 2022, Vol. 42
2022, Vol. 42文章信息
- 张锦堂, 潘志立, 田云海, 侯佳蓝, 彭明春, 段禾祥, 李云芳, 李永萍
- ZHANG Jintang, PAN Zhili, TIAN Yunhai, HOU Jialan, PENG Mingchun, DUAN Hexiang, LI Yunfang, LI Yongping
- 云龙天池国家级保护区云南松种群年龄结构及动态分析
- Age structure and dynamics of Pinus yunnanensis population in Yunlong Tianchi Nature Reserve
- 生态学报. 2022, 42(22): 9091-9099
- Acta Ecologica Sinica. 2022, 42(22): 9091-9099
- http://dx.doi.org/10.5846/stxb202105081208
-
文章历史
- 收稿日期: 2021-05-08
- 网络出版日期: 2022-07-12

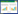




2. 云南大学生态与环境学院, 昆明 650091;
3. 云南省环境科学研究院, 昆明 650034;
4. 云南云龙天池国家级自然保护区管护局, 云龙 672700
2. School of Ecology and Environmental Science, Yunnan University, Kunming 650091, China;
3. Yunnan Institute of Environmental Science, Kunming 650034, China;
4. Management and Protection Bureau of Yunnan Yunlong Tianchi National Nature Reserve, Yunlong 672700, China
云南松(Pinus yunnanensis)是松科(Pinaceae)松属的裸子植物, 是我国西南地区特有的树种, 也是西南地区荒山造林的先锋树种, 具有重要的经济、生态和社会效益[1—3]。云南松林是暖温性针叶林的典型代表, 在滇中高原分布极广。现存的云南松天然林大都是在地带性亚热带半湿润常绿阔叶林遭受破坏后, 在迹地上更新起来的天然次生林, 人工林较少[4]。云南松林是否会被常绿阔叶林所替代是群落演替研究的核心问题。明晰云南松天然林在无人为干扰的自然状态下的种群结构和动态是探究云南松天然林能否实现自我更新的关键, 对探究该地区的森林群落的演替具有重要意义。
目前已有较多的研究关注云南松的种群结构。王磊等[5]的综述认为云南松天然林群落结构单一, 退化的云南松种群缺乏大径级个体, 出现低龄化特征。还有研究也认为云南松林下无小苗、小树不能实现种群更新和维系, 是衰退的种群, 将会被常绿阔叶林所替代[6—7]。而黄小波等[8]和Tang等[9]的研究都认为云南松种群是能够自我更新的种群。何斌等[10]对黔西北地区云南松种群结构及动态进行了研究, 但黔西北地区位于云南松分布的东界, 其研究结果不能确切全面反映云南松种群的发展趋势。因此, 有必要对未受干扰自然状态下的云南松种群的结构和动态进行研究, 从而明晰云南松天然林的发展趋势。
云南云龙天池国家级自然保护区位于大理州云龙县, 是云南松种质资源保护基地。研究认为该保护区内的云南松林是目前留存下来较为原始的云南松森林群落[9], 区内分布有树龄100年以上的云南松个体。该保护区为研究未受人为干扰下云南松的种群结构和动态提供了最佳的试验场所。
本文以云龙天池国家级自然保护区云南松天然种群为研究对象, 采用静态生命表、存活曲线、生存分析和谱分析等方法研究云南松种群结构和动态特征, 旨在阐明云南松天然林的种群发展动态, 为研究该地区的云南松群落可持续发展提供依据。
1 材料与方法 1.1 研究区概况调查区域位于云南省大理州云龙天池国家级自然保护区(25°49′48″—26°14′16″N, 99°11′36″—99°20′34″E)(以下简称“云龙天池保护区”)。全区干湿季分明, 年均气温13.2℃左右, 年均降水量为815.5 mm左右, 海拔为2100.0—3638.9 m, 土壤以棕壤为主。该保护区内的云南松林还伴生少量的其他乔木和灌木, 如:华山松(Pinus armandii)、锐齿槲栎(Quercus aliena)、大白杜鹃(Rhododendron decorum)、南烛(Vaccinium bracteatum)、水红木(Viburnum cylindricum)、银木荷(Schima argentea)、山杨(Populus davidiana)、散毛樱桃(Cerasus patentipila)、乌鸦果(Vaccinium fragile)、西南栒子(Cotoneaster franchetii)、金丝桃(Hypericum monogynum)、野拔子(Elsholtzia rugulosa)、小檗(Berberis amurensis)等, 草本植物主要有草血竭(Polygonum paleaceum)、剪股颖(Agrostis clavata)、一把伞南星(Arisaema erubescens)、荩草(Arthraxon hispidus)、藏象牙参(Roscoea tibetica)、大花夏枯草(Prunella grandiflora)、大戟(Euphorbia pekinensis)、云南兔儿风(Ainsliaea yunnanensis)、狼毒(Stellera chamaejasme)、瓦韦(Lepisorus thunbergianus)、木蓝(Indigofera tinctoria)等。
1.2 样地设置与种群调查对云龙天池保护区进行踏查后, 选择具有代表性的云南松中龄林、近熟林、成熟林和过熟林, 分别布设1个20 m × 50 m的样方。记录每个样方的经纬度、海拔、坡度、坡向、坡位、坡面等信息, 4个样方的基本情况见表 1。对每个样方内的树高> 1.3 m的云南松, 记录其胸径(diameter at breast height, DBH)、树高;对于树高 < 1.3 m的云南松, 划分为幼苗(5 cm≤H < 50 cm)和幼树(50 cm≤H < 130 cm)分别记录株数、树高[9]。
| 样方 Quadrat |
面积 Plot area |
海拔/m Elevation |
坡向/(°) Aspect |
坡度/(°) Slope |
坡位 Slope position |
坡面 Slope surface |
北纬 Northern latitude |
东经 East longitude |
郁闭度 Canopy density |
| 中龄林 Middle-aged stand |
20 m × 50 m | 2577 | 西南243 | 21 | 中 | 凸 | 25°52′20.820″ | 99 °17′13.680″ | 0.80 |
| 近熟林 Near-mature stand |
20 m × 50 m | 2530 | 东南152 | 36 | 下 | 凸 | 25°52′9.012″ | 99°16′21.468″ | 0.80 |
| 成熟林 Mature stand |
20 m × 50 m | 2464 | 东107 | 7 | 下 | 平 | 25°50′56.634″ | 99°16′20.021″ | 0.75 |
| 过熟林 Over-mature stand |
20 m × 50 m | 2649 | 西北313 | 22 | 中 | 平 | 25°52′34.907″ | 99°16′40.847″ | 0.80 |
本研究用种群胸径代替年龄, 绘制种群年龄结构图。胸径级的划分标准参考Tang等[9], 每间隔5 cm为1级, 即DBH < 5 cm(包括树高小于1.3 m的植株), 为第Ⅰ龄级;5 cm≤DBH < 10 cm, 为第Ⅱ龄级;依此类推, 75 cm≤DBH < 80 cm, 为第XVI龄级, 共16级。以龄级作为横坐标, 样方中活立木的株数作为纵坐标, 绘制云南松的龄级结构图。
为定量描述云南松种群动态特征, 参考“植物种群与群落结构动态量化分析方法研究”[11]。其中涉及到的参数及其意义如下:Vn表示从其n到n+1龄级间的个体数量变化动态, Vpi是整个种群年龄结构的数量变化动态指数, V′pi是考虑未来的外部环境干扰的整个种群年龄结构的数量变化动态指数, P是种群在外部环境干扰下承担的风险概率。
1.4 静态生命表的编制及种群的生存分析静态生命表的编制参考“珍稀濒危植物长苞铁杉种群生命表分析”[12], 龄级划分同1.3。表中参数主要有:ax是x龄级内树木现存个体数;lx是标准化存活个体数(一般最小龄级标准化成1000);dx、qx分别是相邻龄级间隔期内树木标准化死亡数和相应的死亡率;Lx是从x进入x+1龄级间隔期间还存活的树木个体数(区间寿命);Tx是x龄级到超过x龄级的树木个体总数(总寿命);ex是进入x龄级后树木个体的平均期望寿命(生命期望);Kx是各年龄组致死力, 即消失率;Sx为存活率。
在编制静态生命表的过程中, 当出现种群的死亡率为负值的情况时采用分段匀滑技术对种群个体的存活数据进行处理[13], x龄级现有的存活数ax经过匀滑修正处理后可以得到ax*值。绘制云南松的存活曲线, 横坐标为与径级对应的龄级, 纵坐标为lnlx。分别采用指数函数Nx=N0e-bx和幂函数Nx=N0x-b模型检验云南松种群的存活曲线类型[12, 14—15]。
计算云南松的种群生存率(S(i))、累计死亡率(F(i))、死亡密度(f(ti))、危险率(λ(ti)), 并绘制生存率、累计死亡率、死亡密度和危险率曲线[16—18], 公式如下:
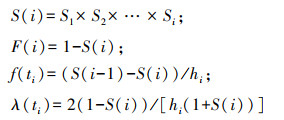
|
式中, Si为存活率, hi为龄级宽度, ti表示第i龄级。
1.5 谱分析谱分析是傅里叶级数(Fourier series)的展开式, Fourier证明过复杂的周期现象可以由不同振幅和相应的谐波组成[19—21], 写成正弦波形式为:
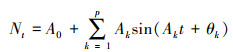
|
式中, Nt为t时刻种群的大小;A0为周期变化的平均值;Ak为各谐波的振幅(k=1, 2, 3, …, p), Ak的大小标志着各周期作用的大小差别;A1为基波的振幅, A2, A3, …, Ap为谐波的振幅, p=n/2是谐波的个数, n为时间系列总长度(龄级总数);ωk和θk分别是谐波频率和相角;T为正弦波的基本周期(时间系列t的最长周期);每个谐波的周期分别是基本周期的1/2, 1/3, …, 1/p。
在编制生命表时, 云南松种群共划分为16个龄级, 所以谐波的个数p=n/2=8。将静态生命表的龄级作为谱分析的级差, 谱分析方法涉及到的参数的计算参照“阔叶红松林红松种群动态的谱分析”[19]。
1.6 数据分析使用Microsoft Excel 2010完成数据的统计整理、制图, 使用SPSS 25完成模型拟合及统计检验。
2 结果与分析 2.1 年龄结构及动态量化云南松种群年龄结构为增长型(倒J型)(图 1), 随着龄级的增大, 云南松个体数逐渐减少。第Ⅰ—Ⅶ龄级个体占调查总数的91.79%, 第Ⅱ龄级和Ⅲ龄级的云南松数目最多, 分别占所有云南松数目的33.05%和23.37%。第Ⅳ龄级(46株)和第Ⅴ龄级(45株)、第Ⅵ龄级(24株)和第Ⅶ龄级(23株)的云南松数目几乎相同。第Ⅸ—XVI龄级的云南松数目较少, 仅占6.11%。

|
| 图 1 云南松种群龄级结构 Fig. 1 Size class structure of P. yunnanensis population Ⅰ:DBH (胸径, diameter at breast height) < 5 cm;Ⅱ:5 cm≤DBH < 10 cm;Ⅲ:10 cm≤DBH < 15 cm;Ⅳ:15 cm≤DBH < 20 cm;Ⅴ:20 cm≤DBH < 25 cm;Ⅵ:25 cm≤DBH < 30 cm;Ⅶ:30 cm≤DBH < 35 cm;Ⅷ:35 cm≤DBH < 40 cm;Ⅸ:40 cm≤DBH < 45 cm;Ⅹ:45 cm≤DBH < 50 cm;Ⅺ:50 cm≤DBH < 55 cm;Ⅻ:55 cm≤DBH < 60 cm;XIII:60 cm≤DBH < 65 cm;XIV:65 cm≤DBH < 70 cm;XV:70 cm≤DBH < 75 cm;XVI:75 cm≤DBH < 80 cm |
种群数量变化动态显示(表 2), 云南松种群动态指数V1、V9、V11和V14均小于0, 表明云南松在第Ⅰ—Ⅱ龄级、第Ⅸ—Ⅹ龄级、第Ⅺ—Ⅻ龄级、第XIV—XV龄级间个体数量减少;V2—V8、V10、V12、V13和V15均大于0, 表明Ⅱ—Ⅸ龄级、第Ⅹ—Ⅺ龄级、第XIII—XIV龄级以及第XVI—XV龄级云南松个体数量呈现增长的结构动态特征。云南松整个种群年龄结构的数量变化动态指数Vpi为28.29%, 种群呈现增长的年龄结构数量动态关系, 考虑随机干扰情况下的动态指数V′pi为1.77%, Vpi和V′pi都大于0, 表明云南松种群为增长型种群结构。Pmax为6.25%表明云南松种群具有一定的抗干扰能力。
| 龄级 Age class |
种群动态指数 Dynamic index |
动态指数值/% Dynamic index value |
龄级 Age class |
种群动态指数 Dynamic index |
动态指数值/% Dynamic index value |
|
| Ⅰ | V1 | -80.89 | Ⅹ | V10 | 42.86 | |
| Ⅱ | V2 | 29.30 | Ⅺ | V11 | -20.00 | |
| Ⅲ | V3 | 58.56 | Ⅻ | V12 | 20.00 | |
| Ⅳ | V4 | 2.17 | XIII | V13 | 100.00 | |
| Ⅴ | V5 | 46.67 | XIV | V14 | -100.00 | |
| Ⅵ | V6 | 4.17 | XV | V15 | 50.00 | |
| Ⅶ | V7 | 56.52 | Vpi | 28.29 | ||
| Ⅷ | V8 | 40.00 | V′pi | 1.77 | ||
| Ⅸ | V9 | -14.29 | Pmax | 6.25 | ||
| V1—V15:相邻龄级数量动态变化指数 quantitative dynamics index of adjacent age class;Vpi:种群数量动态变化指数 quantitative dynamics index of population;V′pi:受外界干扰时种群数量动态变化指数 quantitative dynamics index of population by external interference;Pmax:随机干扰风险概率极大值 the maximum of probability in random disturbance | ||||||
随龄级的增加, 云南松匀滑处理后的存活数(ax*)和标准化存活数(lx)逐渐减小(表 3)。云南松种群的死亡率(qx)和消失率(Kx)反映了该种群随龄级增加的动态特征。死亡率和消失率在第Ⅰ—Ⅷ龄级存在较大的波动性, 第Ⅱ、第Ⅴ、第Ⅶ龄级均出现了峰值。死亡率和消失率从第Ⅸ龄级开始, 随着龄级的增加逐渐增长, 在第XVI龄级处出现最大值。生命期望(ex)反映x龄级内植株个体的平均生存能力, 云南松种群生命期望最高的3个龄级依次为第Ⅵ、第Ⅸ和第Ⅲ龄级, 从第Ⅷ—XVI龄级的生命期望逐渐降低, 第XVI龄级生命期望值最低。存活率(Sx)在第Ⅱ、第Ⅴ和第Ⅶ龄级较低, 分别为0.576、0.533和0.435, 在第Ⅸ—XV龄级期间, 存活率随着龄级的增大逐渐减小。
| 龄级 Age class |
胸径/cm Diameter at breast height |
组中值/cm Mid-value of class |
ax | ax* | lx | lnlx | dx | qx | Lx | Tx | ex | Kx | Sx |
| Ⅰ | 0—5 | 2.5 | 30 | 141 | 1000 | 6.908 | 298 | 0.298 | 851 | 2911 | 2.911 | 0.354 | 0.702 |
| Ⅱ | 5—10 | 7.5 | 157 | 99 | 702 | 6.554 | 298 | 0.424 | 553 | 2060 | 2.934 | 0.552 | 0.576 |
| Ⅲ | 10—15 | 12.5 | 111 | 57 | 404 | 6.002 | 78 | 0.193 | 365 | 1507 | 3.728 | 0.214 | 0.807 |
| Ⅳ | 15—20 | 17.5 | 46 | 46 | 326 | 5.788 | 7 | 0.022 | 323 | 1142 | 3.500 | 0.022 | 0.978 |
| Ⅴ | 20—25 | 22.5 | 45 | 45 | 319 | 5.766 | 149 | 0.467 | 245 | 819 | 2.567 | 0.629 | 0.533 |
| Ⅵ | 25—30 | 27.5 | 24 | 24 | 170 | 5.137 | 7 | 0.042 | 167 | 574 | 3.375 | 0.043 | 0.958 |
| Ⅶ | 30—35 | 32.5 | 23 | 23 | 163 | 5.094 | 92 | 0.565 | 117 | 408 | 2.500 | 0.833 | 0.435 |
| Ⅷ | 35—40 | 37.5 | 10 | 10 | 71 | 4.262 | 14 | 0.200 | 64 | 291 | 4.100 | 0.223 | 0.800 |
| Ⅸ | 40—45 | 42.5 | 6 | 8 | 57 | 4.038 | 7 | 0.125 | 53 | 227 | 4.000 | 0.134 | 0.875 |
| Ⅹ | 45—50 | 47.5 | 7 | 7 | 50 | 3.905 | 7 | 0.143 | 46 | 174 | 3.500 | 0.154 | 0.857 |
| Ⅺ | 50—55 | 52.5 | 4 | 6 | 43 | 3.751 | 7 | 0.167 | 39 | 128 | 3.000 | 0.182 | 0.833 |
| Ⅻ | 55—60 | 57.5 | 5 | 5 | 35 | 3.568 | 7 | 0.200 | 32 | 89 | 2.500 | 0.223 | 0.800 |
| XIII | 60—65 | 62.5 | 4 | 4 | 28 | 3.345 | 7 | 0.250 | 25 | 57 | 2.000 | 0.288 | 0.750 |
| XIV | 65—70 | 67.5 | 0 | 3 | 21 | 3.058 | 7 | 0.333 | 18 | 32 | 1.500 | 0.405 | 0.667 |
| XV | 70—75 | 72.5 | 2 | 2 | 14 | 2.652 | 7 | 0.500 | 11 | 14 | 1.000 | 0.693 | 0.500 |
| XVI | 75—80 | 77.5 | 1 | 1 | 7 | 1.959 | 7 | 1.000 | 4 | 4 | 0.500 | 1.959 | — |
| ax:x龄级内树木现存个体数 individual number of age class x;ax*:匀滑处理后的存活数 the correction value of ax;lx:标准化存活个体数 standardized survivors number;lnlx:标准化存活个体数的自然对数 the natural logarithm of lx;dx:相邻龄级间隔期内树木标准化死亡数 the standardized number of death individuals from age class x to age class x + 1;qx:相邻龄级间隔期内树木死亡率 mortality from age class x to age class x+1;Lx:从x进入x+1龄级间隔期间还存活的树木个体数 the number of surviving individuals from age class x to age class x + 1;Tx:x龄级到超过x龄级的树木个体总数 the total individuals number of age class x and age class elder than x;ex:进入x龄级后树木个体的生命期望 life expectancy of individuals in the age class x;Kx:消失率 vanish rate;Sx:存活率 survival rate | |||||||||||||
云南松种群存活曲线呈现随龄级增大而下降的趋势(图 2)。第Ⅴ—Ⅵ龄级及Ⅶ—Ⅷ龄级下降幅度最大, 仅有13.33%的云南松能从第Ⅴ龄级穿过该环境筛到达第Ⅷ龄级。通过第Ⅷ龄级之后, 云南松种群存活曲线趋于平缓。

|
| 图 2 云南松种群存活曲线 Fig. 2 Survival curve of P. yunnanensis population |
存活曲线拟合模型结果显示, 指数函数的F值(264.547)及相关系数R(0.975)均大于幂函数的F值(48.326)及相关系数R(0.881)(表 4)。因此, 云龙天池保护区的云南松种群更趋向于Deevey-Ⅱ型(对角线型)。Deevey-Ⅱ型有三个亚型(B1, B2和B3亚型), 结合图 2可以看出, 该存活曲线属于其中的B1亚型, 表明各龄级的存活数围绕对角线产生波动, 并且各个龄级的存活数相差较大。
| 存活曲线 Survival curve | 拟合模型 Fitting model | 拟合结果 Fitting result | F | R |
| Deevey-Ⅱ | Nx=N0e-bx | Nx=7.823e-0.072x | 264.547 | 0.975 |
| Deevey-Ⅲ | Nx=N0x-b | Nx=8.997x-0.391 | 48.326 | 0.881 |
| Nx:匀滑后x龄级内存活数 the correction value of ax;N0:种群形成初期的个体存活数 number of surviving individuals in the early formation stage of population | ||||
云南松的生存率随龄级增大而降低, 累计死亡率随龄级增大逐渐增大(图 3)。在Ⅰ—Ⅶ龄级, 种群生存率和累计死亡率变化较大, Ⅶ—XVI龄级生存率和死亡率趋于稳定(图 3)。第Ⅰ龄级的生存率为0.702, 第Ⅶ龄级迅速降低至0.071;第Ⅰ龄级的累计死亡率为0.298, 到第Ⅶ龄级迅速增加为0.929。第Ⅶ—XVI龄级云南松种群的生存率和累计死亡率的增减变缓, 大量个体由于到达生理年龄而死亡, 种群呈现衰退特征, 说明该阶段云南松种群进入生理衰老阶段。

|
| 图 3 云南松种群生存率函数S(i)和累计死亡率函数F(i)曲线 Fig. 3 Survival rate S(i) and cumulative mortality rate F(i) functional curve of P. yunnanensis population |
由图 4可知, 云南松种群的死亡密度在第Ⅰ—Ⅶ龄级存在波动, 之后较平稳降低。危险率呈递增状态, 第Ⅰ—Ⅶ龄级增长较快, 第Ⅷ—XVI龄级增长缓慢。综上, 云南松种群第Ⅰ—Ⅶ龄级生命状态不稳定, 死亡可能性较大, 种群个体数量减少, 第Ⅷ—XVI龄级表现出衰退的特点。这与静态生命表、存活曲线的分析结果是相符的。

|
| 图 4 云南松种群死亡密度f(ti)和危险率λ(ti)曲线 Fig. 4 Mortality density f(ti)and hazard rate λ(ti) functional curve of P. yunnanensis population |
谱分析结果显示, 振幅值呈现逐渐减小的趋势, 其中基波A1最大, 说明云南松种群的具有明显的大周期(表 5)。基本的周期长度为种群本身所固有, 表明云南松种群数量动态变化主要受生物学特征所控制。
| 谐波 Harmonic | A1 | A2 | A3 | A4 | A5 | A6 | A7 | A8 |
| 振幅值 Amplitude value | 1.613 | 0.836 | 0.552 | 0.205 | 0.147 | 0.341 | 0.209 | 0.291 |
| A1:基波振幅 fundamental amplitude;A2—A8:各谐波振幅 amplitude of each harmonic | ||||||||
此外, 云南松种群除了受基波的影响外, 也表现出了其中迭加了明显的小周期波动, 即A6和A8。A6对应的周期为基本周期的1/6, 胸径为5 × 16/6=13.3 cm, 对应第Ⅲ龄级;A8对应的周期为基本周期的1/8, 胸径为5 × 16/8=10 cm, 也对应第Ⅲ龄级。可见, 第Ⅲ龄级(胸径10—15 cm)是影响云南松种群波动的重要阶段。
3 讨论云龙天池自然保护区云南松种群的年龄结构表明其是增长型种群。种群的年龄结构可以作为判断种群是否衰退的依据。一般认为, 具有间歇型和倒J型的种群更新良好[22—23]。本研究中云南松种群胸径级结构属于倒J型的种群, 这与黄小波等[8]和Tang等[9]的研究结果一致。云南松种群数量动态指数Vpi和V′pi大于0, 也表明该云南松种群属于增长型种群。调查中发现云南松林下幼苗较少, 但这不能说明种群是衰退的。种源和更新方式是影响种群持续的主要因素。云南松进入成熟期后每年能够产生大量的种子, 不缺少种源[24]。云南松可以通过林窗、林下实现更新, 也能在火烧后迅速长出幼苗[4, 25]。因此, 云南松是能持续更新的种群。另外, 尽管本研究的样方中存在有常绿阔叶树种, 如银木荷(S. argentea), 但是银木荷在样方中的幼树仅仅30株, 树高均在20 m以下, 无论数量质量都远低于云南松。可见, 云南松种群年龄结构为增长型, 更新方式多样, 种源充足, 短期内不会出现衰退。
云龙天池自然保护区云南松种群也表现出低龄级的个体数量波动大, 死亡率高的特点。结合静态生命表、存活曲线和生存分析函数也表明云南松种群在第Ⅰ—Ⅶ龄级有较为明显的波动。通过谱分析进一步确认, 低龄级的胸径10—15 cm云南松个体之间存在强烈的竞争。之所以出现这样的现象是因为云南松中幼林密度较大且处于生长旺盛的阶段, 对资源与空间的需求较大[26]。云南松是喜光树种, 光照是其竞争的主要资源。金振洲等[4]认为, 随着云南松的生长及林分的郁闭, 缺乏光照会使得长势较差的个体枯死。云南松幼龄林的数量波动是种群通过自疏剔除冗余的过程, 也是种群维系的重要方式[27]。
云南松林是暖温性针叶林的典型代表, 主要分布于滇中高原山地。本研究选择在云南云龙天池自然保护区进行, 该保护区的保护对象是云南松种群, 为进行云南松种群结构研究提供了典型试验场所。对长寿命的木本植物的种群数量动态研究主要以“空间推时间”的方法, 编制静态生命表进行[13, 28—32]。树木静态生命表的编制需要提供树木的确切年龄, 生长锥钻取胸高部位的树芯和WinDENDRO年轮分析系统分析树轮[33]是常用的测定树木的准确年龄的方法。但是这些方法操作复杂, 在野外调查中制约了工作效率提高。树木的年龄和胸径、地径及树高存在正相关关系[34], 实际工作中常常用胸径、地径及树高代替树木年龄[35]。Tang等[9]对该保护区内云南松的年龄和胸径进行拟合后发现该区域云南松的年龄和胸径存在显著的线性相关关系。因此, 本研究用云南松的胸径代替年龄进行种群年龄结构的研究是可行的。
4 结论云南云龙天池自然保护区云南松种群是能够自我更新的种群, 种群动态呈现出低龄级个体数量波动大, 高龄级衰退的特点。建议在云南松林的幼龄林和中龄林适当采取抚育间伐措施促进其种群的生长发育及更新, 维持种群的稳定性。
致谢: 感谢云南云龙天池国家级自然保护区管护局的支持,感谢杨桂天对野外调查采样的帮助。| [1] |
中国科学院中国植物志编辑委员会. 中国植物志. 北京: 科学出版社, 1998, 23(1): 112. DOI:10.3969/j.issn.1001-5965.1998.01.029 |
| [2] |
魏巍, 许玉兰, 李根前, 吕学辉, 陈诗, 蔡年辉. 一年生云南松苗期生长动态规律研究. 西北林学院学报, 2012, 27(1): 75-80. |
| [3] |
邓喜庆, 皇宝林, 温庆忠, 华朝朗, 陶晶. 云南松林在云南的分布研究. 云南大学学报: 自然科学版, 2013, 35(6): 843-848. |
| [4] |
金振洲, 彭鉴. 云南松. 昆明: 云南科技出版社, 2004.
|
| [5] |
王磊, 张劲峰, 马建忠, 魏巍, 胡青. 云南松及其林分退化现状与生态系统功能研究进展. 西部林业科学, 2018, 47(6): 121-130. |
| [6] |
蔡年辉, 李根前, 陆元昌. 云南松纯林近自然化改造的探讨. 西北林学院学报, 2006, 21(4): 85-88, 120. DOI:10.3969/j.issn.1001-7461.2006.04.023 |
| [7] |
王健敏, 刘娟, 陈晓鸣, 温庆忠, 段兆尧, 赖兴会. 云南松天然林及人工林群落结构和物种多样性比较. 林业科学研究, 2010, 23(4): 515-522. DOI:10.13275/j.cnki.lykxyj.2010.04.021 |
| [8] |
黄小波, 李帅锋, 苏建荣, 刘万德, 郎学东. 云龙天池云南松自然种群分布格局分析. 林业科学研究, 2018, 31(4): 47-52. |
| [9] |
Tang C Q, Shen L Q, Han P B, Huang D S, Li S F, Li Y F, Song K, Zhang Z Y, Yin L Y, Yin R H, Xu H M. Forest characteristics, population structure and growth trends of Pinus yunnanensis in Tianchi National Nature Reserve of Yunnan, southwestern China. Vegetation Classification and Survey, 2020, 1: 7-20. |
| [10] |
何斌, 李青, 李仰征, 李望军. 黔西北地区云南松种群数量动态及空间分布格局. 林业资源管理, 2021(2): 149-157. |
| [11] |
陈晓德. 植物种群与群落结构动态量化分析方法研究. 生态学报, 1998, 18(2): 214-217. |
| [12] |
吴承祯, 洪伟, 谢金寿, 吴继林. 珍稀濒危植物长苞铁杉种群生命表分析. 应用生态学报, 2000, 11(3): 333-336. |
| [13] |
江洪. 云杉种群生态学. 北京: 中国林业出版社, 1992.
|
| [14] |
Deevey E S Jr. Life tables for natural populations of animals. The Quarterly Review of Biology, 1947, 22(4): 283-314. |
| [15] |
Hett J M, Loucks O L. Age structure models of balsam fir and eastern hemlock. Journal of Ecology, 1976, 64(3): 1029-1044. |
| [16] |
杨凤翔, 王顺庆, 徐海根, 李邦庆. 生存分析理论及其在研究生命表中的应用. 生态学报, 1991, 11(2): 153-158. |
| [17] |
胡元洁, 巨天珍, 米彩燕, 康晓亚, 张宋智, 马建伟. 小陇山国家级自然保护区华山松种群更新动态分析. 广西植物, 2013, 33(2): 247-252. |
| [18] |
张悦, 易雪梅, 王远遐, 姬兰柱, 吴培莉. 采伐对红松种群结构与动态的影响. 生态学报, 2015, 35(1): 38-45. |
| [19] |
伍业钢, 韩进轩. 阔叶红松林红松种群动态的谱分析. 生态学杂志, 1988, 7(1): 19-23. |
| [20] |
张维, 焦子伟, 任艳利, 杨允菲, 李建东. 新疆野核桃自然保护区不同坡向野核桃种群生命表与波动周期. 西北植物学报, 2015, 35(6): 1229-1237. |
| [21] |
陈远征, 马祥庆, 冯丽贞, 黄勇来, 郑群瑞. 濒危植物沉水樟的种群生命表和谱分析. 生态学报, 2006, 26(12): 4267-4272. |
| [22] |
达良俊, 杨永川, 宋永昌. 浙江天童国家森林公园常绿阔叶林主要组成种的种群结构及更新类型. 植物生态学报, 2004, 28(3): 376-384. |
| [23] |
Ohsawa M. Structural comparison of tropical montane rain forests along latitudinal and altitudinal gradients in south and east Asia. Vegetatio, 1991, 97(1): 1-10. |
| [24] |
Yang J D, Zhang Z M, Dawazhaxi, Wang B, Li Q, Yu Q C, Ou X K, Ali K. Spatial distribution patterns and intra-specific competition of pine (Pinus yunnanensis) in abandoned farmland under the Sloping Land Conservation Program. Ecological Engineering, 2019, 135: 17-27. |
| [25] |
杨文云. 滇中地区云南松天然林群落结构及天然更新规律[D]. 北京: 中国林业科学研究院, 2010.
|
| [26] |
黄小波, 刘万德, 苏建荣, 李帅锋, 郎学东. 云南松天然林的种内和种间竞争. 林业科学研究, 2016, 29(2): 209-215. |
| [27] |
李永萍. 云龙天池省级自然保护区云南松更新和自疏及其冗余机制的研究[D]. 昆明: 云南大学, 2006.
|
| [28] |
卢杰, 郭其强, 郑维列, 徐阿生. 藏东南高山松种群结构及动态特征. 林业科学, 2013, 49(8): 154-160. |
| [29] |
吴语嫣, 李守中, 孙眭涛, 杨贤宇, 黄思彤, 王凯. 长汀水土流失区侵蚀劣地马尾松种群动态. 生态学报, 2019, 39(6): 2082-2089. |
| [30] |
王泳腾, 黄治昊, 王俊, 张童, 崔国发. 燕山山脉黄檗种群结构与动态特征. 生态学报, 2021, 41(7): 2826-2834. |
| [31] |
Shen Z Q, Lu J, Hua M, Tang X Q, Qu X L, Xue J L, Fang J P. Population structure and spatial pattern analysis of Quercus aquifolioides on Sejila Mountain, Tibet, China. Journal of Forestry Research, 2018, 29(2): 405-414. |
| [32] |
Zhao B Q, Guo D G, Shao H B, Bai Z K. Investigating the population structure and spatial pattern of restored forests in an opencast coal mine, China. Environmental Earth Sences, 2017, 76(20): 679. |
| [33] |
程伟, 吴宁, 罗鹏. 岷江上游林线附近岷江冷杉种群的生存分析. 植物生态学报, 2005, 29(3): 349-353. |
| [34] |
Frost I, Rydin H. Spatial pattern and size distribution of the animal-dispersed tree Quercus robur in two spruce-dominated forests. Ecoscience, 2000, 7(1): 38-44. |
| [35] |
何亚平, 费世民, 蒋俊明, 陈秀明, 张旭东, 何飞. 不同龄级划分方法对种群存活分析的影响——以水灾迹地油松和华山松种群生存分析为例. 植物生态学报, 2008, 32(2): 448-455. |