 2022, Vol. 42
2022, Vol. 42文章信息
- 宋思意, 吕思扬, 邱岭军, 王馨逸, 涂利华
- SONG Siyi, Lü Siyang, QIU Lingjun, WANG Xinyi, TU Lihua
- 华西雨屏区常绿阔叶林不同深度土壤氮矿化及酶活性对模拟氮沉降的响应
- Responses of soil nitrogen mineralization and enzyme activities at different depths to nitrogen additions in an evergreen broad-leaved forest
- 生态学报. 2022, 42(22): 9045-9056
- Acta Ecologica Sinica. 2022, 42(22): 9045-9056
- http://dx.doi.org/10.5846/stxb202107211971
-
文章历史
- 收稿日期: 2021-07-21
- 网络出版日期: 2022-07-12





2. 四川农业大学林学院, 长江上游林业生态工程四川省重点实验室, 成都 611130;
3. 中国共产党青神县委员会, 眉山 620000;
4. 四川省眉山市东坡区岷江现代农业示范园区管理委员会, 眉山 620000
2. Sichuan Province Key Laboratory of Ecological Forestry Engineering on the Upper Reaches of the Yangtze River, College of Forestry, Sichuan Agricultural University, Chengdu 611130, China;
3. Qingshen County Committee of The Communist Party of China, Meishan 620000, China;
4. Management Committee of Minjiang Modern Agriculture Demonstration Park, Dongpo District, Meishan 620000, China
过去几十年, 由于人口快速增长、化石燃料的大量燃烧和氮肥过量使用导致向大气输入的活性氮急剧增加, 同时, 通过氮沉降的方式向输入陆地的外源活性氮也相应增加, 这强烈改变了陆地生态系统N循环模式, 可能最终会改变陆地生态系统结构和功能[1]。中国全球氮沉降三氮沉降区之一, 当前的大气平均氮沉降速率远高于全球氮临界负荷水平, 氮沉降区域逐渐从温带向热带亚热带扩展[2]。持续增加的氮沉降导致热带亚热带森林土壤趋近氮饱和状况, 引发的生态环境变化和生物多样性等问题成为生态学家持续关注的热点问题。
土壤氮矿化作用是指土壤有机氮转化为无机氮的过程[3]。氮矿化是森林生态系统氮循环的重要环节之一, 影响着氮素生物地球化学循环和生态系统功能。森林植物所获取的氮素大部分来自土壤[4], 而土壤中的绝大部分氮素需经土壤微生物或动物的分解作用转化为无机态氮, 即氮矿化过程, 才可以被植物吸收利用。目前, 众多模拟氮沉降对土壤氮矿化的影响还没有统一定论, 多数研究表明, 森林土壤氮矿化速率随氮沉降量增加而增加;而部分研究发现, 土壤氮矿化速率先增加后降低, 呈n型, 在模拟氮沉降的初期快速增加到达峰值后开始缓慢降低并逐渐接近对照组水平[5], 即森林土壤氮矿化速率并不随氮沉降增加而持续性加快, 两者并非简单的线性正相关关系[6]。氮沉降的增加不会持续加快氮矿化速率的主要原因是:一方面氮沉降的增加改变了土壤有机质的结构, 减少了土壤微生物分解作用所产生的腐殖质酶量, 使土壤氮矿化相关酶活性降低[7—8], 另一方面氮沉降的增加使得土壤酸化, 土壤氮素淋溶作用增强, 导致土壤有效氮流失, 最终导致土壤氮矿化速率降低。
土壤酶是指土壤中能够催化土壤有机物质的合成与分解反应, 促使土壤养分转化的一类物质, 大多数土壤酶都是特殊的蛋白质类化合物[9]。前人研究发现, 氮沉降对不同生态系统中土壤酶活性影响不同[10]。氮沉降对森林土壤酶活性的影响取决于氮沉降水平、土壤类型、土壤养分、土壤通气性、酶的种类等多种因素[2]。现有研究表明, 氮沉降对森林土壤酶活性的影响机制主要有3方面:(1)氮沉降改变土壤pH值, 影响土壤酶参与反应时的pH值区间, 从而影响森林土壤酶活性;(2)氮沉降通过影响土壤中可利用的养分的种类和含量, 来影响森林土壤酶活性;(3)氮沉降通过改变森林凋落物的输入来影响森林土壤酶活性。土壤氮转化酶是具有特殊生物催化能力的蛋白质, 能促进有机氮转化成植物能直接利用的有效氮[1], 因此氮转化酶是限制森林生产力的重要因素。土壤脲酶、蛋白酶、硝酸还原酶等氮转化酶活性与土壤氮矿化速率以及土壤硝化、反硝化和氨的挥发速率相关[2—3]。不同森林群落, 凋落物基质质量、腐殖质形态、根系系统和吸收营养方式都存在很大差异[4—6], 这些因素可能影响不同森土壤氮转化酶活性, 进而影响土壤氮循环[7]。
课题组以往研究表明, “华西雨屏”区由于区位、地形等原因, 其年均氮湿沉降量高达95 kg N hm-2 a-1, 处于中国最高水平。并且由于氮沉降的空间异质性, 该区域未来几十年氮沉降仍将处于极高水平。本研究样地位于华西雨屏区中心地带, 气候属亚热带湿润季风型山地气候, 年均气温为16.2 ℃, 常年雨水充沛。试验样地自2014年起已经进行7年施氮处理, 长期性外源氮添加使得该森林生态系统出现根下层土壤硝态氮淋溶等氮饱和特征[11]。
土壤深度对土壤氮矿化作用及土壤酶活性的影响不容忽视。研究发现, 0—120 cm范围内不同土壤深度的土壤氮矿化速率明显不同[12]。森林土壤氮矿化速率和土壤酶活性受多种因素的影响, 土壤深度往往间接作用于这些因素, 从而影响土壤氮矿化速率和土壤酶活性。在森林土壤中, 无机氮的垂直移动速率较快, 因此大气氮沉降极有可能对下层森林土壤造成较大影响, 但目前国内外关于大气氮沉降对森林土壤氮循环过程的研究, 多集中在表层土壤。基于以上认识, 本研究——华西雨屏区常绿阔叶林不同深度土壤氮矿化及相关土壤酶活性对模拟氮沉降的响应, 通过模拟在大气氮沉降增加情景下, 研究不同深度土壤氮矿化速率及氮循环相关酶活性对模拟氮沉降的响应及其机制。
1 材料与方法 1.1 试验地概况试验地位于四川省雅安市雨城区碧峰峡(103°00′E, 30°04′N), 该地区气候温和湿润, 属亚热带湿润季风型山地气候, 年均气温为16.2℃。日照时间1040 h, 全年太阳辐射总量3640 MJ/cm2。年均降水量、年蒸发量分别为1772 mm、1011 mm, 全年相对湿度在80%左右。试验地为碧峰峡自然保护区千家山林班, 为亚热带湿性常绿阔叶次生林, 海拔约1026 m, 土壤类型为山地黄壤, 土壤厚度大于60 cm, 研究区内林地条件基本一致。主要树种有木荷(Schima superba)、红淡比(Cleyera japonica)、木姜子(Litsea cubeba)、总状山矾(Symplocos botryantha)、四川山矾(Symplocos setchuensis)、栲(Castanopsis fargesii )、细枝柃(Eurya loquaiana)、白栎(Quercus fabri )、杉木(Cunninghamia lanceolata)等。
1.2 样地设置与N添加处理2013年11月, 根据典型性和代表性原则在四川碧峰峡自然保护区千家山林班设置12块20 m×20 m的样地。用硝酸铵(NH4NO3)模拟氮沉降处理, 共设对照(CK, +0 g N hm-2 a-1)、低氮(LN, +5 g N hm-2 a-1)、高氮(HN, +15 g N hm-2 a-1)3个氮添加水平, 3种处理各4个重复。年施用量均分成12等份。于2014年1月开始, 每月下旬进行定量模拟N沉降处理, 将各样地所需NH4NO3溶解在10 L清水中, 用喷雾器在样地中来回均匀喷洒, CK喷洒等量清水, 对照和处理所用清水均取自同一位置的山间溪水。2017年起, 用NH4Cl和NaNO3混合物代替NH4NO3施肥处理。
1.3 样品采集与培养于2019年7月进行采样。为保证土壤样品具有代表性, 在每块样地中随机选取6个采样点, 每个采样点用土钻取样, 按0—15 cm(上层)、15—30 cm(中层)、30—45 cm(下层)将土壤分层采样。将土壤编号后快速带回实验室。
带回土壤分为需要进行室内培养和不需要进行室内培养的两份。不需进行室内培养的土壤分出一部分挑除可见根、石砾和动物残体后过2 mm筛, 编号装入密封袋, 4℃冰箱贮藏, 尽快对土壤酶活性、硝态氮(nitrate nitrogen, NO3--N)和铵态氮(ammonium nitrogen, NH4+-N)进行测定;另一部分土壤平铺于室内, 自然风干后, 挑除杂质, 过2 mm和0.25 mm筛, 分别编号装入密封袋, 用于测定土壤总有机碳(total organic carbon, TOC)和全氮(total nitrogen, TN)。需要进行室内培养的土壤, 保持其土柱形态, 装入聚乙烯塑料瓶中, 用透气不透水的保鲜膜封口, 置于(20±1)℃恒温培养箱中培养60 d, 培养结束后挑除可见根、石砾和动物残体后过2 mm筛, 编号装入密封袋, 立即测定各指标。
1.4 指标测定测定土壤氮矿化相关指标有:TN、NO3--N、NH4+-N。分别采用半微量凯氏法测定(LY/T 1228—1999)、紫外分光光度法测定、2 mol/L氯化钾(KCl)浸提-靛酚蓝(C18H16N2O)比色法[13]测定。各指标均做3次平行测定。TOC含量参考LY/T 1237—1999, 采用重铬酸钾-硫酸溶液氧化法测定。
测定的土壤酶有:脲酶、蛋白酶、过氧化氢酶和硝酸还原酶。分别采用邻苯三酚(C6H6O3)比色法[14]、加勒斯江比色法、高锰酸钾滴定法测定。硝酸还原酶利用硝酸钾为底物, 厌氧条件下30 ℃恒温培养24 h, 硝态氮还原为亚硝态氮, 产生的亚硝酸根与显色剂反应, 520nm处比色测定酶促反应后生成的亚硝态氮的量, 从而表征硝酸还原酶的活性[15]。
1.5 数据处理与分析土壤氮矿化速率相关计算公式为:
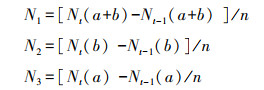
|
式中, N1:土壤净氮矿化速率(mg kg-1 d-1);N2:土壤净氮硝化速率(mg kg-1 d-1);N3:土壤净氮氨化速率(mg kg-1 d-1);Nt:培养60天后铵态氮和硝态氮, Nt-1 :培养前铵态氮和硝态氮;n: 培养天数;a:铵态氮含量(mg/kg);b:硝态氮含量(mg/kg)。
用Excel 2007(Microsoft Corp. Redmond, USA)对试验数据进行整理、均值计算, 用SPSS 20.0(IBM SPSS Inc. Chicago, USA)进行数据统计分析。利用双因素方差分析不同氮处理、不同土壤深度对土壤生化特性各指标的影响。利用Pearson相关分析对各指标进行相关性分析。利用Origin 2018 (Electronic Arts Inc, USA) 进行绘图。统计显著水平设置为α=0.05。
2 结果与分析 2.1 土壤TOC在同一氮处理水平下, 土壤TOC含量随土壤深度的增加而减少, 表现为上层>中层>下层, 三层土壤TOC含量之间有显著差异。在同一土壤深度下, 相较于CK, LN处理降低了土壤TOC含量, HN处理则增加了土壤TOC含量, 但影响均不显著。氮沉降处理和土壤深度的交互效应不显著(图 1)。

|
| 图 1 模拟氮沉降对不同深度土壤碳、氮组分的影响 Fig. 1 Effects of simulated nitrogen deposition on soil carbon and nitrogen components at different depths 不同字母表示处理间差异显著性, 大写字母为同一氮处理下不同土壤深度间的差异显著性, 小写字母为同一土壤深度下不同氮处理间的差异显著性(双因素方差分析, α=0.05);CK:对照 Control Treatment;LN:低氮 Low-Nitrogen Treatment;HN:高氮 High-Nitrogen Treatment; **, P < 0.05, ***, P < 0.01 |
在同一氮处理水平下, 土壤TN含量随土壤深度的增加而减少, 表现为上层>中层>下层, 三层土壤TN含量之间有极显著差异。在同一土壤深度下, 氮沉降处理对土壤TN含量无显著影响;氮沉降处理和土壤深度的交互效应不显著(图 1)。
2.3 土壤硝态氮、铵态氮在同一氮处理水平下, 恒温培养前的土壤硝态氮含量随土壤深度的增加而减少, 表现为上层>中层>下层;在同一土壤深度下, 土壤硝态氮含量随施氮处理的增加而增加, 表现为HN>LN>CK, 氮沉降处理对土壤硝态氮含量有显著影响, 但不同土壤深度对土壤硝态氮含量无显著影响, 两者之间交互效应不显著。在同一氮处理水平下, 恒温培养后的土壤硝态氮含量整体上随土壤深度的增加而减少, 表现为上层>中层>下层, 但在CK处理下, 中层较上层、下层分别增加了135%、474%, 在LN、HN处理下, 上层则远高于中层、下层, 三层土壤硝态氮含量之间有极显著差异;在同一土壤深度下, 上层、下层土壤硝态氮含量均表现为LN>CK>HN, 中层土壤则表现为CK>HN>LN, 氮沉降处理对土壤硝态氮含量影响极显著, 氮沉降处理和土壤深度的交互效应极显著(图 1)。
在同一氮处理水平下, 恒温培养前的土壤铵态氮含量表现为上层>下层>中层, 土壤深度对土壤铵态氮含量的影响极显著;在同一土壤深度下, 土壤铵态氮含量表现为HN>LN>CK, 氮沉降处理对土壤铵态氮含量影响极显著, 氮沉降处理和土壤深度的交互效应不显著。在同一氮处理水平下, 恒温培养后的土壤铵态氮含量表现为上层>下层>中层, 土壤深度对土壤铵态氮含量的影响不显著;在同一土壤深度下, 土壤铵态氮含量表现为HN>CK>LN, 氮沉降处理对土壤铵态氮含量影响不显著, 氮沉降处理和土壤深度的交互效应不显著(图 1)。
2.4 不同深度土壤氮矿化在同一氮处理水平下, 土壤净氮硝化速率整体上大致随土壤深度增加而减缓, 表现为上层>中层>下层, 但在CK处理下则表现为中层>上层>下层, 不同土壤深度土壤净氮硝化速率之间差异显著。A、C两层土壤土壤净氮硝化速率表现为LN>CK>HN, LN处理促进了表层和深层土壤净氮硝化速率, HN处理则抑制了表层和深层土壤净氮硝化速率, 中层土壤异于A、C两层土壤, 其土壤净氮硝化速率表现为CK>LN>HN, 氮沉降处理对土壤净氮硝化速率影响显著, 氮沉降处理和土壤深度的交互效应极显著(图 2)。

|
| 图 2 模拟氮沉降对不同深度土壤净氮硝化、氨化、矿化速率的影响 Fig. 2 Effects of simulated nitrogen deposition on nitrification rate, ammoniation rate and mineralization rate of soil net nitrogen at different depths |
在同一氮处理水平下, 土壤净氮氨化速率随土壤深度增加而减缓, 表现为上层>中层>下层, 不同土壤深度土壤净氮氨化速率之间差异不显著。在同一土壤深度下, 土壤净氮氨化速率表现为CK>HN>LN, 氮沉降处理抑制了土壤净氮氨化速率, LN处理对土壤净氮氨化速率抑制效果强, 土壤净氮氨化速率在高氮处理与自然状态下差别不大, 氮沉降处理对土壤净氮氨化速率影响不显著, 氮沉降处理和土壤深度的交互效应不显著(图 2)。
CK处理下, 土壤氮矿化速率表现为中层>上层>下层, LN处理下表现为上层>下层>中层, HN处理下则表现为上层>中层>下层, 三种施氮处理下土壤矿化速率随土壤深度的变化不同, LN处理下中层土壤氮矿化速率受到明显抑制作用, HN处理下中层和下层土壤氮矿化速率均受抑制, 不同土壤深度对土壤氮矿化速率影响显著。在同一土壤深度下, 氮处理降低了土壤氮矿化速率, 中层土壤中, HN处理下土壤氮矿化速率较CK处理下降低了196%, 氮沉降处理对土壤氮矿化速率影响极显著, 氮沉降处理和土壤深度的交互效应极显著(图 2)。
2.5 不同深度土壤酶活性在同一氮处理水平下, 土壤脲酶活性随土壤深度的增加而降低, 表现为下层 < 中层 < 上层, 不同土壤深度对土壤脲酶活性影响极显著。在同一土壤深度下, 土壤脲酶活性随氮添加的增多而增强, 表现为HN>LN>CK, 氮沉降处理对土壤脲酶活性影响极显著, 氮沉降处理和土壤深度的交互效应极显著(图 3)。

|
| 图 3 模拟氮沉降对不同深度土壤酶活性的影响 Fig. 3 Effects of simulated nitrogen deposition on soil enzyme activities at different depths |
在同一氮处理水平下, 土壤硝酸还原酶活性随土壤深度增加而降低, 表现为下层 < 中层 < 上层, 不同土壤深度对土壤硝酸还原酶活性影响极显著。在同一土壤深度下, 土壤硝酸还原酶活性随氮添加增多而降低, 表现为CK>LN>HN, 氮沉降处理对土壤硝酸还原酶活性影响显著, 氮沉降处理和土壤深度的交互效应不显著(图 3)。
在CK处理下, 土壤蛋白酶活性表现为中层>上层>下层, LN处理下表现为上层>下层>中层, HN处理下则表现为上层>中层>下层, 不同土壤深度对土壤蛋白酶活性影响不显著。上层土壤中土壤蛋白酶活性表现为HN>LN>CK, 中层土壤中表现为CK>HN>LN, 下层土壤中表现为LN>CK>HN, 氮沉降处理对土壤蛋白酶活性影响不显著, 氮沉降处理和土壤深度的交互效应不显著(图 3)。
在同一氮处理水平下, 土壤过氧化氢酶活性随土壤深度的加深而降低, 表现为上层>中层>下层, 不同土壤深度对土壤过氧化氢酶活性影响极显著。在同一土壤深度, 氮沉降处理对土壤过氧化氢酶活性的影响不显著, 氮沉降处理和土壤深度的交互效应不显著(图 3)。
2.6 土壤酶活性与土壤氮矿化速率之间的相关性相关性分析结果表明(表 1), 下层土壤中, 净氮硝化速率、净氮矿化速率均与蛋白酶活性呈现显著正相关关系。上、中两层土壤中并未表现出上述相关系。三层土壤中, 其他三种酶活性与氮矿化速率之间相关性并不显著。
| 土层 Soil layer |
硝酸还原酶活性 Nitrate reductase activity |
脲酶活性 Urease activity |
蛋白酶活性 Proteases activity |
过氧化氢酶活性 Catalase activity |
V1 | V2 | |
| 上层 | 脲酶活性 | -0.535 | |||||
| Upper layer | 蛋白酶活性 | 0.029 | 0.258 | ||||
| 过氧化氢酶活性 | -0.123 | 0.273 | -0.033 | ||||
| V1 | -0.330 | -0.159 | -0.288 | 0.014 | |||
| V2 | -0.114 | -0.201 | 0.256 | -0.346 | 0.192 | ||
| V3 | -0.322 | -0.212 | -0.089 | -0.123 | 0.928** | 0.546 | |
| 脲酶活性 | -0.097 | ||||||
| 中层 | 蛋白酶活性 | 0.437 | -0.228 | ||||
| Middle layer | 过氧化氢酶活性 | -0.228 | 0.154 | -0.238 | |||
| V1 | 0.315 | -0.349 | 0.232 | -0.134 | |||
| V2 | 0.007 | 0.298 | -0.416 | -0.125 | 0.277 | ||
| V3 | 0.341 | -0.240 | 0.162 | -0.148 | 0.977** | 0.458 | |
| 脲酶活性 | -0.265 | ||||||
| 蛋白酶活性 | 0.484 | -0.441 | |||||
| 下层 | 过氧化氢酶活性 | -0.034 | 0.478 | -0.200 | |||
| Sub layer | V1 | 0.436 | -0.237 | 0.634* | -0.069 | ||
| V2 | 0.048 | 0.171 | -0.286 | 0.314 | -0.350 | ||
| V3 | 0.021 | 0.048 | 0.591* | 0.024 | 0.956** | -0.061 | |
| 上层代表 0—15 cm土层;中层代表 15—30 cm土层;下层代表 30—45 cm土层; V1:土壤净氮硝化速率;V2:土壤净氮氨化速率;V3:土壤净氮矿化速率; *, P < 0.05, **, P < 0.01 | |||||||
本研究结果显示, 在CK处理下, 上层土壤TOC含量为(30.51±2.26) g/kg, 与四川相似林分研究中土壤TOC含量大致相同[16—18], 高于北京延庆区松山自然保护区森林土壤TOC含量(27.42 g/kg)[19]、赣南生态林土壤TOC含量(16.47g/kg)[20]。本研究发现, 土壤TOC随土层深度增加显著降低, HN处理显著增加了各层土壤TOC含量, HN处理下, 三层土壤TOC含量均值为28.22 g/kg, LN处理下为18.18 g/kg, CK处理下则为20.75 g/kg。土壤TOC含量主要取决于土壤TOC输入与输出间的平衡[21], 外源氮添加抑制了土壤中木质素的分解、土壤呼吸等利于土壤碳排放的路径, 导致HN处理下土壤TOC含量显著增加[22]。因此, 施氮增加TOC的原因可能是:(1)施氮促进了Mn流失, 导致Mn生物可利用性降低, 从而抑制了锰过氧化物酶活性, 最终抑制木质素的降解;凋落物分解受到抑制[23]。(2)施氮可以稳定异养呼吸, 增强其对微生物分解的抵抗力, 将部分无机氮纳入异养呼吸, 形成一些顽固化合物, 同时对土壤微生物生物量产生负面影响, 最终导致土壤呼吸受到抑制[24]。
土壤TN含量在土壤层中是处于动态变化之中的, 土壤TN含量的增减与土壤中氮元素的积累与消耗密切相关, 土壤有机质的积累与分解作用是影响TN含量的主要因素[25—26]。本研究样地所在的次生常绿阔叶林, 其上层土壤TN含量均值在自然状态下为(5.13±0.09) g/kg, 与瓦屋山次生常绿阔叶林研究中0—10cm腐殖质层土壤TN含量测量值(5.93 g/kg)近似[24]。本研究中, 同一外源氮添加条件下, 土壤TN含量随土壤深度的加深而减少。可能机制是:(1)本研究样地属于亚热带湿性次生常绿阔叶林, 林地内多年生木本植物众多, 表层土壤森林凋落物输入量大, 而凋落物分解产生的有机质难以混入下层土壤[20];(2)随着土壤深度的加深, 参与土壤有机质生化反应过程的土壤微生物与土壤动物数量往往会急剧下降[27—28]。本研究还发现。模拟氮沉降处理下上层、中层TN有增加趋势, 但差异未达统计显著水平, 与该样地氮沉降处理初期[29]试验结果相似, 究其原因可能是模拟氮沉降并未改变该样地的凋落物输入量。因此, 无机氮的增加并未引起土壤有机氮的大量增加, 而多余的无机氮又较容易随水淋溶或通过反硝化过程而损失。
本研究中, 在同一外源氮添加条件下, 室内恒温培养前、后土壤硝态氮、铵态氮含量均随土壤深度增加而减少。研究发现, 土壤深度对培养后土壤硝态氮和培养前土壤铵态氮的含量影响较为显著, 造成下层土壤有效氮含量较低的原因可能是随着土壤深度加深, 下层土壤微生物和土壤动物含量减少[30—31], 对土壤氮元素的转化速率减慢。模拟氮沉降对培养后土壤硝态氮和培养前土壤铵态氮的含量均有显著影响。培养前土壤铵态氮的含量随外源氮添加含量的增加而增加, 与孙宇等[32]在碧峰峡次生常绿阔叶林的研究结果一致, 外源氮添加以直接或间接的方式使土壤无机氮含量增加。培养后土壤硝态氮含量则表现为低氮条件下促进, 高氮条件下抑制的现象, 推测其原因可能是, 本试验样地地处四川盆地西部边缘, 受东南季风和地势的影响, 常年多雨, 长期高氮的施加造成大量土壤硝态氮淋溶流失, 加速了土壤硝态氮的气态损失过程, 最终导致高氮条件抑制土壤硝态氮含量。
3.2 模拟氮沉降对不同深度土壤氮矿化的影响本研究中矿化作用产生的土壤铵态氮的含量略高于土壤硝态氮含量, 该次生常绿阔叶林无机氮的更多以铵态氮的形式存在。土壤净氮氨化速率对模拟氮沉降的响应呈现出与铵态氮变化相似的特点。在同一外源氮添加条件下, 下层土壤净氮氨化速率明显低于上层土壤氨化速率, 但下层土壤净氮氨化速率高于中层土壤, 主要原因可能是, 本试验样地有长达7年的持续施氮过程, 且样地地处四川盆地西部的华西雨屏区, 常年雨水充沛, 导致施加的氮元素随着雨水淋溶到更深层的土壤, 导致下层土壤净氮氨化速率高于中层土壤, 甚至接近表层土壤。
土壤净氮硝化速率对模拟氮沉降的响应呈现出与硝态氮变化相似的特点。土壤净氮硝化速率在三种氮处理下的表现情况各有不同, CK处理下, 中层土壤的净氮硝化速率远高于上层土壤与下层土壤, LN处理下则表现出与土壤净氮氨化速率相同的规律, HN处理下, 土壤净氮硝化速率随土壤深度加深而降低。三个土层的不同响应趋势, 其原因可能试验中是氮添加处理中反硝化作用增强导致的, 这一点在课题组近期试验中被观察到。本试验中, 土壤净氮矿化速率随着土壤深度的加深显著降低, 说明该样地中可供矿化反应的土壤有效氮仍然大量存在于土壤表层[33—34], 而深层土壤氮矿化速率仍然受到土壤微生物和土壤动物的数量、凋落物输入量、土壤氮矿化相关酶活性等多种因素的限制[35—37]
本研究中, 模拟氮沉降显著抑制了土壤净氮矿化速率, 这与大多数前人研究所得出的外源氮添加会促进土壤氮矿化速率结果不同[32]。森林土壤氮除进行矿化作用外, 还进行腐殖化过程, 后者又被称作矿化氮的固持作用[38], 外源氮添加对土壤氮有激发效应, 正效应为矿化, 负效应为固定[39], 推测造成本研究中土壤净氮矿化速率受抑制原因, 可能是外源氮添加条件下两个过程间相互制约的结果。肖银龙等人于2013年在华西雨屏区苦竹林模拟氮沉降试验表明, 外源添加氮显著促进了苦竹林土壤净氮硝化速率, 与该试验仅进行了2年施氮处理不同的是, 本试验样地已经进行了长达7年的施氮处理, 据此推测, 长期性外源氮添加使得森林生态系统处于氮饱和状态, 氮饱和状态下会抑制森林土壤氮矿化过程。徐星凯等人在长白山阔叶红松林进行了时长3年的模拟氮沉降试验, 结果表明, 随着施氮时间的延长, 模拟氮沉降对土壤净氮矿化速率的促进作用明显减弱。程奕等人通过模拟施氮试验, 研究森林生态系统土壤全氮转化对氮沉降的响应, 在较宽的背景氮沉积速率范围内进行了模拟加氮实验, 结果表明, 氮沉降能深刻地改变土壤氮素转化过程和森林生态系统的长期生产力[40]。该试验首次发现, 随着背景氮沉积速率的增加, 氮矿化、硝化和硝态氮固定化的总速率对实验氮添加量的响应由正变为负, 其阈值分别为3.23、6.02、1.90 kg N hm-2 a-1, 高于此值, 施氮量对三种总氮转化速率均有负效应, 矿化速率、硝化速率和硝态氮固定化速率可表观背景氮沉降[40]。在未来氮沉降情景下, 应将背景氮沉降速率纳入生态系统模型, 以更好地预测森林生态系统氮循环[40]。
关于森林土壤氮矿化率的研究表明, 模拟氮沉降可以增加、减少或不影响氮矿化率[40], 当森林接近氮饱和时, 土壤氮矿化可被氮沉积抑制[7, 41—42]。可能的解释是腐殖质降解酶活性的抑制[43]和土壤有机质中化学键结构的改变是由于长期的高氮沉积速率, 这些反过来又降低了细胞外分解代谢酶的有效性[7]。因此, 高氮富集条件通过改变微生物种群规模、微生物群落结构、酶活性和生存策略抑制微生物分解, 并最终减少氮矿化[7, 42]。
3.3 模拟氮沉降对不同深度土壤酶活性的影响本研究中, 模拟氮沉降对土壤脲酶活性的促进作用极显著, 三层土壤中, 脲酶活性均随氮添加的增多而增强, 对土壤氮矿化作用有极大影响, 在前人研究中, 大多数结果表明土壤氮矿化作用与土壤脲酶活性呈极显著正相关关系[44—45], 但本实验中, 土壤脲酶的活性虽增强, 但土壤氮矿化速率受到抑制, 其原因可能是, 影响土壤氮矿化作用的因素不止脲酶活性, 硝酸还原酶活性的降低等因素对其抑制作用强于脲酶的促进作用[46], 最终呈现出模拟氮沉降抑制土壤氮矿化速率, 因此, 对影响土壤氮矿化速率因素的研究应该扩大范围, 更加深入。
本研究中, 模拟氮沉降显著抑制了土壤硝酸还原酶活性, 与前人研究结果相同[10, 47], 三层土壤均呈现出相同规律。硝酸还原酶是影响酸性土壤发生反硝化作用的主要酶, 能够促使土壤中的硝酸根离子转变为亚硝酸根离子。亚硝酸根离子是土壤硝化作用的主要原料, 土壤硝酸还原酶受到外源氮添加的抑制, 导致参与土壤硝化作用亚硝酸根离子减少, 土壤硝化速率降低, 与上述结果一致。因此, 模拟氮沉降使得硝酸还原酶活性受到抑制, 可能是导致外源氮添加抑制土壤氮矿化速率的原因之一。
土壤深度对酶活性的影响, 均呈现出随土壤深度加深, 酶活性减弱现象。造成此现象的原因可能是深层土壤动物和微生物种类及数量都明显少于表层土壤[48—49], 且深层土壤透气性差, 而大多数土壤酶均需要在有氧条件发挥作用[51]。值得一提的是, 大多数前人研究结果表示模拟氮沉降会促进土壤蛋白酶的活性[51], 但本试验中, 模拟氮沉降反而抑制了土壤蛋白酶活性, 且中、下两层土壤的抑制作用强于上层土壤。土壤蛋白酶是参与土壤氮循环的关键酶, 其将有机氮矿化为无机氮, 进而氨化为铵根离子, 是土壤矿化过程的限速酶[52]。因此, 在本研究中, 净氮硝化速率、净氮矿化速率均与蛋白酶活性呈现显著正相关关系, 所以土壤蛋白酶活性的降低也许是造成土壤氮矿化速受到抑制的原因之一。
4 结论本研究结果表明:(1)该常绿阔叶次生林不同深度土壤TN有显著差异;(2)模拟氮沉降对该系统土壤氮矿化总体表现出极显著抑制作用, 其中中层土壤抑制作用最为强烈, 净氮矿化速率主要受硝化过程的影响;(3)氮矿化相关酶活性均随土壤深度的加深而降低, 模拟氮沉降对土壤脲酶活性有极显著促进作用, 对土壤硝酸还原酶活性有显著抑制作用。
| [1] |
Holland E A, Dentener F J, Braswell B H, Sulzman J M. Contemporary and pre-industrial global reactive nitrogen budgets. Biogeochemistry, 1999, 46(1/3): 7-43. DOI:10.1023/A:1006148011944 |
| [2] |
鲁显楷, 莫江明, 张炜, 毛庆功, 刘荣臻, 王聪, 王森浩, 郑棉海, Mori T, 毛晋花, 张勇群, 王玉芳, 黄娟. 模拟大气氮沉降对中国森林生态系统影响的研究进展. 热带亚热带植物学报, 2019, 27(5): 500-522. |
| [3] |
陈伏生, 曾德慧, 范志平, 陈广生, Singh A N. 沙地不同树种人工林土壤氮素矿化过程及其有效性. 生态学报, 2006, 26(2): 341-348. DOI:10.3321/j.issn:1000-0933.2006.02.005 |
| [4] |
熊毅, 李庆揆. 土壤氮素. 北京: 科学出版社, 1988: 464-482.
|
| [5] |
Magill A H, Downs M R, Nadelhoffer K J, Hallett R A, Aber J D. Forest ecosystem response to four years of chronic nitrate and sulfate additions at Bear Brooks Watershed, Maine, USA. Forest Ecology and Management, 1996, 84(1/3): 29-37. |
| [6] |
Thimonier A, Pannatier E G, Schmitt M, Waldner P, Walthert L, Schleppi P, Dobbertin M, Kräuchi N. Does exceeding the critical loads for nitrogen alter nitrate leaching, the nutrient status of trees and their crown condition at Swiss Long-term Forest Ecosystem Research (LWF) sites?. European Journal of Forest Research, 2010, 129(3): 443-461. DOI:10.1007/s10342-009-0328-9 |
| [7] |
Aber J, McDowell W, Nadelhoffer K, Magill A, Berntson G, Kamakea M, McNulty S, Currie W, Rustad L, Fernandez I. Nitrogen saturation in temperate forest ecosystems: hypotheses revisited. BioScience, 1998, 48(11): 921-934. DOI:10.2307/1313296 |
| [8] |
王建林, 钟志明, 王忠红, 陈宝雄, 余成群, 胡兴祥, 沈振西, 大次卓嘎, 张宪洲. 青藏高原高寒草原生态系统土壤碳氮比的分布特征. 生态学报, 2014, 34(22): 6678-6691. |
| [9] |
Saiya-Cork K R, Sinsabaugh R L, Zak D R. The effects of long term nitrogen deposition on extracellular enzyme activity in an Acer saccharum forest soil. Soil Biology and Biochemistry, 2002, 34(9): 1309-1315. DOI:10.1016/S0038-0717(02)00074-3 |
| [10] |
Olander L P, Vitousek P M. Regulation of soil phosphatase and chitinase activityby N and P availability. Biogeochemistry, 2000, 49(2): 175-191. DOI:10.1023/A:1006316117817 |
| [11] |
马豪宇. 模拟氮沉降对湿性常绿阔叶林土壤溶液化学特性的影响[D]. 四川农业大学, 2019. DOI: 10.27345/d.cnki.gsnyu.2019.000732.
|
| [12] |
何文寿, 王菊兰, 李钰. 宁夏不同地区农田土壤矿质氮的空间变异特征. 干旱地区农业研究, 2005, 23(6): 47-55. DOI:10.3321/j.issn:1000-7601.2005.06.010 |
| [13] |
鲍士旦. 土壤农化分析(第三版). 北京: 中国农业出版社, 2000.
|
| [14] |
关松荫. 土壤酶及其研究法. 北京: 农业出版社, 1986.
|
| [15] |
武志杰, 隽英华, 陈利军, 史云峰. 一种检测土壤硝酸还原酶活性的分析方法: 中国, CN101271060B. 2011-06-08.
|
| [16] |
黄从德, 张国庆, 唐宵, 王勇军, 王宪帅. 四川省马尾松人工林土壤有机碳密度研究. 水土保持研究, 2009, 16(2): 46-49, 55-55. |
| [17] |
黄从德, 张健, 杨万勤, 唐宵, 赵安玖. 四川省及重庆地区森林植被碳储量动态. 生态学报, 2008, 28(3): 966-975. DOI:10.3321/j.issn:1000-0933.2008.03.008 |
| [18] |
彭勇, 陈刚, 陈冠陶, 梁政, 涂利华. 模拟氮沉降对瓦屋山常绿阔叶次生林土壤呼吸的影响. 应用与环境生物学报, 2015, 21(4): 733-739. |
| [19] |
聂浩亮, 薄慧娟, 张润哲, 王江, 聂立水, 王丹. 北京海坨山典型林分土壤有机碳含量及有机碳密度垂直分布特征. 林业科学研究, 2020, 33(6): 155-162. |
| [20] |
孙浩, 赖祁华, 刘晓勇, 江柳春, 徐昕, 郑永红. 赣南地区生态公益林5种森林类型土壤有机碳特征. 南方林业科学, 2020, 48(6): 45-48, 63-63. |
| [21] |
彭天驰. 模拟氮沉降对次生湿性常绿阔叶林土壤生化特性的影响[D]. 成都: 四川农业大学, 2018.
|
| [22] |
马秀枝, 王艳芬, 汪诗平, 王金枝, 李长生. 放牧对内蒙古锡林河流域草原土壤碳组分的影响. 植物生态学报, 2005, 29(4): 569-576. |
| [23] |
李曾燕, 邱细容, 陈冠陶, 郑军, 李娟, 涂利华. 多年模拟氮沉降对华西雨屏区苦竹人工林土壤节肢动物的影响. 生态学杂志, 2019, 38(5): 1419-1425. |
| [24] |
Peng Y, Song S Y, Li Z Y, Li S, Chen G T, Hu H L, Xie J L, Chen G, Xiao Y L, Liu L, Tang Y, Tu L H. Influences of nitrogen addition and aboveground litter-input manipulations on soil respiration and biochemical properties in a subtropical forest. Soil Biology and Biochemistry, 2020, 142: 107694. |
| [25] |
彭勇. 模拟氮沉降对瓦屋山次生常绿阔叶林地下碳循环过程和土壤生化特性的影响[D]. 成都: 四川农业大学, 2016.
|
| [26] |
葛晓敏, 唐罗忠, 王瑞华, 李勇, 朱玲, 贾志远, 丁晖. 杨树人工林生态系统凋落物生物量及其分解特征. 生态环境学报, 2017, 26(9): 1457-1464. |
| [27] |
Naseby D C, Pascual J A, Lynch J M. Effect of biocontrol strains of Trichoderma on plant growth, Pythium ultimum populations, soil microbial communities and soil enzyme activities. Journal of Applied Microbiology, 2000, 88(1): 161-169. |
| [28] |
Holt J A. Grazing pressure and soil carbon, microbial biomass and enzyme activities in semi-arid northeastern Australia. Applied Soil Ecology, 1997, 5(2): 143-149. |
| [29] |
李顺. 施氮和凋落物增减对湿性常绿阔叶林土壤呼吸和土壤碳、氮的影响[D]. 成都: 四川农业大学, 2018.
|
| [30] |
王海英, 宫渊波, 陈林武. 嘉陵江上游不同植被恢复模式土壤微生物及土壤酶活性的研究. 水土保持学报, 2008, 22(3): 172-177. |
| [31] |
夏孟婧, 苗颖, 陆兆华, 谢国莉, 裴定宇. 造纸废水灌溉对滨海退化盐碱湿地土壤酶活性的响应. 生态学报, 2012, 32(21): 6599-6608. |
| [32] |
孙宇, 彭天驰, 李顺, 周培, 王婷, 钱学诗, 刘枭鹏, 陈雨芩, 马豪宇, 李曾燕, 涂利华. 模拟氮沉降对湿性常绿阔叶次生林土壤碳氮组分和酶活性的影响. 水土保持学报, 2019, 33(2): 235-243, 250-250. |
| [33] |
Ohrui K, Mitchell M J, Bischoff J M. Effect of landscape position on N mineralization and nitrification in a forested watershed in the Adirondack Mountains of New York. Canadian Journal of Forest Research, 1999, 29(4): 497-508. |
| [34] |
Trofymow J A, Morley C R, Coleman D C, Anderson R V. Mineralization of cellulose in the presence of chitin and assemblages of microflora and fauna in soil. Oecologia, 1983, 60(1): 103-110. |
| [35] |
谢正苗, 卡里德, 黄昌勇, 俞劲炎. 镉铅锌污染对红壤中微生物生物量碳氮磷的影响. 植物营养与肥料学报, 2000, 6(1): 69-74. |
| [36] |
王斌, 陈亚明, 周志宇. 贺兰山西坡不同海拔梯度上土壤氮素矿化作用的研究. 中国沙漠, 2007, 27(3): 483-490. |
| [37] |
林贵刚, 赵琼, 赵蕾, 李慧超, 曾德慧. 林下植被去除与氮添加对樟子松人工林土壤化学和生物学性质的影响. 应用生态学报, 2012, 23(5): 1188-1194. |
| [38] |
鲁彩艳, 陈欣. 土壤氮矿化-固持周转(MIT)研究进展. 土壤通报, 2003, 34(5): 473-477. |
| [39] |
黄东迈, 朱培立, 王志明. 耕作土壤有机态氮内循环中几个问题的商榷//《氮素循环与农业和环境》专辑——氮素循环与农业和环境学术讨论会论文集. 厦门: 科学出版社, 2001: 104-112.
|
| [40] |
Cheng Y, Wang J, Ge Z W, Zhang J B, Cai Y J, Chang S X, Cai Z C, Chen H Y H. Background nitrogen deposition controls the effects of experimental nitrogen addition on soil gross N transformations in forest ecosystems. Biogeochemistry, 2020, 151(2/3): 335-341. |
| [41] |
Corre M D, Beese F O, Brumme R. Soil nitrogen cycle in high nitrogen deposition forest: changes under nitrogen saturation and liming. Ecological Applications, 2003, 13(2): 287-298. |
| [42] |
Corre M D, Brumme R, Veldkamp E, Beese F O. Changes in nitrogen cycling and retention processes in soils under spruce forests along a nitrogen enrichment gradient in Germany. Global Change Biology, 2007, 13(7): 1509-1527. |
| [43] |
Carreiro M M, Sinsabaugh R L, Repert D A, Parkhurst D F. Microbial enzyme shifts explain litter decay responses to simulated nitrogen deposition. Ecology, 2000, 81(9): 2359-2365. |
| [44] |
于群英. 土壤磷酸酶活性及其影响因素研究. 安徽技术师范学院学报, 2001, 15(4): 5-8. |
| [45] |
刘瑞丰, 李新平, 李素俭, 张小虎, 何军. 商洛地区土壤蔗糖酶及过氧化氢酶与土壤养分的关系研究. 干旱地区农业研究, 2011, 29(5): 182-185. |
| [46] |
安韶山, 黄懿梅, 郑粉莉. 黄土丘陵区草地土壤脲酶活性特征及其与土壤性质的关系. 草地学报, 2005, 13(3): 233-237. |
| [47] |
Allison S D, Vitousek P M. Responses of extracellular enzymes to simple and complex nutrient inputs. Soil Biology and Biochemistry, 2005, 37(5): 937-944. |
| [48] |
许华, 阮维斌, 高玉葆, 宋晓艳, 魏宇昆. 根结线虫对黄瓜叶片氮磷含量、土壤pH和酶活性的影响. 应用生态学报, 2010, 21(8): 2038-2044. |
| [49] |
井永苹, 赵莉, 陈小云, 刘满强, 胡锋, 李辉信. 土壤食细菌线虫对菲降解的影响. 土壤, 2010, 42(3): 372-377. |
| [50] |
王启兰, 王溪, 王长庭, 曹广民, 龙瑞军. 高寒矮嵩草草甸土壤酶活性与土壤性质关系的研究. 中国草地学报, 2010, 32(3): 51-56. |
| [51] |
戴全厚, 薛萐, 刘国彬, 勃海峰, 张健. 黄土丘陵区封禁对侵蚀土壤微生物生物量的影响. 土壤学报, 2008, 45(3): 518-525. |
| [52] |
Kamimura Y, Hayano K. Properties of protease extracted from tea-field soil. Biology and Fertility of Soils, 2000, 30(4): 351-355. |