattle and sheep on swards differing in plant diversity. Applied Animal Behaviour Science, 2017, 191: 17-23. DOI:10.1016/j.applanim.2017.02.009 |
| [6] |
Du W C, Yan T, Chang S H, Wang Z F, Hou F J, Du W C, Yan T, Chang S H, Wang Z F, Hou F J. Seasonal hogget grazing as a potential alternative grazing system for the Qinghai-Tibetan plateau: weight gain and animal behaviour under continuous or rotational grazing at high or low stocking rates. The Rangeland Journal, 2017, 39(4): 329-339. DOI:10.1071/RJ16119 |
| [7] |
Yang C T, Tsedan G, Fan Q S, Wang S L, Wang Z F, Chang S H, Hou F J. Behavioral patterns of yaks (Bos grunniens) grazing on alpine shrub meadows of the Qinghai-Tibetan Plateau. Applied Animal Behaviour Science, 2021, 234: 105182. DOI:10.1016/j.applanim.2020.105182 |
| [8] |
侯扶江, 任继周. 甘肃马鹿冬季放牧践踏作用及其对土壤理化性质影响的评价. 生态学报, 2003, 23(3): 486-495. DOI:10.3321/j.issn:1000-0933.2003.03.012 |
| [9] |
Descombes P, Pitteloud C, Glauser G, Defossez E, Kergunteuil A, Allard P M, Rasmann S, Pellissier L. Novel trophic interactions under climate change promote alpine plant coexistence. Science, 2020, 370(6523): 1469-1473. DOI:10.1126/science.abd7015 |
| [10] |
Dennis T S, Unruh-Snyder L J, Neary M K, Nennich T D. Effects of co-grazing dairy heifers with goats on animal performance, dry matter yield, and pasture forage composition. Journal of Animal Science, 2012, 90(12): 4467-4477. DOI:10.2527/jas.2011-4643 |
| [11] |
Xiao X, Zhang T, Angerer J P, Hou F J. Grazing seasons and stocking rates affects the relationship between herbage traits of alpine meadow and grazing behaviors of Tibetan Sheep in the Qinghai-Tibetan Plateau. Animals, 2020, 10(3): 488. DOI:10.3390/ani10030488 |
| [12] |
Lu X Y, Kelsey K C, Yan Y, Sun J, Wang X D, Cheng G W, Neff J C. Effects of grazing on ecosystem structure and function of alpine grasslands in Qinghai-Tibetan Plateau: a synthesis. Ecosphere, 2017, 8(1): e01656. DOI:10.1002/ecs2.1656 |
| [13] |
柴林荣, 孙义, 王宏, 常生华, 侯扶江, 程云湘. 牦牛放牧强度对甘南高寒草甸群落特征与牧草品质的影响. 草业科学, 2018, 35(1): 18-26. |
| [14] |
颜才玉, 孙义, 刘阳, 王召峰, 常生华, 侯扶江. 两个放牧率下藏羊粪在高寒草甸的分解特征. 生态学报, 2018, 38(20): 7208-7215. |
| [15] |
徐长林, 鱼小军, 景媛媛, 肖红, 安晓东, 杨发森, 任宝虎, 周瑞娟. 牦牛粪和藏羊粪浸提液对高寒草甸六种植物种子萌发特性的影响. 生态学杂志, 2014, 33(11): 2988-2994. |
| [16] |
景媛媛, 徐长林, 陈陆军, 鱼小军, 杨海磊, 张建文, 肖红. 高寒草甸冷季牧场牦牛和藏羊粪中植物种子密度和多样性. 生态学杂志, 2014, 33(10): 2603-2609. |
| [17] |
刘玉, 刘振恒, 邓蕾, 武高林. 季节性放牧对草地植物多样性与功能群特征的影响. 草业科学, 2016, 33(7): 1403-1409. |
| [18] |
Wang D L, Wang L, Liu J S, Zhu H, Zhong Z W. Grassland ecology in China: perspectives and challenges. Frontiers of Agricultural Science and Engineering, 2018, 5(1): 24-43. DOI:10.15302/J-FASE-2018205 |
| [19] |
Yuan H, Hou F J. Grazing intensity and soil depth effects on soil properties in alpine meadow pastures of Qilian Mountain in Northwest China. Acta Agriculturae Scandinavica, Section B — Soil & Plant Science, 2015, 65(3): 222-232. |
| [20] |
Sun Y, Angerer J P, Hou F J. Effects of grazing systems on herbage mass and liveweight gain of Tibetan sheep in eastern Qinghai-Tibetan Plateau, China. The Rangeland Journal, 2015, 37(2): 181-190. DOI:10.1071/RJ14062 |
| [21] |
Zhang Y J, Zhang X Q, Wang X Y, Liu N, Kan H M. Establishing the carrying capacity of the grasslands of China: a review. The Rangeland Journal, 2014, 36(1): 1-9. DOI:10.1071/RJ13033 |
| [22] |
Ren J Z, Hu Z Z, Zhao J, Zhang D G, Hou F J, Lin H L, Mu X D. A grassland classification system and its application in China. The Rangeland Journal, 2008, 30(2): 199-209. |
| [23] |
周杰, 赵洪, 袁晓霞, 杨玉婷, 罗开嘉, 郭正刚. 青藏高原边缘区高寒草甸植物群落的特征. 草业科学, 2013, 30(7): 1066-1070. |
| [24] |
Chu H, Zhang C P, Dong Q M, Shang Z H, Degen A A, Yang X X, Yu Y, Yang Z Z, Zhang Y F. The effect of grazing intensity and season on the soil seed bank and its relation with above-ground vegetation on the alpine steppe. Agriculture, Ecosystems & Environment, 2019, 285: 106622. |
| [25] |
Smit H J, Taweel H Z, Tas B M, Tamminga S, Elgersma A. Comparison of techniques for estimating herbage intake of grazing dairy cows. Journal of Dairy Science, 2005, 88(5): 1827-1836. |
| [26] |
金有顺, 侯扶江. 放牧家畜采食量的测定. 动物营养学报, 2020, 32(7): 3012-3030. |
| [27] |
Wang L, Wang D L, Liu J S, Huang Y, Hodgkinson K C. Diet selection variation of a large herbivore in a feeding experiment with increasing species numbers and different plant functional group combinations. Acta Oecologica, 2011, 37(3): 263-268. |
| [28] |
Feng C, Ding S W, Zhang T Y, Li Z Q, Wang D L, Wang L, Liu C, Sun J Y, Peng F G. High plant diversity stimulates foraging motivation in grazing herbivores. Basic and Applied Ecology, 2016, 17(1): 43-51. |
| [29] |
Wang L, Wang D L, He Z B, Liu G F, Hodgkinson K C. Mechanisms linking plant species richness to foraging of a large herbivore. Journal of Applied Ecology, 2010, 47(4): 868-875. |
| [30] |
崔树娟, 布仁巴音, 朱小雪, 白玲, 汪诗平. 不同季节适度放牧对高寒草甸植物群落特征的影响. 西北植物学报, 2014, 34(2): 349-357. |
| [31] |
Filazzola A, Matter S F, Roland J. Inclusion of trophic interactions increases the vulnerability of an alpine butterfly species to climate change. Global Change Biology, 2020, 26(5): 2867-2877. |
| [32] |
牛钰杰, 杨思维, 王贵珍, 刘丽, 花立民. 放牧干扰下高寒草甸物种多样性指数评价与选择. 应用生态学报, 2017, 28(6): 1824-1832. |
| [33] |
Pulungan M A, Suzuki S, Gavina M K A, Tubay J M, Ito H, Nii M, Ichinose G, Okabe T, Ishida A, Shiyomi M, Togashi T, Yoshimura J, Morita S. Grazing enhances species diversity in grassland communities. Scientific Reports, 2019, 9(1): 1-8. |
| [34] |
王志鹏, 张兆杰, 花立民. 不同放牧强度对高寒草甸有毒植物多样性的影响. 草原与草坪, 2019, 39(5): 75-81. |
| [35] |
徐鑫磊, 宋彦涛, 赵京东, 乌云娜. 施肥和刈割对呼伦贝尔草甸草原牧草品质的影响及其与植物多样性的关系. 草业学报, 2021, 30(7): 1-10. |
| [36] |
Török P, Penksza K, Tóth E, Kelemen A, Sonkoly J, Tóthmérész B. Vegetation type and grazing intensity jointly shape grazing effects on grassland biodiversity. Ecology and Evolution, 2018, 8(20): 10326-10335. |
| [37] |
蔡艳, 吕光辉, 何学敏, 蒋腊梅, 王恒方, 滕德雄. 不同利用方式下草地生态系统的多功能性与物种多样性. 干旱地区农业研究, 2019, 37(5): 200-210. |
| [38] |
Cerabolini B, Pierce S, Luzzaro A, Ossola A. Species evenness affects ecosystem processes in situ via diversity in the adaptive strategies of dominant species. Plant Ecology, 2010, 207(2): 333-345. |
| [39] |
周晓松, 朱志红, 李英年, 袁芙蓉, 樊瑞俭. 刈割、施肥和浇水处理下高寒矮嵩草草甸补偿机制. 兰州大学学报: 自然科学版, 2011, 47(3): 50-57. |
| [40] |
Hirata M, Kunieda E, Tobisa M. Short-term ingestive behaviour of cattle grazing tropical stoloniferous grasses with contrasting growth forms. The Journal of Agricultural Science, 2010, 148(5): 615-624. |
| [41] |
Scotton M, Crestani D. Traditional grazing systems in the Venetian Alps: effects of grazing methods and environmental factors on cattle behaviour. Journal of Environmental Management, 2019, 250: 109480. |
| [42] |
Animut G, Goetsch A L, Aiken G E, Puchala R, Detweiler G, Krehbiel C R, Merkel R C, Sahlu T, Dawson L J, Johnson Z B, Gipson T A. Grazing behavior and energy expenditure by sheep and goats co-grazing grass/forb pastures at three stocking rates. Small Ruminant Research, 2005, 59(2/3): 191-201. |