 2022, Vol. 42
2022, Vol. 42文章信息
- 朱昊阳, 李洪宇, 王晓蕾, 姜婷, 孙林, 罗毅
- ZHU Haoyang, LI Hongyu, WANG Xiaolei, JIANG Ting, SUN Lin, LUO Yi
- 黄土高原油松冠层气孔导度和蒸腾变化特征与模拟
- Characteristics and modeling of canopy stomatal conductance and transpiration of Pinus tabuliformis on the Loess Plateau
- 生态学报. 2022, 42(22): 9130-9142
- Acta Ecologica Sinica. 2022, 42(22): 9130-9142
- http://dx.doi.org/10.5846/stxb202106041484
-
文章历史
- 收稿日期: 2021-06-04





2. 中国科学院大学资源与环境学院, 北京 100190
2. College of Resources and Environment, University of Chinese Academy of Sciences, Beijing 100190, China
黄土高原地处干旱半干旱区, 生态环境脆弱、水土流失严重, 是我国重要的生态建设区。近年来随着大规模退耕还林(草)和天然林保护工程的实施, 黄土高原植被类型明显转变[1], 植被覆盖度从1999年的32%增长到了2013年的60%[2]。但植被恢复也造成了新的问题:植被覆盖度的增加引起了黄河流域干支流径流量的持续降低[3];外来树种的引进和高密度种植引起了土壤水分下降, 限制了树木生长[4]。植物蒸腾耗水占森林生态系统总蒸散量的60%以上[5], 有必要研究黄土高原地区造林物种的蒸腾耗水规律及其对环境因子的响应, 以实现人工林可持续经营。
树木以水分利用最优[6]和避免空穴栓塞[7]为目标调节气孔行为、控制蒸腾耗水, 以响应环境变化。冠层气孔导度反映了林冠层气孔状况[8], 是蒸腾和光合等生理过程模拟的关键参数[9]。据此, 结合气象条件、土壤水分条件和植物生理特征[10], 建立冠层气孔导度与环境因子的响应关系, 阐释环境因素对蒸腾的作用机制, 是森林生态系统水循环研究的主要手段之一[11]。
油松(Pinus tabuliformis)根系发达、耐寒抗旱, 是黄土高原主要造林树种之一[12]。已有研究在干旱半干旱区探究了油松冠层蒸腾特征及影响因子[13—15], 并基于Penman-Monteith公式和Jarvis型气孔导度模型模拟了其冠层蒸腾的日内变化[16], 但模型未考虑土壤水分对冠层气孔导度的影响, 难以准确模拟水分胁迫条件下的冠层蒸腾[17]。土壤水分是黄土高原植物生长的重要限制因素[18], 是冠层气孔导度在长时间尺度下的主要限制因子[19]。因此, 需考虑该地区油松冠层气孔导度对土壤水分条件的响应, 以准确模拟其冠层蒸腾。
本文基于树干液流观测数据, 探究黄土高原地区油松冠层气孔导度和蒸腾在不同时间尺度下的变化特征;分析冠层气孔导度和蒸腾对气象因子和土壤水分的响应规律;结合Penman-Monteith公式与Jarvis型模型, 基于冠层气孔导度计算冠层蒸腾, 为开展森林冠层水汽交换过程的机理性研究提供科学依据。
1 材料与方法 1.1 研究区概况研究区位于陕西省扶风县野河山森林保护区(34°31.76′N, 107°54.67′E), 地处中国黄土高原南部, 海拔1090 m。该地区属暖温带半湿润大陆性季风气候区, 年平均气温为12.7℃, 多年平均降水量为580 mm, 主要集中于6—9月, 约占全年降水总量的70%。研究区土壤主要由粉砂壤土组成, 砂粒、粉粒和黏粒含量分别为5.8%, 73.3%和20.9%, 黄土层厚度超过50 m[20]。
野河山森林保护区占地10996hm2, 原为公社集体耕地, 自1985年居民搬迁后, 封育种植刺槐(Robinia pseudoacacia)、油松(Pinus tabuliformis)和侧柏(Platycladus orientalis)等树种。本研究选取于1994年种植的人工油松林作为研究对象, 油松林密度为2800株/hm2, 尚未抚育间伐, 人为扰动弱。乔木冠层郁闭度0.7, 平均树高7.2 m, 平均胸径11.9 cm, 叶面积指数(LAI)为2.2 m2/m2, 林下植被发育较差。
1.2 研究方法 1.2.1 油松树干液流观测Granier[21]提出TDP测量方法能长期、准确、稳定地获取树干液流数据, 已被广泛地应用于植物蒸腾耗水及冠层气孔导度的研究中。按照树木生长良好、树干通直的基本原则, 选取多株油松作为典型样树, 在2015年至2018年生长季内(4—10月)进行树干液流测定(表 1)。树干液流采用SF插针式液流测量系统(Campbell Scientific Inc. Logan, Utah, USA)测定, 将探针统一安装在树干南向离地面高1.3 m处, 安装时去除油松表面硬化树皮, 用铝箔和防雨装置覆盖探头以避免太阳辐射和雨水的影响。数据采用CR1000(Campbell Scientific Inc. Logan, Utah, USA)数据采集器记录, 数据测定和存储时间间隔为10 min。
| 编号 Number |
胸径 Diameter at breast height/cm |
边材厚度 The thickness of the sapwood/cm |
边材面积 Sapwood area/cm2 |
| 1 | 11.5 | 5.31 | 73.63 |
| 2 | 13.1 | 5.56 | 86.81 |
| 3 | 12.1 | 5.28 | 78.52 |
| 4 | 9.8 | 4.61 | 60.15 |
在样地附近另选取8棵油松, 于树干胸径处利用生长锥取样确定边材面积, 拟合胸径-边材面积关系式(Ac=0.96·DBH1.73, R2=0.79, n= 12)。
1.2.2 环境因子观测气象数据由自动气象观测装置同步监测。降水量(P, mm)观测装置为T-200B (Geonor, Norway)型雨雪量计, 布设于无遮蔽林间空地内。其他气象传感器位于架设在样地附近的16 m高塔上:太阳辐射(Rad, W/m2)传感器型号为CNR4 (Kippzonen, Netherlands), 空气相对湿度(RH, %)和气温(T, ℃) 传感器型号为HMP155A (Vaisala, Finland), 风速观测装置为CSAT3 (Campbell Scientific Inc. Logan, UT, USA) 型三维超声风速仪。数据采集器为CR3000 (Campbell Scientific Inc. Logan, UT, USA), 所有气象因子测定和存储时间间隔均为10 min。
土壤体积含水量(θ, cm3/cm3)测定传感器型号为HydraProbe Ⅱ(Stevens, USA), 测定深度设定为5, 15, 35, 50, 80, 110, 140, 180 cm和230 cm, 其中样地土壤230 cm深处存在浆石层。数据采集器为CR1000 (Campbell Scientific Inc. Logan, UT, USA), 数据测定和存储时间间隔为10 min。利用环刀取原状土样, 使用离心机法(CR21G)测定土壤水分特征曲线。
1.3 数据处理 1.3.1 环境因子预处理冠层饱和水汽压亏缺(VPD, kPa)计算采用公式:
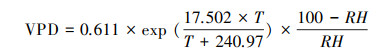
|
(1) |
式中, RH为空气相对湿度(%), T为气温(℃)。
考虑到油松根系在不同深度土层内分布的差异, 采用根区土壤有效含水率(Relative Extractable Water, REW)表征土壤水的可利用程度:
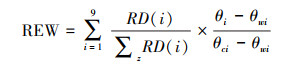
|
(2) |
式中, θi为i层土壤含水量(cm3/cm3);θci为i层田间持水量(cm3/cm3), 根据所测得土壤水分特征曲线, 取土壤水势为-33 kPa时的土壤含水量;θwi为i层萎蔫含水量(cm3/cm3), 取土壤水势为-1500 kPa时的土壤含水量;RD(i) 为i层土层的细根生物量密度(Fine Root Biomass Density, FRB), 取值参考黄土高原地区油松根系分布的相关研究[22]。
1.3.2 林分冠层蒸腾计算采用Granier[21]提出的公式及参数值计算油松小时液流速率:
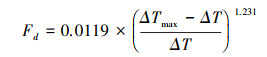
|
(3) |
式中, Fd为液流速率(g cm-2 h-1);ΔT为两探针之间的实测温差(℃);ΔTmax为两探针之间的日最大温差(℃)。
分别将不同时滞(15, 30, 60 min)的树干液流速率与太阳辐射和VPD进行相关分析并比较相关系数, 结果表明油松树干液流并无明显时滞。将太阳辐射低于5 W/m2时产生的液流定义为夜间液流[23], 发现样地油松夜间液流仅占3.5%, 低于Chen等[24]计算的油松夜间液流占比(12.3%—13.1%)。因此, 忽略夜间液流对油松蒸腾计算的影响。
结合胸径-边材面积关系式与平均液流速率计算林分小时冠层蒸腾速率:
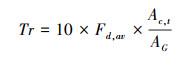
|
(4) |

|
(5) |
式中, Fd, i为第i棵树液流速率(g cm-2 h-1);Ac, i为第i棵树的边材面积;Tr为林分冠层蒸腾(mm/h);Fd, av为平均液流速率(g cm-2 h-1);Ac, t为林分边材总面积(m2);AG为林分面积(m2)。将林分小时冠层蒸腾速率在日内合计得到林分日冠层蒸腾速率。
1.3.3 冠层平均气孔导度计算本文采用Monteith等[25]基于Penman-Monteith公式提出的简化公式计算冠层平均气孔导度:
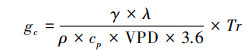
|
(6) |
式中, gc为冠层平均气孔导度(mm/s);γ为干湿表常数(kPa/℃);λ为汽化潜热(MJ/kg);ρ为常温下平均大气密度(kg/m3);cp为定压比热容(MJ kg-1 ℃-1);VPD为饱和水汽压差(kPa)。该公式适用于冠层与林下通风条件较好, 且冠层表面空气动力学导度远大于冠层平均气孔导度的情况。
1.4 模型构建 1.4.1 冠层气孔导度与冠层蒸腾模型Jarvis型模型假设各环境因子对叶片气孔的影响相互独立, 将气孔导度表示为与环境因子相关的经验方程的连乘, 可直观反映植物叶片气孔导度与辐射、饱和水汽压差和土壤含水量等环境因子的关系。本文采用Jarvis型模型模拟冠层平均气孔导度:
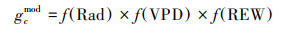
|
(7) |
式中, gcmod为模拟冠层平均气孔导度(mm/s), f(Rad)、f(VPD)、f(REW) 分别为gc对太阳辐射、水汽压亏缺、根区土壤有效含水率的响应方程。
gc对Rad的响应方程选取了Stewart[11]提出的驱动方程形式:
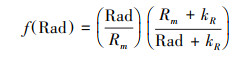
|
(8) |
式中, Rm为最大太阳辐射, 通常设定为1000 W/m2。
gc对VPD的响应方程形式有多种, 常用的指数或双曲线函数形式难以确定气孔导度最大值, 因此选取对数函数形式[26]:
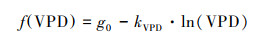
|
(9) |
式中, g0为参比冠层平均气孔导度(VPD=1 kPa时对应的气孔导度)。
土壤水分对植物气孔导度的胁迫作用通常存在阈值[27], 故选取S型函数形式土壤水分胁迫方程[28]:
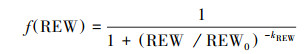
|
(10) |
式中, REW0为1/2最大气孔导度时的根区土壤有效含水量, 即f(REW0)=0.5;土壤水分胁迫阈值点(REWt)定义为f(REWt)=0.95。
根据公式(6)可得, 由气孔导度计算冠层蒸腾计算公式:
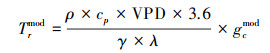
|
(11) |
将观测数据分为数据集A(2015、2016年数据, n=7759)和数据集B(2017、2018年数据, n=10207):利用数据集A分析gc和各环境因子之间的关系并拟合响应方程, 率定模型参数;利用数据集B检验模型, 并使用纳什效率系数(NSE)、均方根误差(RMSE)、平均绝对误差(MAE)三个指标评价模型模拟效果。
NSE常被用来评定模型模拟的精度, 取值范围从负无穷到1, 其值越接近1表明模型拟合度越高, 模拟效果越好[29]。其计算公式如下:
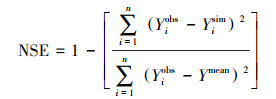
|
(12) |
式中, Yiobs为第i个观测值, Yisim为第i个模拟值, Ymean为观测值均值。
RMSE和MAE能够反应模拟的误差情况, 当RMSE和MAE小于测量数据标准差的一半则可认为模拟效果较好[29]。其计算公式如下:
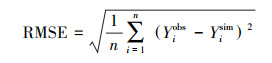
|
(13) |

|
(14) |
式中, Yiobs为第i个观测值, Yisim为第i个模拟值。
2 结果与分析 2.1 冠层平均气孔导度与冠层蒸腾变化特征2015—2018年生长季内(4—10月), 油松日均gc为2.02 mm/s, 日均Tr为1.25 mm/d, 样地油松日均单株耗水量为4.44 kg d-1株-1;生长季总蒸腾耗水量均值为195.47 mm。
Rad和VPD为gc日内变化的主要影响因子, 偏相关系数分别为0.65和-0.41(表 2)。晴天典型日过程显示(图 1), gc和Tr日内呈单峰型变化, 与Rad基本同步, 其日变化可以分为三个阶段:自6:00开始, gc和Tr随Rad的上升迅速升高, 在10:00达到峰值, 此时Tr为0.23 mm/h, gc为7.06 mm/s;此后, Tr相对稳定, 而gc随VPD的增加显著减少;17:00之后, gc和Tr随Rad与VPD迅速下降。
| 环境因子 Environment factors |
太阳辐射 Rad |
水汽压亏缺 VPD |
气温 T |
降水量 P |
根区土壤有效含水率 REW |
| 偏相关系数(小时尺度) Partial coefficient (hourly) |
0.65** | -0.41** | 0.08** | 0.06** | 0.20** |
| 偏相关系数(日尺度) Partial coefficient (daily) |
0.38** | -0.33** | 0.137** | -0.04 | 0.46** |
| Rad:太阳辐射Solar radiation;VPD:水汽压亏缺Vapor pressure deficit;T:气温Air temperature;P:降雨量Rainfall;REW:根区土壤有效含水率Relative extractable water;** P < 0.01, * P < 0.05 | |||||

|
| 图 1 黄土高原油松冠层平均气孔导度、冠层蒸腾及环境因子晴天典型日变化特征(2015年7月26日到30日) Fig. 1 Diurnal variation of mean canopy stomatal conductance and canopy transpiration of Pinus tabuliformis and environmental factors on the Loess Plateau between July 26, 2015 and July 30, 2015 |
REW为gc年内变化的主要控制因子, 偏相关系数为0.46(表 2)。以2015—2018年生长季实测数据为例分析gc年内变化过程, 由图 2可知:gc与REW在晴天同步降低, 这是由于REW随着植被蒸腾耗水持续减少, 植被调节气孔控制水分散失以适应水分胁迫;当REW受降水补给增加后, gc也同步增加。4—6月土壤水分较低时, gc和Tr与REW变化趋势一致, 在5月共同到达生长季最低点。7—10月土壤水分充足时, gc保持稳定, Tr在7月达到峰值(42.67 mm)后随Rad和VPD的下降而减少。

|
| 图 2 黄土高原油松冠层平均气孔导度、冠层蒸腾及环境因子典型(2016) 与多年平均年内变化 Fig. 2 Typical (2016) and multi-year average annual variation of mean canopy stomatal conductance and canopy transpiration of Pinus tabuliformis and environmental factors on the Loess Plateau |
基于实测环境因子与gc拟合响应方程(表 3, 图 3), 在参数率定阶段, NSE=0.81, RMSE=1.14, MAE=0.85。
| 参数 Parameters |
参数描述 Parameter descriptions |
拟合值 Fitted values |
参数 Parameters |
参数描述 Parameter descriptions |
拟合值 Fitted values |
|
| kR | 气孔导度太阳辐射驱动系数 | 298.64 | kVPD | 气孔导度水汽压亏缺驱动系数 | 5.29 | |
| Rm | 最大太阳辐射 | 1000.00 | REW0 | 半气孔导度土壤有效含水率 | 0.30 | |
| g0 | 参比冠层平均气孔导度 | 7.40 | kREW | 气孔导度土壤水分影响系数 | 7.00 |

|
| 图 3 冠层平均气孔导度对环境因子的响应 Fig. 3 The response of hourly mean canopy stomatal conductance to environmental factors |
gc随Rad的升高而增加。在Rad较小时gc对Rad更加敏感, Rad超过300 W/m2后, gc随太阳辐射增加趋于饱和。
gc随VPD的增加而减少。利用对数函数能较好的描述gc与VPD的关系, g0拟合值为7.40 mm/s。
gc对REW的响应存在阈值(REWt= 0.45)。阈值点之前, gc随REW的减少迅速下降, 二者相关性较高, REW0为0.30;在REW达到0.45之后二者相关性较低。
利用数据集B的实测环境因子数据检验gc模拟效果, 结果表明拟合较好(NSE=0.80), 能准确模拟gc日内变化(图 4)。

|
| 图 4 小时冠层平均气孔导度(DOY 174-187)测量值与模拟值比较 Fig. 4 Comparison between measured and simulated hourly mean canopy stomatal conductance (DOY 174-187) gc:冠层平均气孔导度Mean canopy stomatal conductance |
结合Jarvis型冠层气孔导度模型和Penman-Monteith公式构建冠层蒸腾模型, 利用数据集B进行检验。结果表明, 冠层小时与日蒸腾量模拟效果较好, NSE分别为0.78和0.76(图 5)。基于2015—2018年气象和土壤水分数据模拟冠层蒸腾, 求得生长季内林分蒸腾耗水总量均值为197.05 mm, 占同期降雨量的39.11%。

|
| 图 5 黄土高原油松小时、日冠层蒸腾量模拟结果 Fig. 5 Hourly and daily canopy transpiration simulation results of Pinus tabuliformis on the Loess Plateau Tr:冠层蒸腾Canopy transpiration |
油松gc和Tr日内变化主要受Rad和VPD影响(图 1, 表 2)。Rad能调节保卫细胞的离子吸收和有机质积累, 从而改变其渗透压, 控制气孔;另外, Rad也可以促进叶片光合作用, 使细胞间二氧化碳浓度减少, 从而刺激保卫细胞, 影响气孔开闭[30]。这种响应机制可以确保气孔仅在能够进行光合作用时张开, 以提高光合水分利用效率[6]。因此, 油松gc和Tr日内变化的上升(6:00—10:00)和下降(15:00之后)阶段, Rad控制效果明显(图 1)。VPD能从影响气孔和改变水汽扩散速率两方面控制蒸腾[10]。因此在图 1中, 10:00之后, 随VPD升高, 尽管gc开始下降, 但由于水势梯度的增加, Tr仍保持稳定。受气象因子调控, 样地油松gc和Tr在日内呈现单峰曲线。与栓皮栎[31]、侧柏[8]、樟子松[32]等Tr日内呈现双峰型的树种相比, 油松倾向于采用等水调节策略[33], 更能适应长期的干旱胁迫[12]。
油松gc和Tr年内变化主要受土壤水分控制(图 2, 表 2)。土壤水分亏缺会导致植物体内脱落酸浓度升高, 叶片保卫细胞膨压降低, 促使气孔关闭[30]。这种响应机制使得植物蒸腾耗水不会超过土壤可供水量[7], 可以避免由叶片—根系水势差过大导致的木质部栓塞。因此, gc和Tr的年内变化与REW同步(图 2), 与陈胜楠等[15]和Jian等[14]对油松的研究结果一致。在4—5月份, 样地日降雨量均小于5 mm, 难以有效补充土壤水分, 所以gc和Tr随REW在5月共同达到生长季最低点(图 2)。
3.2 油松冠层气孔导度模拟通过模拟油松gc, 能够得到反映油松用水策略的特征参数。Oren等[34]指出, 植被对VPD的敏感度随g0的增加而提高, 且kVPD与g0存在线性关系, 斜率约为0.6。样地油松g0为7.40 mm/s, 与美洲山杨[35]、桉树[36]等树种相近, 高于马占相思[37]、荷木[19]等树种(表 4)。样地油松kVPD与g0的比值为0.71, 与陈胜楠等[15](0.74)对油松的研究结果相近, 均高于0.6(图 6)。因此, 与同等g0的树种相比, 油松kVPD更大。这表明随VPD的升高, 油松gc降低更为迅速, 能够通过调节气孔避免高VPD引起的过度蒸腾, 对环境水分变化较敏感。kREW与REWt分别代表了植被对土壤水分变化的敏感性和土壤水分对油松产生胁迫的关键阈值。样地油松kREW(7.00)与黄土高原地区华北落叶松相似[18], REWt(0.45)与柠条相同[42]。kR代表油松对太阳辐射的敏感性, 与白柏松(257.99)相似[10]。本研究得到的模型参数值符合油松耐旱的基本特征, 能够为评估黄土高原油松冠层气孔导度和蒸腾变化特征提供重要参考。
| 植物种类 Species |
代码 Code |
g0 / (mm/s) |
kVPD/ (mm s-1 ln(kPa)-1) |
来源 Sources |
| 油松Pinus tabuliformis | Pt1 | 7.4 | 5.29 | 本文 |
| Pt2 | 1.03 | 0.77 | 陈胜楠等, 2020[15] | |
| 南欧油松Pinus pinaster | Pp | 3.36 | 2.41 | Granier等[38] |
| 樟子松Pinus sylvestris | Ps | 6.39 | 3.78 | Korner等[34] |
| 马尾松Pinus contorta | Pc | 5.14 | 2.17 | Sandford等[39] |
| 美洲山杨Populus tremuloides | Pt′ | 8.11 | 5.26 | Dang等[35] |
| 桉树Eucalyptus pauciflora | Ep | 7.99 | 4.72 | Korner等[36] |
| 柠檬桉Eucalyptus citriodora | Ec | 1.92 | 1.32 | 胡彦婷等[40] |
| 荷木Schima superba | Ss | 3.85 | 2.04 | 曹庆平等[19] |
| 马占相思Acacia mangium | Am | 2.19 | 1.76 | 赵平等[41] |

|
| 图 6 不同树种冠层气孔导度对水汽压亏缺增高的敏感度(kVPD) 与参比冠层气孔导度(g0) 的关系 Fig. 6 Sensitivity of canopy stomatal conductance (kVPD) of different tree species to vapor pressure deficit in relation to reference canopy stomatal conductance (g0) Pt1—2:油松Pinus tabuliformis;Pp:南欧油松Pinus pinaster;Ps:樟子松Pinus sylvestris;Pc:马尾松Pinus contorta;Pt′:美国山杨Populus tremuloides;Ep:桉树Eucalyptus pauciflora;Ec:柠檬桉Eucalyptus citriodora;Ss:荷木Schima superba;Am:马占相思Acacia mangium |
油松gc和Tr在不同土壤水分条件下对环境因子的响应存在显著差异[33], 因此本研究构建Jarvis型模型时考虑了REW对gc和Tr的影响。然而部分研究指出冠层气孔导度或植物耗水与土壤水分之间关系较弱[14, 43], 在构建Jarvis模型时未考虑REW, 其可能原因是:(1)在较小时间尺度(小时尺度)gc主要受气象因子的影响, 与REW关系较弱[37]; (2)土壤水分充足时REW的限制作用不明显[44];(3)植物根系较深时, 深根吸水对蒸腾贡献更大[45—46], 仅考虑浅层土壤(如表层20 cm)计算的REW与gc关系不明显[17]。本研究开展于黄土高原, 水分是该地区植物生长的主要限制因素[47], 研究时段内REW低于胁迫阈值的时间占比为68%。此外, 样地土层深厚, 油松根系发达[22], 深层土壤吸水对其蒸腾有重要贡献[45—46]。通过对比考虑REW前后gc和Tr的模拟效果(图 7), 可以发现:在土壤水分胁迫发生时, gc和Tr随REW减少持续下降, 而不考虑REW的模拟结果未出明显变化, 对gc和Tr产生了高估。因此, 为准确模拟黄土高原地区人工林的蒸腾耗水, 在构建Jarvis型气孔导度模型时需要考虑REW。

|
| 图 7 根区土壤有效含水率对冠层气孔平均导度和冠层蒸腾模拟的影响 Fig. 7 Effects of relative extractable water on mean canopy stomatal conductance and canopy transpiration (公式一:gc=f(Rad)f(VPD)f(REW);公式二:gc=f(Rad)f(VPD)) |
气温对于gc有显著影响[48], 但是由于VPD与气温通常高度相关[49], 同时考虑VPD与气温对gc的影响不符合Jarvis模型的基本假设, 将会导致gc的低估[50], 降低模拟效果[10]。因此, 本研究构建Jarvis型模型时并未考虑气温的影响。
3.3 油松冠层蒸腾特征研究区油松日均单株耗水量为4.44 kg d-1株-1, 与韩磊[16]计算结果相近, 远低于阔叶树种如橡树(10 kg d-1株-1)[51]和杨树(> 6.4 kg d-1株-1)[52], 也低于同为耐旱树种的樟子松(> 5.31 kg d-1株-1)[32]和刺槐(6.61 kg d-1株-1)[20]。结果表明, 油松单株耗水更少, 更适合黄土高原水分限制区的植被恢复。
4 结论本文基于黄土高原实测油松液流和环境因子数据, 分析了油松gc和Tr的变化特征和影响因素, 以准确模拟黄土高原地区油松冠层气孔导度和蒸腾耗水, 得到以下结论:
(1) 油松gc和Tr存在明显的日内和年内变化, gc日均值为2.02 mm/s, Tr日均值为1.25 mm/d, 日均单株耗水量为4.44 kg d-1株-1;油松耗水量在7月份达到峰值(42.67 mm), 生长季(4—10月)总蒸腾耗水量年均值为195.47 mm。
(2) 油松gc的主要影响因素在不同时间尺度存在差异:小时尺度, Rad为gc的主要驱动因子, 当Rad高于300 W/m2时, 驱动作用减弱, VPD为gc的主要控制因子, 参比冠层平均气孔导度为7.40 mm/s;日尺度, REW为gc的主要限制因子, 当REW低于0.45时限制作用明显。
(3) 结合Penman-Monteith公式和Jarvis型气孔导度模型能够有效模拟油松gc和Tr。
| [1] |
Feng X M, Fu B J, Piao S L, Wang S, Ciais P, Zeng Z Z, Lü Y, Zeng Y, Li Y, Jiang X H, Wu B F. Revegetation in China's Loess Plateau is approaching sustainable water resource limits. Nature Climate Change, 2016, 6(11): 1019-1022. DOI:10.1038/nclimate3092 |
| [2] |
Chen Y P, Wang K B, Lin Y S, Shi W Y, Song Y, He X H. Balancing green and grain trade. Nature Geoscience, 2015, 8(10): 739-741. DOI:10.1038/ngeo2544 |
| [3] |
Li S, Liang W, Fu B J, Lü Y H, Fu S Y, Wang S, Su H M. Vegetation changes in recent large-scale ecological restoration projects and subsequent impact on water resources in China's Loess Plateau. Science of the Total Environment, 2016, 569/570: 1032-1039. DOI:10.1016/j.scitotenv.2016.06.141 |
| [4] |
Jia X X, Shao M A, Zhu Y J, Luo Y. Soil moisture decline due to afforestation across the Loess Plateau, China. Journal of Hydrology, 2017, 546: 113-122. DOI:10.1016/j.jhydrol.2017.01.011 |
| [5] |
Jasechko S, Sharp Z D, Gibson J J, Birks S J, Yi Y, Fawcett P J. Terrestrial water fluxes dominated by transpiration. Nature, 2013, 496(7445): 347-350. DOI:10.1038/nature11983 |
| [6] |
Wong S C, Cowan I R, Farquhar G D. Leaf conductance in relation to assimilation in Eucalyptus pauciflora sieb. ex spreng: influence of irradiance and partial pressure of carbon dioxide. Plant Physiology, 1978, 62(4): 670-674. DOI:10.1104/pp.62.4.670 |
| [7] |
Jackson R B, Sperry J S, Dawson T E. Root water uptake and transport: using physiological processes in global predictions. Trends in Plant Science, 2000, 5(11): 482-488. DOI:10.1016/S1360-1385(00)01766-0 |
| [8] |
刘文娜, 贾剑波, 余新晓, 贾国栋, 侯贵荣. 华北山区侧柏冠层气孔导度特征及其对环境因子的响应. 应用生态学报, 2017, 28(10): 3217-3226. |
| [9] |
高冠龙, 张小由, 常宗强, 鱼腾飞, 赵虹. 植物气孔导度的环境响应模拟及其尺度扩展. 生态学报, 2016, 36(6): 1491-1500. |
| [10] |
Whitley R, Medlyn B, Zeppel M, Macinnis-Ng C, Eamus D. Comparing the Penman-Monteith equation and a modified Jarvis-Stewart model with an artificial neural network to estimate stand-scale transpiration and canopy conductance. Journal of Hydrology, 2009, 373(1/2): 256-266. |
| [11] |
Stewart J B. Modelling surface conductance of pine forest. Agricultural and Forest Meteorology, 1988, 43(1): 19-35. DOI:10.1016/0168-1923(88)90003-2 |
| [12] |
温杰, 陈云明, 唐亚坤, 吴旭, 谢育利, 崔高阳. 黄土丘陵区油松、沙棘生长旺盛期树干液流密度特征及其影响因素. 应用生态学报, 2017, 28(3): 763-771. |
| [13] |
Fang S M, Zhao C Y, Jian S Q. Canopy transpiration of Pinus tabuliformis plantation forest in the Loess Plateau region of China. Environmental Earth Sciences, 2016, 75(5): 1-9. |
| [14] |
Jian S Q, Wu Z N, Hu C H. Estimation of water use of Pinus tabuliformis Carr. in Loess Plateau of Northwest China. Journal of Hydrology and Hydromechanics, 2019, 67(3): 271-279. DOI:10.2478/johh-2019-0013 |
| [15] |
陈胜楠, 孔喆, 陈立欣, 刘清泉, 刘平生, 张志强. 半干旱区城市环境下油松林分蒸腾特征及其影响因子. 生态学报, 2020, 40(4): 1269-1280. |
| [16] |
韩磊. 黄土半干旱区主要造林树种蒸腾耗水及冠层蒸腾模拟研究[D]. 北京: 北京林业大学, 2011.
|
| [17] |
Jiao L, Lu N, Fang W W, Li Z S, Wang J, Jin Z. Determining the independent impact of soil water on forest transpiration: a case study of a black locust plantation in the Loess Plateau, China. Journal of Hydrology, 2019, 572: 671-681. DOI:10.1016/j.jhydrol.2019.03.045 |
| [18] |
Li Z H, Yu P T, Wang Y H, Webb A A, He C, Wang Y B, Yang L L. A model coupling the effects of soil moisture and potential evaporation on the tree transpiration of a semi-arid larch plantation. Ecohydrology, 2017, 10(1): e1764. DOI:10.1002/eco.1764 |
| [19] |
曹庆平, 赵平, 倪广艳, 朱丽薇, 牛俊峰, 曾小平. 华南荷木林冠层气孔导度对水汽压亏缺的响应. 生态学杂志, 2013, 32(7): 1770-1779. |
| [20] |
Ma C K, Luo Y, Shao M G, Li X D, Sun L, Jia X X. Environmental controls on sap flow in black locust forest in Loess Plateau, China. Scientific Reports, 2017, 7: 13160. DOI:10.1038/s41598-017-13532-8 |
| [21] |
Granier A. Evaluation of transpiration in a Douglas-fir stand by means of sap flow measurements. Tree Physiology, 1987, 3(4): 309-320. DOI:10.1093/treephys/3.4.309 |
| [22] |
Qi Y L, Wei W, Li J R, Chen C G, Huang Y Y. Effects of terracing on root distribution of Pinus tabuliformis Carr. forest and soil properties in the Loess Plateau of China. Science of the Total Environment, 2020, 721: 137506. DOI:10.1016/j.scitotenv.2020.137506 |
| [23] |
Daley M J, Phillips N G. Interspecific variation in nighttime transpiration and stomatal conductance in a mixed New England deciduous forest. Tree Physiology, 2006, 26(4): 411-419. DOI:10.1093/treephys/26.4.411 |
| [24] |
Chen Z, Zhang Z Q, Sun G, Chen L X, Xu H, Chen S N. Biophysical controls on nocturnal sap flow in plantation forests in a semi-arid region of Northern China. Agricultural and Forest Meteorology, 2020, 284: 107904. DOI:10.1016/j.agrformet.2020.107904 |
| [25] |
Monteith J, Unsworth M. Principles of Environmental Physics: Plants, Animals, and the Atmosphere[M]. Academic Press, 2013.
|
| [26] |
Oren R, Sperry J S, Ewers B E, Pataki D E, Phillips N, Megonigal J P. Sensitivity of mean canopy stomatal conductance to vapor pressure deficit in a flooded Taxodium distichum L. forest: hydraulic and non-hydraulic effects. Oecologia, 2001, 126(1): 21-29. DOI:10.1007/s004420000497 |
| [27] |
Leuning R. A critical appraisal of a combined stomatal-photosynthesis model for C3 plants. Plant, Cell and Environment, 1995, 18(4): 339-355. DOI:10.1111/j.1365-3040.1995.tb00370.x |
| [28] |
孙林, 管伟, 王彦辉, 徐丽宏, 熊伟. 华北落叶松冠层平均气孔导度模拟及其对环境因子的响应. 生态学杂志, 2011, 30(10): 2122-2128. |
| [29] |
Moriasi D N, Arnold J G, Liew M W V, Bingner R L, Harmel R D, Veith T L. Model evaluation guidelines for systematic quantification of accuracy in watershed simulations. Transactions of the ASABE, 2007, 50(3): 885-900. DOI:10.13031/2013.23153 |
| [30] |
Lambers H, Chapin F S Ⅲ, Pons T L. Plant Physiological Ecology. New York, NY: Springer New York, 2008.
|
| [31] |
聂立水, 李吉跃, 翟洪波. 油松、栓皮栎树干液流速率比较. 生态学报, 2005, 25(8): 1934-1940. DOI:10.3321/j.issn:1000-0933.2005.08.015 |
| [32] |
陈彪, 陈立欣, 刘清泉, 刘平生, 张志强. 半干旱地区城市环境下樟子松蒸腾特征及其对环境因子的响应. 生态学报, 2015, 35(15): 5076-5084. |
| [33] |
Jia G D, Liu Z Q, Chen L X, Yu X X. Distinguish water utilization strategies of trees growing on earth-rocky mountainous area with transpiration and water isotopes. Ecology and Evolution, 2017, 7(24): 10640-10651. DOI:10.1002/ece3.3584 |
| [34] |
Oren R, Sperry J S, Katul G G, Pataki D E, Ewers B E, Phillips N, Schäfer K V R. Survey and synthesis of intra-and interspecific variation in stomatal sensitivity to vapour pressure deficit. Plant, Cell and Environment, 1999, 22(12): 1515-1526. DOI:10.1046/j.1365-3040.1999.00513.x |
| [35] |
Dang Q L, Margolis H A, Coyea M R, Sy M, Collatz G J. Regulation of branch-level gas exchange of boreal trees: roles of shoot water potential and vapor pressure difference. Tree Physiology, 1997, 17(8/9): 521-535. |
| [36] |
K rner C, Cochrane P M. Stomatal responses and water relations of Eucalyptus pauciflora in summer along an elevational gradient. Oecologia, 1985, 66(3): 443-455. DOI:10.1007/BF00378313 |
| [37] |
赵平, 饶兴权, 马玲, 蔡锡安, 曾小平. 马占相思林冠层气孔导度对环境驱动因子的响应. 应用生态学报, 2006, 17(7): 1149-1156. |
| [38] |
Granier A, Loustau D. Measuring and modelling the transpiration of a maritime pine canopy from sap-flow data. Agricultural and Forest Meteorology, 1994, 71(1/2): 61-81. |
| [39] |
Sandford A P, Jarvis P G. Stomatal responses to humidity in selected conifers. Tree Physiology, 1986, 2(1_2_3): 89-103. |
| [40] |
胡彦婷, 赵平, 牛俊峰, 孙振伟, 朱丽薇. 三种植被恢复树种的冠层气孔导度特征及其对环境因子的敏感性. 应用生态学报, 2015, 26(9): 2623-2631. |
| [41] |
赵平, 刘惠, 孙谷畴. 4种植物气孔对水汽压亏缺敏感度的种间差异. 中山大学学报: 自然科学版, 2007, 46(4): 63-68. |
| [42] |
She D L, Xia Y Q, Shao M G, Peng S Z, Yu S G. Transpiration and canopy conductance of Caragana korshinskii trees in response to soil moisture in sand land of China. Agroforestry Systems, 2013, 87(3): 667-678. |
| [43] |
Tie Q, Hu H C, Tian F Q, Guan H D, Lin H. Environmental and physiological controls on sap flow in a subhumid mountainous catchment in North China. Agricultural and Forest Meteorology, 2017, 240/241: 46-57. |
| [44] |
Bernier P Y, Bréda N, Granier A, Raulier F, Mathieu F. Validation of a canopy gas exchange model and derivation of a soil water modifier for transpiration for sugar maple (Acer saccharum Marsh.) using sap flow density measurements. Forest Ecology and Management. |
| [45] |
Zhao Y L, Wang Y Q, He M N, Tong Y P, Zhou J X, Guo X Y, Liu J Z, Zhang X C. Transference of Robinia pseudoacacia water-use patterns from deep to shallow soil layers during the transition period between the dry and rainy seasons in a water-limited region. Forest Ecology and Management, 2020, 457: 117727. |
| [46] |
Liu Z H, Jia G D, Yu X X, Lu W W, Sun L B, Wang Y S, Baheti Z. Morphological trait as a determining factor for Populus simonii Carr. to survive from drought in semi-arid region. Agricultural Water Management. |
| [47] |
张涵丹, 卫伟, 陈利顶, 于洋, 杨磊, 贾福岩. 典型黄土区油松树干液流变化特征分析. 环境科学, 2015, 36(1): 349-356. |
| [48] |
韩磊, 何俊, 齐拓野, 田佳, 孙兆军, 展秀丽. 宁夏河东沙区侧柏冠层气孔导度对环境因子的响应及其模拟. 生态学杂志, 2018, 37(9): 2862-2868. |
| [49] |
罗紫东, 关华德, 章新平, 刘娜, 张赐成, 王婷. 桂花树冠层气孔导度模型的优化及其参数分析. 生态学报, 2016, 36(13): 3995-4005. |
| [50] |
Wang H L, Guan H D, Liu N, Soulsby C, Tetzlaff D, Zhang X P. Improving the Jarvis-type model with modified temperature and radiation functions for sap flow simulations. Journal of Hydrology, 2020, 587: 124981. |
| [51] |
Wullschleger S D, Hanson P J, Tschaplinski T J. Whole-plant water flux in understory red maple exposed to altered precipitation regimes. Tree Physiology, 1998, 18(2): 71-79. |
| [52] |
莫康乐, 陈立欣, 周洁, 方显瑞, 康满春, 张志强. 永定河沿河沙地杨树人工林蒸腾耗水特征及其环境响应. 生态学报, 2014, 34(20): 5812-5822. |