 2022, Vol. 42
2022, Vol. 42文章信息
- 陈松鹤, 向晓玲, 雷芳, 邹乔生, 艾代龙, 郑亭, 黄秀兰, 樊高琼
- CHEN Songhe, XIANG Xiaoling, LEI Fang, ZOU Qiaosheng, AI Dailong, ZHENG Ting, HUANG Xiulan, FAN Gaoqiong
- 秸秆覆盖配施氮肥根际土真菌群落及其与小麦产量的关系
- Relationship between rhizosphere fungal community and wheat yield under straw mulching combined with nitrogen fertilizer
- 生态学报. 2022, 42(21): 8751-8761
- Acta Ecologica Sinica. 2022, 42(21): 8751-8761
- http://dx.doi.org/10.5846/stxb202112143553
-
文章历史
- 收稿日期: 2021-12-14
- 网络出版日期: 2022-03-21

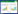




土壤微生物是影响肥力与质量状况的关键因素[1], 对土壤肥力形成与发展、植物发育生长与保持生态稳定等多个领域有关键的影响[2—3]。在四川丘陵旱地雨养农业区土壤有机质缺乏, 秸秆覆盖还田由土壤微生物驱动分解释放养分, 提高土壤有机质含量[4]。相较于细菌, 真菌有更强的吸收氮素和磷素及分解土壤中植物残体和复杂化合物的能力[3, 5]。因此, 研究秸秆覆盖与氮肥定位施用土壤真菌群落变化及其驱动因素对提升石灰性紫色土肥力和促进小麦生产具有重要意义。
秸秆作为一种有机肥源富含有机质及各营养元素, 农业生产中应用较为广泛[6]。秸秆还田不仅提高土壤速效养分的含量[4, 7], 而且为土壤微生物的生长和繁殖提供适宜的碳源和氮源[8—9], 从而提高了微生物多样性[8]和改变其群落结构[10]。真菌作为土壤中常见的微生物具有很强的降解复杂化合物能力[11], 在秸秆降解的过程中, 真菌的分解作用占据优势[12]。前人研究表明[13—15], 秸秆还田有助于提高真菌群落的丰富度, 改变真菌群落结构, 但也有研究认为秸秆还田会降低真菌Alpha多样性[16]。化学氮肥是提高作物产量常用肥料, 但长期施用氮肥会降低真菌群落多样性和改变其群落结构[17—18]。此外, 目前有关土壤真菌的研究主要集中在驱动真菌群落结构变化的土壤环境因素、土壤类型、施肥方式和栽培措施等方面[2, 16, 19—21]。秸秆覆盖和施氮对土壤真菌群落的影响虽有研究[22], 但针对秸秆覆盖配施氮肥如何影响根际真菌群落结构及促进作物生产力研究较少。
本研究基于石灰性紫色土连续4年秸秆覆盖与施氮定位试验, 采用Illumina Miseq测序技术, 研究真菌群落结构与多样性对秸秆覆盖与施氮的响应, 并结合土壤养分揭示其驱动因素及与小麦产量的关系, 为石灰性紫色土培肥地力和提高小麦生产力提供科学理论依据。
1 材料与方法 1.1 研究区概况该定位试验于2015年在四川省眉山市仁寿县珠嘉镇踏水村(30°04′ N, 104°13′ E)进行试验地属亚热带季风湿润气候, 丘陵地貌, 海拔482 m, 年均降雨量1009.4 mm, 年均气温17.4 ℃。土壤类型为石灰性紫色土。前茬作物为夏玉米, 研究对象冬小麦为后茬作物。2015年试验开始时土壤(0—20 cm)基础肥力:pH 7.82 (土∶水= 1∶2.5), 有机质16.9 g/kg, 全氮0.83 g/kg, 全磷0.86 g/kg, 全钾14.0 g/kg, 碱解氮68.2 mg/kg, 速效磷9.3 mg/kg, 速效钾158.2 mg/kg。
1.2 试验设计与方法试验采用二因素裂区设计, 主区为玉米秸秆粉碎覆盖(SM)与不覆盖(NSM);裂区为两种氮水平为:0(N0)和180(N1)kg/hm2, 共计4个处理, 每个处理均设4次重复, 小区面积为30 m2(6 m×5 m)。所有试验小区氮肥为尿素(N 46.2%), 磷肥和钾肥分别为过磷酸钙(P2O5 12%)和氯化钾(K2O 60%), 磷钾肥用量均为75 kg/hm2。60%的氮与所有磷、钾肥作为基肥撒施, 剩余氮肥于小麦拔节期作为追肥。于每年8月下旬实施玉米机械粉碎均匀撒于试验小区表面, 秸秆粉碎长度约5 cm, 覆盖量为8 000 kg/hm2。并于2018年10月底免耕开沟播种小麦(川麦104), 基本苗为2.25×106株/hm2。于2019年5月上旬收获。其他栽培管理措施同当地大田生产。
1.3 样品采集于2019年小麦开花期采集土壤样品, 此时期根际效应最为强烈[23]。每个处理采集15株小麦植株, 将整株小麦挖出, 采用抖根法将附着在根系上的土壤作为根际土, 使用冷藏箱将样品运输至实验室, 并在-80℃下储存, 直至DNA提取。同时采用“S”形5点采样法采集0—20 cm非根际土混为1个土样, 在室内通风处自然风干, 用于土壤养分测定。于小麦成熟期选定4 m2进行小麦产量测定。
1.4 项目测定与方法 1.4.1 土壤理化性状测定采用重铬酸钾-容量法测定有机碳(SOC), 凯氏定氮法测定全氮(TN), 碱解氮(AN)采用碱解扩散法, 速效磷(AP)使用0.5 mol/L NaHCO3浸提-钼锑抗比色法, 速效钾(AK)采用火焰光度法测定, 土壤pH值(水土比2.5∶1)采用电位法测定[24]。采用氯化钾(KCl)浸提-靛酚蓝比色法测定铵态氮(NH4+-N)和双波长分光光度法测定土壤硝态氮(NO3--N)[25]。
1.4.2 土壤DNA的提取和内源转录间隔区(ITS)基因的高通量测序准确称取0.5g新鲜土壤使用美国MOBIO公司生产的Fast DNA Spin Kit for Soil试剂盒参照说明书提取土壤总DNA。采用2%琼脂糖凝胶电泳检测提取DNA的纯度以及浓度。实验使用真菌18S引物为ITS5F: 5′-GGAAGTAAAAGTCGTAACAAGG-3′, ITS2R: 5′-GCTGCGTTCTTCATCGATGC-3′。扩增体系为25 μL, 主要构成为4 μL 5× FastPfu Buffer, 2 μL dNTPs(2.5 mmol/L), 正反引物各0.8 μL(5 μmol/L), 0.4 μL FastPfu Polymerase和10 ng模板DNA, ddH2O进行补充至体积。扩增条件所设定的具体参数是98℃预变性2 min, 设定为98℃持续变性处理10 s, 设定为55℃进行退火处理并持续30 s, 设定为72℃进行延伸处理持续30 s, 持续30个循环。各样本均进行3次重复的操作, 对同样本聚合酶链式反应(PCR)产物进行有效混合之后, 运用2%琼脂糖凝胶进行电泳测定工作, 完成前述操作后进行回收扩增子的处理。PCR扩增物用AgencourtAMPure珠纯化(Beckman Coulter, 使用PicoGreen dsDNA试剂盒(Invitrogen, Carlsbad, CA, USA))开展定量分析, 以同样浓度进行混合处理。采用Illumina公司MiSeq测序仪完成序列分析(由上海派森诺生物科技股份有限公司测定)。获得原始序列后进行质量控制, 在97%序列相似性水平上聚类成可操作的分类单元(OTUs), 对照RDP和UNITE数据库进行分类注释, 获取对应的真菌分类学信息。测序获得的具体数据已传递到NCBI SRA数据库, 具体编号参数是:SRP346982。
1.5 数据统计使用QIIME2 2019.4 (https://docs.qiime2.org/2019.4/tutorials/)按照官方教程中的描述处理序列。使用Microsoft Excel 2019整理数据, IBM SPSS Statistics 26进行相应的双因素方差分析和相关分析, 采用Fisher氏最小显著差检验法(LSD)进行平均值显著性多重比较(P < 0.05)。基于Bray-Curtis距离的主坐标分析(PCoA)和冗余分析(RDA)使用R v4.1.0软件的vegan包进行;利用主成分分析(PCA)分析真菌群落与小麦产量的关系;线性判别分析效应量(LEfSe)分析不同条件下微生物群落的特征, 以线性判别分析(LDA)值≥ 4.5被认为是每个处理的显著差异物种。采用Origin 2021绘图。
2 结果与分析 2.1 秸秆覆盖与施氮对小麦产量及土壤养分的影响秸秆覆盖和施氮提高了小麦产量(图 1), 差异显著。秸秆覆盖处理较不覆盖处理小麦产量增加40.3%。不覆盖施氮处理(NSMN1)比无覆盖不施氮处理(NSMN0)小麦产量显著提高75%;覆盖施氮处理(SMN1)较覆盖不施氮处理(SMN0)小麦产量显著提高92%。

|
| 图 1 秸秆覆盖与施氮处理下冬小麦产量 Fig. 1 Winter wheat yield under straw mulching and nitrogen application NSM和SM分别表示无秸秆覆盖和秸秆覆盖处理;N0表示不施氮, N1表示施氮量180 kg/hm2;不同小写字母表示NSM和SM处理内氮水平之间差异显著(P < 0.05);**表示极显著(P < 0.01) |
如表 1,SM处理相较于NSM处理土壤有机碳(SOC)含量增加44.4%, 差异极显著;施氮SOC含量有所增加, 但差异不显著。秸秆覆盖和施氮对土壤全氮(TN)、碱解氮(AN)、氨态氮(NH4+-N)、硝态氮(NO3--N)、速效磷(AP)和速效钾(AK)含量有显著或极显著的影响。SM处理较NSM处理土壤TN、AN、NH4+-N、NO3--N、AP和AK含量分别增加6.32%、7.69%、29.9%、24.6%和19.3%, NO3--N含量降低21.9%。无论秸秆覆盖与否, 施氮显著提高了土壤TN、AN、NH4+-N、NO3--N、AP和AK的含量。在不覆盖条件下, 与NSMN0处理相比, NSMN1处理土壤TN含量提高18.4%, AN含量提高12.1%、NH4+-N含量提高105.4%、NO3--N含量提高121.0%、AP含量提高10.0%和AK含量提高4.3%, 差异显著;覆盖处理下, SMN1处理较SMN0处理土壤TN含量提高14.9%, AN含量提高12.7%、NH4+-N含量提高56.1%、NO3--N含量提高95.5%、AP含量提高11.8%和AK含量提高1.7%, 差异显著。秸秆覆盖、施氮及其交互作用对土壤pH均无显著影响。
| 处理 Treatment |
氮水平 N level |
pH | 有机碳 SOC/(g/ kg) |
全氮 TN/(g/kg) |
碱解氮 AN/(mg/kg) |
氨态氮 NH4+-N/(mg/kg) |
硝态氮 NO3--N/(mg/kg) |
速效磷 AP/(mg/kg) |
速效钾 AK/(mg/kg) |
| NSM | N0 | 8.02a | 9.12a | 0.87b | 57.30b | 1.84b | 6.58b | 7.51b | 163.86b |
| N1 | 8.04a | 9.64a | 1.03a | 64.21a | 3.78a | 14.54a | 8.26a | 170.94a | |
| Mean | 8.03 | 9.53 | 0.95 | 60.76 | 2.81 | 10.56 | 7.89 | 167.40 | |
| SM | N0 | 8.01a | 13.62a | 0.94b | 61.53b | 2.85b | 5.58b | 8.44b | 198.08a |
| N1 | 8.06a | 13.90a | 1.08a | 69.32a | 4.45a | 10.91a | 9.44a | 201.49a | |
| Mean | 8.04 | 13.76 | 1.01 | 65.43 | 3.65 | 8.25 | 8.94 | 199.79 | |
| F值 | M | 0.02 n.s. | 262.33** | 35.93** | 235.06** | 91.74** | 69.43** | 34.25* | 1790.80** |
| N | 1.11 n.s. | 1.26 n.s. | 137.72** | 134.76** | 368.33** | 455.80** | 49.51** | 4.95* | |
| M×N | 0.24 n.s. | 0.13 n.s. | 0.64 n.s. | 0.47 n.s. | 3.50 n.s | 16.65** | 0.96 n.s. | 0.61 n.s. | |
| NSM和SM分别表示无秸秆覆盖和秸秆覆盖处理;M和N分别表示秸秆覆盖处理和施氮处理;N0表示不施氮, N1表示施氮量180 kg/hm2;不同小写字母表示NSM和SM处理内氮水平之间差异显著(P < 0.05);*和**表示P < 0.05和P < 0.01; n.s.表示没有显著差异 | |||||||||
由图 2可知, 秸秆覆盖和施氮对小麦根际真菌群落丰富度(Chao1指数)和多样性(Shannon指数)均有显著或极显著的影响。秸秆覆盖(SM)相对无覆盖(NSM)处理有利于提高小麦根际真菌群落丰富度与多样性。无论秸秆覆盖与否, 施氮处理(N1)与不施氮(N0)处理相比显著降低了真菌群落丰富度和多样性;真菌群落丰富度与多样性以SMN0处理最高。真菌群落以子囊菌门(Ascomycota)、担子菌门(Basidiomycota)与被孢霉门(Mortierellomycota)为所有处理的优势门(图 3), 其相对丰度分别为26%—49%、4.61%—26.9%和2.71%—14.5%。SM处理相较NSM处理显著增加担子菌门和降低被孢霉门相对丰度, 子囊菌门的相对丰度无显著差异(表 2)。在NSM条件下, 施氮显著增加子囊菌门的相对丰度, 担子菌门与被孢霉门相对丰度显著降低, 在SM处理中, 施氮处理显著降低子囊菌门的相对丰度, 担子菌门的相对丰度显著增加, 被孢霉门的相对丰度降低, 但无显著差异。在属水平上(图 3;表 2), SM处理较NSM处理显著增加了光盖伞属(Psilocybe)、弯孢菌属(Curvularia)和黑孢霉属(Nigrospora)的相对丰度, 被孢霉属(Mortierella)、球腔菌属(Mycosphaerella)、帚枝霉属(Sarocladium)、镰刀菌属(Fusarium)和柱霉属(Scytalidium)的相对丰度显著降低, 蛋白单胞属(Pyrenochaetopsis)的相对丰度无显著差异。在NSM处理中, 施氮处理较不施氮处理显著增加了球腔菌属、弯孢菌属和帚枝霉属的相对丰度, 显著降低被孢霉属和柱霉属的相对丰度;在SM处理中施氮则显著增加光盖伞属、弯孢菌属、帚枝霉属和柱霉属的相对丰度, 显著降低被孢霉属的相对丰度。

|
| 图 2 秸秆覆盖与施氮处理下冬小麦根际真菌群落多样性 Fig. 2 Alpha-diversity of bacterial communities in wheat rhizosphere soil under straw mulching and nitrogen fertilization treatments NSM和SM分别表示无秸秆覆盖和秸秆覆盖处理;N0表示不施氮,N1表示施氮量180 kg/ hm2;不同小写字母表示NSM和SM处理内氮水平之间差异显著(P < 0.05), **表示极显著(P < 0.01) |

|
| 图 3 秸秆覆盖和施氮处理下小麦根际土壤细菌群落组成 Fig. 3 Bacterial community composition in wheat rhizosphere soil under straw mulching and nitrogen fertilization treatments |
| 处理 Treatment |
门Phylum | ||||||||
| Ascomycota | Basidiomycota | Mortierellomycota | |||||||
| M | 2.9n.s. | 843.8** | 63.6** | ||||||
| N | 16.6** | 206.6** | 15.5** | ||||||
| M×N | 110.9** | 597.8** | 8.7n.s. | ||||||
| 处理 Treatment |
属Genus | ||||||||
| Psilocybe | Mortierella | Mycosphaerella | Curvularia | Sarocladium | Pyrenochaetopsis | Scytalidium | Fusarium | Nigrospora | |
| M | 3427.5** | 647.4** | 1077.3** | 1198.6** | 446.2** | 0.6n.s. | 171.5** | 52.7** | 430.0** |
| N | 2385.3** | 221.7** | 14.0** | 935.1** | 14.0** | 0.3n.s. | 137.4** | 0.7n.s. | 7.9* |
| M×N | 2384.4** | 116.6** | 14.3** | 815.5** | 0.01n.s. | 0.7n.s. | 45.2** | 0.1n.s. | 1.6n.s. |
| M和N分别表示秸秆覆盖处理和施氮处理;*和**分别表示P < 0.05和P < 0.01;n.s.表示差异不显著 | |||||||||
采用主坐标分析(PCoA), 第一、二主坐标轴对相应结构变异产生的解释量为37.8%和22.8%(图 4)。相同处理的4个重复之间, 得出的实际距离参数接近, 结构相似度较高。SM处理与NSM处理明显区分开, NSMN0和NSMN1处理在PC1轴上的投影较为接近, SMN1和SMN0处理在PC1轴上的投影与不覆盖处理两种施氮水平明显区分开, 说明秸秆覆盖还田改变了土壤真菌群落结构。通过组间群落差异分析(图 5), SM处理和NSM处理分别有15个和14个类群有显著差异。对NSM处理起重要作用的是被孢霉属和根霉属, SM处理主要是柱霉属、光盖伞属和弯孢菌属。说明秸秆覆盖和施氮改变了土壤中真菌特征类群。

|
| 图 4 秸秆覆盖与施氮处理下主坐标分析 Fig. 4 Principal coordinate analysis of straw mulching and nitrogen application |

|
| 图 5 秸秆覆盖和施氮肥处理下小麦根际土壤真菌的组间群落差异(LEfSe)分析 Fig. 5 LEfSe analysis fungi in wheat rhizosphere soil under straw mulching and nitrogen fertilization treatments |
在属水平上, 利用冗余分析方法研究了土壤理化性质与真菌群落组成的关系(图 6)。SM和NSM处理间群落组成的差异与SOC、AK、AP、NH4+-N和AN极显著相关, 与TN显著相关。通过主成分分析(图 7), 在NSM处理, 小麦产量与柱霉属和根瘤菌属高度相关;SM处理中, 光盖伞属、柱霉属和弯孢菌属与小麦产量高度相关。

|
| 图 6 秸秆覆盖和施氮肥处理下小麦根际土壤真菌群落和环境因子的冗余分析(RDA) Fig. 6 Redundancy analysis (RDA) of the fungal community and environmental factors in the wheat rhizosphere soil under straw mulching and nitrogen fertilization treatments SOC、TN、AN、NH4+-N、NO3--N、AP、AK和pH分别表示有机质、全氮、碱解氮、氨态氮、硝态氮、速效磷、速效钾和pH值;*和**分别表示P < 0.05和P < 0.01 |

|
| 图 7 秸秆覆盖和施氮处理下小麦根际土壤真菌群落与小麦产量的主成分分析 Fig. 7 Principal component analysis (PCA) of fungal communities in wheat rhizosphere soil and wheat yield under straw mulching and nitrogen application |
土壤有机碳(SOC)是反映土壤肥力变化的关键指标, 影响土壤的物理、化学以及生物特征[26]。先前的研究表明[27—28], 秸秆还田和氮肥配施土壤SOC含量显著增加。本试验中, 秸秆覆盖处理相较于不覆盖处理显著提高了土壤SOC含量, 且施氮比不施氮处理土壤SOC含量有所增加, 可能是由于小麦分蘖凋亡增加了对土壤的C输入, 同时减少分解过程中C的损失[29], 提高了土壤SOC含量。同时, 秸秆覆盖处理较不覆盖处理土壤TN、AN和NH4+-N含量显著增加, 主要由于外源氮添加和秸秆分解释放大量有机碳, 增加了土壤活性有机碳和氮, 缓解了微生物的营养限制, 进一步促进了土壤固氮和转化[30-31], 提高了土壤活性氮含量。此外, 无覆盖不施氮处理土壤TN含量比试验定位初土壤TN含量高出0.04g/kg, 是免耕播种减少了土壤氮素矿化损失以及前茬作物玉米种植氮残留。本研究中, 秸秆覆盖处理土壤NO3--N含量在开花时降低, 是小麦从拔节到开花的生长阶段对氮素需求旺盛[32], 而秸秆覆盖促进了小麦对硝酸盐的吸收, 从而减少土壤中NO3--N的残留。赵小军等[33]研究表明, 秸秆还田有利于促进土壤磷素向中等和高活性磷形态转化, 最终土壤速效磷含量提高[34]。在本研究区域, 石灰性紫色土土壤中磷素多与Ca2+结合[35], 形成作物难以利用的磷形态积累在土壤中, 降低了磷的有效性。本试验中秸秆覆盖处理较不覆盖处理土壤速效磷含量增加了24.6%, 促进了磷的有效性。此外, 钾元素大部分储存在作物秸秆中[16, 36], 在玉米秸秆分解过程中大量的钾元素进入土壤, 本研究中秸秆覆盖和施氮均促进土壤速效钾含量的增加。本试验条件下, 秸秆覆盖还田和施氮提高了石灰性紫色土土壤养分含量, 培肥了地力, 促进了小麦增产。
3.2 秸秆覆盖与施氮优化了根际真菌群落结构, 提高冬小麦生产力真菌是土壤中重要的分解者, 对土壤中植物残体和纤维素、半纤维素、木质素等难降解有机物具有较强的分解能力[3]和吸收转化氮素的能力[37]。覆盖处理提高真菌群落多样性, 施氮则降低真菌群落多样性, 这与秸秆覆盖后的有机质增加、土壤温度及湿度等存在非常密切的联系[38—39]。本研究中, 秸秆覆盖相比单施氮肥增加了真菌群落的多样性, 从而提高了土壤真菌群落结构的稳定性。有研究表明[40—41], 长期采用有机无机肥配施真菌群落以子囊菌门、担子菌门与接合菌门为主;在本研究中, 子囊菌门、担子菌门与被孢霉门为优势门, 该结论和之前报道一致[42]。宋秀丽等[39]研究得出, 和旋耕、深松不覆盖的处理相比, 深松与覆盖处理之后有助于子囊菌门相对丰度大幅增加。靳玉婷等[43]在水稻-油菜轮作发现秸秆还田配施化肥土壤中子囊菌门和担子菌门真菌群落的相对丰度提高。本试验中, 子囊菌门的相对丰度以秸秆覆盖不施氮处理最高和覆盖施氮处理最低, 覆盖处理降低被孢霉门的相对丰度, 担子菌门的相对丰度增加, 说明担子菌门更适宜秸秆覆盖提供的生长环境。有研究表明[44], 担子菌门是植物凋落物顽固性成分的主要分解者。因此, 在秸秆覆盖条件下担子菌门的相对丰度增加更有利于秸秆的分解。前人研究表明[15, 45], 秸秆还田显著增加弯孢菌属、镰刀菌属、被孢霉属的相对丰度;而本研究中, 秸秆覆盖增加了光盖伞属、弯孢菌属和黑孢霉属的相对丰度。光盖伞属、弯孢菌属和黑孢霉属具有降解木质素和纤维素的功能[46—48], 说明此类真菌菌属在秸秆覆盖下相对丰度增加利于秸秆分解, 促进养分释放。
微生物群落的多样性与结构变化可较为理想地判断土壤质量变化[49]。土壤pH值、有机碳、速效磷、速效钾等单个或多个因素均会影响真菌群落结构变化[12, 16, 50]。本研究中土壤SOC、AK、AP、AN、TN和NH4+-N含量是影响石灰性紫色土真菌群落结构变异的主要因素, 主要是秸秆覆盖还田增加了土壤碳、氮、钾养分的含量, 促进磷的有效性提高[4], 为真菌生长提供重要的碳源与能量, 影响其结构与多样性。此外, 本研究中秸秆覆盖配施氮肥条件光盖伞属、柱霉属和弯孢菌属与小麦产量增加相关, 从而提高小麦生产力。
4 结论覆盖与氮肥配施有利于提高土壤SOC、TN、AN、NH4+-N、AP和AK含量, 促进土壤真菌群落结构的优化, 增加了子囊菌门和担子菌门的相对丰度及光盖伞属、弯孢菌属和黑孢霉属的相对丰度。土壤SOC、AK、AP、AN、TN和NH4+-N含量是促进土壤真菌群落结构改变的主要环境因子。光盖伞属、柱霉属和弯孢菌属在秸秆覆盖配施氮肥条件下是提高小麦产量的主要真菌菌群。秸秆覆盖配施氮肥能提高土壤养分量, 调控微生物的结构组成和多样性, 对四川丘陵旱地石灰性紫色土培肥地力和提高作物生产力具有重要意义。
| [1] |
郑勇, 高勇生, 张丽梅, 何园球, 贺纪正. 长期施肥对旱地红壤微生物和酶活性的影响. 植物营养与肥料学报, 2008, 14(2): 316-321. |
| [2] |
徐万里, 唐光木, 葛春辉, 王西和, 刘骅. 长期施肥对新疆灰漠土土壤微生物群落结构与功能多样性的影响. 生态学报, 2015, 35(2): 468-477. |
| [3] |
王小玲, 马琨, 伏云珍, 汪志琴, 安嫄嫄. 免耕覆盖及有机肥施用对土壤真菌群落组成及多样性的影响. 应用生态学报, 2020, 31(3): 890-898. |
| [4] |
Chen S H, Gao R C, Xiang X L, Yang H K, Ma H L, Zheng T, Xiao Y, Zhang X, Li H, Fan G Q, Yu Y. Straw mulching and nitrogen application altered ammonia oxidizers communities and improved soil quality in the alkaline purple soil of Southwest China. AMB Express, 2021, 11(1): 52. DOI:10.1186/s13568-021-01211-x |
| [5] |
Peršoh D. Plant-associated fungal communities in the light of meta'omics. Fungal Diversity, 2015, 75: 1-25. DOI:10.1007/s13225-015-0334-9 |
| [6] |
Dhaliwal S S, Naresh R K, Gupta R K, Panwar A S, Mahajan N C, Singh R, Mandal A. Effect of tillage and straw return on carbon footprints, soil organic carbon fractions and soil microbial community in different textured soils under rice-wheat rotation: a review. Reviews in Environmental Science and Biotechnology, 2020, 19: 103-115. DOI:10.1007/s11157-019-09520-1 |
| [7] |
董亮, 田慎重, 王学君, 孙泽强, 郑东峰, 刘盛林, 董晓霞, 郭洪海, 罗加法. 秸秆还田对土壤养分及土壤微生物数量的影响. 中国农学通报, 2017, 33(11): 77-80. |
| [8] |
Hao M M, Hu H Y, Liu Z, Dong Q L, Sun K, Feng Y P, Li G, Ning T Y. Shifts in microbial community and carbon sequestration in farmland soil under long-term conservation tillage and straw returning. Applied Soil Ecology, 2019, 136: 43-54. DOI:10.1016/j.apsoil.2018.12.016 |
| [9] |
Su Y, Lv J L, Yu M, Ma Z H, Xi H, Kou C L, He Z C, Shen A L. Long-term decomposed straw return positively affects the soil microbial community. Journal of Applied Microbiology, 2020, 128(1): 138-150. DOI:10.1111/jam.14435 |
| [10] |
Zhao S C, Qiu S J, Xu X P, Ciampitti I A, Zhang S Q, He P. Change in straw decomposition rate and soil microbial community composition after straw addition in different long-term fertilization soils. Applied Soil Ecology, 2019, 138: 123-133. DOI:10.1016/j.apsoil.2019.02.018 |
| [11] |
Nair A, Ngouajio M. Soil microbial biomass, functional microbial diversity, and nematode community structure as affected by cover crops and compost in an organic vegetable production system. Applied Soil Ecology, 2012, 58: 45-55. DOI:10.1016/j.apsoil.2012.03.008 |
| [12] |
李鹏, 李永春, 史加亮, 郑宪清, 武国干, 蒋玮, 赵凯, 明凤, 潘爱虎, 吕卫光, 唐雪明. 水稻秸秆还田时间对土壤真菌群落结构的影响. 生态学报, 2017, 37(13): 4309-4317. |
| [13] |
张翰林, 白娜玲, 郑宪清, 李双喜, 张娟琴, 张海韵, 周胜, 孙会峰, 吕卫光. 秸秆还田与施肥方式对稻麦轮作土壤细菌和真菌群落结构与多样性的影响. 中国生态农业学报(中英文), 2021, 29(3): 531-539. |
| [14] |
辛励, 陈延玲, 刘树堂, 刘锦涛, 袁铭章, 南镇武. 长期定位秸秆还田对土壤真菌群落的影响. 华北农学报, 2016, 31(5): 186-192. |
| [15] |
李月, 张水清, 韩燕来, 李世莹, 王祎, 李慧, 李芳, 谭金芳. 长期秸秆还田与施肥对潮土酶活性和真菌群落的影响. 生态环境学报, 2020, 29(7): 1359-1366. |
| [16] |
李红宇, 王志君, 范名宇, 刘梦红, 吕艳东, 刘丽华. 秸秆连续还田对苏打盐碱水稻土养分及真菌群落的影响. 干旱地区农业研究, 2021, 39(2): 15-23. |
| [17] |
Paungfoo-Lonhienne C, Yeoh Y K, Kasinadhuni N R P, Lonhienne T G A, Robinson N, Hugenholtz P, Ragan M A, Schmidt S. Nitrogen fertilizer dose alters fungal communities in sugarcane soil and rhizosphere. Scientific Reports, 2015, 5: 8678. |
| [18] |
Edwards I P, Zak D R, Keliner H, Eisenlord S D, Pregitzer K S. Simulated atmospheric N deposition alters fungal community composition and suppresses ligninolytic gene expression in a northern hardwood forest. PLoS One, 2011, 6(6): e20421. |
| [19] |
郭梨锦, 曹凑贵, 张枝盛, 刘天奇, 李成芳. 耕作方式和秸秆还田对稻田表层土壤微生物群落的短期影响. 农业环境科学学报, 2013, 32(8): 1577-1584. |
| [20] |
沈吉成, 李玲玲, 谢军红, 彭正凯, Eunice E, 邓超超, 王进斌, 颉健辉. 耕作措施对陇中麦豆轮作系统土壤真菌群落的影响. 中国土壤与肥料, 2019(6): 94-98, 153-153. |
| [21] |
丁建莉, 姜昕, 马鸣超, 关大伟, 赵百锁, 魏丹, 曹凤明, 李力, 李俊. 长期有机无机肥配施对东北黑土真菌群落结构的影响. 植物营养与肥料学报, 2017, 23(4): 914-923. |
| [22] |
Chen Q Y, Liu Z J, Zhou J B, Xu X P, Zhu Y J. Long-term straw mulching with nitrogen fertilization increases nutrient and microbial determinants of soil quality in a maize-wheat rotation on China's Loess Plateau. Science of the Total Environment, 2021, 775: 145930. |
| [23] |
Chen W X, Johnson D W, Fu S L. Rhizosphere effects on decomposition: controls of plant species, phenology, and fertilization. Soil Science Society of America Journal, 2003, 67(5): 1418-1427. |
| [24] |
鲍士旦. 土壤农化分析 (第三版). 北京: 中国农业出版社, 2000.
|
| [25] |
Chen S H, Xiang X L, Ma H L, Penttinen P, Zhao J R, Li H, Gao R C, Zheng T, Fan G Q. Straw mulching and nitrogen fertilization affect diazotroph communities in wheat rhizosphere. Frontiers in Microbiology, 2021, 12: 658668. |
| [26] |
Liu X Q, Herbert S J, Hashemi A M, Zhang X, Ding G. Effects of agricultural management on soil organic matter and carbon transformation-a review. Plant, Soil and Environment, 2006, 52(12): 531-543. |
| [27] |
Akhtar K, Wang W Y, Ren G X, Khan A, Feng Y Z, Yang G H, Wang H Y. Integrated use of straw mulch with nitrogen fertilizer improves soil functionality and soybean production. Environment International, 2019, 132: 105092. |
| [28] |
Wang L, Yuan X L, Liu C, Li Z G, Chen F, Li S Q, Wu L H, Liu Y. Soil C and N dynamics and hydrological processes in a maize-wheat rotation field subjected to different tillage and straw management practices. Agriculture, Ecosystems & Environment, 2019, 285: 106616. |
| [29] |
Xu C H, Xu X, Ju C H, Chen H Y H, Wilsey B J, Luo Y Q, Fan W. Long-term, amplified responses of soil organic carbon to nitrogen addition worldwide. Global Change Biology, 2021, 27(6): 1170-1180. |
| [30] |
Blanco-Canqui H, Claassen M M, Presley D R. Summer cover crops fix nitrogen, increase crop yield, and improve soil-crop relationships. Agronomy Journal, 2012, 104(1): 137-147. |
| [31] |
Dong Q, Dang T H, Guo S L, Hao M D. Effects of mulching measures on soil moisture and N leaching potential in a spring maize planting system in the southern Loess Plateau. Agricultural Water Management, 2019, 213: 803-808. |
| [32] |
Liang W X, Zhang Z C, Wen X X, Liao Y C, Liu Y. Effect of non-structural carbohydrate accumulation in the stem pre-anthesis on grain filling of wheat inferior grain. Field Crops Research, 2017, 211: 66-76. |
| [33] |
赵小军, 李志洪, 刘龙, 崔婷婷. 种还分离模式下玉米秸秆还田对土壤磷有效性及其有机磷形态的影响. 水土保持学报, 2017, 31(1): 243-247. |
| [34] |
Yan C, Zhan H Q, Yan S S, Dong S K, Ma C M, Song Q L, Gong Z P, Barbie M. Effects of straw retention and phosphorous fertilizer application on available phosphorus content in the soil solution during rice growth. Paddy and Water Environment, 2016, 14: 61-69. |
| [35] |
Li H, Huang G, Meng Q, Ma L, Yuan L, Wang F, Zhang W, Cui Z, Shen J, Chen X, Jiang R, Zhang F. Integrated soil and plant phosphorus management for crop and environment in China. A review. Plant and Soil, 2011, 349(1/2): 157-167. |
| [36] |
宋大利, 侯胜鹏, 王秀斌, 梁国庆, 周卫. 中国秸秆养分资源数量及替代化肥潜力. 植物营养与肥料学报, 2018, 24(1): 1-21. |
| [37] |
Wu T H. Can ectomycorrhizal fungi circumvent the nitrogen mineralization for plant nutrition in temperate forest ecosystems?. Soil Biology and Biochemistry, 2011, 43(6): 1109-1117. |
| [38] |
李彤, 王梓廷, 刘露, 廖允成, 刘杨, 韩娟. 保护性耕作对西北旱区土壤微生物空间分布及土壤理化性质的影响. 中国农业科学, 2017, 50(5): 859-870. |
| [39] |
宋秀丽, 盘雨薇, 孙士明, 王俊河, 靳晓燕, 庞爱国, 于晓波, 金新月, 郝天乐, 宋翔雨, 杨德光. 真菌群落多样性及组成结构对不同耕作模式的响应机制. 玉米科学, 2021: 1-13. (2021-01-09). https://kns.cnki.net/KCMS/detail/detail.aspx?dbcode=CJFQ&dbname=CAPJLAST&filename=YMKX2021040900I.
|
| [40] |
吴宪, 王蕊, 胡菏, 修伟明, 李刚, 赵建宁, 杨殿林, 王丽丽, 王欣奕. 潮土细菌及真菌群落对化肥减量配施有机肥和秸秆的响应. 环境科学, 2020, 41(10): 4669-4681. |
| [41] |
Ma A Z, Zhuang X L, Wu J M, Cui M M, Lv D, Liu C Z, Zhuang G Q. Ascomycota members dominate fungal communities during straw residue decomposition in arable soil. PLoS One, 2013, 8(6): e66146. |
| [42] |
萨如拉, 杨恒山, 邰继承, 高聚林, 李媛媛. 秸秆还田条件下腐熟剂对不同质地土壤真菌多样性的影响. 中国生态农业学报(中英文), 2020, 28(7): 1061-1071. |
| [43] |
靳玉婷, 李先藩, 蔡影, 胡宏祥, 刘运峰, 付思伟, 张博睿. 秸秆还田配施化肥对稻-油轮作土壤酶活性及微生物群落结构的影响. 环境科学, 2021, 42(8): 3985-3996. |
| [44] |
Lynch M D J, Thorn R G. Diversity of basidiomycetes in Michigan agricultural soils. Applied and Environmental Microbiology, 2006, 72(11): 7050-7056. |
| [45] |
Wicklow D T, Roth S, Deyrup S T, Gloer J B. A protective endophyte of maize: Acremonium zeae antibiotics inhibitory to Aspergillus flavus and Fusarium verticillioides. Mycological Research, 2005, 109(Pt 5): 610-618. |
| [46] |
Capelari M, Zadrazil F. Lignin degradation and In vitro digestibility of wheat straw treated with Brazilian tropical species of white rot fungi. Folia Microbiologica, 1997, 42(5): 481-487. |
| [47] |
Melo M D, Araujo A C A, Chogi M A N, Duarte I C S. Cellulolytic and lipolytic fungi isolated from soil and leaf litter samples from the cerrado (Brazilian Savanna). Revista De Biologia Tropical, 2018, 66(1): 237-245. |
| [48] |
Devi R, Kaur T, Kour D, Rana K L, Yadav A, Yadav A N. Beneficial fungal communities from different habitats and their roles in plant growth promotion and soil health. Microbial Biosystems, 2020, 5(1): 21-47. |
| [49] |
Bai N L, Zhang H L, Li S X, Zheng X Q, Zhang J Q, Zhang H Y, Zhou S, Sun H F, Lv W G. Long-term effects of straw and straw-derived biochar on soil aggregation and fungal community in a rice-wheat rotation system. PeerJ, 2019, 6: e6171. |
| [50] |
代红翠, 张慧, 薛艳芳, 高英波, 钱欣, 赵海军, 成浩, 李宗新, 刘开昌. 不同耕作和秸秆还田下褐土真菌群落变化特征. 中国农业科学, 2019, 52(13): 2280-2294. |