 2022, Vol. 42
2022, Vol. 42文章信息
- 黄一鑫, 程艳霞
- HUANG Yixin, CHENG Yanxia
- 森林光环境对4种乔木幼树光合和光谱反射特性的影响
- Photosynthetic characteristics and spectral reflectance characteristics of four natural tree saplings under forest light environment
- 生态学报. 2022, 42(22): 9121-9129
- Acta Ecologica Sinica. 2022, 42(22): 9121-9129
- http://dx.doi.org/10.5846/stxb202105171289
-
文章历史
- 收稿日期: 2021-05-17
- 网络出版日期: 2022-07-12





光不仅是影响植物生长、发育的主要生态因子, 也是影响植物存活和分布的重要环境因素[1—2]。森林光环境在空间和时间上均存在异质性, 且与林下幼树的存活、生长和分布等密切相关[3—4]。林窗作为森林生态系统得以长期维持的重要驱动力之一[5], 对于维持群落的结构、功能以及更新起着重要的作用[6]。林窗和林缘的形成导致光照因子的急剧变化, 形成不同的光环境[7]。除了空间上的变化, 随着演替的进程, 郁闭的林冠遮挡了大部分入射的太阳光, 使得林下幼树接收到的光照减少[8], 而幼树作为维持森林生物多样性和群落结构更新重要的环节[9], 为了适应弱光环境, 会通过改变其光合生理特性和生理特性来适应环境[10]。有研究表明, 弱光环境下植物通过降低比叶重(leaf mass per area, LMA), 增加叶绿素含量来提高对低光的截获[10—14], 同时升高表观量子效率(apparent quantum yield, AQY), 降低光补偿点(light compensation point, LCP), 光饱和点(light saturation point, LSP), 最大净光合速率(maximum net photosynthetic rate, Pnmax)和暗呼吸效率(dark respiration rate, Rd), 以提高叶片对光能的利用效率并减小呼吸作用的能量消耗, 以保证最大限度地进行光合碳积累, 维持弱光下的碳平衡[3, 13, 15—16]。但也有研究发现弱光条件下植物叶片LCP增大, AQY、Rd、Pnmax、LSP无显著变化[13, 17—18]。说明不同植物以不同的方式来响应其生存的异质光环境, 不同植物叶片光合特性对不同光环境的响应不尽相同[19]。目前关于植物对不同光环境响应的研究多为控制试验条件下培育出幼苗、幼树移栽到不同光环境下进行再生长或人为遮阴模拟自然条件下不同光环境[3, 14, 17—18, 20]。控制变量试验虽能模拟出自然条件下不同的光环境, 但无法完全还原自然条件下植株对不同环境的响应。在野外除了光照的总量在不同生境中发生变化, 由于林冠层分布模式不同, 林窗和林缘往往具有不同的受光模式[13], 同时野外环境中还混杂水分、土壤和种间竞争等非生物和生物因子, 使得野外植物在长期生长中光合特性的表现与控制条件下出现不一致现象[21]。因此, 研究植物在不同天然森林光环境下的光合生理特性, 可以在一定程度上解释植物演替的一些机制问题, 有助于了解天然林组成树种的生态适应性, 为植被恢复、天然林保护等提供理论依据。
叶片作为植物进行光合作用的主要器官, 通过改变其外在或内在的特征, 以适应不同光环境的变化[4, 22—23], 因此叶片特征是植物响应光环境能力的一个较好表征[24]。近年来, 植物反射光谱分析技术快速发展, 并在植物生理生态研究中广泛应用[25]。相关研究表明, 叶片的光谱反射率曲线和光谱反射指数与植物生理特性联系紧密, 能较好评估植物生长状况、光合作用、受胁迫等情况[26—27]。以往研究主要对不同光环境下植株光合特性的探究, 少有将光合特性和光谱特性结合的探究, 本文以吉林蛟河阔叶红松林为研究地, 通过对林窗、林缘、林下3种不同自然光环境下的常见树种幼树光合特性和光谱特性的探究, 以期从生理生态角度探讨它们对环境的适应能力和响应机制, 为深入研究阔叶红松林林木更新, 群落动态和演替提供参考。
1 研究区概况和研究方法 1.1 研究区概况研究地点位于吉林省林业实验区国有林保护中心的阔叶红松林动态监测样地, 坐标为43°57′N, 127°45′E, 平均海拔459m。该区域属大陆性季风气候, 年平均降水量为695.9mm, 降水集中在7、8月份, 年平均气温3.8℃, 最热月7月平均气温21.7℃, 最冷月1月平均气温是-18.6℃。林下土壤为山地暗棕色森林土, 土层厚20—80cm。研究区域植被属于长白山植物区系, 阔叶红松林是主要的森林类型, 样地内乔木树种主要有红松(Pinus koraiensis)、蒙古栎(Quercus mongolica)、紫椴(Tiliaamurensis)、春榆(Ulmus japonica)、色木槭(Acer mono)、白牛槭(Acer mandshuricum)、水曲柳(Fraxinus mandshurica)、拧筋槭(Acer triflorum Komarov)等[28]。
1.2 研究方法试验于2020年8月树木旺盛生长期进行。在吉林蛟河阔叶红松林动态监测样地内, 经踏选, 在林下、林缘、大林窗环境中选择3块样地。各样地随机选取1.8m左右天然更新、生长状况良好、大小相对一致的拧筋槭、白牛槭、色木槭和红松幼树各3株。各样地光环境情况见表 1。
| 光环境 Forest light environment |
相对光合有效辐射 Relative photosynthetically active radiation/% |
叶面积指数 Leaf area index |
| 林下Understory | 7.58±1.90b | 4.17±0.12a |
| 林缘Forest edge | 8.48±2.48b | 3.11±0.11b |
| 林窗Forest gap | 13.89±1.64a | 2.45±0.16c |
| 光环境数据为均值±标准误差, 不同小写字母表示样地间的光环境差异(P<0.05) | ||
于天气晴朗的08:30—11:00进行光响应曲线的测定, 所有叶片均进行离体测定。用高枝剪在每棵样树的中上部剪下二级枝后, 立刻将枝条断口插入清水中进行水下剪枝, 以维持导管内部水势, 防止叶片失水失活。每棵样树选择3片成熟健康叶片进行光响应曲线测定[29]。采用Li_6800红蓝光源提供的标准光强, 设定空气流速为0.5L/min, 叶片温度25℃, 叶室中相对湿度50%, CO2浓度400μmol/mol。诱导光强约2000μmol m-2 s-1, 诱导时间30min。测定时光强由强到弱依次设定光量子通量密度(PFD)为;2000、1500、1200、1000、800、600、400、200、150、100、50、20、0μmol m-2 s-1(红松);1200、1000、750、500、250、150、100、75、50、25、10、5、0μmol m-2 s-1(其他树种), 测定时每一光强下最小等待时间120s, 最大等待时间240s。数据导出后, 采用非直角双曲线模型[30]对测量得到的光响应数据进行拟合, 模型形式如下:
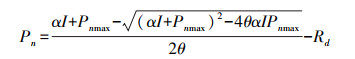
|
式中, α是光响应曲线的初始斜率;θ为曲线的曲率;Pn为净光合速率, μmol m-2 s-1;I为入射光强, μmol m-2 s-1;应用此模型拟合各树种光响应曲线, 得出表观量子率(AQY)、暗呼吸速率(Rd)、最大净光合速率(Pnmax)、光补偿点(LCP)、光饱和点(LSP)。
1.2.2 光谱数据采集与处理在选定的目标树采下新鲜枝条, 选取3片健康成熟的叶片, 用英国PPSYSTEM公司的Unispec-SC光谱分析仪进行测量。波长范围为310—1130 nm, 卤光灯光强设为100%, 扫描波长间隔为3.3nm, 设置整合时间4ms, 每片叶选取3个点进行重复测量, 每个点重复扫描3次[29]。利用Multispec 5.1软件对光谱仪测得的数据进行处理, 取3次重复的平均值, 根据相应的计算公式得到不同光环境下每棵目标树叶片的叶绿素归一化指数(chlorophyll normalized index, Chl NDI)、光化学反射指数(photochemical reflectance index, PRI)、结构不敏感植被光谱指数(structure-insensitive pigment index, SIPI)和叶片含水量指数(water index, WI), 公式中R值代表叶片对不同波长的光的反射率, 数字代表波长。

|
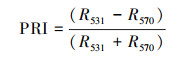
|
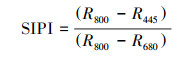
|
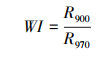
|
采用IBM SPSS Statistics 25.0软件对试验数据进行统计分析, 采用单因素方差分析(one-way ANOVA)和最小显著差异法(LSD)比较不同数据组间的差异(α=0.05)。利用Origin2018c进行图表绘制。图表中数据为平均值±标准差。
2 研究结果 2.1 不同光环境下各树种光响应曲线比较由图 1所示, 在3种光照条件下, 4种常见树种净光合速率(net photosynthetic rate, Pn)随光合有效辐射(photosynthetically active radiation, PAR)增大的变化趋势一致, 当光照强度在0—200μmol m-2 s-1时, 各树种幼树Pn几乎呈线性增长, 随着光照强度加强, Pn的增长速度减缓, 当光照强度达到光饱和点时Pn的增长趋于平缓。除白牛槭外, 林窗内其余树种Pn增幅大于林缘和林下幼树Pn增幅, 林缘和林下环境幼树Pn增幅差异较小;3种光环境下白牛槭幼树Pn增幅差异不大。

|
| 图 1 不同光环境下各树种光响应曲线 Fig. 1 Light response curves of tree species under different light environments |
根据模型回归结果得到3种光环境下4种树种的光响应参数。从表 2可以看出4种植物的光响应参数随着光强的变化呈现不同的趋势, 随着光照强度的降低, 4种常见树种LSP、LCP和Pnmax存在不同程度的减少, 其中林窗内白牛槭、色木槭和红松幼树LSP显著高于林下幼树(P<0.05), 林窗内拧筋槭、白牛槭和色木槭LCP显著高于林下幼树, 林窗内拧筋槭和红松幼树Pmax显著高于林下幼树, 林窗内色木槭幼树Pnmax显著高于林缘和林下(P<0.05)。不同环境下白牛槭和色木槭AQY并没有显著差异, 而林下红松幼树AQY显著高于林缘和林窗幼树(P<0.05), 白牛槭和色木槭Rd随着光照强度降低呈现减小趋势, 但差异未达到显著水平, 林下红松幼树Rd显著大于林缘幼树。
| 树种 Tree species |
生境 Habitat |
光响应参数Light response parameter | ||||
| LSP/ (μmol m-2 s-1) | LCP/ (μmol m-2 s-1) | AQY/ (μmol m-2 s-1) | Rd/ (μmol m-2 s-1) | Pnmax/ (μmol m-2 s-1) | ||
| 拧筋槭 | 林窗 | 389.70±131.75Ba | 13.80±4.58Ba | 0.042±0.011Aa | 0.56±0.20Ba | 6.06±1.79Ba |
| A. Komarov | 林缘 | 393.60±83.58Ba | 9.94±1.66Bb | 0.025±0.011Ab | 0.23±0.14Cb | 5.00±1.12Bab |
| 林下 | 304.11±123.02Ba | 8.16±2.74Bb | 0.046±0.016Aa | 0.47±0.17Ba | 4.50±1.23Bb | |
| 白牛槭 | 林窗 | 353.20±86.35Ba | 14.40±5.03Ba | 0.041±0.020Aa | 0.53±0.23Ba | 4.87±1.35Ba |
| A.mandshuricum | 林缘 | 337.03±47.09Ba | 12.00±2.94Bab | 0.039±0.016Aa | 0.52±0.19Ba | 4.58±1.29Ba |
| 林下 | 259.68±67.98Bb | 8.16±2.82Bb | 0.041±0.010ABa | 0.35±0.15Ba | 4.13±1.46Ba | |
| 色木槭 | 林窗 | 428.91±63.87Ba | 14.00±6.15Ba | 0.030±0.010Ba | 0.42±0.18Ba | 4.67±1.01Ba |
| A.mono | 林缘 | 285.60±98.70Bb | 18.91±9.27Ba | 0.030±0.020Aa | 0.49±0.16Ba | 3.27±0.80Bb |
| 林下 | 307.50±66.28Bb | 11.10±2.85Bb | 0.030±0.010Ba | 0.36±0.15Ba | 3.71±0.56Bb | |
| 红松 | 林窗 | 1118.67±332.94Aa | 37.67±11.24Aa | 0.029±0.008Bb | 0.98±0.37Aab | 12.09±2.14Aa |
| P.koraiensis | 林缘 | 960.33±484.60Aab | 32.53±19.70Aa | 0.027±0.010Ab | 0.76±0.29Ab | 9.74±3.56Ab |
| 林下 | 730.18±307.24Ab | 34.55±17.55Aa | 0.035±0.008ABa | 1.08±0.52Aa | 9.02±2.32Ab | |
| LSP:光饱和点Light saturation point;LCP:光补偿点Light compensation point;AQY:表观量子效率Apparent quantum yield;Rd:暗呼吸速率Dark respiratory rate;Pnmax:最大净光合速率Maximum net photosynthetic rate;数据为平均值±标准误差, 不同小写字母表示不同光环境之间的差异显著性(P < 0.005), 不同大写字母表示不同树种之间的差异显著性(P < 0.005) | ||||||
叶绿素归一化指数(Chl NDI)与叶绿素的含量有很好的正相关关系, Chl NDI值越高表示叶绿素含量越高[6, 31], 如表 3所示, 不同光环境下拧筋槭、色木槭、红松幼树的Chl NDI值随环境光强的增大而上升, 其中林窗拧筋槭、色木槭和红松幼树的Chl NDI值显著低于林下幼树(P<0.05), 表明不同光环境中林下拧筋槭、色木槭和红松含有更多的叶绿素。而林窗、林缘白牛槭Chl NDI值显著大于林下(P<0.05), 预示着光照更好环境下的白牛槭具有更高的叶绿素含量。PRI与叶黄素循环色素含量及光合辐射利用效率有关, 当PRI高时, 光合速率就高[32]。各树种幼树PRI随着环境光强减小均呈现下降趋势, 其中拧筋槭PRI在不同光环境间差异达到显著水平(P<0.05), 这表明光照条件较差环境下的植物具有更高的光合速率。结构不敏感植被光谱指数(SIPI)是与类胡萝卜素的含量高度相关且与叶片结构无关的指数, 反映出类胡萝卜素与叶绿素含量的比值以及树种受到胁迫的状况[33—34], 林缘内拧筋槭、色木槭和红松幼树SIPI值低于林窗和林下幼树, 其中拧筋槭和色木槭幼树差异达到显著(P<0.05), 林窗白牛槭幼树SIPI值显著高于林缘、林下幼树。WI是由Penuelas等人建立, 与植物水分浓度有很高的相关性, WI值越大表示植物水分浓度越高, 不同光环境下各树种幼树的WI值呈现林窗<林缘<林下的趋势, 其中3种槭属幼树WI值在不同光环境间存在显著性差异(P<0.05), 这表明环境光照强度越低, 植物水分浓度越高。
| 树种 Tree species |
生境 Habitat |
光谱反射指数Spectral reflectance index | |||
| Chl NDI | PRI | SIPI | WI | ||
| 拧筋槭 | 林窗 | 0.388±0.030Cb | 0.059±0.010Dc | 0.698±0.029Ba | 0.973±0.010Ac |
| A.Komarov | 林缘 | 0.418±0.058Cb | 0.073±0.015Cb | 0.627±0.048Cb | 0.979±0.005Ab |
| 林下 | 0.467±0.019Aa | 0.087±0.007Ba | 0.712±0.016Ba | 0.988±0.003Aa | |
| 白牛槭 | 林窗 | 0.485±0.043Aa | 0.082±0.011Ba | 0.761±0.017Aa | 0.984±0.003Ac |
| A.mandshuricum | 林缘 | 0.488±0.043Aa | 0.082±0.012Ba | 0.721±0.012Ab | 0.986±0.003Ab |
| 林下 | 0.449±0.045Bb | 0.083±0.016Ba | 0.717±0.017Bb | 0.992±0.002Aa | |
| 色木槭 | 林窗 | 0.414±0.020Bb | 0.072±0.010Cb | 0.746±0.008Aa | 0.926±0.008Ac |
| A.mono | 林缘 | 0.470±0.022Ba | 0.088±0.012Ba | 0.709±0.014Bb | 0.982±0.005Ab |
| 林下 | 0.479±0.042Aa | 0.091±0.010Ba | 0.744±0.009Aa | 0.989±0.002Aa | |
| 红松 | 林窗 | 0.120±0.042Db | 0.121±0.023Aa | -0.355±0.151Ca | 0.816±0.057Ba |
| P.koraiensis | 林缘 | 0.127±0.070Db | 0.120±0.022Aa | -0.392±0.212Da | 0.827±0.065Ba |
| 林下 | 0.153±0.030Ca | 0.122±0.022Aa | -0.326±0.088Ca | 0.837±0.038Ba | |
| Chl NDI:叶绿素归一化指数, chlorophyll normalized index;PRI:光化学反射指数, photochemical reflectance index;SIPI:结构不敏感植被光谱指数, structure-insensitive pigment index;WI:叶片含水量指数, water index;数据为平均值±标准误差, 不同小写字母表示不同光环境之间的差异显著性(P < 0.005), 不同大写字母表示不同树种之间的差异显著性(P < 0.005) | |||||
如图 2所示, 林窗内各槭属树种间光合参数均没有显著差异, 红松幼树各光合参数显著高于槭属树种幼树。多重比较分析表明, 林窗内各树种的Chl NDI值表现为白牛槭>色木槭>拧筋槭>红松, 各树种间指数存在显著差异。而林窗内各树种的PRI值同样差异显著, 表现为红松>白牛槭>色木槭>拧筋槭。白牛槭和色木槭的SIPI值显著高于其他树种, 红松的SIPI值最低。林窗内各阔叶树种间的WI值并没有显著差异, 针叶树种(红松)的WI值显著低于其他树种。

|
| 图 2 林窗内各树种光谱反射指数 Fig. 2 Spectral reflectance index of tree species in forest gap 不同大写字母表示不同树种之间的差异显著性(P < 0.005) |
不同光环境会影响植物生理生态特性, 尤其是光合作用[22, 35]。植物的光合适应性是指在低光条件下光捕获最大化和在高光条件下光合最优化的能力表现, 其中LCP和LSP代表植物光照强度的利用范围, 分别反映植物叶片对弱光和强光的利用能力[2—3]。各树种幼树叶片的LCP和LSP随着生长环境光强的下降而减小, 对不同光环境具有一定的可塑性, 林下各树种幼树LSP显著低于林窗内幼树, 说明遮阴条件下幼树光合能力随着光照的增强更易达到最大效率, 林下3种槭属幼树LCP显著低于林窗内幼树, 说明3种槭属幼树可通过降低光补偿点来适应光辐射强度低的环境, 从而更好地利用弱光。AQY代表叶片对弱光的利用能力, 其值越大说明利用弱光的能力越强[20—21]。本研究中红松的AQY随着环境光强的下降而显著增大, 说明红松幼树对林下低光环境有较强的适应能力, 具有一定的耐阴性[36], Rd反映了植物在黑暗条件下的呼吸速率, 相关研究表明弱光环境下植物通过降低Rd来保持碳平衡[16, 37]。但也有部分研究表示植物Rd并不受光环境的影响, 植物可能通过其他非光合特性的改变而非光合特性的改变来适应光环境的变化[3, 38]。本研究中不同树种幼树Rd随光环境的变化并没有出现明显的变化规律, 各环境间差异较小, 与杨莹等对化香幼苗的研究相似[39]。Pnmax作为反映植物光合潜力的重要指标, 目前相关研究对于Pnmax随光强的变化规律并不统一, 有的研究认为植物通过提高光合速率来利用强光, 防止光组织被破坏[40], 如王振兴等对闽楠的研究发现, 闽楠幼树Pnmax随光环境的提高而提高[3], 而唐星林等对闽楠幼苗的研究发现Pnmax随着遮光率的增大而增大, 遮阴条件下闽楠叶片有较大的光合能力[41], 王暾等对四种天然灌木的研究也得到相似的结果[21]。这说明不同植物对遮阴的适应策略不同, 而且不同年龄植物的需光特性也可能存在差异。本研究中林下各树种Pnmax均小于林窗内幼树, 除白牛槭外其他树种均差异显著, 说明林下幼树具有较低的光合能力, 这可能是树种为了适应林下弱光环境的一种表现。
相对于传统化学分析法, 通过光谱分析法测定色素不仅准确、无损且高效, 还可以同时分析出多种色素含量、水分含量以及光合等生理状况[25]。叶绿素是叶片进行光合作用的重要色素, 叶片中叶绿素含量的变化可以反映出植物对环境因子的响应[41]。本研究中除白牛槭外, 其他树种幼树叶片绿素含量随环境光强的减少而升高, 说明较低的光照强度可以促进植物叶绿素的合成, 植物通过提高对光能的利用效率来更好的适应低光照环境[21, 41]。本研究还发现林下树种的PRI值普遍高于光条件更好环境下树种, 说明林下幼树光合速率大于其他光环境, 出现这一现象可能是由于林窗、林缘环境由于受到较强的太阳辐射, 水分挥发使地面更为干燥, 加之温度的上升致使植物的呼吸作用加快[32], 此外相关研究表明叶片含水量的变化对叶片净光合速率的影响显著[42]。本试验结果显示各树种植物叶片WI值在不同光环境之间均存在显著差异, 光照更强环境下植物叶片的WI值显著低于光环境较差的植物叶片, 说明林下各树种叶片水分浓度、水势显著高于光照条件较好的环境。各树种叶片水分的这种响应可能跟弱光环境下植物蒸腾作用的减弱有关, 此外叶温的降低也可使得植物叶片水分耗散减小[11]。这种响应机制的另一种解释是, 随着遮阴加重, 土壤含水量也随之增加, 使得植物根吸水效率提高[43], 相关研究表明幼树叶水势与其土壤含水率之间有互为指示的关系[44]。由于本试验并没有对不同光环境下土壤含水率进行测定, 所以此响应的具体原因有待进一步验证。4种树种幼树SIPI值在不同光环境下表现出林缘幼树小于林窗和林下, 其中3种槭属差异达到显著水平, 说明林缘下幼树受到相对较小的环境胁迫[32], 相对于林下低光环境, 林缘更好的光照环境有利于树木的光合作用, 使得树木受到胁迫相对较小, 此外林缘环境树木的更新不及林窗环境, 树种间的竞争较林窗环境也更小。红松幼树各光谱反射指数在不同光环境间并没有显著差异, 说明红松幼树生长适应范围较广。
在林窗中, 红松具有最高的LCP、LSP、Pnmax、Rd和PRI, 说明红松具有较高的光合潜力, 更加适应高光生长环境[40], 是典型的演替前期阳生植物, 也是作为阔叶红松林重要建群种树种的重要原因。林窗内红松叶片叶绿素含量最少, 但光合速率最高, 这可能与红松叶片结构有关, 与刘伟国等人[32]的研究结果相似。林窗内白牛槭Chl NDI在所有树种中最高, 同时PRI值在3种槭属树种中也最高, 说明白牛槭具有较高的光合速率, 所以白牛槭应该在林窗内适应性较好、分布较多, 这与事实相符。林窗内白牛槭的SIPI值也最大, 正好印证林窗内白牛槭适应性较好, 分布较多, 相互竞争也较大, 从而面临着更大的环境胁迫[32]。林窗内拧筋槭Chl NDI、PRI值显著低于其他槭属树种, 表现为弱阳性特点, 说明拧筋槭对林窗环境的适应性不如其他3种乔木。
4 结论本研究从植物生理生态的角度对吉林蛟河阔叶红松林不同森林光环境下4种常见树种的生理特性进行了探究。研究发现4种乔木幼树通过降低Pnmax、LSP、LCP的光合策略来适应林下低光环境, 低光环境下拧筋槭和色木槭幼树通过增加叶片叶绿素含量以增强弱光适应性, 且拥有相对更高的光合速率。红松幼树具有一定耐阴性, 在不同光环境下光合和光谱特性差异不显著, 说明其在不同光环境下适应性较好, 也是红松能成为阔叶红松林重要建群种的重要原因。林窗环境中红松具有最大的光合潜力, 适应林窗高光环境, 此外白牛槭也表现出较好的适应性, 说明红松和白牛槭能在林窗环境内较好更新并逐步占据林冠层。
| [1] |
Gspaltl M, Bauerle W, Binkley D, Sterba H. Leaf area and light use efficiency patterns of Norway spruce under different thinning regimes and age classes. Forest Ecology and Management, 2013, 288: 49-59. DOI:10.1016/j.foreco.2011.11.044 |
| [2] |
罗光宇, 陈超, 李月灵, 金则新. 光照强度对濒危植物长序榆光合特性的影响. 生态学杂志, 2021, 40(4): 980-988. DOI:10.13292/j.1000-4890.202104.013 |
| [3] |
王振兴, 朱锦懋, 王健, 汪滢, 卢钰茜, 郑群瑞. 闽楠幼树光合特性及生物量分配对光环境的响应. 生态学报, 2012, 32(12): 3841-3848. |
| [4] |
程晶, 刘济明, 王灯, 王姝, 李丽霞, 陈敬忠. 喀斯特特有植物罗甸小米核桃幼苗对光照强度的可塑性响应. 应用与环境生物学报, 2021, 27(1): 23-30. DOI:10.19675/j.cnki.1006-687x.2020.09012 |
| [5] |
Silbernagel J, Moeur M. Modeling canopy openness and understory gap patterns based on image analysis and mapped tree data. Forest Ecology and Management, 2001, 149(1/2/3): 217-233. |
| [6] |
Chazdon R L. Sunflecks and their importance to forest understorey plants. Advances in Ecological Research, 1988, 18: 1-63. |
| [7] |
Leakey A D B, Press M C, Scholes J D. Patterns of dynamic irradiance affect the photosynthetic capacity and growth of dipterocarp tree seedlings. Oecologia, 2003, 135(2): 184-193. DOI:10.1007/s00442-003-1178-7 |
| [8] |
Parelle J, Roudaut J P, Ducrey M. Light acclimation and photosynthetic response of beech (Fagus sylvaticaL.) saplings under artificial shading or natural Mediterranean conditions. Annals of Forest Science, 2006, 63(3): 257-266. DOI:10.1051/forest:2006004 |
| [9] |
刘福德, 王中生, 张明, 王文进, 安树青, 郑建伟, 杨文杰, 张世挺. 海南岛热带山地雨林幼苗幼树光合与叶氮、叶磷及比叶面积的关系. 生态学报, 2007, 27(11): 4651-4661. DOI:10.3321/j.issn:1000-0933.2007.11.033 |
| [10] |
招礼军, 权佳惠, 朱栗琼, 黄曈, 金赟. 不同生境下濒危植物膝柄木幼树的生态适应性. 广西植物, 2021, 1-11. |
| [11] |
马天光, 李向义, 林丽莎, 李磊, 李世民, 王波. 遮阴对骆驼刺叶性状和水分生理的影响. 生态学报, 2018, 38(23): 8466-8474. |
| [12] |
张云, 夏国华, 马凯, 李根有, 代英超, 严彩霞. 遮阴对堇叶紫金牛光合特性和叶绿素荧光参数的影响. 应用生态学报, 2014, 25(7): 1940-1948. |
| [13] |
杜宁, 张秀茹, 王炜, 陈华, 谭向峰, 王仁卿, 郭卫华. 荆条叶性状对野外不同光环境的表型可塑性. 生态学报, 2011, 31(20): 6049-6059. |
| [14] |
陈斌, 刘筱玮, 贾琳, 杨扬, 何淼. 光强对4种鸭跖草科植物生长和光合特性的影响. 生态学报, 2022(04): 1-12. |
| [15] |
蔡建国, 韦孟琪, 章毅, 魏云龙. 遮阴对绣球光合特性和叶绿素荧光参数的影响. 植物生态学报, 2017, 41(5): 570-576. |
| [16] |
双升普, 张金燕, 寸竹, 武洪敏, 孟珍贵, 李龙根, 陈军文. 光照强度驱动典型阴生植物三七的生理生态响应特征. 生态学报, 2022(9): 1-17. |
| [17] |
刘柿良, 马明东, 潘远智, 魏刘利, 何成相, 杨开茂. 不同光环境对桤木幼苗生长和光合特性的影响. 应用生态学报, 2013, 24(2): 351-358. |
| [18] |
王凯, 朱教君, 于立忠, 孙一荣, 张金鑫. 光环境对胡桃楸幼苗生长与光合作用的影响. 应用生态学报, 2010, 21(4): 821-826. |
| [19] |
Rabelo G R, Vitória  P, da Silva M V A, Cruz R A, Pinho E I B, Ribeiro D R, Freitas A V, Cunha M. Structural and ecophysiological adaptations to forest gaps. Trees, 2013, 27(1): 259-272. |
| [20] |
尚三娟, 王义婧, 王楠, 杨吉兰, 徐胜, 何兴元, 陈玮. 光照强度对紫斑牡丹生理及生长特性的影响. 生态学杂志, 2020, 39(9): 2963-2973. |
| [21] |
王暾, 郭晋平, 刘宁, 张芸香. 森林光环境对4种天然灌木的光合作用和形态的影响. 林业科学, 2011, 47(6): 56-63. |
| [22] |
胡启鹏, 郭志华, 李春燕, 马履一. 不同光环境下亚热带常绿阔叶树种和落叶阔叶树种幼苗的叶形态和光合生理特征. 生态学报, 2008, 28(7): 3262-3270. |
| [23] |
徐海东, 熊静, 成向荣, 凌高潮, 郑文华, 胡卫明, 虞木奎. 麻栎和闽楠幼苗叶功能性状及生物量对光照和施肥的响应. 生态学报, 2021, 41(6): 2129-2139. |
| [24] |
Poorter L, Bongers F. Leaf traits are good predictors of plant performance across 53 rain forest species. Ecology, 2006, 87(7): 1733-1743. |
| [25] |
乔雪涛, 何欣燕, 何俊, 范秀华. 不同秸秆填埋量对盐碱土水盐运移及垂柳反射光谱的影响. 生态学报, 2018, 38(22): 8107-8117. |
| [26] |
方晓雨, 张宏琴, 姜超, 卢文敏, 范秀华. 3种乔木叶片反射光谱与光合特性随树高的变化. 应用与环境生物学报, 2013, 19(2): 383-388. |
| [27] |
姜超, 黄珍, 方晓雨, 程艳霞. 长白山5种槭属植物光合及反射光谱特性. 应用与环境生物学报, 2013, 19(4): 713-717. |
| [28] |
何怀江, 张忠辉, 张春雨, 郝珉辉, 姚杰, 解蛰, 高海涛, 赵秀海. 采伐强度对东北针阔混交林林分生长和物种多样性的短期影响. 林业科学, 2019, 55(2): 1-12. |
| [29] |
胡乘风, 陈巧玲, 乔雪涛, 程艳霞. 阔叶红松林主要树种光合与光谱反射特性及初级生产力研究. 北京林业大学学报, 2020, 42(5): 12-24. |
| [30] |
Farquhar G D, Von C S, Berry J A. Models of photosynthesis. Plant Physiology, 2001, 125(1): 42-45. |
| [31] |
Sims D A, Gamon J A. Relationships between leaf pigment content and spectral reflectance across a wide range of species, leaf structures and developmental stages. Remote Sensing of Environment, 2002, 81(2/3): 337-354. |
| [32] |
刘伟国, 范秀华. 长白山红松阔叶林主要树种光谱反射特征对光环境的响应. 生态学报, 2009, 29(9): 4720-4726. |
| [33] |
Merzlyak M N, Gitelson A A, Chivkunova O B, Rakitin V Y. Non-destructive optical detection of pigment changes during leaf senescence and fruit ripening. Physiologia Plantarum, 1999, 106(1): 135-141. |
| [34] |
Penuelas J, Pinol J, Ogaya R, Filella I. Estimation of plant water concentration by the reflectance Water Index WI (R900/R970). International Journal of Remote Sensing, 1997, 18(13): 2869-2875. |
| [35] |
张兰, 王静, 张金峰, 邓晓娟, 罗永红, 闫兴富. 辽东栎幼苗生长和生理特性对光照强度的响应. 中南林业科技大学学报, 2021, 41(11): 73-81. |
| [36] |
Richardson A D, Berlyn G P. Spectral reflectance and photosynthetic properties of Betula papyrifera (Betulaceae) leaves along an elevational gradient on Mt. Mansfield, Vermont, USA. American Journal of Botany, 2002, 89(1): 88-94. |
| [37] |
刘泽彬, 程瑞梅, 肖文发, 郭泉水, 王娜. 遮荫对中华蚊母树苗期生长及光合特性的影响. 林业科学, 2015, 51(2): 129-136. |
| [38] |
Gamage H K, Jesson L. Leaf heteroblasty is not an adaptation to shade: seedling anatomical and physiological responses to light. New Zealand Journal of Ecology, 2007, 31(2): 245-254. |
| [39] |
杨莹, 王传华, 刘艳红. 光照对鄂东南2种落叶阔叶树种幼苗生长、光合特性和生物量分配的影响. 生态学报, 2010, 30(22): 6082-6090. |
| [40] |
管铭, 金则新, 王强, 李月灵, 左威. 千岛湖次生林优势种植物光合特性对不同光环境的响应. 应用生态学报, 2014, 25(6): 1615-1622. |
| [41] |
唐星林, 姜姜, 金洪平, 周晨, 刘光正, 杨桦. 遮阴对闽楠叶绿素含量和光合特性的影响. 应用生态学报, 2019, 30(9): 2941-2948. |
| [42] |
冯晓钰, 周广胜. 夏玉米叶片水分变化与光合作用和土壤水分的关系. 生态学报, 2018, 38(1): 177-185. |
| [43] |
唐钢梁, 李向义, 林丽莎, 李磊, 鲁建荣. 骆驼刺在不同遮阴下的水分状况变化及其生理响应. 植物生态学报, 2013, 37(4): 354-364. |
| [44] |
衣英华, 樊大勇, 谢宗强, 陈芳清. 模拟淹水对枫杨和栓皮栎气体交换、叶绿素荧光和水势的影响. 植物生态学报, 2006, 30(6): 960-968. |