 2022, Vol. 42
2022, Vol. 42文章信息
- 温智峰, 魏识广, 李林, 叶万辉, 练琚愉
- WEN Zhifeng, WEI Shiguang, LI Lin, YE Wanhui, LIAN Juyu
- 亚热带不同纬度森林群落物种空间分布格局
- Spatial distribution patterns of forest community species at different latitudes in the subtropics
- 生态学报. 2022, 42(22): 9153-9164
- Acta Ecologica Sinica. 2022, 42(22): 9153-9164
- http://dx.doi.org/10.5846/stxb202106121570
-
文章历史
- 收稿日期: 2021-06-12
- 网络出版日期: 2022-07-12





2. 广西师范大学珍稀濒危动植物生态与环境保护教育部重点实验室, 桂林 541006;
3. 华南植物园中国科学院退化生态系统植被恢复与管理重点实验室, 广州 510650;
4. 中国科学院核心植物园植物生态中心, 广州 510650
2. Key Laboratory of Ecology of Rare and Endangered Species and Environment Protection (Guangxi Normal University), Ministry of Education, Guilin 541006, China;
3. Key Laboratory of Vegetation Restoration and Management of South China Botanical Garden of Degraded Ecosystems, Chinese Academy of Science, Guangzhou 510650, China;
4. Center of Plant Ecology, Core Botanical Gardens, Chinese Academy of Sciences, Guangzhou 510650, China
物种个体的空间分布是生物多样性监测和研究的核心, 个体在群落中聚集或分散的程度, 体现了物种对生境的适应情况[1]。植物个体的空间分布格局是种群自身特性、种间关系和生境因素综合作用的结果[2], 是研究种群特征、种间关系、种群与生境因子相互影响的重要手段[3], 因此群落中物种的空间分布格局及其形成机制一直是植物生态学研究的热点问题[4]。揭示植物群落中物种的空间分布格局及其成因, 可以为群落的更新、演替及多样性维持机制提供重要线索[5]。
森林群落中物种个体呈现的空间分布格局是由多种生态过程共同作用的结果[6], 也是森林群落中多种多样性维持机制综合作用的最直接体现[4]。因此, 通过了解物种个体在群落中的分布可以获取种群的空间分布特征, 能够进一步揭示格局产生的生态学过程, 例如:研究生境异质性、扩散限制、密度制约、随机作用等过程对群落中物种空间分布格局形成的作用[7], 可以解释不同群落物种共存机制和多样性的维持状况[8]。
物种空间分布格局的研究主要集中在群落中的优势种。优势种既作为群落“骨架”, 又影响群落类型[9], 对群落的结构具有决定作用[10]。分析群落中优势种群的空间分布格局, 可以阐明群落生境的形成和维持机制, 群落的稳定性以及更新和演替的规律[11—12]。此外, 由于优势种在群落中具有数量优势, 满足空间点格局统计模型的样本精度要求, 所以通常被选作代表群落多样性格局分布状况的研究对象。相反, 群落中低多度物种的空间格局分析鲜有报道, 现状为:一方面由于低多度物种个体数量少, 传统意义上以物种为单位统计其空间分布格局时受限于模型统计的样本量不足, 为确保统计精度而舍弃低多度物种;另一方面, 研究发现群落中物种的丰富度格局主要由大量的低多度物种决定, 在物种多样性丰富的群落中低多度物种所占的比例不容忽视[13]。低多度物种是群落生物多样性中非常重要的组分, 研究其空间分布格局及成因, 可以为生物多样性保护提供可靠依据, 特别是为濒危物种保护提供理论基础, 所以对群落的空间分布格局相关研究需要把优势物种和低多度物种同时纳入研究范围。
目前, 关于高丰富度群落中物种空间格局分布的研究大多都来自于热带地区[14], 美国Smithsonian研究院CTFS(The Center for Tropical Forest Science)已经建成热带森林生物多样性监测网络, 进行了森林生态系统群落分布、结构组成和物种多样性等方面的研究[15]。为了揭示其他气候带生境中的物种分布和物种共存机制, 我国的生态学家借鉴CTFS大样地建设方法, 建成了中国森林生物多样性监测网络, 取得了一系列研究成果, 如:在温带, 对长白山次生杨桦林种群的空间点格局分析结果证明了密度制约是调节温带树木空间分布格局的主要机制[16];北京东灵山落叶阔叶林群落中物种的空间格局分布研究发现了树木径级大小在空间格局分布上互补且不同径级个体占据不同生态位[17]。在亚热带, 对古田山大样地群落的研究证实了种子性状是影响物种空间分布格局的重要因素[18];对鼎湖山常绿阔叶林群落的分析证明了聚集分布是其主要空间分布格局类型[14], 大径级物种个体的空间分布趋向于随机分布[15]。在热带, 尖峰岭热带山地雨林物种多样性空间分布格局具有明显的尺度效应[19], 小径级植株比大径级植株对生境变化更加敏感, 物种空间格局受到多种因素影响[20]。以上研究大多是分散在各个纬度带上的独立讨论, 所得结果虽然较好地解释了对应地带群落的研究问题, 但是由于缺乏研究尺度上的连续性, 结论有待基于不同纬度群落的连续研究进行验证和拓展。
本文以我国亚热带区域4个纬度上6个1 hm2样地群落中所有胸径(DBH)≥1 cm的物种为研究对象, 分析各群落中物种空间分布格局的尺度效应及其随纬度变化规律;检验各群落物种多度、物种胸径空间聚集程度之间的相关性及其纬度变化规律;最后探讨影响亚热带森林群落物种分布格局形成的因素。旨在揭示我国亚热带森林群落的物种空间分布格局及其成因, 探讨其共性规律和个性差异, 为该区域的生物多样性保护和监测工作提供科学依据。
1 研究区域 1.1 研究区域概况本研究选取中国亚热带季风气候区域4个纬度带上6个1 hm2典型森林群落, 物种数据来源于中国科学院华南植物园生态研究中心的数据共享, 调查时间跨度为2010年到2012年[21]。各群落基本信息见表 1。
| 群落 Community |
经度 Longitude |
纬度 Latitude |
海拔 Altitude/m |
总物种数 Total number of species |
总物种多度 Total species abundance |
多度≥10株物种数 Number of species with abundance≥10 |
单个体物种数 Number of individual species |
| 井冈山常绿阔叶林 Jinggang Mountain evergreen broad-leaved forest |
114°11′E | 26°23′N | 450—500 | 110 | 3947 | 60 | 23 |
| 猫儿山针阔混交林 Maoer Mountain mixed needle and broad forest |
110°30′E | 25°56′N | 1900—1950 | 81 | 2650 | 33 | 12 |
| 猫儿山常绿阔叶林 Maoer Mountain evergreen broad-leaved forest |
110°30′E | 25°56′N | 1450—1500 | 125 | 2465 | 47 | 13 |
| 南岭常绿阔叶林 Nanling Mountain evergreen broad-leaved forest |
112°75′E | 24°39′N | 1000—1100 | 165 | 2893 | 51 | 34 |
| 鼎湖山常绿阔叶林 Dinghu Mountain evergreen broad-leaved forest |
112°31′E | 23°10′N | 250—300 | 95 | 3608 | 37 | 19 |
| 鼎湖山针阔混交林 Dinghu Mountain mixed needle and broad forest |
112°31′E | 23°10′N | 30—40 | 60 | 3120 | 23 | 17 |
各样地的建设和野外调查均按照CTFS国际标准进行。6个面积1 hm2的样地均采用相邻格子法设置25个20 m×20 m的样方, 每个样方再细分为16个5 m×5 m的小样方。以5 m×5 m小样方为基本单元, 对样方内植株进行每木标尺测量, 标记每个胸径(DBH)≥1 cm的个体及其分枝, 记录物种的名称、胸径、坐标、生长状况等信息, 并建立专项数据库。
2 研究方法 2.1 空间点格局分析方法采用环形取样方式, 研究种群空间分布格局采用Ripley′s K(t)方程的变换L(t)方程进行分析[22], 方程的公式表达如下:

|
(1) |
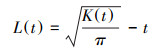
|
(2) |
式中, A为样地面积;n为样地内物种个数;t为距离尺度;Uij为植物两点i和j之间的距离;当Uij≤t时, Ir(Uij)=1, 反之为0;Wij为以i为圆心Uij为半径的圆在A中弧长与整个圆周长的比值, 表示一个植物被观测到的概率[23]。
Ripley′s K(t)方程可以用来解释以t为半径圆中的物种分布模式, 而在实际中为了稳定方差, 简化解释, 经常使用L(t)方程进行分析[24]。L(t)方程以植物个体在样地中的实际坐标为基本数据, 在分析过程中能最大限度地利用个体在二维空间上的坐标, 其最大优势在于多尺度的空间格局分析。在均质泊松分布假设下, L(t)的期望值等于0, 如果L(t)>0, 则物种呈聚集分布, 如果L(t)<0, 物种则呈均匀分布, 若L(t)=0, 则为随机分布[23]。本文选用均质泊松分布零模型(CSR), 通过Monte-Carlo随机排列999次得到99%的置信区间计算分布的上下包迹线, 若计算出的L(t)值曲线在上包迹线以上时, 为显著聚集分布格局, 若在上下包迹线之间时, 为显著随机分布格局, 若在下包迹线以下时, 为显著均匀分布格局。由于L(t)方程采用环形取样法, 样地最大边长为100 m, 为消除边缘效应, 取样的最大尺度设置为50 m。
2.2 低多度与单个体物种的空间点格局分析为了满足L(t)方程的统计精度, 将各群落中多度≥10的植物以物种为单位统计, 进行空间点格局统计分析;多度 < 10的物种则划分为单个体种组(多度=1)和低多度种组(多度=2—9), 满足统计精度后分别对其进行空间点格局统计分析。
空间点格局分析使用R 4.0.0软件及其下的“splancs”包来编程计算(R Core Team 2020. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL https://www.R-project.org/.)。
2.3 多度和胸径与空间聚集程度的关系分析因为相邻尺度的L(t)值之间高度相关, 所以本研究选取L10(10 m取样尺度上的统计值)作为表征各群落物种空间聚集程度的指标[1]。物种多度与空间聚集程度指标L10的关系采用双变量Pearson相关性分析方法。对各群落的物种多度进行自然对数转换, 并用多度的对数值与空间聚集程度指标L10进行曲线拟合。物种胸径(DBH)与空间聚集程度的关系采用最大DBH、最小DBH和平均DBH与空间聚集程度指标L10进行多元线性回归分析。
使用SPSS 26进行相关性分析和多元线性回归分析, 使用Origin 2018软件绘图。
3 结果 3.1 各群落多度≥10株物种空间分布格局及其变化规律各群落中物种显著聚集分布比例具有相同的尺度变化趋势, 均为随尺度先增大后降低(图 1)。其中, 猫儿山和鼎湖山2个针阔混交林群落在20—25 m尺度时显著聚集分布比例达到峰值, 4个常绿阔叶林群落均在10 m尺度时达到峰值。显著聚集分布比例变化幅度随纬度由高到低呈现先增后减的趋势, 纬度从高到低各群落的变化幅度为:井冈山常绿阔叶林33.33%;猫儿山针阔混交林和常绿阔叶林分别为39.39%、53.19%;南岭常绿阔叶林43.14%;鼎湖山常绿阔叶林和针阔混交林分别为29.73%、27.27%。

|
| 图 1 各群落中物种显著聚集分布比例随尺度变化规律 Fig. 1 The change of the proportion of significant aggregation distribution of species in each community with scale |
在不同取样尺度上各群落物种空间分布格局类型比例见表 2。不同纬度的群落均在小尺度上以显著聚集分布为主, 大尺度上以随机分布为主, 显著均匀分布仅出现在较大取样尺度时的部分群落, 占比极少。具有明显尺度效应。4个常绿阔叶林群落以30 m作为主要格局类型变化的尺度节点, 取样尺度 < 30 m时以显著聚集分布为主, 尺度≥30 m时以随机分布为主;2个针阔混交林群落, 主要格局类型变化的尺度节点明显更大。
| 群落 Community |
空间分布格局类型 Types of spatial distribution pattern |
尺度Scale/m | |||||||||
| 5 | 10 | 15 | 20 | 25 | 30 | 35 | 40 | 45 | 50 | ||
| 井冈山常绿阔叶林 | 显著聚集分布 | 55.00 | 60.00 | 56.67 | 50.00 | 46.67 | 43.33 | 38.33 | 35.00 | 31.67 | 26.67 |
| Jinggang Mountain evergreen | 随机分布 | 45.00 | 40.00 | 43.33 | 50.00 | 53.33 | 56.67 | 60.00 | 65.00 | 68.33 | 73.33 |
| broad-leaved forest | 显著均匀分布 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 1.67 | 0.00 | 0.00 | 0.00 |
| 猫儿山针阔混交林 | 显著聚集分布 | 63.64 | 72.73 | 69.70 | 72.73 | 69.70 | 66.67 | 63.64 | 51.52 | 39.39 | 33.33 |
| Maoer Mountain mixed needle | 随机分布 | 36.36 | 27.27 | 30.30 | 27.27 | 30.30 | 33.33 | 36.36 | 48.48 | 57.58 | 63.64 |
| and broad forest | 显著均匀分布 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 3.03 | 3.03 |
| 猫儿山常绿阔叶林 | 显著聚集分布 | 61.70 | 70.21 | 65.96 | 61.70 | 53.19 | 48.94 | 40.43 | 38.30 | 29.79 | 17.02 |
| Maoer Mountain evergreen | 随机分布 | 38.30 | 29.79 | 34.04 | 38.30 | 46.81 | 51.06 | 59.57 | 59.57 | 65.96 | 80.85 |
| broad-leaved forest | 显著均匀分布 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 2.13 | 4.26 | 2.13 |
| 南岭常绿阔叶林 | 显著聚集分布 | 47.06 | 58.82 | 49.02 | 45.10 | 41.18 | 29.41 | 23.53 | 19.61 | 15.69 | 15.69 |
| Nanling Mountain evergreen | 随机分布 | 52.94 | 41.18 | 50.98 | 54.90 | 58.82 | 70.59 | 76.47 | 80.39 | 82.35 | 84.31 |
| broad-leaved forest | 显著均匀分布 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 1.96 | 0.00 |
| 鼎湖山常绿阔叶林 | 显著聚集分布 | 54.05 | 67.57 | 56.76 | 56.76 | 54.05 | 48.65 | 48.65 | 48.65 | 43.24 | 37.84 |
| Dinghu Mountain evergreen | 随机分布 | 45.95 | 32.43 | 43.24 | 43.24 | 45.95 | 51.35 | 51.35 | 51.35 | 56.76 | 62.16 |
| broad-leaved forest | 显著均匀分布 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| 鼎湖山针阔混交林 | 显著聚集分布 | 50.00 | 50.00 | 50.00 | 59.09 | 59.09 | 54.55 | 45.45 | 45.45 | 40.91 | 31.82 |
| Dinghu Mountain mixed needle | 随机分布 | 50.00 | 50.00 | 50.00 | 40.91 | 40.91 | 45.45 | 54.55 | 54.55 | 59.09 | 68.18 |
| and broad forest | 显著均匀分布 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
各群落中单个体种组空间点格局分析结果显示(图 2):不同纬度群落中单个体种组均以随机分布为主, 格局随尺度变化规律不明显。仅井冈山常绿阔叶林群落在取样尺度接近50 m时出现显著均匀分布, 以及南岭常绿阔叶林群落在20—35 m尺度表现为显著聚集分布, 其余群落所有尺度均呈现随机分布。单个体物种的空间分布点图验证了单个体种组在大多数尺度上随机分布的空间格局(图 3)。

|
| 图 2 各群落中单个体种组的空间点格局分析结果 Fig. 2 Spatial pattern analysis results of species in each community |

|
| 图 3 各群落中单个体物种的空间分布点图 Fig. 3 Scatter plot of individual species spatial distribution in each community |
不同纬度群落低多度种组空间点格局呈现小尺度上显著聚集分布, 大尺度上随机分布甚至出现显著均匀分布的特点, 具有明显的尺度效应, 但不同纬度群落间尺度效应存在差异(图 4)。低多度种组在井冈山常绿阔叶林所有取样尺度均为随机分布, 其余群落主要格局类型变化尺度各不相同:猫儿山、南岭和鼎湖山3个常绿阔叶林群落以35 m为主要格局类型变化的尺度节点;猫儿山和鼎湖山2个针阔混交林群落主要格局类型变化的尺度节点更大, 为40—45m。规律性与各群落非低多度种组的物种相同。

|
| 图 4 各群落中低多度种组的空间点格局分析结果 Fig. 4 Spatial pattern analysis results of low abundance species in each community |
物种多度与空间聚集程度指标L10的双变量相关性分析表明(表 3):物种多度与L10均表现出负相关关系, 纬度从高到低对应群落的相关性分析P值表现出数值递增的规律, P值表征的负相关关系从极显著变化到显著, 最后到不显著。具体表现为:井冈山常绿阔叶林、猫儿山针阔混交林和常绿阔叶林群落的物种多度与L10极显著负相关(井冈山常绿阔叶林P=0.001, 猫儿山针阔混交林P=0.006, 猫儿山常绿阔叶林P=0.010);南岭常绿阔叶林群落的物种多度与L10显著负相关(P=0.048);鼎湖山常绿阔叶林和针阔混交林群落的物种多度与L10的负相关关系不显著。
| 群落 Community |
P | Pearson相关性系数 Pearson correlation coefficient |
| 井冈山常绿阔叶林Jinggang Mountain evergreen broad-leaved forest | 0.001 | -0.418** |
| 猫儿山针阔混交林Maoer Mountain mixed needle and broad forest | 0.006 | -0.467** |
| 猫儿山常绿阔叶林Maoer Mountain evergreen broad-leaved forest | 0.010 | -0.374** |
| 南岭常绿阔叶林Nanling Mountain evergreen broad-leaved forest | 0.048 | -0.278* |
| 鼎湖山常绿阔叶林Dinghu Mountain evergreen broad-leaved forest | 0.190 | -0.221 |
| 鼎湖山针阔混交林Dinghu Mountain mixed needle and broad forest | 0.061 | -0.406 |
| *表示相关性显著P < 0.05, **表示相关性极显著P < 0.01 | ||
将多度的对数值与L10进行多次拟合后发现, 幂函数的拟合优度(R2)明显高于线性拟合和其他曲线拟合方式。拟合后的幂函数曲线验证了物种多度与空间聚集程度的负相关关系(图 5), 各群落中物种的空间聚集程度随着多度的增大而降低, 物种空间聚集程度的递减速率随着多度的增大逐渐降低。

|
| 图 5 各群落物种多度与空间聚集程度的关系 Fig. 5 Relationship between species abundance and spatial aggregation degree of each community |
各群落中物种平均DBH、最大DBH、最小DBH与空间聚集程度指标L10相关性分析表明(表 4):各群落物种平均DBH与空间聚集程度L10的相关性系数均为负;最大DBH与L10的相关性系数主要为负;最小DBH与L10的相关性系数主要为正。显著性检验验证了猫儿山常绿阔叶林群落的物种平均DBH与L10极显著负相关;井冈山和猫儿山2个常绿阔叶林群落的物种最大DBH与L10极显著负相关;其他群落的相关关系均未达到显著性检验水平, 不具统计学意义。
| 群落 Community |
胸径类型 Diameter at breast height (DBH) type |
P | Pearson相关性系数 Pearson correlation coefficient |
| 井冈山常绿阔叶林 | 平均DBH | 0.136 | -0.195 |
| Jinggang Mountain evergreen broad-leaved forest | 最小DBH | 0.126 | 0.200 |
| 最大DBH | 0.013 | -0.318* | |
| 猫儿山针阔混交林 | 平均DBH | 0.433 | -0.141 |
| Maoer Mountain mixed needle and broad forest | 最小DBH | 0.583 | 0.099 |
| 最大DBH | 0.797 | 0.047 | |
| 猫儿山常绿阔叶林 | 平均DBH | 0.013 | -0.359* |
| Maoer Mountain evergreen broad-leaved forest | 最小DBH | 0.813 | 0.035 |
| 最大DBH | 0.011 | -0.367* | |
| 南岭常绿阔叶林 | 平均DBH | 0.694 | -0.056 |
| Nanling Mountain evergreen broad-leaved forest | 最小DBH | 0.171 | 0.194 |
| 最大DBH | 0.062 | -0.263 | |
| 鼎湖山常绿阔叶林 | 平均DBH | 0.278 | -0.183 |
| Dinghu Mountain evergreen broad-leaved forest | 最小DBH | 0.835 | -0.035 |
| 最大DBH | 0.337 | -0.162 | |
| 鼎湖山针阔混交林 | 平均DBH | 0.725 | -0.080 |
| Dinghu Mountain mixed needle and broad forest | 最小DBH | 0.688 | -0.078 |
| 最大DBH | 0.731 | -0.125 | |
| *表示相关性显著P < 0.05 | |||
同时, 对物种的平均DBH、最大DBH、最小DBH与L10进行多元线性回归分析, 发现随着纬度的由高到低, 各群落物种胸径对空间聚集程度的解释率总体趋势降低(图 6), 由最高纬度的19.7%(井冈山常绿阔叶林)降低到了最低纬度的3.9%(鼎湖山针阔混交林), 说明物种胸径对空间聚集程度的影响在高纬度群落更加明显。

|
| 图 6 各群落的物种胸径对空间聚集程度的解释率 Fig. 6 The explanation rate of species diameter at breast height of each community to spatial aggregation degree |
本研究中亚热带不同纬度森林群落物种的分布格局规律验证了物种空间分布格局具有尺度依赖性[25], 并且不同群落中物种空间分布格局的成因存在差异[26]。随取样尺度增大, 显著聚集分布比例先增大后减小, 小尺度上以显著聚集分布为主, 大尺度上以随机分布为主, 符合物种在森林群落中空间分布的一般规律[4]。物种显著聚集分布比例的变化幅度能够在一定程度上反映森林群落空间分布格局的稳定性[27], 不同纬度各群落物种显著聚集分布比例随取样尺度的变化趋势虽然相同, 但变化幅度存在差异:纬度最高的井冈山常绿阔叶林群落变化幅度处于中间水平, 处于中间纬度的猫儿山常绿阔叶林、针阔混交林以及南岭常绿阔叶林群落变化幅度较大, 纬度最低的鼎湖山常绿阔叶林和针阔混交林群落变化幅度较小, 推测是由于纬度最低群落具有较高的生物多样性, 因此具有较高稳定性和较强抵御能力。在同一纬度, 不同海拔的森林群落林型会存在差异, 而本研究发现猫儿山和鼎湖山的常绿阔叶林和针阔混交林群落物种空间分布格局存在差异性, 常绿阔叶林群落显著聚集分布比例随尺度变化幅度明显高于针阔混交林群落;针阔混交林群落主要格局类型变化的尺度节点明显大于常绿阔叶林群落(图 1), 推测林型的不同可以通过影响林分密度以及群落物种组成, 进而影响物种空间分布格局, 使其有所差异[28], 因此认为林型也是群落影响物种空间分布格局的因素之一。
目前普遍认为生境异质性、扩散限制、密度制约以及随机作用等是影响群落物种空间分布格局的重要因素和群落多样性维持的驱动机制[29]。森林群落的生物多样性分布具有明显的纬度梯度规律[30], 纬度不同时, 群落中的物种类型、气温、太阳辐射强度、地形因子以及其他环境因素均会随之发生改变, 进而引起物种的空间分布格局随纬度梯度的变化, 这可以解释不同纬度的群落虽然空间分布格局变化的趋势相同, 但变化幅度存在差异的原因, 说明生境异质性作用在亚热带森林群落中普遍存在。
聚集分布是群落优势种主要空间分布格局类型[1], 在亚热带区域6个群落中多度≥10株的物种空间分布格局也在大部分取样尺度呈现聚集分布(表 1)。而为了更全面的了解群落中所有物种的空间分布格局, 还需要关注低多度物种在群落中的分布。低多度物种由于数量稀少, 很少像优势种那样存在较强的种内竞争排斥作用[31], 小尺度上形成聚集分布格局有助于发挥低多度种群的群体效应, 增大种间竞争力来维持种群稳定[32], 但是由于低多度物种个体数量有限, 所以往往在大尺度上会表现为随机分布或均匀分布。这与本研究中低多度物种空间分布格局规律性一致:不同纬度亚热带森林群落中的低多度种组空间分布格局在小尺度上也是显著聚集分布, 随尺度增大出现随机分布, 各群落均存在物种分布的尺度效应, 但在不同纬度上存在差异(图 4)。纬度最高的井冈山常绿阔叶林群落有别于其他群落, 其低多度种组在所有尺度上均表现出随机分布格局, 未检测到聚集分布。本研究以单个体种组为单位统计分析后发现各群落中单个体物种之间的分布是随机的, 没有检测出预期的聚集分布趋势, 所体现出的随机分布还需要进一步结合亲缘种组或与单个体物种邻近物种的关系等进行深入研究。
4.2 物种多度与空间聚集程度的关系热带森林群落中的研究表明, 相同取样面积时低多度物种空间聚集程度明显高于高多度物种[1]。本研究各群落物种多度与空间聚集程度指标L10的分析结果验证了以上规律在亚热带森林群落中同样适用。由于优势种在群落中数量较多, 种间具有较强竞争排斥作用[31], 高密度聚集情况下种内竞争凸显, 受密度制约作用, 反映为空间聚集程度降低;而相对于广布的优势种, 低多度种对生境依赖性更强, 反映为空间聚集程度提高。但不同纬度各群落物种多度对空间聚集程度的影响水平并不相同, 各群落物种多度与空间聚集程度指标L10的负相关关系随纬度从高到低逐渐减弱, 说明物种多度对空间聚集程度的影响逐渐降低。推测是由于纬度降低, 生境多样性和复杂性随着群落生产力增大而提高[33], 这使影响物种空间聚集程度的因素增多, 从而使物种多度对空间聚集程度影响水平显著性降低。
4.3 物种胸径与空间聚集程度的关系群落中物种个体生长早期的空间分布格局一般表现为明显的聚集性分布, 随着个体径级的增大, 种内竞争加剧[29], 导致空间聚集程度降低, 表明胸径与空间聚集程度存在负相关关系。但本研究中不同纬度森林群落物种胸径和空间聚集程度的规律性并不完全与之相同:仅纬度较高的井冈山常绿阔叶林和猫儿山常绿阔叶林、针阔混交林群落物种胸径与空间聚集程度指标L10间显著负相关, 纬度较低群落物种胸径与空间聚集程度指标L10相关性均不显著。这说明在纬度较高的群落中, 物种胸径是影响空间聚集程度的重要因素之一, 而随纬度降低, 生境复杂性增强, 胸径对空间聚集程度的影响逐渐减弱。此外, 多元线性回归发现, 随着纬度由高到低, 物种胸径对空间聚集程度的解释率逐渐降低, 由19.7%降低到3.9%, 这一规律同样反映了胸径对空间聚集程度的影响随纬度降低而减少。群落中物种胸径和空间聚集程度的关系亦受到监测尺度、生境类型和环境因子的影响, 需综合考虑其他因素得出更可靠的规律。
4.4 亚热带森林群落的物种空间分布格局形成原因森林群落物种空间分布格局的形成受到多种机制影响[34], 而生境异质性被认为是大尺度上主要的影响因素[35], 对群落中物种空间分布和多样性格局形成及维持有重要作用[36]。本研究中不同纬度群落物种空间分布格局所表现出来的差异性有力的支持了这一观点, 例如不同纬度各群落显著聚集分布比例随取样尺度的变化幅度不同以及各群落低多度种组的空间分布格局存在差异。除生境因素以外, 物种的种子性状和扩散模式不同也会导致空间分布格局产生差异[18], 所以扩散限制同样被认为是群落格局有所差异的原因。有研究发现群落中物种的空间分布格局可能由生境异质性与扩散限制的联合作用决定[37], 但生境异质性的贡献高于扩散限制[38]。本研究各群落的物种多度和胸径与物种空间聚集程度的负相关关系也从不同侧面反映出密度制约作用也是亚热带区域物种分布格局形成的影响因素, 而大尺度上的随机分布以及单个体物种的随机分布也暗示着随机作用对群落物种分布格局有所影响。综上所述, 推测影响亚热带森林群落物种空间分布格局形成的主要原因是由于因纬度变化引起的生境异质性, 而林型、扩散限制、密度制约作用和随机作用为次要原因。
5 结论本研究对我国6个不同纬度亚热带森林群落物种空间分布格局及其成因进行研究, 所得结论如下:(1)亚热带不同纬度群落中多度≥10株物种的空间分布格局表现出相似的尺度依赖性规律:显著聚集分布比例随尺度增大而降低, 不同纬度群落的变化幅度存在差异, 纬度较低的群落稳定性更强。(2)各群落中低多度种组空间分布格局表现出小尺度显著聚集分布, 大尺度随机分布的尺度效应, 但尺度效应在不同纬度群落间存在差异;各群落单个体种组空间分布格局均以随机分布为主。(3)不同纬度各群落物种多度与空间聚集程度之间存在负相关关系, 物种多度对空间聚集程度的影响随纬度从高到低逐渐减弱。(4)较高纬度群落物种胸径与空间聚集程度之间存在负相关关系;随纬度从高到低, 物种胸径对空间聚集程度影响水平逐渐降低。对亚热带不同纬度森林群落物种空间分布格局形成原因的进一步探讨, 认为由纬度差异引起的生境异质性是影响群落物种空间分布格局产出过程的主要因素, 同时林型、扩散限制、密度制约和随机作用是影响各群落物种空间格局及多样性维持的次要因素。本研究通过不同纬度群落物种空间分布格局的分析和讨论, 揭示了亚热带森林群落物种空间分布格局类型及其成因, 为该区域的生物多样性保护工作提供了依据。
致谢: 中国科学院华南植物园黄忠良研究员、曹洪麟研究员对样地调查数据给予支持, 广东鼎湖山森林生态系统国家野外科学观测研究站、猫儿山国家自然保护区提供帮助,特此致谢。| [1] |
Condit R, Ashton P S, Baker P, Bunyavejchewin S, Gunatilleke S, Gunatilleke N, Hubbell S P, Foster R B, Itoh A, LaFrankie J V, Lee H S, Losos E, Manokaran N, Sukumar R, Yamakura T. Spatial patterns in the distribution of tropical tree species. Science, 2000, 288(5470): 1414-1418. DOI:10.1126/science.288.5470.1414 |
| [2] |
李明辉, 何风华, 刘云, 潘存德. 林分空间格局的研究方法. 生态科学, 2003, 22(1): 77-81. |
| [3] |
白聪, 闫明, 毕润成, 何艳华. 山西太岳山兴唐寺红柄白鹃梅群落优势种的空间格局分析. 植物生态学报, 2014, 38(12): 1283-1295. |
| [4] |
Greig-Smith P. Quantitative Plant Ecology. Berkeley: University of California Press, 1983, 54-57. |
| [5] |
石蒙蒙. 长白山次生林云冷杉幼苗幼树空间分布与关联性[D]. 北京: 北京林业大学, 2019.
|
| [6] |
马芳, 王顺忠, 冯金朝, 桑卫国. 北京东灵山优势种群树木死亡对空间格局与生境的影响. 生态学报, 2018, 38(21): 7669-7678. |
| [7] |
冯云, 马克明, 张育新, 祁建. 北京东灵山地区辽东栎(Quercus liaotungensis)种群生活史特征与空间分布. 生态学杂志, 2009, 28(8): 1443-1448. |
| [8] |
倪瑞强, 唐景毅, 程艳霞, 张春雨, 陈贝贝, 何怀江, 夏富才. 长白山云冷杉林主要树种空间分布及其关联性. 北京林业大学学报, 2013, 35(6): 28-35. |
| [9] |
李立, 陈建华, 任海保, 米湘成, 于明坚, 杨波. 古田山常绿阔叶林优势树种甜槠和木荷的空间格局分析. 植物生态学报, 2010, 34(3): 241-252. |
| [10] |
Heegaard E, Gjerde I, Saetersdal M. Contribution of rare and common species to richness patterns at local scales. Ecography, 2013, 36(8): 937-946. DOI:10.1111/j.1600-0587.2013.00060.x |
| [11] |
刘贵峰, 程伟燕, 刘玉平, 张淑娟. 大青沟自然保护区主要森林群落优势种的种群结构与动态. 西北植物学报, 2014, 34(4): 843-850. |
| [12] |
魏识广, 李林, 许睿, 黄忠良, 曹洪麟. 井冈山植物群落优势种空间分布格局与种间关联. 热带亚热带植物学报, 2015, 23(1): 74-80. |
| [13] |
Berg Å, Tjernberg M. Common and rare Swedish vertebrates-distribution and habitat preferences. Biodiversity & Conservation, 1996, 5(1): 101-128. |
| [14] |
Li L, Huang Z L, Ye W H, Cao H L, Wei S G, Wang Z G, Lian J Y, Sun I F, Ma K P, He F L. Spatial distributions of tree species in a subtropical forest of China. Oikos, 2009, 118(4): 495-502. |
| [15] |
叶万辉, 曹洪麟, 黄忠良, 练琚愉, 王志高, 李林, 魏识广, 王章明. 鼎湖山南亚热带常绿阔叶林20公顷样地群落特征研究. 植物生态学报, 2008, 32(2): 274-286. |
| [16] |
安璐, 吴兆飞, 范春雨, 张春雨, 赵秀海. 长白山次生杨桦林种群空间点格局及密度制约效应. 生态学报, 2021, 41(4): 1461-1471. |
| [17] |
马芳, 王顺忠, 冯金朝, 桑卫国. 北京东灵山暖温带落叶阔叶林枯立木与活立木空间分布格局. 生态学报, 2018, 38(16): 5717-5725. |
| [18] |
姚蓓, 余建平, 刘晓娟, 米湘成, 马克平. 亚热带常绿阔叶林种子性状对木本植物聚集格局的影响. 生物多样性, 2015, 23(2): 157-166. |
| [19] |
李艳朋, 许涵, 李意德, 骆土寿, 陈德祥, 周璋, 林明献, 杨怀. 海南尖峰岭热带山地雨林物种多样性空间分布格局的尺度效应. 植物生态学报, 2016, 40(9): 861-870. |
| [20] |
梁爽, 许涵, 林家怡, 李意德, 林明献. 尖峰岭热带山地雨林优势树种白颜树空间分布格局. 植物生态学报, 2014, 38(12): 1273-1282. |
| [21] |
邹顺, 耿卫欣, 张倩媚, 周国逸, 刘世忠, 褚国伟. 1992-2015年鼎湖山季风常绿阔叶林乔木物种组成数据集. 中国科学数据, 2019, 4(4): 161-167. |
| [22] |
Diggle P. Statistical analysis of spatial point patterns. London: Academic Press, 1983.
|
| [23] |
张金屯. 数量生态学 (2版). 北京: 科学出版社, 2011: 299-302.
|
| [24] |
Li L, Wei S G, Huang Z L, Ye W H, Cao H L. Spatial patterns and interspecific associations of three canopy species at different life stages in a subtropical forest, China. Journal of Integrative Plant Biology, 2008, 50(9): 1140-1150. |
| [25] |
王本洋, 余世孝, 王永繁. 种群分布格局的各向异性分析. 中山大学学报(自然科学版), 2006, 45(2): 83-87. |
| [26] |
张钦弟, 张金屯, 张斌, 程佳佳, 田世广, 苏日古嘎. 东灵山华北落叶松人工林的分布格局及环境解释. 武汉植物学研究, 2010, 28(5): 577-582. |
| [27] |
周梦丽, 张青, 亢新刚, 郭韦韦, 季蕾, 李成富. 基于空间结构指数的不同森林群落稳定性研究. 植物科学学报, 2016, 34(5): 724-733. |
| [28] |
玉宝, 乌吉斯古楞, 王百田, 王立明, 达布希拉图. 兴安落叶松天然林不同林分结构林木水平分布格局特征研究. 林业科学研究, 2010, 23(1): 83-88. |
| [29] |
吴初平, 袁位高, 盛卫星, 黄玉洁, 陈庆标, 沈爱华, 朱锦茹, 江波. 浙江省典型天然次生林主要树种空间分布格局及其关联性. 生态学报, 2018, 38(2): 537-549. |
| [30] |
李林, 魏识广, 练琚愉, 曹洪麟. 亚热带不同纬度植物群落物种多样性分布规律. 生态学报, 2020, 40(4): 1249-1257. |
| [31] |
毛子昆, 郝占庆, 原作强, 蔺菲, 叶吉, 匡旭, 王绪高. 物种聚集分布与种间关系的多度不对称性. 中国科学: 生命科学, 2020, 50(4): 381-390. |
| [32] |
韩路, 王海珍, 彭杰, 莫治新. 塔里木河上游天然胡杨林种群空间分布格局与动态研究. 西北植物学报, 2007, 27(8): 1668-1673. |
| [33] |
郝亚军. 中国东部森林群落物种多样性纬度梯度格局形成机制研究[D]. 杭州: 浙江大学, 2020.
|
| [34] |
张忠华, 胡刚, 倪健. 茂兰喀斯特常绿落叶阔叶混交林树种的空间分布格局及其分形特征. 生态学报, 2015, 35(24): 8221-8230. |
| [35] |
Takyu M, Aiba S I, Kitayama K. Effects of topography on tropical lower montane forests under different geological conditions on Mount Kinabalu, Borneo. Plant Ecology, 2002, 159(1): 35-49. |
| [36] |
朱强, 艾训儒, 吴漫玲, 黄小, 王进, 彭宗林. 七姊妹山常绿落叶阔叶混交林物种多样性格局及其环境解释. 西北植物学报, 2019, 39(8): 1491-1502. |
| [37] |
沈国春. 生境异质性和扩散限制对亚热带和热带森林物种多样性维持的作用[D]. 杭州: 浙江大学, 2010.
|
| [38] |
李林, 魏识广, 马姜明, 叶万辉, 练琚愉. 生境异质性和扩散限制对南亚热带常绿阔叶林群落物种多样性的相对作用. 林业科学, 2020, 56(10): 1-10. |