 2022, Vol. 42
2022, Vol. 42文章信息
- 林晓雪, 黄佳芳, 李慧, 仝川
- LIN Xiaoxue, HUANG Jiafang, LI Hui, TONG Chuan
- 闽江河口芦苇沼泽和短叶茳芏沼泽生态系统含碳温室气体的年收支
- Annual carbon gases budgets for Phragmites australis marsh and Cyperus malaccensis marsh in the Minjiang River estuary
- 生态学报. 2022, 42(22): 9186-9198
- Acta Ecologica Sinica. 2022, 42(22): 9186-9198
- http://dx.doi.org/10.5846/stxb202107272036
-
文章历史
- 收稿日期: 2021-07-27
- 网络出版日期: 2022-07-12





2. 湿润亚热带生态-地理过程教育部重点实验室, 福建师范大学, 福州 350007;
3. 福建闽江河口湿地生态系统国家定位观测研究站(国家林草局), 福州 350215
2. Key Laboratory of Humid Subtropical Eco-geographical Process, Ministry of Education, Fujian Normal University, Fuzhou 350007, China;
3. Wetland Ecosystem Research Station of Minjiang Estuary, State Forestry and Grassland Administration, Fuzhou 350215, China
滨海感潮沼泽湿地是全球天然湿地的重要组成部分, 是全球生产力较高的植物群落类型, 它与滨海红树林和海草床生态系统共同组成了海岸带蓝碳生态系统[1—2]。滨海感潮沼泽湿地植物通过光合作用固定大气中二氧化碳(CO2), 构成植物的地上/地下碳库及土壤碳库, 在减缓全球变缓趋势中发挥着巨大的、不可替代的作用[3]。
湿地生态系统碳收支或称碳平衡及CO2净交换量(net ecosystem exchange of CO2, NEE of CO2)的测定是准确估算湿地生态系统碳汇规模的基础和关键环节。湿地生态系统碳收支、CO2净交换量的主要测定方法为通量塔涡度协方差法[4—6], 此外, 透明箱以及黑布遮盖+红外气体分析仪相结合的方法在没有涡度相关通量塔的湿地研究中也得到较广泛的应用[7—10]。由于研究区域可达性以及观测条件的限制, 目前鲜见对滨海感潮沼泽生态系统碳收支, 包括NEE、生态系统呼吸(ecosystem respiration, ER)和甲烷(CH4)排放通量等过程的系统性测定数据, 较少的研究报道集中在对于感潮沼泽湿地生态系统CO2净交换量的观测[6, 11], Yan等利用2个涡度相关通量塔, 结合MODIS数据研究了长江口感潮沼泽湿地生态系统CO2净交换量的变化, 近岸和近海点感潮沼泽湿地生态系统NEE分别为-2.02 g C m-2 d-1和-1.42 g C m-2 d-1, 平均NEE为-1.75 g C m-2 d-1[12]。Vázquez-Lule和Vargas[13]利用3年的涡度相关通量塔数据分析表明:感潮盐沼生态系统NEE在各物候期间差异显著, 且该感潮盐沼是一个净的排向大气的碳源, 这一结果也挑战了人们普遍认为的滨海感潮沼泽湿地是一个净的自然蓝碳碳汇的认知。滨海感潮沼泽具体是大气中的碳汇还是碳源, 除了要考虑感潮沼泽CH4排放的影响, 即沼泽CH4排放是否会抵消部分沼泽生态系统CO2的净吸收以外, 河口区从感潮淡水-半咸水-盐沼这一盐度梯度上的不同湿地均需要全面和系统的测定, 而这方面研究鲜见报道。
本研究运用明、暗箱+红外气体分析仪/气相色谱相结合的方法, 并使用不同遮光率的布罩住静态箱模拟不同光照条件, 对闽江河口鳝鱼滩分布的半咸水芦苇沼泽和短叶茳芏沼泽湿地生态系统CO2气体的瞬时NEE、ER和CH4排放通量进行测定, 并在月和年尺度上估算2个湿地生态系统含碳气体收支及固碳量, 以期厘清闽江河口主要土著种沼泽湿地生态系统的碳汇规模, 回答沼泽湿地CH4排放在多大程度上抵消沼泽湿地生态系统CO2净吸收产生的固碳效益,为科学评估我国海岸带感潮沼泽湿地生态系统蓝碳固碳能力, 以及滨海感潮沼泽湿地生态系统管理和生态系统服务功能的评价和维系提供科学依据。
1 材料与方法 1.1 研究区域概况研究区位于福建闽江河口鳝鱼滩湿地中部的中潮滩(26°0′36″N—26°3′42″N, 119°34′12″E—119°41′40″E)。研究区与采样点位置如图 1。研究区属于亚热带海洋性季风气候区, 温暖湿润。冬季盛行东北风或东风, 夏季盛行东南风。区内雨量充沛, 年平均降水量为1346 mm, 集中于3—9月[14]。潮汐为正规半日潮, 涨落潮历时基本相等, 比值为1∶1.15。闽江河口鳝鱼滩湿地分布的主要土著种沼泽植物有芦苇(Phragmites australis)、短叶茳芏(Cyperus malaccensis)和海三棱藨草(Scirpus×triqueter)。本研究选择鳝鱼滩中部中潮滩相邻的芦苇沼泽湿地和短叶茳芏沼泽湿地为研究对象开展碳收支的研究。研究区从涨潮开始至落潮结束平均潮水水淹时间为3.5—4 h, 在非涨落潮阶段, 地表完全出露。研究区5—12月潮水盐度均值为4.2‰[15], 故本研究中的芦苇沼泽湿地和短叶茳芏沼泽湿地均为半咸水(brackish)沼泽湿地。研究区芦苇群落结构单一, 为单一优势种群落, 芦苇植株高度夏季达到最高(1.6—1.8m), 地上生物量在夏季也达到峰值(1524.8±78.9)g/m2, 土壤(0—50 cm)有机碳含量为(22.1±2.0)g/kg[15]。研究区短叶茳芏群落结构简单, 也为单一优势种群落, 3月初进入生长期, 6—9月生长旺盛, 7月份植株高度1.5 m, 地上生物量为(1062.4±129.6)g/m2, 土壤(0—50 cm)有机碳含量为(19.7±0.7)g/kg[14]。

|
| 图 1 研究区与采样点图 Fig. 1 Location of the study area and sampling site |
本研究的测定时间为2019年8月—2020年7月, 期间2020年2月—4月因疫情原因未能测定。在选定的芦苇沼泽湿地及其临近的短叶茳芏沼泽湿地内分别选取3个样地(3个重复), 搭建木质栈桥通往研究样地。在每个样地布置透明静态箱, 透明静态箱由底座、中箱和顶箱(在中箱高度不够时使用, 特别是在夏季)三部分组成, 底座由PVC材料制成, 规格(长×宽×高)为35 cm×35 cm×30 cm, 底座顶部设有密封水槽。在采样前15 d将底座布设在样地中。中箱与顶箱均为透明亚克力材料制作。中箱的规格(长×宽×高)均为35 cm×35 cm×120 cm, 中箱顶部也带有水槽, 在箱体两侧各设1个采气孔用于连接CO2气体分析仪。顶箱规格(长×宽×高)为35 cm×35 cm×50(或100)cm。顶箱顶部及中箱侧壁各安装1个小风扇以混匀箱内气体。采气时将中箱、顶箱依次叠放到底座上方, 罩住植物, 在底座与中箱顶部水槽中加入水以起到密封的作用。
为了估算月和年尺度上的NEE动态时间变化, 需测定不同光照情景下的NEE和ER。本研究每月选择一个晴天, 在中午光强较高时段测定。使用不同遮光率的布遮住透明静态箱以模拟不同光照强度, 分别测定有光状况下(透光率分别为100%(不遮光), 70%, 45%, 15%)瞬时净生态系统CO2通量, 即CO2净交换通量(NEE)。在有光状况测定结束后, 最后用透光率为0%的黑布遮住静态箱, 测定黑暗状况下的生态系统呼吸(ER)[7—9]。用遮光布罩住静态箱后, 通过软管与CO2/H2O气体分析仪(LI-850, 美国)相连, 平衡0.5 min后, 测定每个遮光率下箱内的CO2浓度, 每个遮光率下共测定2 min。测定完某一遮光率下的箱内CO2浓度后, 取下遮光布以及静态箱中箱或顶箱, 使静态箱内气体与大气平衡后, 时间间隔为1.5 min, 再遮上其它遮光率的布罩, 测定其它光照强度下的NEE, 5个不同光强处理的NEE测定45 mins内完成。NEE和ER根据静态箱内CO2浓度的变化率计算得出[6, 9]。NEE负值表示生态系统从大气中净吸收CO2, 是大气中CO2的汇;NEE正值表示生态系统向大气中净排放CO2, 是大气中CO2的源。
1.3 CH4排放通量的测定静态暗箱+气相色谱法是测定湿地生态系统CH4排放通量的常用方法[16—17]。在CO2通量的测定结束后, 对于静态暗箱(用遮光率为100%的布罩住静态箱), 用60 mL注射器抽取箱内气样, 采样量为50 mL, 装入真空气袋, 每隔15 min采集1次, 共抽气4次。整个采气过程向底座和中箱间及中箱和顶箱间的水槽加水密封。气样立刻带回实验室测定。
气体样品及时运回实验室, 利用气相色谱仪(岛津GC-2010, 日本)测定CH4气体浓度。检测器为FID (氢焰离子化检测器)。测气时使用氢气作为燃气, 流速为47 mL/min, 载气为N2, 流速20 mL/min, 检测器温度为130℃, 分离柱温度为60℃。用南京特种气体厂生产的浓度为2.15, 51.58和101 μL/L的CH4标准气体校正仪器。CH4排放通量计算公式[14—15]如下:
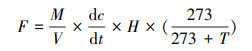
|
(1) |
式中, F为CH4排放通量(mg m-2 h-1);M为CH4摩尔质量(g/mol);V为标准状态下的气体摩尔体积(22.4 L/mol);dc/dt为CH4浓度变化率;T为采样箱内温度(℃); H为采样箱的箱高(m)。测量的浓度数据只有在线性回归系数r2> 0.90才视为有效数据[14]并计算CH4排放通量。
1.4 环境因子的测定测定不同光强下NEE的同时, 同步使用温度计测定箱内气温, 同时使用Light Scout 3415FX光合有效辐射计(Spectrum, 美国)测定箱内光合有效辐射值(PAR)。同时, 每15 min观测一次气温, 每10 min观测一次光合有效辐射, 二者数据均读自样地附近的闽江河口湿地生态系统定位观测站(国家林草局)的自动气象站(Campbell, 美国)(距离研究样地约30 m)。与采集气样同步, 原位使用2265FS土壤电导/温度记录仪(Spectrum, 美国)测定各样地土壤温度。
1.5 年尺度上NEE, ER和GEE的估算总光合作用吸收量的表征指标较多, 包括GPP(gross primary productivity)[18—19], GPP(gross photosynthetic production)[20], 或GEE(gross ecosystem exchange)[9, 21—22], 或GEP(gross ecosystem production)[5], 这些指标的实质是一致的, 均表征生态系统光合作用对大气中碳的总吸收量。本研究选用GEE。GEE为负值, 表示生态系统光合作用吸收的总碳量[22]。基于不同光照情景下测定的瞬时NEE和ER值可计算湿地生态系统总光合作用碳吸收量(GEE), 然后建立GEE与光合有效辐射值(PAR)的拟合关系模型, 以及ER与气温的拟合关系, 最后根据GEE与NEE和ER的关系式[9, 22], 计算日、月和年尺度上的NEE值[9]。
NEE与ER计算GEE的公式见公式2[22]。
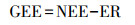
|
(2) |
GEE与PAR进行直角双曲线拟合。
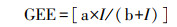
|
(3) |
式中:GEE, μmol CO2 m-2 s-1; I为PAR(μmol m-2 s-1); a, b为常数[9, 23]。
10 min 1次的PAR数据由样地附近的闽江河口湿地生态系统定位观测站(国家林草局)的自动气象站读取, 根据公式3计算每天每10 min一次的GEE值, 并将每天内所有GEE数值求和累加得出该天的光合作用碳总吸收量, 然后将各月所有天的光合作用碳总吸收量累积相加便得到各月的光合作用碳总吸收量(g CO2 m-2月-1), 最后各月光合作用碳总吸收量相加, 便得到年GEE。
为了估算日ER动态时间变化, 将正午测定的ER与气温按以下指数方程进行拟合[9, 22]:
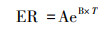
|
(4) |
ER为生态系统呼吸速率(μmol C m-2 s-1), T为气温(℃), A, B为经验常数[24]。
15 min 1次的气温数据由样地附近的闽江河口湿地生态系统定位观测站(国家林草局)的自动气象站读取, 利用公式4得出每15min一次的ER值, 将每个月所有数值求和累加得出该月累积的生态系统呼吸总量(g CO2 m-2 month-1), 最后各月相加, 便得到年ER值。
NEE月累计值通过GEE与ER月累计值计算得出, 将各月的NEE数值求和累加得到年NEE(g CO2/m2)。2020年2月—4月因疫情原因未能测定正午的NEE和ER, 考虑到2月、3月植物还未进入生长期, 故2020年2月、3月的GEE、ER与PAR关系的拟合是代入2020年1月的GEE数据, 2020年4月则是利用2020年5月的GEE数据进行拟合。
1.6 年尺度上的湿地生态系统蓝碳碳汇的估算根据每月测定的1次的CH4排放通量, 估算日、月和年的研究样地CH4排放通量, 并根据全球变暖潜力(global warming potential, GWP), 在百年尺度上单位CH4的CO2当量为25, 将年CH4排放通量转化为CO2当量排放通量(CO2-equivalent flux)。
CO2净生态系统交换通量与CH4排放通量的代数和, 即为沼泽湿地生态系统固碳量(carbon sequestration), 即湿地生态系统蓝碳碳汇, 公式如下:
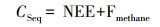
|
(5) |
CSeq: 湿地生态系统固碳量; NEE: 湿地生态系统CO2净交换量; Fmethane: 湿地生态系统CH4排放通量的CO2当量通量值。
滨海感潮沼泽湿地全面的碳收支, 还包括潮汐和海水带来或带走的横向碳输入/出通量, 包括DOC、DIC、POC水平通量, 考虑到目前没有成熟的测量水平通量的方法, 故本研究在计算感潮沼泽湿地生态系统蓝碳碳汇时没有考虑横向的碳输入和碳输出。
1.7 统计分析数据处理采用Microsoft Excel 2016、IBM SPSS Statistics 21、Origin 2018进行数据的计算、分析以及制图。利用Excel 2016进行通量值及其平均值、标准误差计算, 利用SPSS软件进行CO2通量、CH4排放通量与各环境因子之间的相关性分析。图表及文字中的数据均以平均值±标准误差的形式表示。
2 结果 2.1 温度和PAR的变化2019年8月—2020年7月闽江河口鳝鱼滩湿地PAR日平均值波动范围为27.31—636.6 μmol m-2 s-1, 月平均值最高值出现在2019年8月(456.0 μmol m-2 s-1), 最低值出现在2020年1月(167.4μmol m-2 s-1), 夏季值PAR平均值明显高于冬季。日平均气温波动范围为8.91—32.4℃, 2020年7月份平均气温最高(30.2℃), 2020年2月份平均气温最低(12.7℃)(图 2)。

|
| 图 2 闽江河口鳝鱼滩日平均气温、PAR日均值和日最大值动态变化 Fig. 2 Variations of daily mean air temperature, daily mean and daily maximum photosynthetically active radiation in the Shanyutan, Minjiang River estuary |
芦苇沼泽各测定日正午瞬时GEE、ER和NEE变化如图 3所示。研究期间不同月份测定日正午时刻芦苇沼泽NEE具有明显的时间动态变化, 不同月份间差异显著(P < 0.001)。5个月份为负值, 表现为大气CO2的汇, 2019年8月NEE负值最小((-48.30±3.32)μmol m-2 s-1), 即芦苇沼泽从大气中净吸收(固定)CO2的量最大;4个月份为正值, 表现为大气CO2的一个“弱源”, 接近“中立”状况, 其中2020年7月, NEE正值最大((1.73±0.18)μmol m-2 s-1)。芦苇沼泽ER同样具有明显的时间动态变化, 不同月份间差异显著(P < 0.001)。2020年6月ER值最大((23.00±1.50)μmol m-2 s-1), 2019年12月ER值最小((1.67±0.33)μmol m-2 s-1)。芦苇沼泽GEE也呈明显的时间变化, 不同月份间差异显著(P < 0.001)。2020年12月GEE负值最大((-0.70±0.33)μmol m-2 s-1), 2019年8月GEE值最小((-67.65±5.49)μmol m-2 s-1)。研究期间芦苇沼泽所有测定日正午时段GEE、ER和NEE的均值分别为(-20.70±4.30)、(10.60±1.7)、(-10.46±3.36)μmol m-2 s-1。

|
| 图 3 闽江河口芦苇沼泽、短叶茳芏沼泽各测定日正午GEE、ER和NEE的动态变化 Fig. 3 Midday GEE, ER, and NEE in the P. australis marsh, and C. malaccensis marsh of the Minjiang River estuary |
短叶茳芏沼泽各测定日正午瞬时GEE、ER和NEE变化如图 3所示。研究期间不同月份测定日正午短叶茳芏沼泽NEE具有明显的时间动态变化, 不同月份间差异显著(P < 0.001)。各测定月份均为负值, 均表现为大气CO2的汇, 其中, 2019年9月NEE负值最小((-24.01±2.84)μmol m-2 s-1), 即该时刻短叶茳芏沼泽从大气中净吸收(固定)CO2的量最大。短叶茳芏湿地ER同样具有明显的时间动态变化, 不同月份间差异显著(P < 0.001), 2020年6月ER值最大((18.67±1.99)μmol m-2 s-1), 2019年12月ER值最小((2.16 ±0.06)μmol m-2 s-1)。短叶茳芏沼泽GEE值也呈明显的时间变化, 不同月份间差异显著(P < 0.001), 2019年12月GEE负值最大((-3.61±0.04)μmol m-2 s-1), 2020年6月GEE负值最小((-35.27±4.93)μmol m-2 s-1)。研究期间短叶茳芏沼泽所有测定日正午时段GEE、ER和NEE的均值分别为(-16.98±2.29)、(7.61±1.11)、(-9.38±1.66)μmol m-2 s-1。
独立样本T检验结果表明芦苇沼泽正午瞬时NEE值显著低于短叶茳芏沼泽(P < 0.05), 即具有较强的固碳能力, 芦苇沼泽正午瞬时ER值显著高于短叶茳芏湿地(P < 0.001), GEE与短叶茳芏沼泽差异不显著(P>0.05)。
2.3 芦苇沼泽、短叶茳芏沼泽CH4排放通量芦苇沼泽、短叶茳芏沼泽CH4排放通量的时间变化如图 4所示。芦苇沼泽CH4排放通量介于0.009—0.033 μmol m-2 s-1之间, 2019年8月CH4排放通量达到最大值, 2019年12月为最小值。短叶茳芏沼泽CH4排放通量介于0.005—0.030 μmol m-2 s-1之间, 2019年8月CH4排放通量达到最大值, 2019年12月为最小值。研究期间, 芦苇沼泽和短叶茳芏沼泽各测定日CH4排放通量均值分别为(0.019±0.002)、(0.015±0.002)μmol m-2 s-1。芦苇湿地不同月份间排放通量差异显著(P < 0.05)。芦苇沼泽与短叶茳芏沼泽甲烷排放通量差异不显著(P > 0.05)。

|
| 图 4 闽江河口芦苇沼泽和短叶茳芏沼泽CH4排放通量的时间变化 Fig. 4 Variations of the CH4 fluxes from the P. australis marsh and C. malaccensis marsh of the Minjiang River estuary |
对各月测定日芦苇沼泽、短叶茳芏沼泽GEE(这里将GEE转化为正值)与PAR的进行直角双曲线拟合[8], 图 5为7月份芦苇沼泽、短叶茳芏沼泽各自3个样地GEE与PAR的拟合曲线。研究时段所有月份测定日芦苇沼泽、短叶茳芏沼泽ER与大气温度的拟合曲线见图 6, 二者ER均与大气温度呈指数回归关系。

|
| 图 5 7月份测定日不同光照条件下芦苇沼泽、短叶茳芏沼泽3个样地GEE与PAR的拟合光响应曲线 Fig. 5 Light response curves created for the P. australis and C. malaccensis marshes in July at different light intensities |

|
| 图 6 所有月份测定日芦苇沼泽、短叶茳芏沼泽生态系统呼吸与气温的拟合曲线 Fig. 6 Relationship between ecosystem respiration of CO2 in the P. australis and C. malaccensis marshes and air temperature for the entire study period |
芦苇沼泽2019年8月至2020年7月各月累积的GEE、ER和NEE如图 7所示。研究期间各月芦苇沼泽NEE具有明显的时间动态变化, 不同月份间差异显著(P < 0.001), 在春、夏、秋季各月均表现为负值, 即这3个季节表现为大气CO2的汇, 只有在冬季11、12月则转化为大气CO2的源。2019年8月NEE值((-2997.79±210.77)g CO2/m2)最小, 即8月份芦苇沼泽大气CO2汇的功能最强, 2019年11月NEE值((134.20±27.64)g CO2/m2)最大, 表现为大气CO2源。各月份芦苇沼泽ER均为正值, 不同月份间差异显著(P < 0.001)。2020年7月ER值最大((778.47±36.26)g CO2/m2), 2020年2月ER值最小((117.13±5.89)g CO2/m2)。芦苇沼泽各月份间GEE差异显著(P < 0.001)。2019年12月GEE值最大((-48.07±31.93)g CO2/m2), 2019年8月GEE值最小((-3679.10±200.27)g CO2/m2)。研究期间, 芦苇沼泽GEE、ER和NEE月均值分别为(-830.1±166.67)、(364.97±39.48)、(-465.17±140.26)g CO2/m2。芦苇沼泽年GEE、ER和NEE分别为(-9961.76±544.59)、(4379.68±205.82)、(-5582.08±369.01)g CO2/m2。

|
| 图 7 芦苇沼泽、短叶茳芏沼泽月尺度整合的GEE、ER和NEE的动态变化 Fig. 7 Monthly-integrated rates of GEE, ER, and NEE for the P. australis mrsh, and C. malaccensis marsh of the Minjiang River estuary |
短叶茳芏沼泽2019年8月至2020年7月各月的GEE、ER和NEE如图 7所示。研究期间短叶茳芏沼泽各月份NEE具有明显的时间动态变化, 不同月份间差异显著(P < 0.001)。2019年4—7月及2020年8—10月NEE为负值, 在这些月份表现为大气CO2的汇, 而在11—1月的非生长季, 表现为正值, 即转化为大气CO2的源, 其中, 2020年6月NEE值最小((-830.06±228.33)g CO2/m2), 表现为最强的大气CO2汇, 2019年11月NEE值最大((132.15±25.68)g CO2/m2), 表现为较强的大气CO2源。各月份短叶茳芏沼泽ER均为正值, 研究期间变化趋势呈现冬季低, 夏季高, 不同月份间差异显著(P < 0.001)。2020年7月ER值最大((679.18±21.04)g CO2/m2), 2020年2月ER值最小((268.22±7.29)g CO2/m2)。短叶茳芏沼泽不同月份间GEE差异显著(P < 0.001)。2019年11月GEE值最大((-236.89±28.05)g CO2/m2), 2020年6月GEE值最小((-1444.13±223.76)g CO2/m2)。研究期间, 短叶茳芏沼泽GEE、ER和NEE的月均值分别为(-688.44±68.16)、(446.54±24.29)、(-241.90±51.96)g CO2/m2。短叶茳芏沼泽年GEE、ER和NEE分别为(-8261.30±417.69)、(5358.48±100.66)、(-2902.82±498.72)g CO2/m2。
独立样本T检验结果表明芦苇沼泽月GEE值显著低于短叶茳芏湿地(P < 0.01), 即其光合作用碳吸收能力强于短叶茳芏沼泽, 芦苇沼泽月ER值显著低于短叶茳芏沼泽(P < 0.01), 芦苇沼泽月NEE值显著低于短叶茳芏沼泽(P < 0.05), 即具有较强的固碳能力。
根据各月份测定日测定的芦苇沼泽、短叶茳芏沼泽CH4通量, 估算月CH4排放通量, 然后估算出2个沼泽CH4年排放通量分别为(9.52±1.32)、(6.90±1.41)g CH4/m2, 最后, 根据100年尺度下单位CH4的全球变暖潜势(GWP)是CO2的25倍, 将芦苇沼泽、短叶茳芏沼泽CH4年尺度排放通量转化为CO2当量排放通量, 计算结果为芦苇沼泽和短叶茳芏沼泽的年CH4排放通量分别为(210.56±32.04)g CO2-eq/m2和(172.50±35.19)g CO2-eq/m2。根据公式5, 计算得出闽江河口芦苇沼泽、短叶茳芏沼泽生态系统的年固碳量分别为(5371.52±336.97)g CO2-eq/m2和(2730.32±503.67)g CO2-eq/m2, 在年尺度上, 闽江河口芦苇沼泽、短叶茳芏沼泽生态系统均表现为较强的碳汇。
3 讨论 3.1 沼泽生态系统碳收支本研究发现:闽江河口半咸水芦苇沼泽和短叶茳芏沼泽NEE具有明显的季节变化, 在春夏表现为明显的大气中CO2的汇, 而在冬季则转化为大气中CO2的源, 在年尺度综合考虑NEE和CH4排放, 2个沼泽湿地均为较强的碳汇, 两者的年固碳量分别为(5371.52±336.97)g CO2-eq/m2和(2730.32±503.67)g CO2-eq/m2。
目前国际上对于内陆沼泽湿地碳收支的研究多集中在北方泥炭沼泽。Sottocornola and Kiely运用通量塔涡度协方差法测定了爱尔兰西南部毡状酸沼湿地的碳收支, 研究结果也表明NEE具有明显的季节变化, 5—9月为大气中CO2的汇, 11—1月份为大气中CO2的源, 3、4和10月份未显示为明显的CO2汇或源特征[4]。Hendriks等[25]利用通量塔涡度协方差法测定北方泥炭草甸CO2净通量, 利用静态箱法测定CH4排放通量, 结果表明:生长季尺度NEE表现为净吸收, 综合考虑泥炭草甸湿地CH4排放通量, 在100年尺度上泥炭草甸湿地年固碳量为86 g CO2-eq/m2, 是一个很弱的湿地碳汇。Waddington and Roulet[26]运用透明箱以及黑布遮盖方法测定了瑞典北部泥炭藓(Sphagnum)占优势的泥炭沼泽碳收支(包括NEE和CH4排放通量), 结果表明:1992年研究样地为碳汇(-2.0 g C/m2), 但是1993年却表现为一个碳源(7.6 g C/m2)。Bäckstrand等[7]运用自动箱法研究了瑞典北部土壤湿度和冻融梯度上分布的不同优势植物泥炭沼泽碳收支的变化, 结果表明:永久冻土干沼泽、中等融化的泥炭藓沼泽和基本解冻且土壤湿度较高的羊胡子泥炭沼泽(Eriophorum spp.)年净含碳气体收支(annual net C gas balance)分别为30、-29、-3.1 g C/m2, 永久冻土干沼泽为大气中CO2的源, 中等融化的泥炭藓沼泽和土壤湿度较高的羊胡子泥炭沼泽是大气中CO2的汇。从以上研究的结果可以看出:北方泥炭沼泽年尺度上基本表现为大气中CO2的汇, 但是综合考虑CH4排放, 其年固碳量规模均较小, 年固碳量远低于本研究中位于亚热带河口的芦苇沼泽和短叶茳芏沼泽。北方地区较低的年均气温以及较低的植物生产力是其年固碳量低的主要原因。
Guo等运用涡度协方差通量塔法研究了长江口崇明岛互花米草(Spartina)群落、芦苇群落(Phragmites)和藨草群落(Scirpus)镶嵌生态系统垂直方向CO2净交换通量与潮汐的关系, 但是并没有估算其年固碳量[6]。马安娜和陆健健运用开路式涡度相关系统测定了长江口崇明岛西滩芦苇沼泽8月份的NEE, 结果表明:芦苇沼泽8月份NEE平均值为-0.06 mg m-2 s-1[27], 远低于本研究中芦苇沼泽8月份的NEE均值(-2.13 mg m-2 s-1)。邢庆会等[17]利用涡度相关法测定了黄河口潮间盐地碱蓬(Suaeda salsa)湿地生长季(4—10月) NEE和ER的变化, 结果表明NEE具有明显的季节变化, 整个生长季表现为CO2的汇, NEE为-22.3 g C/m2。比较本研究与长江口芦苇沼泽、黄河口碱蓬湿地的研究结果, 可以发现闽江河口芦苇沼泽、短叶茳芏沼泽具有相对较高的碳汇能力。一个有趣的结果是Chu等[28]综合考虑NEE和CH4排放通量, 研究表明香蒲(Typha)和睡莲(Nymphaea)为优势种的淡水沼泽湿地CH4排放通量年均值为49.7 g C-CH4/m2, NEE年均值仅为-21.0 g C-CO2/m2, 即该淡水沼泽年尺度上表现为一个碳源。综上研究结果, 我们可以推断出以下结论:综合考虑CH4排放, 内陆泥炭沼泽以及滨海感潮沼泽生态系统的年尺度上的碳收支较为复杂, 具体表现为:(1)整体上看, 无论是内陆泥炭沼泽, 还是滨海感潮沼泽基本表现为碳汇;(2)可能存在着年际间碳源/汇功能的转化;(3)复杂的环境条件, 可能使某些沼泽为碳汇, 而另一些沼泽为碳源。
3.2 植物及环境因子对沼泽生态系统碳通量影响Waddington and Roulet[26]测定了北方泥炭藓(Sphagnum)沼泽的NEE、GEE和CH4排放通量, 结果发现锈色泥炭藓(Sphagnum fuscum)占优势的泥炭沼泽以上碳通量值与泥炭藓(Sphagnum majus)占优势的泥炭沼泽明显不同, 锈色泥炭藓泥炭沼泽GEE值高于泥炭藓沼泽。本研究中芦苇沼泽年NEE负值显著小于短叶茳芏沼泽, 即芦苇沼泽的碳汇强度显著高于短叶茳芏沼泽。我们推测可能与2种植物的生理生态特征有关, 短叶茳芏的叶片很窄, 宽度仅为3—8 mm, 主要靠叶鞘和茎进行光合作用, 其光合作用能力可能低于芦苇植株。芦苇沼泽各测定日正午ER值显著高于短叶茳芏沼泽, 这与仝川等[29]对闽江河口潮汐沼泽的研究结果一致。但本研究中发现把ER日瞬时值尺度上移至月尺度, 芦苇沼泽ER月值却低于短叶茳芏沼泽, 这可能是由于二者的温度敏感性不同导致(图 6)。在年尺度上, 芦苇沼泽因其光合作用碳吸收量高于于短叶茳芏沼泽, 而其生态系统呼吸低于短叶茳芏沼泽, 综合考虑, 短叶茳芏沼泽固碳能力弱于芦苇沼泽。
本研究中, 芦苇沼泽、短叶茳芏沼泽各测定日正午ER表现出相似的月动态规律, 虽然测定日正午瞬时ER与月值最大值、最小值出现的月份不一致, 但均表现为夏季强, 冬季弱。2个沼泽测定日正午瞬时ER均以6月最大, 12月最小, 而月值则均以7月最大, 2月最小。本研究中无论正午时刻的ER还是月尺度整合的ER, 均表现为夏季强, 冬季弱。这与多数研究结果相同[18, 30—31]。温度是影响ER的环境因子之一, 湿地生态系统ER随土壤温度的升高而增加的结果在较多的文献中得到报道[17, 32—35], 本研究中, 芦苇测定日ER与大气温度(P < 0.001, r=0.889), 土壤温度(P < 0.001, r=0.955)均具有显著的正相关关系, 短叶茳芏沼泽测定日ER与大气温度(P < 0.001, r=0.642), 土壤温度(P < 0.001, r = 0.787)均具有显著的正相关关系。土壤酶活性及微生物活性是影响生态系统呼吸的主要环境因子, 随着温度的升高, 土壤酶活性及微生物活性均增加[35], 进而造成生态系统呼吸的增加。
本研究中, 芦苇沼泽测定日正午瞬时GEE值、月值均以12月最大, 8月最小;短叶茳芏沼泽测定日正午瞬时GEE值、月值均以6月最小, 最大值分别出现在12月和11月。虽然最大、最小值出现月份略有不同, 但总体上表现为植物生长季光合能力大于非生长季。2个沼泽月尺度整合的NEE仅在11月和12月表现为碳源, 其余月份均为碳汇。就正午瞬时通量而言, 短叶茳芏沼泽正午NEE在所有观测月中均表现为负值, 均具有固碳能力。而当上移整合到月尺度, 在11月至3月的非生长季, 短叶茳芏湿地基本表现为碳源, 主要可能原因与短叶茳芏在非生长季大量倒伏, 植物光合作用受限有关。
本研究中2个沼泽生态系统GEE与PAR均较好地符合直角双曲线关系(图 5), 这与以往很多研究结果一致[9, 18]。但是, 两个沼泽的GEE与PAR拟合光响应曲线趋势不一样, 分析原因可能是由于芦苇为C3植物, 短叶茳芏为C4植物, 通常C4植物的光饱和点高于C3植物, 因而芦苇较早到达光饱和状态, 从而导致两者GEE与PAR的拟合光响应曲线趋势不一致。光合有效辐射是影响植物光合作用的关键因素, PAR对沼泽生态系统NEE, GEE等碳收支关键过程均具有重要影响[5, 13, 36—37]。
本研究中, 芦苇沼泽与短叶茳芏沼泽CH4排放通量差异不显著。Tong等[36]在闽江河口连续3年的观测发现芦苇沼泽与短叶茳芏沼泽CH4排放通量在不同年份表现出芦苇沼泽CH4排放通量大于短叶茳芏沼泽或两者差异不显著。温度是影响沼泽湿地CH4排放通量的关键环境因子。本研究中, 芦苇沼泽CH4排放通量与大气温度(P < 0.01, r=0.504), 土壤温度(P < 0.05, r=0.513)均具有显著的正相关关系;但是, 短叶茳芏沼泽CH4排放通量与大气温度, 土壤温度相关性均不显著(P > 0.05), 原因尚不清楚。一般认为沼泽湿地CH4排放通量通常随温度升高而升高[38], 其原因包括温度影响沼泽湿地土壤产甲烷微生物活性[39], 提高甲烷产生速率从而提高CH4排放通量。
4 结论闽江河口半咸水芦苇沼泽、短叶茳芏沼泽CO2通量、CH4排放通量均具有明显的季节变化特征。2个沼泽湿地除冬季表现为大气中CO2的源, 春、夏、秋季均为大气中CO2的汇。综合考虑CH4排放通量, 闽江河口半咸水芦苇沼泽、短叶茳芏沼泽生态系统年固碳量分别为(5371.52±336.97)g CO2-eq/m2和(2730.32±503.67)g CO2-eq/m2, 均具有很强的碳汇功能。因此, 在我国滨海湿地生态保护与生态恢复实践中, 对于滨海感潮沼泽应给予更多的关注和保护。
| [1] |
Howard J, Sutton-Grier A, Herr D, Kleypas J, Landis E, Mcleod E, Pidgeon E, Simpson S. Clarifying the role of coastal and marine systems in climate mitigation. Frontiers in Ecology and the Environment, 2017, 15(1): 42-50. DOI:10.1002/fee.1451 |
| [2] |
章海波, 骆永明, 刘兴华, 付传城. 海岸带蓝碳研究及其展望. 中国科学: 地球科学, 2015, 45(11): 1641-1648. |
| [3] |
Macreadie P I, Costa M D P, Atwood T B, Friess D A, Kelleway J J, Kennedy H, Lovelock C E, Serrano O, Duarte C M. Blue carbon as a natural climate solution. Nature Reviews Earth & Environment, 2021, 2(12): 826-839. |
| [4] |
Sottocornola M, Kiely G. An Atlantic blanket bog is a modest CO2 sink. Geophysical Research Letters, 2005, 32(23): L23804. DOI:10.1029/2005GL024731 |
| [5] |
Bonneville M C, Strachan I B, Humphreys E R, Roulet N T. Net ecosystem CO2 exchange in a temperate cattail marsh in relation to biophysical properties. Agricultural and Forest Meteorology, 2008, 148(1): 69-81. DOI:10.1016/j.agrformet.2007.09.004 |
| [6] |
Guo H Q, Noormets A, Zhao B, Chen J Q, Sun G, Gu Y J, Li B, Chen J K. Tidal effects on net ecosystem exchange of carbon in an estuarine wetland. Agricultural and Forest Meteorology, 2009, 149(11): 1820-1828. DOI:10.1016/j.agrformet.2009.06.010 |
| [7] |
Bäckstrand K, Crill P M, Jackowicz-Korczyñski M, Mastepanov M, Christensen T R, Bastviken D. Annual carbon gas budget for a subarctic peatland, Northern Sweden. Biogeosciences, 2010, 7(1): 95-108. DOI:10.5194/bg-7-95-2010 |
| [8] |
Golovatskaya E A, Dyukarev E A. Carbon budget of oligotrophic mire sites in the Southern Taiga of Western Siberia. Plant and Soil, 2009, 315(1/2): 19-34. |
| [9] |
Wilson B J, Mortazavi B, Kiene R P. Spatial and temporal variability in carbon dioxide and methane exchange at three coastal marshes along a salinity gradient in a Northern Gulf of Mexico estuary. Biogeochemistry, 2015, 123(3): 329-347. DOI:10.1007/s10533-015-0085-4 |
| [10] |
Järveoja J, Nilsson M B, Gažovič M, Crill P M, Peichl M. Partitioning of the net CO2 exchange using an automated chamber system reveals plant phenology as key control of production and respiration fluxes in a boreal peatland. Global Change Biology, 2018, 24(8): 3436-3451. DOI:10.1111/gcb.14292 |
| [11] |
贺文君, 韩广轩, 许延宁, 张希涛, 王安东, 车纯广, 孙宝玉, 张孝帅. 潮汐作用下干湿交替对黄河三角洲盐沼湿地净生态系统CO2交换的影响. 应用生态学报, 2018, 29(1): 269-277. |
| [12] |
Yan Y E, Guo H Q, Gao Y, Zhao B, Chen J Q, Li B, Chen J K. Variations of net ecosystem CO2 exchange in a tidal inundated wetland: coupling MODIS and tower-based fluxes. Journal of Geophysical Research: Atmospheres, 2010, 115(D15): D15102. DOI:10.1029/2009JD012838 |
| [13] |
Vázquez-Lule A, Vargas R. Biophysical drivers of net ecosystem and methane exchange across phenological phases in a tidal salt marsh. Agricultural and Forest Meteorology, 2021, 300: 108309. DOI:10.1016/j.agrformet.2020.108309 |
| [14] |
仝川, 姚顺, 王维奇, 黄佳芳, 张林海, 章文龙, 曾从盛. 中国东南沿海短叶茳芏潮汐沼泽湿地甲烷动态. 中国科学: 地球科学, 2012, 42(5): 723-735. |
| [15] |
仝川, 曾从盛, 王维奇, 闫宗平, 杨红玉. 闽江河口芦苇潮汐湿地甲烷通量及主要影响因子. 环境科学学报, 2009, 29(1): 207-216. DOI:10.13671/j.hjkxxb.2009.01.022 |
| [16] |
Hirota M, Tang Y H, Hu Q W, Hirata S, Kato T, Mo W H, Cao G M, Mariko S. Methane emissions from different vegetation zones in a Qinghai-Tibetan Plateau wetland. Soil Biology and Biochemistry, 2004, 36(5): 737-748. DOI:10.1016/j.soilbio.2003.12.009 |
| [17] |
Tong C, Morris J T, Huang J F, Xu H, Wan S A. Changes in pore-water chemistry and methane emission following the invasion of Spartina alterniflora into an oliogohaline marsh. Limnology and Oceanography, 2018, 63(1): 384-396. DOI:10.1002/lno.10637 |
| [18] |
Han G X, Xing Q H, Yu J B, Luo Y Q, Li D J, Yang L Q, Wang G M, Mao P L, Xie B H, Mikle N. Agricultural reclamation effects on ecosystem CO2 exchange of a coastal wetland in the Yellow River Delta. Agriculture, Ecosystems & Environment, 2014, 196: 187-198. |
| [19] |
邢庆会, 韩广轩, 于君宝, 吴立新, 杨利琼, 毛培利, 王光美, 谢宝华. 黄河口潮间盐沼湿地生长季净生态系统CO2交换特征及其影响因素. 生态学报, 2014, 34(17): 4966-4979. |
| [20] |
Lloyd C R. Annual carbon balance of a managed wetland meadow in the Somerset Levels, UK. Agricultural and Forest Meteorology, 2006, 138(1/4): 168-179. |
| [21] |
Urbanski S, Barford C, Wofsy S, Kucharik C, Pyle E, Budney J, Mckain K, Fitzjarrald D, Czikowsky M, Munger J W. Factors controlling CO2 exchange on timescales from hourly to decadal at Harvard Forest. Journal of Geophysical Research: Biogeosciences, 2007, 112(G2): G02020. |
| [22] |
Malone S L, Starr G, Staudhammer C L, Ryan M G. Effects of simulated drought on the carbon balance of everglades short-hydroperiod marsh. Global Change Biology, 2013, 19(8): 2511-2523. DOI:10.1111/gcb.12211 |
| [23] |
Whiting G J, Bartlett D S, Fan S M, Bakwin P S, Wofsy S C. Biosphere/atmosphere CO2 exchange in tundra ecosystems: community characteristics and relationships with multispectral surface reflectance. Journal of Geophysical Research: Atmospheres, 1992, 97(D15): 16671-16680. DOI:10.1029/91JD01027 |
| [24] |
Miller W D, Neubauer S C, Anderson I C. Effects of sea level induced disturbances on high salt marsh metabolism. Estuaries, 2001, 24(3): 357-367. DOI:10.2307/1353238 |
| [25] |
Hendriks D M D, Van Huissteden J, Dolman A J, Van Der Molen M K. The full greenhouse gas balance of an abandoned peat meadow. Biogeosciences, 2007, 4(3): 411-424. DOI:10.5194/bg-4-411-2007 |
| [26] |
Waddington J M, Roulet N T. Carbon balance of a boreal patterned peatland. Global Change Biology, 2000, 6(1): 87-97. DOI:10.1046/j.1365-2486.2000.00283.x |
| [27] |
马安娜, 陆健健. 长江口崇西湿地生态系统的二氧化碳交换及潮汐影响. 环境科学研究, 2011, 24(7): 716-721. |
| [28] |
Chu H S, Chen J Q, Gottgens J F, Ouyang Z T, John R, Czajkowski K, Becker R. Net ecosystem methane and carbon dioxide exchanges in a Lake Erie coastal marsh and a nearby cropland. Journal of Geophysical Research: Biogeosciences, 2014, 119(5): 722-740. DOI:10.1002/2013JG002520 |
| [29] |
仝川, 鄂焱, 廖稷, 姚顺, 王维奇, 黄佳芳, 张林海, 杨红玉, 曾从盛. 闽江河口潮汐沼泽湿地CO2排放通量特征. 环境科学学报, 2011, 31(12): 2830-2840. |
| [30] |
Miao G F, Noormets A, Domec J C, Fuentes M, Trettin C C, Sun G, Mcnulty S G, King J S. Hydrology and microtopography control carbon dynamics in wetlands: implications in partitioning ecosystem respiration in a coastal plain forested wetland. Agricultural and Forest Meteorology, 2017, 247: 343-355. DOI:10.1016/j.agrformet.2017.08.022 |
| [31] |
Song C C, Xu X F, Tian H Q, Wang Y Y. Ecosystem-atmosphere exchange of CH4 and N2O and ecosystem respiration in wetlands in the Sanjiang Plain, Northeastern China. Global Change Biology, 2009, 15(3): 692-705. DOI:10.1111/j.1365-2486.2008.01821.x |
| [32] |
杨利琼, 韩广轩, 于君宝, 吴立新, 朱敏, 邢庆会, 王光美, 毛培利. 黄河三角洲芦苇湿地生长季净生态系统CO2交换及其环境调控机制. 应用生态学报, 2013, 24(9): 2415-2422. |
| [33] |
Zhang F W, Liu A H, Li Y N, Zhao L, Wang Q X, Du M Y. CO2 flux in alpine wetland ecosystem on the Qinghai-Tibetan Plateau, China. Acta Ecologica Sinica, 2008, 28(2): 453-462. DOI:10.1016/S1872-2032(08)60024-4 |
| [34] |
Wang C L, Yu G R, Zhou G Y, Yan J H, Zhang L M, Wang X, Tang X L, Su X M. CO2 flux evaluation over the evergreen coniferous and broad-leaved mixed forest in Dinghushan, China. Science in China Series D: Earth Sciences, 2006, 49(2): 127-138. |
| [35] |
Cox P M, Betts R A, Jones C D, Spall S A, Totterdell I J. Acceleration of global warming due to carbon-cycle feedbacks in a coupled climate model. Nature, 2000, 408(6809): 184-187. DOI:10.1038/35041539 |
| [36] |
Yan Y E, Zhao B, Chen J Q, Guo H Q, Gu Y J, Wu Q H, Li B. Closing the carbon budget of estuarine wetlands with tower-based measurements and MODIS time series. Global Change Biology, 2008, 14(7): 1690-1702. DOI:10.1111/j.1365-2486.2008.01589.x |
| [37] |
Gomez-Casanovas N, Delucia N J, Delucia E H, Blanc-Betes E, Boughton E H, Sparks J, Bernacchi C J. Seasonal controls of CO2 and CH4 dynamics in a temporarily flooded subtropical wetland. Journal of Geophysical Research: Biogeosciences, 2020, 125(3): e2019JG005257. |
| [38] |
Tong C, Wang W Q, Huang J F, Gauci V, Zhang L H, Zeng C S. Invasive alien plants increase CH4 emissions from a subtropical tidal estuarine wetland. Biogeochemistry, 2012, 111(1/3): 677-693. |
| [39] |
Deng Y C, Liu P F, Conrad R. Effect of temperature on the microbial community responsible for methane production in alkaline NamCo wetland soil. Soil Biology and Biochemistry, 2019, 132: 69-79. DOI:10.1016/j.soilbio.2019.01.024 |