 2022, Vol. 42
2022, Vol. 42文章信息
- 邓杨, 王瑶, 郭洋楠, 李欣颖, 陆兆华, 张萌, 陈睿, 吴皓宇, 刘育达
- DENG Yang, WANG Yao, GUO Yangnan, LI Xinying, LU Zhaohua, ZHANG Meng, CHEN Rui, WU Haoyu, LIU Yuda
- 干旱胁迫下柳枝稷在露天矿区土壤中的种子萌发和生长特性
- Effect of drought stress on the seed germination and growth characteristics of switchgrass
- 生态学报. 2022, 42(22): 9175-9185
- Acta Ecologica Sinica. 2022, 42(22): 9175-9185
- http://dx.doi.org/10.5846/stxb202107091854
-
文章历史
- 收稿日期: 2021-07-09
- 网络出版日期: 2022-07-12





2. 神华神东煤炭集团有限责任公司技术研究院, 榆林 719315;
3. 河南省矿山生态修复工程技术中心, 郑州 450000
2. Institude of Technology, ShenhuaShendong Coal Group Company Limited, Yulin 719315, China;
3. Henan Mine Ecological Restoration Engineering Technology Research Center, Zhenzhou 450000, China
煤炭在我国一次性能源消费结构中占主导地位, 在国民经济发展和人民生活中起着极其重要的作用[1]。我国煤矿主要分布在气候干燥、降雨量少、水资源短缺的干旱半干旱区域[2]。煤炭的开采促进经济发展的同时也造成了严重的生态破坏。由煤炭开采造成的物种资源破坏、水土流失、土地荒漠化现象, 严重影响矿区及周边农牧民的生活生产和地方经济的可持续发展[3]。因此, 选择抗逆性强、生产性能好的牧草品种是保障矿区植被恢复重建、建设生态文明的关键[4]。而能源植物的种植, 既可缓解当地生物质原材料的短缺, 又能改善生态环境, 是兼顾生态与经济效益的有效植被恢复方式[5]。
在干旱区, 植物都必须具备复杂的生存机制和避逆策略, 才能在特定环境中生存[6]。柳枝稷(Panicum virgatum L.)作为多年生C 4草本植物, 不仅具有适应性强, 生长速度快, 产量高, 生产成本低, 水分利用效率高的优点, 并且根系发达, 有很好的水土保持效果, 还有助于提高土壤有机质含量, 具有良好的生态效益[7—8]。目前, 柳枝稷已经在我国的挖沙废弃地、重金属污染土地、盐碱地等受污染土壤上进行引种, 均表现出较好的生态适应性和较高的生产潜力[9—10]。若能将其引种到矿区土壤中, 不仅有利于矿区生态修复, 还能提供优质的生物质能, 但目前柳枝稷矿区环境的生态适应性研究仍较少。
植物的种子萌发及出苗阶段对逆境的适应能力是决定该植被能否在矿区成功建植及延续的关键[11]。植被生态恢复与种子萌发和种苗存活有直接关系[12]。目前, 研究柳枝稷种子萌发特性的成果很多。何学青[13]等通过PEG-6000模拟水分胁迫对柳枝稷种子萌发的影响发现, 柳枝稷种子发芽率、发芽指数及活力指数随着水分胁迫程度的增加而下降。抗旱性植被在种子萌发和种苗生长阶段都会采取复杂的避逆策略应对恶劣环境[14]。珊丹[15]等研究发现柳枝稷种苗能通过增加游离脯氨酸及提高可溶性糖含量来缓解土壤干旱胁迫的伤害。范希峰[16]等研究表明柳枝稷种苗能在华北地区沙化贫瘠的边际土壤正常生长发育。对于干旱且贫瘠的矿区土壤基质下柳枝稷的萌发特性以及种苗生长状态尚不清楚。
对柳枝稷生长特性的研究也有很多报道。郑敏娜[17]对4种禾草植被的抗旱性研究表明, 柳枝稷的抗旱性明显高于另外三种。赵春桥[18]等研究表明, 干旱胁迫显著抑制植被的生长, 而根冠比显著增加, 表现出了柳枝稷的生存策略。徐炳成等[19]和左海涛等[20]分别以北京潮褐土、兰州灰钙土和陕北安塞地区黄绵土为基质的研究结果表明, 水分胁迫显著抑制柳枝稷生长, 使其干物质积累下降、株高和分蘖数减少、净光合速率下降、蒸腾速率降低。而在矿区土壤基质下, 柳枝稷生长发育对干旱胁迫的响应机制研究较少。
针对上述问题, 本文通过研究柳枝稷在矿区土壤中的种子萌发和种苗生长发育对干旱胁迫响应, 以室内模拟试验探究柳枝稷对矿区土壤环境的生态适应性, 为干旱矿区柳枝稷的大规模引种和栽培提供科学依据, 对矿区生态修复具有重要意义。
1 材料与方法 1.1 试验材料试验材料为柳枝稷种子和幼苗, 品种均为Cave-in-Rock。以锡林浩特市北电胜利矿区内排土场土壤为矿区土壤基质(以下简称矿区土), 以黄河三角洲生态环境研究中心温室棕壤土为非矿区土壤基质(以下简称非矿区土), 理化性质见表 1, 其中天然含水率为当年7月份测得。栽培容器为高23.5 cm, 上直径25.5 cm, 下直径17 cm的塑料花盆, 花盆底部平均分布有3个小孔(孔径为1.5 cm), 以保证透气性。为了防止土壤沿小孔漏出, 在盆内底部铺两层纱布, 底部并配以托盘。
| 土壤类型 Soil type |
田间最大持水量 Maximum field water holding capacity/% |
天然含水率 Natural moisture content/% |
pH | 电导率 Electrical conductivity/ (μs/cm) |
含盐量 Salt content/% |
有机质 Organic matter/ (g/kg) |
全碳 Total carbon/ (g/kg) |
全氮 Total nitrogen/ (g/kg) |
全磷 Total phosphorus/ (g/kg) |
| 矿区土Mining soil | 27 | 5.7 | 8.84 | 394 | 0.17 | 6.74 | 8.95 | 0.21 | 0.29 |
| 非矿区土Non-mining soil | 30 | 11.4 | 7.76 | 258 | 0.09 | 12.96 | 17.48 | 0.74 | 0. 68 |
将矿区土壤混匀置于培养皿中铺平, 用烘干法设置4个不同的水分梯度(即对照组、轻度胁迫、中度胁迫、重度胁迫分别为田间最大持水量的80%、60%、40%、20%), 每个梯度设置5组平行试验。选取大小均匀、籽粒饱满、无机械损伤、成熟健康的种子, 冲洗干净后用70%酒精消毒3 min, 蒸馏水冲洗3次, 用吸水纸吸干表面水分用于试验。将种子埋入土下2 mm, 每个培养皿30粒种子, 置于恒温30℃、相对湿度50%、12 h光暗交替的培养箱中。为防止水分蒸发, 用塑料薄膜封口, 每天透气1次。每天将种子连同培养皿用电子天平称重, 采用人工称重法补充消耗的水分, 保持培养皿中土壤含水量不变, 期间不施加任何肥料。种子萌发以胚芽露出土表面视为发芽。每天同一时间记录种子发芽数, 当连续4 d不再有种子发芽时视为发芽试验结束。
1.2.2 盆栽试验盆栽试验在黄河三角洲生态环境研究中心温室内采用盆栽土培法进行。于2018年10月将直径、长度大致相同的柳枝稷移栽于花盆中, 每盆5株, 每株3个分蘖, 经过6个月缓苗期后用于试验。2019年5月5日开始干旱胁迫处理。用环刀法测定土壤田间最大持水量, 并以此设置4种水分梯度:对照组(田间最大持水量的80%)、轻度胁迫(田间最大持水量的60%)、中度胁迫(田间最大持水量的40%)、重度胁迫(田间最大持水量的20%)。采用称重法控制水分, 每天下午5:00点称重, 并补水达到设定的土壤含水量。由于植物生长, 每隔7 d在补水后1—2 h内用烘干法测定一次土壤含水量, 对数据进行校正。除水分处理外, 其他栽培管理措施完全一致。每个处理设置5个平行, 共40盆。试验期间每天测定温室内温度、湿度。干旱胁迫60 d后进行生长指标的测定。
1.3 指标测定与方法 1.3.1 种子萌发指标测定干旱胁迫下种子萌发测定发芽势、发芽率、发芽指数以及种苗的芽长、根长、鲜重, 根据每天记录的发芽数计算种子活力指数。

|
(1) |

|
(2) |
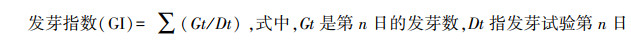
|
(3) |
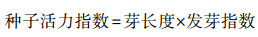
|
(4) |
形态参数:植株的生长指标(形态变化、叶片颜色变化和叶片数量变化)于盆栽试验开始后, 定期观察并拍照记录。枯叶率为叶片面积的50%出现干枯萎蔫的叶片数[21]。分蘖数为地面以下或近地面处所发生的分枝数量[22]。每个处理随机选取5株柳枝稷挂牌, 每30 d对挂牌植株用卷尺测量记录植株露出土壤表面到顶端的垂直高度, 并计算其相对生长高度。试验结束后, 将柳枝稷整株取出, 将植株地上与地下部分分离, 用卷尺测量根基部至最长须根尖端的自然长度作为根长。采用量筒排水法测定根系体积。并记录每株柳枝稷的根数。

|
(5) |
生物量计算:盆栽试验结束后, 将柳枝稷整株取出, 将其分为地上和地下两部分, 洗净后于105℃下杀青30 min, 80℃下烘干至恒重, 称重, 分别得到地上生物和地下生物量, 并计算总生物量和根冠比。

|
(6) |

|
(7) |
叶片水分计算:每个处理取相同部位的叶片测定。用分析天平称得叶片鲜重为W1, 在黑暗中浸入蒸馏水中24 h后, 用干净的纱布吸干叶片表面多余水分, 称得饱和鲜重W2。然后将叶片放入烘箱105 ℃杀青、80 ℃烘干至恒重W3。根据以下公式计算叶片相对含水量和叶片水分饱和亏缺。

|
(8) |
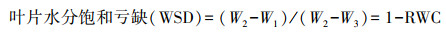
|
(9) |
采用SPSS 26.0和Origin 2021进行数据分析, 试验数据为平均值±标准误差。对同一土壤基质在不同干旱胁迫程度下植物株高、枯叶率、根冠比、根长、根体积及根数进行单因素方差分析, 显著性水平为P=0.05, 并用Duncan新复极差法进行多重比较。对不同土壤基质在同一干旱胁迫程度下植被枯叶率、根冠比、根长、根体积及根数进行独立样本的T检验分析。
2 结果与分析 2.1 干旱胁迫对柳枝稷种子萌发的影响由表 2可知, 随着干旱胁迫程度的增加, 柳枝稷种子的各萌发指标均呈现先增后减的变化趋势, 且柳枝稷在轻度胁迫下种子各萌发指标达到最大值, 在重度胁迫下柳枝稷种子各萌发指标明显低于其余三组, 表明其受到明显抑制。种苗的生长发育在重度胁迫下表现最好, 种苗根长和鲜重在重度胁迫下达到最大值, 比对照分别高了63.7%、30.2%。轻度胁迫下, 柳枝稷各种苗生长指标最低, 表明轻度胁迫对柳枝稷种苗的生长有一定的抑制作用, 在重度胁迫下具有一定的适应能力。
| 萌发指标 Germination indicators |
对照 Control treatment |
轻度胁迫 Mild stress |
中度胁迫 Moderate stress |
重度胁迫 Severe stress |
| 发芽总数Total number of germinations/个 | 40 | 45 | 31 | 5 |
| 发芽率Germination rate/% | 26.67 | 30.00 | 20.67 | 3.33 |
| 发芽势Germinative force/% | 3.33 | 6.67 | 6.67 | 0 |
| 发芽指数Germination Index | 3.16 | 4.73 | 3.17 | 1.11 |
| 活力指数Vitality index | 72.67 | 61.21 | 65.75 | 24.19 |
| 根长Root length/cm | 18.96±1.35 | 11.28±1.20 | 15.71±2.12 | 31.03±3.20 |
| 芽长Bud length/cm | 23.00 ±2.45 | 12.94±0.84 | 20.74±2.07 | 21.80±3.45 |
| 芽重Bud weight/mg | 3.51±0.18 | 1.80±0.07 | 3.80±0.10 | 3.90±0.11 |
| 种苗鲜重Seedling fresh weight/mg | 6.92±0.19 | 0.30±0.11 | 6.50±0.16 | 9.01±0.21 |
如图 1所示, 随着干旱胁迫时间的增加, 柳枝稷在不同土壤基质下的形态变化相似。非矿区土壤比矿区土壤营养物质更丰富, 因此柳枝稷在非矿区土壤基质下每株叶片数更多, 叶片颜色更绿, 植株更高, 总体生长更加旺盛。在干旱胁迫初期, 柳枝稷在重度胁迫下出现缺水症状, 开始萎蔫, 叶片开始卷曲。干旱胁迫30 d后, 对照、轻度胁迫和中度胁迫的植株长势差别不大, 重度胁迫植株生长缓慢, 叶尖变黄、边缘起皱, 并出现叶片下垂、叶片脱落现象, 植株整体干枯变黄。

|
| 图 1 干旱胁迫30 d后柳枝稷生长情况 Fig. 1 The growth of switchgrass after 30 days of drought stress |
如图 2所示, 干旱胁迫60 d后, 对照组的柳枝稷长势最好, 轻度胁迫和中度胁迫下长势次之, 重度胁迫下的植株半数叶片发黄干枯, 底部叶片发黄状况更为严重, 部分叶片脱落, 萎蔫现象严重, 但未出现死亡植株。由此可以看出, 贫瘠的矿区土壤环境会使柳枝稷的生长发育受到限制, 而干旱胁迫会进一步抑制柳枝稷的生长, 但在长时间内重度干旱下不会使其死亡, 体现了柳枝稷的抗旱性和抗贫瘠性。

|
| 图 2 干旱胁迫60 d后柳枝稷生长情况 Fig. 2 The growth of switchgrass after 60 days of drought stress |
由表 3可知, 随着干旱胁迫的加剧, 两种土壤基质下柳枝稷的相对生长高度呈现先升后降的趋势, 在轻度胁迫下最大, 并且株高在胁迫30 d及60 d后在轻度胁迫下达到最大值。两种土壤基质中的柳枝稷在重度胁迫下柳枝稷同时出现负增长, 表明对柳枝稷的生长产生了抑制作用。非矿区土中的柳枝稷普遍高于矿区土中的柳枝稷。矿区土壤基质和非矿区土壤基质下柳枝稷相对生长高度在对照组相差最大。结果表明, 干旱胁迫对柳枝稷的株高有一定影响。适当的土壤含水量会提高柳枝稷的相对生长量, 极度缺水时, 则会抑制柳枝稷的生长。
| 土壤类型 Soil type |
干旱胁迫程度 Drought stress |
胁迫前株高 Plant height before stress /cm |
胁迫30 d Coercion for 30 days |
胁迫60 d Coercion for 60 days |
|||
| 株高 Plant height/cm |
相对生长高度 Relative growth height/cm |
株高 Plant height/cm |
相对生长高度 Relative growth height/cm |
||||
| 矿区土 | 对照 | 52.21±0.34a | 58.15±0.21ab | 5.94±0.04b | 64.95±0.05b | 12.74±0.23bc | |
| Mining soil | 轻度胁迫 | 54.84±0.04a | 61.16±0.36b | 6.23±0.13b | 71.73±0.42b | 16.89±0.16c | |
| 中度胁迫 | 53.95±0.72a | 56.29±0.05ab | 2.34±0.08ab | 60.29±0.31ab | 6.34±0.47b | ||
| 重度胁迫 | 51.49±0.65a | 48.70±0.23a | -2.79±0.32a | 48.15±0.11a | -3.34±0.35a | ||
| 非矿区土 | 对照 | 71.20±0.06a | 83.45±0.31b | 12.25±0.17c | 97.89±0.37c | 26.69±0.24c | |
| Non-mining soil | 轻度胁迫 | 68.69±0.17a | 82.4±0.02b | 13.71±0.22c | 90.63±0.25c | 21.94±0.42c | |
| 中度胁迫 | 65.72±0.31a | 68.03±0.05a | 2.31±0.16ab | 75.07±0.36b | 9.35±0.12b | ||
| 重度胁迫 | 67.23±0.12a | 63.00±0.11a | -4.23±0.08a | 61.85±0.33a | -5.38±0.52a | ||
| 不同小写字母表示柳枝稷相同土壤类型下不同土壤水分柳枝稷株高差异显著(P < 0.05); 图中数据取平均值±标准误差(n=10) | |||||||
如表 4所示, 干旱胁迫前柳枝稷在矿区土壤基质下每株分蘖数约为4个, 在非矿区土基质下约为5—7个。干旱胁迫30 d后, 矿区土壤基质中的柳枝稷每株分蘖数无变化, 非矿区土壤基质中的柳枝稷在轻度胁迫和中度胁迫下均相对增加约1个分蘖。干旱胁迫60 d后, 矿区土壤基质中的柳枝稷在中度胁迫和重度胁迫下分蘖数均减少约1个, 非矿区土壤基质中柳枝稷分蘖数除重度胁迫外增加约1个。试验表明, 干旱胁迫对柳枝稷的分蘖数无明显影响。
| 土壤类型 Soil type |
干旱胁迫程度 Drought stress |
胁迫前株高 Plant height before stress /cm |
胁迫30 d Coercion for 30 days |
胁迫60 d Coercion for 60 days |
|||
| 株高 Plant height/cm |
相对生长高度 Relative growth height/cm |
株高 Plant height/cm |
相对生长高度 Relative growth height/cm |
||||
| 矿区土 | 对照 | 3.72±0.20 | 3.72±0.20 | 4.04±0.20 | 3.72±0.20 | 3.72±0.20 | |
| Mining soil | 轻度胁迫 | 3.80±0.28 | 3.76±0.32 | 3.92±0.24 | 3.80±0.28 | 3.76±0.32 | |
| 中度胁迫 | 3.76±0.20 | 3.76±0.30 | 3.04±0.20 | 3.76±0.20 | 3.76±0.30 | ||
| 重度胁迫 | 3.96±0.24 | 3.88±0.30 | 2.92±0.30 | 3.96±0.24 | 3.88±0.30 | ||
| 非矿区土 | 对照 | 7.04±0.08 | 6.96±0.08 | 7.96±0.08 | 7.04±0.08 | 6.96±0.08 | |
| Non-mining soil | 轻度胁迫 | 4.88±0.16 | 6.04±0.23 | 6.08±0.20 | 4.88±0.16 | 6.04±0.23 | |
| 中度胁迫 | 5.92±0.15 | 6.88±0.10 | 6.92±0.10 | 5.92±0.15 | 6.88±0.10 | ||
| 重度胁迫 | 5.04±0.15 | 5.08±0.10 | 5.08±0.27 | 5.04±0.15 | 5.08±0.10 | ||
图 3表明, 干旱胁迫30 d后, 柳枝稷枯叶率随干旱胁迫的加剧而升高, 且在各干旱胁迫处理之间差异显著。矿区土中柳枝稷在对照、轻度、中度、重度胁迫下枯叶率分别为11.26%、17.89%、21.15%、42.07%, 非矿区土中柳枝稷枯叶率为5.59%、8.47%、11.70%、23.60%。干旱胁迫60 d时, 随着干旱胁迫的加剧, 矿区土中的柳枝稷枯叶率在中度胁迫之前无显著差异, 但在重度胁迫下柳枝稷枯叶率显著上升, 其枯叶率是对照组的4倍, 表明柳枝稷在中度胁迫下有明显的耐受阶段。在非矿区土中, 柳枝稷的枯叶率在轻度胁迫下最低, 在重度胁迫下最高, 且与其他处理差异显著。

|
| 图 3 柳枝稷枯叶率对土壤水分梯度胁迫的响应 Fig. 3 Response of switchgrass withered leaf rate to soil water gradient stress 不同大写字母表示在相同土壤水分下不同土壤类型之间差异显著(P < 0.05), 不同小写字母表示柳枝稷枯叶率在相同土壤类型下不同土壤水分差异显著(P < 0.05); 图中数据取平均值±标准误差(n=3) |
如图 4所示, 对照组和轻度胁迫下矿区土壤基质下柳枝稷的叶片含水量稍高于非矿区土壤基质, 而在中度和重度胁迫下表现相反。随着干旱胁迫的加剧, 矿区土壤基质中的柳枝稷叶片含水量呈现先升后降的趋势。对照、轻度胁迫和中度胁迫柳枝稷叶片相对含水量较高, 重度胁迫下其叶片相对含水量较低。表明柳枝稷在一定干旱胁迫下, 能保持叶片水分含量稳定, 但在重度胁迫下叶片缺水更加严重。

|
| 图 4 柳枝稷叶片含水量对土壤水分梯度胁迫的响应 Fig. 4 Response of switchgrass leaf water content to soil water gradient stress |
柳枝稷在矿区土和非矿区土中的地上生物量、地下生物量存在显著差异。非矿区土壤基质下的柳枝稷地上生物量比矿区土中柳枝稷的地上生物量高4—5倍。在不同干旱胁迫下, 矿区土基质下柳枝稷的地上生物量轻度胁迫达到最高, 在重度胁迫下最低, 地下生物量在对照组下为最高。随着干旱胁迫加剧, 矿区土壤基质下柳枝稷总生物量变化不明显, 但明显低于非矿区土基质中柳枝稷总生物量。
除对照组外, 矿区土壤基质中柳枝稷的根冠比随干旱胁迫的加剧逐渐增大, 而非矿区土壤基质中柳枝稷的根冠比变化不大。矿区土壤基质下, 柳枝稷的根冠比在各干旱胁迫下均比在非矿区土壤基质中大, 体现了柳枝稷对矿区土壤的适应性。
2.2.7 干旱胁迫对柳枝稷根长、根体积、根数的影响如图 6所示, 随着干旱胁迫的加剧, 在矿区土壤环境中柳枝稷的根长变化不大, 在中度胁迫下达到最长, 重度胁迫下最短。两种土壤基质中柳枝稷根体积越来越小, 分布范围为5.57—14.55 cm3。除重度胁迫外非矿区土壤中柳枝稷根体积均大于矿区土壤中柳枝稷的根体积。从差异性分析可以看出, 矿区土中柳枝稷根体积在各处理间无显著差异, 非矿区土中柳枝稷根体积存在显著差异。矿区土壤基质下柳枝稷根数先降后升, 中度胁迫下根数最少, 约为20根, 重度胁迫下最多, 约为27根。非矿区土壤基质下柳枝稷根数随着干旱程度加剧呈小幅波动变化, 各处理根数差异不显著。在各处理中, 非矿区土里的柳枝稷的根数均多于矿区土。

|
| 图 5 柳枝稷生物量对土壤水分梯度胁迫的响应 Fig. 5 Response of switchgrass biomass to soil water gradient stress DW: 干重dry weight;AGB:地上生物量aboveground biomass;BGB:地下生物量belowground biomass;不同大写字母表示在不同土壤类型下相同土壤水分差异显著(P < 0.05);不同小写字母表示柳枝稷根冠比在相同土壤类型下不同土壤水分差异显著(P < 0.05); 图中数据取平均值±标准误差(n=3) |

|
| 图 6 柳枝稷根系指标对土壤水分梯度胁迫的响应 Fig. 6 Response of switchgrass root index to soil water gradient stress |
种子萌发对环境的适应能力决定了植物种群的繁衍与发展。种子能在水分胁迫下萌发, 并保持较高的出苗率和干物质积累, 是对不良环境的适应和耐受[23]。而面对干旱、贫瘠的矿区土壤环境, 种子能否正常萌发及出苗是判断其是否能成为矿区生态修复品种的重要因素之一[24]。有研究表明, 水分胁迫显著降低冷蒿[25]、长芒草[26]等耐旱植物的发芽率, 且在高渗溶液中, 各品种的发芽势和萌发指数均会显著下降。在营养物质相对匮乏的矿区土壤环境中, 柳枝稷在轻度胁迫下其各种子萌发指标达到最高, 说明柳枝稷种子具有良好的抗贫瘠性和抗旱性。而在对照组柳枝稷种子各萌发指标均低于轻度胁迫, 这可能是因为土壤水分过多产生了水涝胁迫[27]。由此可见, 柳枝稷种子对干旱胁迫具有一定抵抗力。在本研究中, 重度胁迫土壤含水率和土壤天然含水率相差不大, 因此需要适当人工补水, 提高柳枝稷种子的萌发指标。
植物根系是植物直接吸收水分的重要器官, 它对植物的耐旱功能具有至关重要的作用[28]。纵深发达的根系系统可使植物充分吸收利用贮存在土壤中的水分, 使植物度过干旱期[29]。在本研究中, 在柳枝稷种子萌发效果最好的轻度胁迫下, 其种苗的相关生理指标反而表现最差, 这是因为在土壤水分含量合适的环境中, 柳枝稷种子根部为了吸收水分而不断伸长, 通过降低芽长减少其对养分的消耗, 反映了柳枝稷种苗对干旱的适应性。这与对干旱胁迫下高粱[30]、紫花苜蓿[31]等耐旱植物相关研究结果一致。因此, 综合柳枝稷种子萌发状况和种苗生长特性, 表明柳枝稷种子和种苗具有较强的抗旱性, 但在极端干旱的环境下, 需要及时补充适量的水分, 保证种子的正常萌发。
3.2 干旱胁迫对矿区土壤环境下柳枝稷生长特性的影响植物在受到干旱胁迫时会通过改变自身形态适应环境的变化[32]。耐旱植被调整自身生理结构以减少体内水分散失和增强水分吸取能力[33]。株高、分蘖数、枯叶率可作为衡量生长状态的重要指标[34]。有研究表明, 干旱胁迫使玉米[35]、蒙古莸[36]及羊草[37]等耐旱植被株高降低、枯叶率增加, 与柳枝稷的研究结果一致。叶片含水量反映了植物水分状况及植物保水力, 叶片相对含水量可以有效反映植物叶片水分状况及植物叶片保水能力[38]。干旱胁迫下, 植被叶片相对含水量越高或水分饱和亏缺越低, 其耐旱性越强[39]。在本研究中, 柳枝稷在非矿区土壤环境下叶片的保水能力以及耐旱性强于矿区土壤, 因此非矿区土壤基质中植被生长状况要好于矿区土壤中。在对照、轻度和中度干旱胁迫下, 柳枝稷叶片含水量未表现出显著性差异, 表明柳枝稷叶片具有较强的保水能力。
根系的生长发育是植物适应环境因子变化的自我调节过程[40]。植物根系是植物吸收水分的重要器官, 是植物与环境接触的重要界面[41]。苜蓿[42]、柠条[43]和沙打旺[44]等抗旱性强的植被往往具有更加发达的根系。对柳枝稷的研究表明, 柳枝稷面临轻度和中度干旱胁迫时会通过伸长根长吸收到深层土壤的水分, 重度胁迫虽然抑制了柳枝稷根长生长, 但柳枝稷会通过增加根数的方式维持养分的摄取。柳枝稷对不同的干旱胁迫会有不同的应对方式, 与朱毅[45]等研究结果一致。土壤类型同样影响着柳枝稷根系的发育。矿区土壤含盐量高、养分匮乏, 导致在其环境下柳枝稷根数、根体积、地下生物量等指标均低于非矿区土壤, 与左海涛[20]等研究结果一致。
综上所述, 矿区土壤环境下柳枝稷的生长指标较差, 干旱胁迫在一定程度上抑制柳枝稷的生长发育, 但其凭借较强的叶片保水能力以及根系对环境变化的应对机制, 使其具有在贫瘠干旱的矿区土壤基质中维持一定生物量的能力。
3.3 柳枝稷作为矿区生态恢复牧草品种的可行性矿区生态恢复的关键是植被恢复。选择抗逆性强、适应矿区环境的物种对排土场进行植被重建和生态恢复, 对维持露天矿区生态系统平衡具有重要意义[46]。柳枝稷作为新型能源植物, 具有较强的抗旱性和抗贫瘠性[47]。本试验中, 重度胁迫下矿区土壤含水率与自然含水率相差不大。矿区土壤基质中的柳枝稷在重度胁迫下其种子各萌发指标均偏低, 并且其在四组干旱胁迫中最高发芽率仅为30%, 相比于其他草本[48—49]仍较低, 但其种苗生理状况比较好。由此可见, 采用种子繁殖的方式将柳枝稷引种到矿区土壤环境中具有一定的局限性。通过对其生长特性的研究表明, 面对不同干旱胁迫, 除了生物量和根系指标外, 柳枝稷在两种土壤基质下的生理状况变化规律几乎一致。在长达60 d的重度干旱胁迫下, 柳枝稷的生长形态及各生理指标均有所下降, 但并未出现死亡。因此, 柳枝稷能凭借其抗逆性适应矿区土壤环境。由此可见, 面对矿区土壤环境, 柳枝稷具有较强的抗逆性, 将柳枝稷作为矿区生态修复的牧草品种具有一定的可行性。
| [1] |
魏同. 中国煤炭工业可持续发展的系统分析. 中国煤炭经济学院学报, 1996(1): 3-10. |
| [2] |
范英宏, 陆兆华, 程建龙, 周忠轩, 吴钢. 中国煤矿区主要生态环境问题及生态重建技术. 生态学报, 2003(10): 2144-2152. |
| [3] |
刘胜洪, 周玲艳, 杨妙贤, 刘文, 粱红. 十种耐逆植物在和平县稀土矿区生态修复中的应用. 天津农业科学, 2013, 19(7): 92-96. |
| [4] |
卞正富. 矿区土地复垦界面要素的演替规律及其调控研究. 中国土地科学, 1999(2): 7-12. |
| [5] |
李永庚, 蒋高明. 矿山废弃地生态重建研究进展. 生态学报, 2004(1): 95-100. |
| [6] |
黄文琳, 张强, 孔冬冬, 顾西辉, 孙鹏, 胡畔. 1982-2013年内蒙古地区植被物候对干旱变化的响应. 生态学报, 2019, 39(13): 4953-4965. |
| [7] |
侯新村, 范希峰, 武菊英, 左海涛. 纤维素类能源草在京郊地区的经济效益与生态价值评价. 草业学报, 2011, 20(6): 12-17. |
| [8] |
曲涛, 南志标. 作物和牧草对干旱胁迫的响应及机理研究进展. 草业学报, 2008(2): 126-135. |
| [9] |
Sanderson M A, Reed R L, Mclaughlin S B, Wullschleger S D, Tischler C R. Switchgrass as a sustainable bioenergy crop. BIORESOURCE TECHNOL, 1996, 56(1): 83-93. DOI:10.1016/0960-8524(95)00176-X |
| [10] |
刘吉利, 朱万斌, 谢光辉, 林长松, 程序. 能源作物柳枝稷研究进展. 草业学报, 2009, 18(3): 232-240. |
| [11] |
贺慧, 燕玲, 郑彬. 5种荒漠植物种子萌发特性及其吸水特性的研究. 干旱区资源与环境, 2008(1): 184-188. DOI:10.3969/j.issn.1003-7578.2008.01.036 |
| [12] |
张威震, 刘忠宽, 赵贺靖, 刘桂霞. 种苗建植的影响因素. 草业科学, 2016, 33(3): 479-488. |
| [13] |
何学青, 呼天明, 许岳飞, 冯雪, 方燕利, 龙明秀. PEG-6000模拟水分胁迫对柳枝稷种子萌发及幼苗生长的影响. 草地学报, 2014, 22(3): 556-563. |
| [14] |
曾彦军, 王彦荣, 萨仁, 田雪梅. 几种旱生灌木种子萌发对干旱胁迫的响应. 应用生态学报, 2002(8): 953-956. |
| [15] |
珊丹, 何京丽, 邢恩德, 梁占岐, 高鹏程. 干旱胁迫对柳枝稷幼苗生理特征的影响. 国际沙棘研究与开发, 2014, 12(1): 33-38. |
| [16] |
范希峰, 侯新村, 左海涛, 武菊英, 段留生. 边际土地类型及移栽方式对柳枝稷苗期生长的影响. 草业科学, 2010, 27(1): 97-102. |
| [17] |
郑敏娜, 李向林, 万里强, 何峰, 席翠玲, 陈瑞祥. 四种暖季型禾草对水分胁迫的生理响应. 中国农学通报, 2009, 25(9): 114-119. |
| [18] |
赵春桥, 陈敏, 侯新村, 朱毅, 武菊英, 范希峰. 干旱胁迫对柳枝稷生长与生理特性的影响. 干旱区资源与环境, 2015, 29(3): 126-130. |
| [19] |
徐炳成, 山仑, 黄瑾, 黄占斌. 柳枝稷和白羊草苗期水分利用与根冠比的比较. 草业学报, 2003(4): 73-77. |
| [20] |
左海涛, 李继伟, 郭斌, 宋尚有, 高旺盛. 盐分和土壤含水量对营养生长期柳枝稷的影响. 草地学报, 2009, 17(6): 760-766. |
| [21] |
范苏鲁, 苑兆和, 冯立娟, 王晓慧, 丁雪梅, 甄红丽. 干旱胁迫对大丽花生理生化指标的影响. 应用生态学报, 2011, 22(3): 651-657. |
| [22] |
李春喜, 赵广才, 代西梅, 姜丽娜, 尚玉磊. 小麦分蘖变化动态与内源激素关系的研究. 作物学报, 2000(6): 963-968. |
| [23] |
孙艳茹, 石屹, 陈国军, 闫慧峰. PEG模拟干旱胁迫下8种绿肥作物萌发特性与抗旱性评价. 草业学报, 2015, 24(3): 89-98. |
| [24] |
Platt W J. The Colonization and Formation of Equilibrium Plant Species Associations on Badger Disturbances in a Tall-Grass Prairie. ECOL MONOGR, 1975, 45(3): 285-305. |
| [25] |
李阳, 丁芳兵, 魏莹, 李倩. 水分胁迫对冷蒿种子萌发和幼苗生长的影响. 广西林业科学, 2021, 50(4): 434-439. |
| [26] |
孙清琳, 张瑞红, 易三桂, 杨柳, 李巧燕, 周继华, 来利明, 姜联合, 郑元润. 长芒草(Stipa bungeana)种子萌发与出苗对关键环境因子的响应. 生态学报, 2019, 39(6): 2034-2042. |
| [27] |
赵可夫. 植物对水涝胁迫的适应. 生物学通报, 2003(12): 11-14. |
| [28] |
Davies W J, Zhang J H. Root Signals and the Regulation of Growth and Development of Plants in Drying Soil. Annu. Rev. Plant Physiol. Plant Mol. Biol, 1991, 42(1): 55-76. |
| [29] |
王竞红, 张秀梅, 陈艾, 周蕴薇, 陈鹏, 江远芳. 紫穗槐幼苗根系生理特性和解剖结构对PEG-6000模拟干旱的响应. 生态学报, 2018, 38(2): 511-517. |
| [30] |
王志恒, 魏玉清, 邹芳, 黄思麒, 杨秀柳, 郝正刚. PEG-6000和盐碱胁迫对甜高粱种子萌发影响研究. 种子, 2019, 38(5): 39-43. |
| [31] |
李文娆, 张岁岐, 山仑. 水分胁迫下紫花苜蓿和高粱种子萌发特性及幼苗耐旱性. 生态学报, 2009, 29(6): 3066-3074. |
| [32] |
席璐璐, 缑倩倩, 王国华, 宋冰. 荒漠绿洲过渡带一年生草本植物对干旱胁迫的响应. 生态学报, 2021, 41(13): 5425-5434. |
| [33] |
Basu S, Ramegowda V, Kumar A, Pereira A. Plant adaptation to drought stress. F1000Research, 2016, 5: F1000-F1554. |
| [34] |
李庆康, 马克平. 植物群落演替过程中植物生理生态学特性及其主要环境因子的变化. 植物生态学报, 2002(S1): 9-19. |
| [35] |
郑盛华, 严昌荣. 水分胁迫对玉米苗期生理和形态特性的影响. 生态学报, 2006(4): 1138-1143. |
| [36] |
谢乾瑾, 夏新莉, 刘超, 尹伟伦. 水分胁迫对不同种源蒙古莸光合特性与生长的影响. 林业科学研究, 2010, 23(4): 567-573. |
| [37] |
李怡, 侯向阳, 武自念, 任卫波, 赵青山, 梁潇. 羊草种质资源抗旱性综合评价. 中国草地学报, 2019, 41(1): 75-82. |
| [38] |
史燕山, 骆建霞, 王煦, 赵坤平, 刘玉冬, 黄俊轩. 5种草本地被植物抗旱性研究. 西北农林科技大学学报(自然科学版), 2005(5): 130-134. |
| [39] |
吴阿贵. 干旱胁迫下红叶李与绿叶李抗旱生理特性比较. 现代农业科技, 2007(16): 21-22. |
| [40] |
Carrow R N. Drought avoidance characteristics of diverse tall fescue cultivars. CROP SCI, 1996, 36(2): 371-377. |
| [41] |
丁红, 张智猛, 戴良香, 康涛, 慈敦伟, 宋文武. 干旱胁迫对花生根系生长发育和生理特性的影响. 应用生态学报, 2013, 24(6): 1586-1592. |
| [42] |
张翠梅, 师尚礼, 刘珍, 杨帆, 张振科. 干旱胁迫对不同抗旱性苜蓿品种根系形态及解剖结构的影响. 草业学报, 2019, 28(5): 79-89. |
| [43] |
左志严, 格日乐, 乌仁图雅, 王娟. 内蒙古中西部3种乡土植物根系抗拉力学特性的对比研究. 水土保持通报, 2015, 35(2): 52-57. |
| [44] |
许爱云, 曹兵, 谢云. 干旱风沙区煤炭基地12种草本植物对干旱胁迫的生理生态响应及抗旱性评价. 草业学报, 2020, 29(10): 22-34. |
| [45] |
朱毅, 范希峰, 武菊英, 段留生, 侯新村. 水分胁迫对柳枝稷生长和生物质品质的影响. 中国农业大学学报, 2012, 17(2): 59-64. |
| [46] |
杨勤学, 赵冰清, 郭东罡. 中国北方露天煤矿区植被恢复研究进展. 生态学杂志, 2015, 34(4): 1152-1157. |
| [47] |
杜菲, 杨富裕, M·D·Casler, 张蕴薇. 美国能源草柳枝稷的研究进展. 安徽农业科学, 2010, 38(35): 20334-20339. |
| [48] |
杨景宁, 王彦荣. PEG模拟干旱胁迫对四种荒漠植物种子萌发的影响. 草业学报, 2012, 21(6): 23-29. |
| [49] |
王慧慧, 王普昶, 赵钢, 孙秋, 龙忠富, 张瑜. 干旱胁迫下白刺花种子大小与萌发对策. 生态学报, 2016, 36(2): 335-341. |