 2022, Vol. 42
2022, Vol. 42文章信息
- 华咏乐, 谢燕锦, 殷宝法, 魏万红
- HUA Yongle, XIE Yanjin, YIN Baofa, WEI Wanhong
- 内蒙古草原布氏田鼠与绵羊的营养生态位研究
- Trophic niches of Brandt's vole (Lasiopodomys brandtii) and sheep (Ovis aries) in the Inner Mongolia Grassland
- 生态学报. 2022, 42(21): 8618-8627
- Acta Ecologica Sinica. 2022, 42(21): 8618-8627
- http://dx.doi.org/10.5846/stxb202109142578
-
文章历史
- 收稿日期: 2021-09-14
- 网络出版日期: 2022-06-21

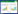




2. 江苏省动物重要疫病与人兽共患病防控协同创新中心, 扬州 225009
2. Jiangsu Co-Innovation Center for Prevention and Control of Important Animal Infectious Diseases and Zoonoses, Yangzhou 225009, China
布氏田鼠(Lasiopodomys brandtii)是内蒙古典型草原的啮齿动物之一[1], 在维持草原生物多样性和生态系统功能方面发挥着重要作用[2—6]。然而, 当种群数量过多时, 其啃食及挖掘活动破坏了优良牧草, 导致草地生态系统日趋退化[7—8]。因此, 确定布氏田鼠种群数量动态的变化机制及影响因素对于合理管理其种群数量、维护草地生态系统稳定具有非常重要的意义。
放牧是人类利用草原的主要活动方式之一。相关研究结果表明, 家畜的践踏、采食能够降低植物的高度和植被的覆盖度[6, 9—10], 为喜栖低矮稀疏环境的布氏田鼠提供了有利的栖息条件[8], 促进其种群数量增加, 认为过度放牧可能是造成布氏田鼠种群数量爆发的主要因素[11—12]。也有学者认为, 放牧能降低植被的高度和盖度, 破坏啮齿动物的洞穴系统, 从而增加啮齿类被天敌捕食的风险[13—14];同时, 放牧活动也增加了与啮齿动物的食物资源竞争强度, 进而对其种群数量产生负面效应[15]。由于这些工作缺乏严格的野外控制实验, 因此, 不同的学者所得到的结果各异。
植物为动物提供食物, 因此植物多样性的变化直接影响动物食物的丰富度[16];反之动物的选择性采食也会影响植物群落的物种组成、物种多样性及生产力[17—19], 这种变化可能从食物资源方面改变食草动物间的关系。当植物资源丰富时, 动物面临的食物竞争压力小, 会因营养生态位的分离而降低种间竞争[20—22];当植物多样性较低时, 食草动物间营养生态位重叠度增加, 最终导致激烈的种间竞争[23—24]。内蒙古草原同时承受着放牧和布氏田鼠的干扰, 它们的食性及生态位关系影响着小哺乳动物的种群数量和群落结构, 也影响着绵羊的食物资源, 但二者之间的营养生态位分化以及时间格局鲜有报道。因此, 本研究利用大型野外围栏放牧控制实验, 采用粪便显微分析法测定布氏田鼠和绵羊的食谱组成及季节性变化, 分析两种食草动物间的生态位重叠与分离, 探讨绵羊对布氏田鼠食性选择的影响以及二者之间竞争与共存关系, 以回答2个问题:(1)植物资源的变化是否影响布氏田鼠和绵羊的食性选择?(2)营养生态位的重叠是否引起绵羊和布氏田鼠间的食物竞争、并对布氏田鼠种群数量产生影响?
1 研究地点本研究在内蒙古锡林浩特市毛登牧场草原动物研究站(44.11°N, 116.27°E)建立的大型野外围栏中进行。实验区气候属于温带半干旱大陆性气候, 春季温暖, 冬季寒冷;年平均温度为2 ℃, 气温日较差大, 年降水量约241 mm, 75%的降水集中在生长季(5—8月)。实验区内主要的植物优势种有羊草(Leymus chinensis)、克氏针茅(Stipa krylovii)和糙隐子草(Cleistogenes squarrosa)等, 植物覆盖度约50%。
2 材料与方法 2.1 围栏设计研究期间, 共建立了24个60 m×80 m的大型铁皮围栏, 镀锌铁皮深入到地下1 m, 高出地上1.4 m, 以防止布氏田鼠、绵羊和其他动物进出围栏, 同时围栏上方和四周都覆盖尼龙网(网眼为10 cm), 以有效阻隔猫头鹰等鸟类的捕食作用。本实验随机选择8个围栏作为田鼠处理和放牧+田鼠处理, 每种处理4个围栏。5月初清空围栏内的所有动物, 每个围栏中分别放入13对年龄、大小相近的布氏田鼠作为建群种;根据当地绵羊放牧强度标准[25], 设置为40只绵羊每两周放牧一天, 该放牧强度为适度放牧。
2.2 植物群落结构调查及粪便样品收集2012年6—9月, 每月在每个围栏中随机设置5个调查样带, 采用1 m×1 m的样方框调查统计每种植物的株数和覆盖度, 并分种收集0.3 m×0.3 m的地上植物部分, 带回实验室于55 ℃烘箱中烘干至恒重, 称量作为该植物的地上生物量。此外, 采集围栏出现的所有植物种类放入采集袋中, 带到实验室内放在55 ℃烘箱烘48 h至恒重, 作为组织显微分析的对照样本。
每月采用标志重捕法调查每个围栏内布氏田鼠的数量, 同时收集活捕笼内田鼠新排出的粪便, 根据标记流放的记录对粪便样品进行编号。绵羊摄入食物后, 食物中绝大多数不能消化的部分将在24 h后排出体外[26—27], 每月进行放牧处理时, 在每个围栏随机选10只绵羊, 晚上归牧24 h后利用集粪袋收集绵羊的粪便, 每月2次。非放牧时间绵羊在围栏附近草地活动。根据野外重捕记录选取布氏田鼠成体(每个围栏每月10份, 雌雄各半)和绵羊的粪便放在55 ℃烘箱烘48 h至恒重, 以备下一步的组织显微分析。
2.3 显微制片及镜检将烘干的植物样品按种粉碎, 然后按照殷宝法等[28]的方法制备植物对照装片。粪便显微装片的制备方法同植物参考装片。每份粪便样品制作2张显微装片。粪样、对照植物显微装片都在10倍物镜(放大100倍)下镜检, 根据对照植物表皮细胞的形状结构及排列方式、导管纤维的类型等特征, 确定视野中粪样残留碎片的植物种类。每张粪便装片随机选择10个互不重叠的视野, 分种记录每个视野中出现的可辨认植物碎片种类及数量, 将10个视野中的同种植物碎片数量进行累加, 作为一个统计样本。
2.4 数据处理与分析 2.4.1 植物资源的相对丰富度的计算参考蒋志刚等[29]的公式计算每种植物资源的相对丰富度(RAi):
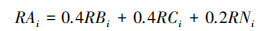
|
式中, RBi、RCi、RNi分别为种i的相对生物量、相对盖度和相对株数, 其计算公式分别为:
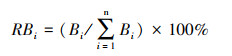
|
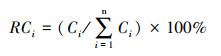
|
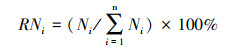
|
式中,Bi、Ci、Ni为种i的平均生物量、平均盖度、平均株数。
本研究中, 将相对丰富度超过5%的植物定义为围栏中优势植物。
2.4.2 动物对植物的利用频率的计算将镜检下粪便中各植物种类的相对密度(RDi)作为动物对植物i的利用频率, 其计算公式如下[30]:

|
本研究中, 将利用频率超过10%的植物定义为主要食物。
2.4.3 选择性指数的计算应用选择性指数公式[31—32]计算布氏田鼠和绵羊对种i的选择性指数:
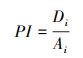
|
式中,PI为选择性指数, 与动物的喜食程度正相关, Di为植物i在食物干重中所占的比例, Ai为该种植物在植被丰富度中所占的比例。PI>1.0说明该种植物为动物的喜食植物[7]。
2.4.4 多样性的计算应用Shannon-Wiener指数计算植物多样性和动物食物多样性[32]:
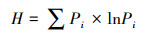
|
式中, Pi为物种i的相对生物量。
2.4.5 生态位宽度和生态位重叠度的计算参考殷宝法等[28]的公式计算食草动物在食物利用维上的生态位宽度:
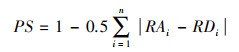
|
式中, RAi和RDi分别为i资源位上的资源相对丰富度和被利用频率。
生态位重叠用Smith[33]推荐的生态位重叠公式计算:
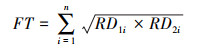
|
式中, FT为生态位重叠度, RD1i和RD2i分别为种1、种2在i资源位上的利用频率。
2.4.6 统计分析利用软件R 3.5.3中的线性混合模型(LMMS)分析放牧和月份及其交互作用对布氏田鼠喜食植物种数以及禾本植物生物量的影响, 其中把放牧处理、月份处理作为固定效应, 把围栏作为随机效应, 若存在交互作用, 则用单因素方差分析进行两两比较。利用Graphpad Prism 9对围栏植物多样性和两种动物食物多样性进行相关性分析并画图。文中实验数值以平均数±标准误差(Mean ± SE)表示, 以P<0.05作为差异显著的标准。
3 结果及分析 3.1 围栏中的植物组成布氏田鼠围栏中共发现18种植物(表 1), 其中, 8月份植物种数最多;糙隐子草和克氏针茅的相对丰富度在4个月均超过20%, 皱叶糙苏、刺藜和羊草(除7月)均超过5%, 为围栏中的优势植物。
| 植物种 Plant species |
6月 June | 7月 July | 8月 August | 9月 September | |||||||
| 田鼠 Vole |
放牧+田鼠 Grazing+Vole |
田鼠 Vole |
放牧+田鼠 Grazing+Vole |
田鼠 Vole |
放牧+田鼠Grazing+Vole | 田鼠 Vole |
放牧+田鼠 Grazing+Vole |
||||
| 糙隐子草 Cleistogenes squarrosa |
34.79±6.12 | 42.73±6.12 | 43.34±5.96 | 29.63±5.55 | 38.29±5.23 | 29.02±4.75 | 39.56±6.30 | 28.65±5.40 | |||
| 克氏针茅 Stipa krylovii |
22.53±4.60 | 17.19±5.06 | 24.41±5.78 | 15.89±4.74 | 21.23±4.60 | 13.17±4.37 | 25.58±6.38 | 17.13±5.03 | |||
| 皱叶糙苏 Paraphlomis lanceolata |
13.42±4.43 | 3.22±1.83 | 8.42±2.91 | 0.47±0.33 | 9.91±3.47 | 0.46±0.23 | 9.49±3.39 | 0.71±0.39 | |||
| 刺藜 Dysphania aristata |
10.70±1.30 | 11.55±2.15 | 11.21±1.53 | 29.24±4.14 | 12.05±1.69 | 41.98±4.58 | 5.44±0.96 | 34.88±5.54 | |||
| 羊草 Leymus chinensis |
6.62±2.93 | 11.15±2.89 | 1.56±0.49 | 14.54±4.19 | 6.32±3.15 | 7.64±3.56 | 8.34±3.97 | 10.03±2.57 | |||
| 狗尾巴草 Setaria viridis |
4.50±0.84 | 3.09±0.43 | 0.42±0.16 | 1.36±0.38 | 0.35±0.12 | 2.41±0.94 | 0.41±0.31 | 0.77±0.22 | |||
| 松叶猪毛菜 Salsola laricifolia |
1.57±0.78 | 0.35±0.19 | 2.13±0.78 | 0.49±0.31 | 3.13±1.65 | 0.43±0.18 | 4.22±1.74 | 0.32±0.16 | |||
| 瓣蕊唐松草 Thalictrum petaloideum |
1.41±0.64 | 0.18±0.13 | 0.69±0.41 | 0.24±0.16 | 0.27±0.11 | 0.03±0.03 | - | - | |||
| 灰藜 Chenopodium album |
1.06±0.45 | 2.30±0.81 | 2.20±1.36 | 1.42±0.56 | 1.70±1.27 | 0.73±0.27 | 2.30±1.62 | 1.80±0.89 | |||
| 阿氏旋花 Convolvulus ammannii |
0.98±0.39 | 2.57±2.20 | 0.25±0.14 | 0.32±0.21 | 0.86±0.47 | 0.04±0.04 | - | 0.03±0.03 | |||
| 蒙古猪毛菜 Salsola ikonnikovii |
0.89±0.39 | 0.10±0.09 | 4.51±2.55 | 0.09±0.09 | 4.44±2.57 | 0.06±0.06 | 2.32±1.28 | - | |||
| 苔草 Carex tristachya |
0.69±0.38 | 4.02±1.40 | 0.77±0.43 | 3.35±1.89 | 0.67±0.26 | 1.38±0.57 | 1.10±0.46 | 1.29±0.55 | |||
| 栉叶蒿 Neopallasia pectinata |
0.43±0.22 | 0.93±0.36 | - | 1.15±0.65 | 0.30±0.21 | 0.32±0.13 | 0.14±0.14 | 0.26±0.10 | |||
| 紫花苜蓿 Medicago sativa |
0.41±0.19 | 0.20±0.11 | - | 0.03±0.03 | 0.07±0.05 | 0.11±0.05 | 0.03±0.03 | 0.05±0.05 | |||
| 独行菜 Lepidium apetalum |
- | 0.31±0.23 | 0.08±0.05 | 0.61±0.16 | 0.03±0.03 | 0.14±0.06 | 0.37±0.34 | 0.15±0.07 | |||
| 打碗花 Calystegia hederacea |
- | 0.10±0.10 | - | - | - | - | - | - | |||
| 乳白黄芪 Astragalus galactites |
- | - | 0.02±0.02 | - | 0.13±0.07 | - | - | 0.08±0.06 | |||
| 画眉草 Eragrostis pilosa |
- | - | - | 0.97±0.60 | 0.01±0.01 | 1.64±0.65 | 0.70±0.70 | 3.81±1.94 | |||
| 大籽蒿 Artemisia sieversiana |
- | - | - | 0.16±0.09 | - | 0.11±0.06 | - | - | |||
| 老鹳草 Geranium wilfordii |
- | - | - | 0.05±0.05 | 0.25±0.18 | - | - | - | |||
| 虎尾草 Chloris virgata |
- | - | - | - | - | 0.26±0.26 | - | - | |||
| 车前 Plantago asiatica |
- | - | - | - | - | 0.06±0.04 | - | - | |||
| 细叶鸢尾 Iris tenuifolia |
- | - | - | - | - | - | - | 0.04±0.04 | |||
| “-”表示研究区域内未调查到这种植物 | |||||||||||
放牧+布氏田鼠围栏中共发现23种植物(表 1), 其中, 8月份植物种数最多;糙隐子草的相对丰富度在4个月均超过20%, 克氏针茅、刺藜、羊草(除8月)均超过10%, 为围栏中的优势植物。
3.2 布氏田鼠的食性选择研究期间, 布氏田鼠共采食围栏内的10种植物(表 2);其中羊草、克氏针茅、糙隐子草为其主要采食植物, 占总食物的比例超过90%。不同月份下布氏田鼠的采食植物种数不同, 尤其在植物资源量降低的9月, 布氏田鼠的采食物种数最高(8种)。
| 植物种 Plant species |
6月 June | 7月 July | 8月 August | 9月 September | |||||||
| 田鼠 Vole |
放牧+田鼠 Grazing+Vole |
田鼠 Vole |
放牧+田鼠 Grazing+Vole |
田鼠 Vole |
放牧+田鼠 Grazing+Vole |
田鼠 Vole |
放牧+田鼠 Grazing+Vole |
||||
| 羊草 L. chinensis | 62.56±1.59 | 62.55±1.53 | 60.18±1.39 | 58.70±1.56 | 8.60±1.04 | 9.78±1.17 | 25.21±1.69 | 28.23±1.78 | |||
| 克氏针茅 S. krylovii | 25.27±2.08 | 24.45±1.52 | 22.29±1.66 | 18.47±1.63 | 68.62±1.58 | 70.84±1.47 | 65.13±1.95 | 61.48±2.50 | |||
| 糙隐子草 C. squarrosa | 9.03±1.67 | 12.25±1.57 | 14.73±1.69 | 18.15±1.71 | 14.70±1.54 | 12.86±1.62 | 1.11±0.48 | 1.93±0.58 | |||
| 皱叶糙苏 P. lanceolata | 1.28±0.45 | - | 0.90±0.38 | 1.14±0.40 | 1.99±0.68 | 1.27±0.49 | 3.20±0.79 | 0.44±0.30 | |||
| 狗尾巴草 S. viridis | 1.03±0.40 | 0.19±0.19 | - | - | - | - | - | - | |||
| 苔草 C. tristachya | 0.84±0.43 | 0.56±0.32 | 1.69±0.61 | 3.55±0.68 | 5.74±0.85 | 4.85±0.85 | 3.11±0.68 | 5.45±0.96 | |||
| 栉叶蒿 N. pectinata | - | - | - | - | 0.35±0.24 | 0.17±0.17 | - | 1.48±0.48 | |||
| 刺藜 D. aristata | - | - | - | - | - | - | 1.78±0.66 | 0.64±0.36 | |||
| 紫花苜蓿 M. sativa | - | - | - | - | - | 0.23±0.23 | 0.23±0.23 | 0.19±0.19 | |||
| 蒙古猪毛菜 S. ikonnikovii | - | - | - | - | - | - | 0.23±0.23 | - | |||
布氏田鼠在不同月份的喜食植物不同(表 3), 6月份喜食羊草和克氏针茅, 7月份喜食羊草和苔草, 8月份喜食羊草、克氏针茅和苔草, 9月份喜食羊草、克氏针茅、苔草以及紫花苜蓿。
| 植物种 Plant species |
6月 June | 7月 July | 8月 August | 9月 September | |||||||
| 田鼠 Vole |
放牧+田鼠 Grazing+Vole |
田鼠 Vole |
放牧+田鼠 Grazing+Vole |
田鼠 Vole |
放牧+田鼠 Grazing+Vole |
田鼠 Vole |
放牧+田鼠 Grazing+Vole |
||||
| 羊草 L. chinensis | 9.45 | 5.61 | 38.58 | 4.04 | 1.36 | 1.28 | 3.02 | 2.81 | |||
| 克氏针茅 S. krylovii | 1.12 | 1.42 | 0.91 | 1.16 | 3.23 | 5.38 | 2.55 | 3.59 | |||
| 糙隐子草 C. squarrosa | 0.26 | 0.29 | 0.34 | 0.61 | 0.38 | 0.44 | 0.03 | 0.07 | |||
| 皱叶糙苏 P. lanceolata | 0.10 | 0 | 0.11 | 2.43 | 0.2 | 2.76 | 0.34 | 0.61 | |||
| 狗尾巴草 S. viridis | 0.23 | 0.06 | 0 | 0 | 0 | 0 | 0 | 0 | |||
| 苔草 C. tristachya | 1.22 | 0.14 | 2.20 | 1.06 | 8.58 | 3.51 | 2.81 | 4.23 | |||
| 栉叶蒿 N. pectinata | 0 | 0 | 0 | 0 | 1.14 | 0.52 | 0 | 5.68 | |||
| 刺藜 D. aristata | 0 | 0 | 0 | 0 | 0 | 0 | 0.33 | 0.02 | |||
| 紫花苜蓿 M. sativa | 0 | 0 | 0 | 0 | 0 | 2.10 | 7.03 | 3.97 | |||
| 蒙古猪毛菜 S. ikonnikovii | 0 | 0 | 0 | 0 | 0 | 0 | 0.10 | 0 | |||
研究期间, 绵羊共采食围栏内的8种植物(表 4)。绵羊的主要食物为羊草、克氏针茅、糙隐子草、苔草以及蒙古猪毛菜。绵羊6月份喜食羊草、克氏针茅、紫花苜蓿, 7月份喜食羊草、阿氏旋花、紫花苜蓿和蒙古猪毛菜, 8月份喜食羊草、糙隐子草、苔草、栉叶蒿, 9月份喜食其食谱中除糙隐子草以外的所有植物。
| 植物种 Plant species |
6月 June | 7月 July | 8月 August | 9月 September | |||||||
| 利用频率/% Foraging frequency |
选择性指数 Preference index |
利用频率/% Foraging frequency |
选择性指数 Preference index |
利用频率/% Foraging frequency |
选择性指数 Preference index |
利用频率/% Foraging frequency |
选择性指数 Preference index |
||||
| 羊草 L. chinensis | 68.12±1.99 | 6.11 | 62.11±1.88 | 4.27 | 33.96±2.19 | 4.45 | 26.42±1.94 | 2.63 | |||
| 克氏针茅 S. krylovii | 17.26±1.54 | 1.00 | 3.47±0.76 | 0.22 | 5.31±1.44 | 0.40 | 47.83±2.00 | 2.79 | |||
| 糙隐子草 C. squarrosa | 10.89±1.48 | 0.25 | 21.15±2.47 | 0.71 | 41.50±2.51 | 1.43 | 18.41±2.82 | 0.64 | |||
| 苔草 C. tristachya | 1.19±0.45 | 0.30 | 3.13±0.79 | 0.94 | 10.49±1.08 | 7.59 | 5.96±1.20 | 4.63 | |||
| 阿氏旋花 C. ammannii | 1.71±0.66 | 0.67 | 0.48±0.33 | 1.50 | - | 0 | 0.28±0.28 | 9.21 | |||
| 紫花苜蓿 M. sativa | 0.83±0.48 | 4.08 | 0.81±0.39 | 24.02 | - | 0 | 1.11±0.53 | 22.81 | |||
| 蒙古猪毛菜 S. ikonnikovii | - | 0 | 16.88±1.08 | 189.52 | - | 0 | - | 0 | |||
| 栉叶蒿 N. pectinata | - | 0 | - | 0 | 8.75±1.61 | 27.42 | - | 0 | |||
分析围栏内植物多样性与绵羊和布氏田鼠的食物多样性之间的关系, 6月、8月和9月的植物多样性与两种食草动物的食物多样性显著正相关, 但在7月无显著相关关系(图 1)。

|
| 图 1 布氏田鼠和绵羊食物多样性与围栏植物多样性之间的关系 Fig. 1 Relationships between plant diversity and dietary diversity of Brandt′s vole and sheep |
放牧显著降低了围栏内禾本植物的生物量(F=19.199, P < 0.001; 图 2), 禾本科生物量在不同月份存在显著差异(F=28.889, P < 0.001), 8月份最高。放牧和月份对禾本植物的生物量没有显著的交互作用(F=1.211, P=0.308)。

|
| 图 2 放牧和不同月份处理对禾本植物地上生物量(g/m2)的影响 Fig. 2 Effects of grazing and different months on aboveground biomass(g/m2) of plants from Gramineae family |
放牧显著提高了布氏田鼠的喜食植物种数(F=13.170, P < 0.001; 图 3), 放牧和月份对布氏田鼠的喜食物种数有显著的交互作用(F=3.122, P=0.026), 与6月的布氏田鼠围栏相比, 7月(P=0.011)和8月(P=0.049)的放牧处理显著增加了布氏田鼠喜食植物种数;与7月的布氏田鼠围栏相比, 7月(P < 0.001)、8月(P=0.006)和9月(P=0.049)的放牧处理也显著增加了布氏田鼠喜食植物种数。

|
| 图 3 放牧和不同月份处理对布氏田鼠喜食物种数的影响 Fig. 3 Effect of grazing and different months on the number of plant species favored by Brandt′s vole 不同小写字母表示组内差异性显著(P < 0.05) |
布氏田鼠处理和放牧+布氏田鼠处理下布氏田鼠的营养生态位宽度结果表明(图 4), 除9月外, 放牧+布氏田鼠处理下布氏田鼠的生态位宽度都大于布氏田鼠处理。

|
| 图 4 6—9月布氏田鼠和放牧+布氏田鼠处理下布氏田鼠的营养生态位宽度的变化 Fig. 4 Changes of trophic niche breadth of Brandt′s vole under Brandt′s vole and Grazing+Brandt′s vole treatment |
放牧+布氏田鼠围栏内, 绵羊和布氏田鼠的营养生态位重叠度均较高(图 5), 除了8月份, 其它月份的营养生态位重叠度都高于0.9, 说明在布氏田鼠和绵羊之间可能存在激烈的食物竞争。

|
| 图 5 6—9月放牧+布氏田鼠处理下布氏田鼠和绵羊营养生态位重叠度的变化 Fig. 5 Changes of trophic niche overlap values of Brandt′s vole and sheep under Grazing+Brandt′s vole treatment |
为了探究家畜采食对布氏田鼠食性选择和种群数量的影响, 本研究采用粪便显微分析法, 结合大型围栏放牧控制实验, 从食物资源利用的角度探讨了布氏田鼠和绵羊之间的竞争与共存关系。研究结果表明, 禾本科植物是布氏田鼠和绵羊的主要食物来源, 两种动物的食物多样性与植物物种多样性显著正相关, 其食性选择取决于食物资源的变化, 但具体的食性选择存在明显的季节性变化, 并且两者的喜食植物的种类存在一定的差异, 说明两者的营养生态位存在一定程度的分离, 这可能是它们能够在该地区长期共存的重要原因。然而, 布氏田鼠和绵羊之间存在较高的营养生态位重叠, 在6、7、9月份, 营养生态位重叠度都超过0.9。生态位重叠程度反映了不同物种利用资源的相似程度以及它们之间的潜在竞争程度[34—35], 根据竞争排斥原理[36], 这将不利于布氏田鼠种群数量的增长, 这与Li等[6]在该地区的研究结果相一致, 即持续的放牧活动能显著地降低布氏田鼠的种群数量。因此, 本研究支持食物竞争假说, 在家畜放牧压力下, 小型啮齿动物的数量会减少[37]。
一些研究表明, 食物的数量和质量是影响一些啮齿类动物种群动态的关键因素[38]。在本研究中, 放牧活动极显著地降低了围栏内禾本科植物的地上生物量(图 2)。禾本科植物是布氏田鼠主要的食物来源, 占其总食物的比例超过90%, 因此禾本科植物地上生物量的降低无疑会降低布氏田鼠食物的数量。草地群落中植物的多样性造成动物对其利用程度的差异, 使得不同食草动物具有不同食性选择策略, 进而又改变植物群落结构[39—40]。本研究中布氏田鼠和绵羊的食物多样性与食物资源多样性正相关, 这意味着群落中植物种数、均匀度等发生改变时, 它们的食谱也将改变, 这与汪诗平[32, 41]的研究结果一致。当植物资源丰富时, 草食性动物可以优先选择喜食植物;当植物资源短缺时会增加先前不喜食植物的采食强度[7]。本研究中, 放牧活动显著增加了布氏田鼠的喜食植物的物种数(图 3)和营养生态位宽度(图 4)。导致这一现象的原因可能是两者之间较高的营养生态位重叠度。绵羊的竞争性采食降低了布氏田鼠喜食植物的数量, 迫使布氏田鼠增加对一些不喜食植物的采食强度, 从而使一些不喜食植物变为喜食植物。王桂明等[1]的研究结果表明, 布氏田鼠喜食的植物种类较少, 具有明显的狭食性, 偏好蛋白质含量高的植物。因此, 放牧条件下, 布氏田鼠喜食植物的物种数增加会降低布氏田鼠的食物质量。Li等[6]在该地区的研究结果显示, 放牧活动提高了布氏田鼠食物中的次生化合物含量, 使其食物的适口性降低。这些研究结果表明, 放牧活动可能是通过降低布氏田鼠的食物数量和质量, 进而对布氏田鼠个体发育和种群数量产生负面效应。
布氏田鼠是许多肉食动物的主要食物资源, 在维持草原生物多样性和生态系统稳定等方面发挥着重要的作用[2—6]。本研究从食物资源利用的角度分析了放牧活动对布氏田鼠种群动态的影响途径, 为合理管理布氏田鼠种群提供一定的科学资料, 将来的研究需要进一步分析不同放牧强度对布氏田鼠食性选择和种群动态的影响。
| [1] |
王桂明, 周庆强, 钟文勤, 王广和. 布氏田鼠(Microtus brandti)的食性. 兽类学报, 1992, 12(1): 57-64. |
| [2] |
Campos C M, Campos V E, Giannoni S M, Rodríguez D, Albanese S, Cona M I. Role of small rodents in the seed dispersal process: Microcavia australis consuming Prosopis flexuosa fruits. Austral Ecology, 2017, 42(1): 113-119. |
| [3] |
Davidson A D, Detling J K, Brown J H. Ecological roles and conservation challenges of social, burrowing, herbivorous mammals in the world's grasslands. Frontiers in Ecology and the Environment, 2012, 10(9): 477-486. DOI:10.1890/110054 |
| [4] |
Valkó O, Tölgyesi C, Kelemen A, Bátori Z, Gallé R, Rádai Z, Bragina T M, Bragin Y A, Deák B. Steppe Marmot (Marmota bobak) as ecosystem engineer in arid steppes. Journal of Arid Environments, 2021, 184: 104244. DOI:10.1016/j.jaridenv.2020.104244 |
| [5] |
Qin Y, Chen J J, Yi S H. Plateau pikas burrowing activity accelerates ecosystem carbon emission from alpine grassland on the Qinghai-Tibetan Plateau. Ecological Engineering, 2015, 84: 287-291. |
| [6] |
Li G L, Yin B F, Wan X R, Wei W H, Wang G M, Krebs C J, Zhang Z B. Successive sheep grazing reduces population density of Brandt's voles in steppe grassland by altering food resources: a large manipulative experiment. Oecologia, 2016, 180(1): 149-159. DOI:10.1007/s00442-015-3455-7 |
| [7] |
刘贵河, 王国杰, 汪诗平, 韩建国, 宛新荣, 郝树广. 内蒙古荒漠草原主要草食动物食性及其营养生态位. 生态学报, 2013, 33(3): 856-866. |
| [8] |
内蒙古锡林郭勒盟卫生防疫站. 布氏田鼠的生态研究. 动物学报, 1975, 21(1): 30-39. |
| [9] |
蒯晓妍, 邢鹏飞, 张晓琳, 梁艳, 王常慧, 董宽虎. 短期放牧强度对半干旱草地植物群落多样性和生产力的影响. 草地学报, 2018, 26(6): 1283-1289. |
| [10] |
张宇, 阿斯娅·曼力克, 辛晓平, 张荟荟, 热娜·阿布都克力木, 闫瑞瑞, 郭美兰. 禁牧与放牧对新疆温性草原群落结构、生物量及牧草品质的影响. 草地学报, 2020, 28(3): 815-821. |
| [11] |
钟文勤, 周庆强, 王广和, 孙崇潞, 周丕义, 刘文智, 贾永平. 布氏田鼠鼠害生态治理方法的设计及其应用. 兽类学报, 1991, 11(3): 204-212. |
| [12] |
Davidson A D, Ponce E, Lightfoot D C, Fredrickson E L, Brown J H, Cruzado J, Brantley S L, Sierra-Corona R, List R, Toledo D, Ceballos G. Rapid response of a grassland ecosystem to an experimental manipulation of a keystone rodent and domestic livestock. Ecology, 2010, 91(11): 3189-3200. DOI:10.1890/09-1277.1 |
| [13] |
Steen H, Mysterud A, Austrheim G. Sheep grazing and rodent populations: evidence of negative interactions from a landscape scale experiment. Oecologia, 2005, 143(3): 357-364. |
| [14] |
Torre I, Díaz M, Martínez-Padilla J, Bonal R, Viñuela J, Fargallo J A. Cattle grazing, raptor abundance and small mammal communities in Mediterranean grasslands. Basic and Applied Ecology, 2007, 8(6): 565-575. |
| [15] |
Parsons E W R, Maron J L, Martin T E. Elk herbivory alters small mammal assemblages in high-elevation drainages. Journal of Animal Ecology, 2013, 82(2): 459-467. |
| [16] |
王德利, 王岭. 草食动物与草地植物多样性的互作关系研究进展. 草地学报, 2011, 19(4): 699-704. |
| [17] |
李文怀, 郑淑霞, 白永飞. 放牧强度和地形对内蒙古典型草原物种多度分布的影响. 植物生态学报, 2014, 38(2): 178-187. |
| [18] |
韩梦琪, 王忠武, 靳宇曦, 康静, 李江文, 王悦华, 王舒新, 韩国栋. 短花针茅荒漠草原物种多样性及生产力对长期不同放牧强度的响应. 西北植物学报, 2017, 37(11): 2273-2281. |
| [19] |
朱爱民, 韩国栋, 康静, 闫宝龙, 赵坤, 朱毅. 长期不同放牧强度下短花针茅荒漠草原物种多样性季节性动态变化. 草地学报, 2019, 27(4): 1013-1021. |
| [20] |
Lovari S, Pokheral C P, Jnawali S R, Fusani L, Ferretti F. Coexistence of the tiger and the common leopard in a prey-rich area: the role of prey partitioning. Journal of Zoology, 2015, 295(2): 122-131. |
| [21] |
Hofmann R R. Evolutionary steps of ecophysiological adaptation and diversification of ruminants: a comparative view of their digestive system. Oecologia, 1989, 78(4): 443-457. |
| [22] |
Chesson P. Mechanisms of maintenance of species diversity. Annual Review of Ecology and Systematics, 2000, 31(1): 343-366. |
| [23] |
岳闯, 郭乾伟, 张卓然, 李鑫, 满都呼, 袁帅, 付和平, 武晓东, 金国, 刘建文, 李永善. 内蒙古典型草原布氏田鼠营养生态位及其种间关系. 兽类学报, 2020, 40(5): 424-434. |
| [24] |
Zielinski W J, Duncan N P. Diets of sympatric populations of American martens (Martes americana) and fishers (Martes pennanti) in California. Journal of Mammalogy, 2004, 85(3): 470-477. |
| [25] |
Schönbach P, Wan H W, Gierus M, Bai Y F, Müller K, Lin L J, Susenbeth A, Taube F. Grassland responses to grazing: effects of grazing intensity and management system in an Inner Mongolian steppe ecosystem. Plant and Soil, 2011, 340(1): 103-115. |
| [26] |
达来, 敖特根, 占布拉, 小林裕志, 杉捕俊弘, 宇都牧生. 绵羊粪中排出的牧草种子及其发芽研究. 中国草地, 1994(5): 23-27. |
| [27] |
娜丽克斯·外里, 王树林, 赵刚, 杨洁晶, 吕艳萍, 鲁为华. 4种豆科牧草种子对绵羊消化道作用的反应. 草业科学, 2016, 33(8): 1566-1573. |
| [28] |
殷宝法, 淮虎银, 张镱锂, 周乐, 魏万红. 可可西里地区藏羚羊、藏原羚和藏野驴的营养生态位. 应用生态学报, 2007, 18(4): 766-770. |
| [29] |
蒋志刚, 夏武平. 高原鼠兔食物资源利用的研究. 兽类学报, 1985, 5(4): 251-262. |
| [30] |
高中信, 金昆, 马建章, 陈华豪. 呼伦贝尔草原黄羊冬季食性的研究. 兽类学报, 1995, 15(3): 203-208. |
| [31] |
Batzli G O, Pitelka F A. Nutritional ecology of microtine rodents: food habits of lemmings near barrow, alaska. Journal of Mammalogy, 1983, 64(4): 648-655. |
| [32] |
汪诗平. 不同放牧率下绵羊的食性及食物多样性与草地植物多样性间的关系. 生态学报, 2001, 21(2): 237-243. |
| [33] |
Smith E P. Niche breadth, resource availability, and inference. Ecology, 1982, 63(6): 1675-1681. |
| [34] |
Colwell R K, Futuyma D J. On the measurement of niche breadth and overlap. Ecology, 1971, 52(4): 567-576. |
| [35] |
Murie D J. Comparative feeding ecology of two sympatric rockfish congeners, Sebastes caurinus (copper rockfish) and S. maliger (quillback rockfish). Marine Biology, 1995, 124(3): 341-353. |
| [36] |
孙儒泳. 动物生态学原理 (第二版). 北京: 北京师范大学出版社, 1992: 332-338.
|
| [37] |
Augustine D J, Springer T L. Competition and facilitation between a native and a domestic herbivore: trade-offs between forage quantity and quality. Ecological Applications, 2013, 23(4): 850-863. |
| [38] |
Yin B F, Li G L, Wan X R, Shang G Z, Wei W H, Zhang Z B. Large manipulative experiments reveal complex effects of food supplementation on population dynamics of Brandt's voles. Science China Life Sciences, 2017, 60(8): 911-920. |
| [39] |
王旭, 王德利, 刘颖, 巴雷, 孙伟, 张宝田. 羊草草地生长季放牧山羊采食量和食性选择. 生态学报, 2002, 22(5): 661-667. |
| [40] |
王德利. 植物与草食动物之间的协同适应及进化. 生态学报, 2004, 24(11): 2641-2648. |
| [41] |
汪诗平. 不同放牧季节绵羊的食性及食物多样性与草地植物多样性间的关系. 生态学报, 2000, 20(6): 951-957. |