 2022, Vol. 42
2022, Vol. 42文章信息
- 李晓玲, 杨进, 孙雷, 黄应平, 朱士江, 吕坤, 胥焘, 黄桂云, 张国禹
- LI Xiaoling, YANG Jin, SUN Lei, HUANG Yingping, ZHU Shijiang, LÜ Kun, XU Tao, HUANG Guiyun, ZHANG Guoyu
- 中华蚊母树在干旱-水淹交叉胁迫下形态和活性氧代谢的适应机制
- Adaptive mechanism of morphology and reactive oxygen species metabolism of Distylium chinense seedlings to alternate drought and submergence stresses
- 生态学报. 2022, 42(19): 7966-7977
- Acta Ecologica Sinica. 2022, 42(19): 7966-7977
- http://dx.doi.org/10.5846/stxb202008062050
-
文章历史
- 收稿日期: 2020-08-06

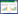




2. 三峡大学生物与制药学院, 宜昌 443002;
3. 中国长江三峡集团有限公司长江珍稀植物研究所, 宜昌 443000
2. College of Biological and Pharmaceutical Science, Three Gorges University, Yichang 443002, China;
3. Rare Plants Research Institute of Yangtze River, Three Gorges Corporation, Yichang 443000, China
随着全球气候变暖, 植物应对干旱和水淹胁迫的生理生态适应机制引起了生态学者的广泛关注[1—2]。由于三峡大坝的修建, 形成了水位落差将近30m、面积达349km2的季节性涨落的消落带[3]。同时, 三峡库区夏季暴雨洪水和高温干旱频发, 促使水位变化频率加快, 导致部分消落带在短期内形成水淹和干旱交叉胁迫现象[4]。这种大幅度、高频次的水位涨落将严重影响三峡库区的安全、健康和可持续发展[5]。因此, 深入研究植物对干旱-水淹交叉胁迫下的特异性生理生态适应机制, 对高效保护库区的生态环境、充分利用土著生物改善消落带生态具有重要意义。
植物体内代谢会产生活性氧ROS(Reactive Oxygen Species), 包括超氧阴离子自由基(O2·)、过氧化氢(H2O2)、羟自由基(· OH)和单线态氧(1O2), 正常情况下, ROS的产生与清除处于动态平衡状态[6]。但是, 逆境下会产生冗余ROS, 导致植物细胞氧化还原状态的失衡, 对细胞具有毒害作用, 可能会导致细胞的死亡[7—9]。植物体内存在ROS清除系统(酶促与非酶促系统)[10—12]。研究表明脯氨酸(Pro)[13—14]、脱落酸(ABA)[15—16]、可溶性糖[如葡萄糖(Glu)][17]等外源物质参与植物的应激状态, 维持植物ROS的代谢平衡。同时, 水淹胁迫会对植物形态产生较大影响[18], 水淹胁迫下, 植物形态产生不定根、皮孔、基茎增加、叶片减少等变化, 以维持正常的生理代谢[19]。目前, 植物在干旱或水淹等非生物胁迫下产生ROS, 通过超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、抗坏血酸过氧化物酶(APX)等酶促系统清除ROS以增强抗氧化能力、提高植物的抗逆性的研究较为深入[8, 20—21], 但目前的研究主要集中在单一逆境胁迫下植物ROS的清除, 关于植物在干旱-水淹交叉胁迫下ROS的清除报道较少。
中华蚊母树(Distylium chinense)是金缕梅科(Hamamelidaceae)蚊母树属(Distylium)多年生常绿河岸带小灌木[22], 是三峡库区河岸带的优势树种和表征种[23], 具有耐淹性好、适应性强等特点, 其根系发达、生长快, 分布广泛, 是三峡库区消落带理想的防沙固土树种。同时, 也是一种较好的园林绿化植物[24]。目前研究主要集中在半淹或全淹等单一因子对中华蚊母树形态、光合作用等方面的变化[25—26], 而关于干旱-水淹交叉胁迫下, 中华蚊母树形态及ROS代谢的生理适应机制尚未见报道。本文通过研究干旱-水淹交叉胁迫及恢复生长阶段施加不同外源物质(Glu、ABA、Pro), 对中华蚊母树幼苗的形态及ROS清除的状况, 拟解决以下3个问题:(1)作为三峡库区河岸带优势树种的中华蚊母树, 在遭受干旱-水淹交叉胁迫时, 其在形态学方面产生哪些适应性机制?(2)在遭受干旱-水淹交叉胁迫后, 如果中华蚊母树能够存活和恢复生长, 那么中华蚊母树的ROS清除系统会产生哪些变化?(3)胁迫恢复阶段, 施加外源物质, 是否对中华蚊母树的恢复生长产生影响?通过对上述问题的回答以阐明中华蚊母树对干旱-水淹交叉胁迫的形态和ROS代谢的适应机制, 可为进一步研究如何提高其在消落带生境存活率提供基础科学数据。
1 材料和方法 1.1 实验材料2017年9月中下旬采集野外种群随机成熟的种子, 共计2000粒, 种植在15cm×13cm的塑料盆中让其自然萌发生长, 种植基质选用腐殖质土和壤土(按1 ∶ 4的比例混合均匀, 土壤pH为6.25, 土壤含水量为27.32%, 土壤容重为1.38g/cm3), 盆内土层厚度约为12cm, 所有幼苗具有相同的生长环境, 且给予除草等常规管理。生长一年后, 用于室外模拟实验, 实验设置在三峡大学生态实验基地(111°32′E, 30°73′N, 海拔高度134m, 年平均降雨量992.1—1404.1mm)进行。
1.2 实验设计于2018年12月从移栽苗中挑选长势和大小相近的450盆进行实验处理。将实验用苗随机分为5组, 每组90盆, 每组再随机分为30盆, 共3个重复, 每个重复30盆。其中, 10盆用于形态学调查, 20盆用于生理生化指标的测定。共设置4个阶段, 阶段1为干旱阶段, 包括对照组(常规供水, CK)、4组干旱(简称D)处理, 共计5个处理组, 每个处理组3个重复, 每个重复30盆, 每个处理组为90盆。对照组CK即为正常供水组, 土壤含水量保持在田间持水量的80%(土壤含水量采用称重法确定)。干旱时间设置4周;阶段2为干旱恢复阶段, 将阶段1的CK组继续常规供水, D处理组于干旱阶段结束当天施加3种不同外源物质(4mg/L ABA[27]、30mg/L Pro[28]、360mg/L Glu[29]), 叶面均匀喷施ABA(简称A)和Pro(简称P)、根部浇灌Glu(简称G)以及胁迫后不施加外源物质(简称N), 见图 1。此时, 干旱恢复处理(简称R)组包括常规供水(CK)、施加脱落酸(该处理简称D-R-A)、施加脯氨酸(该处理简称D-R-P)、施加葡萄糖(该处理简称D-R-G)及胁迫恢复不施加外源物质(该处理简称D-R-N)5个不同处理组。干旱恢复生长设置3周;干旱恢复阶段结束后, 立即将幼苗浸水进行水淹(F)阶段。阶段3为水淹阶段, 将阶段2的CK处理组常规供水(CK), 其他4个处理组进行水淹处理, 共5个处理组, 水淹时间设置9周;阶段4为水淹恢复阶段, 将阶段3的处理组(不包括CK)施加3种外源物质, 此时, 水淹恢复处理组包括常规供水(CK)、施加脱落酸(该处理简称D-R-A-F-R-A)、施加脯氨酸(该处理简称D-R-P-F-R-P)、施加葡萄糖(D-R-G-F-R-G)及不施加外源物质(该处理简称D-R-N-F-R-N)5个不同处理组, 水淹恢复生长设置3周。

|
| 图 1 实验设计图 Fig. 1 Experimental design drawing A:脱落酸Abscisic acid;P:脯氨酸Proline;G:葡萄糖Glucose;C:对照Control;N:进行胁迫处理但恢复阶段不施加外源物质Stress treatment was carried out but no allogenic materials were applied in the recovery stage |
干旱及水淹处理植株放于80cm×50cm×40cm的塑料篮内, 每个塑料篮放15盆, 共计24个塑料篮, 3种外源物质4个处理(不包括CK), 每个处理3次重复, 每个重复30盆, 共360株, 干旱、水淹及正常处理放置于13个水池(2m×1.5m×2.5m)中进行(干旱6个水池、水淹6个水池、正常处理1个水池);对照处理进行常规供水, 定期更换水池中的水, 实验示意图见图 1。盆内土层厚度约为12cm, 所有植株置于三峡大学生态实验基地生长。干旱从2018年10月25日开始, 干旱采取自然连续干旱模式, 每5天采用称重法测一次土壤含水量, 直至土壤含水量达到30%停止干旱, 11月22日停止干旱复水恢复至12月13日, 继续进行水淹处理至2019年2月14日停止水淹恢复至3月7日。采取裂区设计的方法, 水池随机放置植株和塑料篮, 每两周随机轮换一次位置, 以此减少位置效应的影响。同时, 由于模拟用水相对清澈, 为充分模拟库区消落带水体浑浊带来的低光强环境, 在水池上方布设一层遮阳网, 使到达水下植物的光照强度小于10μmol m-2 s -1[30]。所有植株实验期间均不施肥。
在干旱及水淹处理的最后一天, 与干旱及水淹恢复处理的最后一天, 于每个重复内随机采集长势良好的叶片10片, 洗净, 并用定性滤纸擦干后迅速放入液氮, 速冻5min, 放置于超低温冰箱(-78℃)保存, 用于生理生化指标的测定。
1.3 形态特征的测定每个处理期最后一天, 测定用于形态学调查的10株植物的株高、基径、不定根数、叶片数及嫩枝数。
1.4 生理指标的测定参照Giannopolitis等[31]的方法提取粗酶液, 取0.5g叶片于预冷的研钵中, 加5mL 100mmol/L PBS和(pH 7.8)提取液冰浴研磨, 4℃、13000g离心10min, 离心2次, 上清液即为酶提取液, 可用于测定SOD、CAT、APX、O2·。
参照Giannopolitis等[31]的方法检测SOD活性。50μL酶液加入0.3mL 13mmol/L Met、0.3mL 1.3μmol/L核黄素、0.3mL 63μmol/L NBT、1.5mL 50 mmol/L PBS(pH 10.2)、0.3mL 0.1mmol/L EDTA、0.25mL蒸馏水。于4000 lx日光下反应20min, 测定560nm处吸光值。
参照Wang等[32]的方法检测CAT活性。0.5mL酶液中加入2mL 50mmol/L PBS (pH7.0)、0.5mL 40mmol/L H2O2, 于240nm下测定H2O2的分解率。
参照Kanamoto[33]的方法检测APX活性。0.1mL酶液中加入0.1mL 0.1mmol/L EDTA、0.1mL 0.5mmol/L ASA、1.6mL 50mmol/L PBS(pH 7.0), 加入0.1mL 0.1mmol/L H2O2启动反应, 测定290nm处吸光值。
参照王爱国等[34]的方法测定O2·含量0.5mL酶提取液中加入0.5mL 50 mmol/LPBS (pH 7.8)、0.1mL10mmol/L盐酸羟胺, 25℃显色30min, 再加入1mL 58 mmol/L对氨基苯磺酸、1mL 7mmol/L萘胺, 25℃水浴30min。加入等体积三氯甲烷萃取色素, 并于10000r/min离心3min, 于530nm测定上层吸光度。使用0.5mL PBS代替酶提取液, 做空白对照。绘制标准曲线, 计算O2·含量。
参照Bates等[35]的方法测定Pro含量。称取0.5g叶片, 加入10mL 3%磺基水杨酸沸水提取10min, 3000r/min离心10min。取2mL清液, 加入2mL冰醋酸、2mL 2.5%酸性茚三酮, 沸水浴100min, 加入甲苯萃取色素, 于520nm测定甲苯相吸光度。采用脯氨酸标准品绘制标准曲线, 计算Pro含量。· OH采用ELISA试剂盒(南京建成)测定。
1.5 数据分析采用R语言对数据进行图表绘制和统计分析。水淹、干旱胁迫对幼苗的影响, 采用aov()函数进行单因素方差分析;利用corrgram包中的corrgram()函数作出相关系数矩阵图, 以检验施加不同外源物质对ROS清除的影响, 采用multcomp包中的glht()函数进行单因素方差分析, 并进行多重比较;为了比较ROS清除系统的相对重要性, 采用psych包的principal()函数进行PCA分析, 以对ROS清除系统进行检验。采用Excel 2010和SPSS 19.0数据分析软件对形态指标的差异性采用单因素方差分析(One-way ANOVA, Duncan′s multiple range test多重比较)。
2 结果与分析 2.1 干旱-水淹交叉胁迫对中华蚊母树形态学特征的影响经过干旱4周恢复至正常生长, 后再水淹胁迫9周对中华蚊母树幼苗形态学特征产生了显著影响(P < 0.05)。中华蚊母树幼苗经过4周干旱胁迫后, 株高、基茎、不定根数、叶片数及嫩枝数与对照没有显著差异, 但经过后期水淹胁迫9周后, 较对照组株高显著降低, 生长减缓, 基茎显著增加, 不定根也显著增加, 叶片脱落, 嫩枝略有增加但不显著(表 1)。结果表明干旱-水淹交叉胁迫过程中幼苗形态特征的变化, 有利于幼苗适应后期水淹胁迫的低氧条件, 表明前期干旱胁迫促进了对后期水淹胁迫的耐受性。
| 处理 Treatment |
株高/cm Plant height |
基茎/cm Base stem diameter |
不定根数 Number of adventitious |
叶片数 Leaf number |
嫩枝数 Number of epicormic shoots |
| 对照Control | 17.23±0.85a | 1.86±0.03b | 207.83±2.24b | 18.83±0.15a | 1.32±0.28a |
| 干旱Drought | 17.41±0.62a | 1.87±0.36b | 210.83±1.81b | 17.33±0.42b | 1.38±0.18a |
| 水淹Flooding | 15.25±0.38b | 2.34±0.42a | 353.71±2.15a | 17.23±0.72b | 1.50±0.27a |
| 不同处理间标有不同字母者表示差异显著(P < 0.05) | |||||
干旱和水淹交叉胁迫, 对中华蚊母树幼苗ROS及ROS清除具有显著影响(图 2)。在干旱和水淹胁迫下, 中华蚊母树幼苗O2·与· OH等ROS含量均显著升高, SOD、CAT、APX等抗氧化系统酶活性及Pro等抗氧化非生物小分子均显著升高。其中, 干旱阶段SOD、CAT、APX及Pro等抗氧化物质分别高于对照93.94%、120.16%、157.35%、223.31%, 而第二次逆境胁迫水淹阶段的SOD、CAT、APX及Pro等抗氧化物质分别高于对照44.98%、84.08%、39.6%、131.14%, 均达到显著水平。结果表明, 前期干旱胁迫提高了中华蚊母树幼苗对后期水淹胁迫的耐受能力。

|
| 图 2 干旱和水淹交叉胁迫对中华蚊母树幼苗ROS含量及ROS清除的影响 Fig. 2 Effects of alternate drought and flooding stresses on ROS content and ROS clearance of D. chinense seedlings ROS: 活性氧Reactive oxygen species; 不同处理间标有不同字母者表示差异显著(P < 0.05) |
由图 3可以看出, 解除胁迫后的恢复生长阶段, 施加ABA、Pro、Glu等外源物质, 中华蚊母树幼苗SOD、CAT、APX等酶促系统与Pro等非酶促系统对ROS的清除表现出协同性。蓝色表示正相关, 红色表示负相关, 颜色越深, 相关性越强, 可以看出SOD与CAT及APX具有较强的正相关性, Pro与APX、CAT、· OH表现出较强的正相关性, SOD与O2·表现出较强的负相关性。

|
| 图 3 恢复期施加不同外源物质, 中华蚊母树酶促系统、非酶促系统、ROS的相关系数矩阵图 Fig. 3 The matrix diagram of correlation coefficient of enzymatic system, non-enzymatic system and ROS of D. chinensewith different exogenous substances during the recovery growth O2·:超氧阴离子自由基Superoxide anion free radical;· OH:羟自由基Hydroxyl radical; SOD:超氧化物歧化酶Superoxide dismutas; ;APX:抗坏血酸过氧化物酶Ascorbate peroxidase;CAT: 过氧化氢酶Catalase; Pro:脯氨酸Proline |
干旱与水淹恢复阶段施加ABA、Pro、Glu等外源物质, 中华蚊母树幼苗ROS含量及ROS清除系统产生了差异(图 4和图 5)。外源添加Pro, SOD、CAT、APX活性升高, · OH与含量降低;外源添加Glu, SOD、CAT、APX活性未升高, 但· OH与含量降低;外源添加ABA, 内源Pro含量升高。说明外源添加Pro, 可能是通过增强SOD、CAT、APX等保护酶的活性, 进而清除ROS;外源添加Glu, 保护酶未增强, 但植株具有清除ROS的其他途径;外源添加ABA, 可诱导植株内源Pro的生成, 进而可清除· OH。同时, 外源施加ABA, 与内源Pro含量表现出强的正相关性。

|
| 图 4 干旱胁迫恢复阶段施加不同的外源物质(ABA、Glu、Pro) 对中华蚊母树幼苗ROS含量及ROS清除的影响 Fig. 4 Effects of different exogenous substances (ABA, Glu, Pro) on ROS content and ROS clearance of D. chinense seedlings during the recovery growth of drought stress D-R-A:干旱-恢复施加脱落酸Drought-recovery with abscisic acid (ABA);D-R-N:干旱-恢复不施加任何物质Drought-recovery with no any matter;D-R-G:干旱-恢复施加葡萄糖Drought-recovery with glucose(Glu);CK:对照Control;D-R-P:干旱-恢复施加脯氨酸Drought-recovery with proline (Pro) |

|
| 图 5 干旱-水淹恢复阶段施加不同的外源物质(ABA、Glu、Pro) 对中华蚊母树幼苗ROS含量及ROS清除的影响 Fig. 5 Effects of different exogenous substances (ABA, Glu, Pro) on ROS content and ROS clearance of D. chinense seedling during the recovery growth of drought-flooding stresses D-R-A-F-R-A:干旱-恢复施加脱落酸-水淹-恢复施加脱落酸Drought-recovery with abscisic acid (ABA)-flooding-recovery with ABA;D-R-N-F-R-N:干旱-恢复不施加任何物质-水淹-恢复不施加任何物质Drought-recovery with no any matter-flooding-recovery with with no any matter; D-R-G-F-R-G: 干旱-恢复施加葡萄糖-水淹-恢复施加葡萄糖Drought-recovery with glucose(Glu)-flooding-recovery with Glu;CK: 对照Control; D-R-P-F-R-P:干旱-恢复施加脯氨酸-水淹-恢复施加脯氨酸Drought-recovery with proline (Pro)-flooding-recovery with Pro |
水淹胁迫后, 水分过多会引起土壤通气性差和植株根际周围缺氧[19]。这种不良的根际环境会影响根系对水分和矿物质的吸收和转运, 从而引起植株生长减慢和叶片衰老脱落。胁迫后, 植株的生长状况可作为判断植株幼苗水淹耐受能力的重要指标之一[36-37]。经过4周干旱胁迫恢复后再水淹胁迫9周对中华蚊母树的生长影响显著(表 1)。中华蚊母树在形态学上对水分胁迫表现出了适应性。面临水淹胁迫, 植物可能具有两种策略:耐受性与逃避策略[38]。中华蚊母树幼苗在遭受水淹胁迫时, 叶片的减少可能有利于植株的存活, 植株通过脱落叶片的耐受性策略来应对厌氧代谢等造成的巨大消耗, 以此保证根部的存活。同时中华蚊母树植株为适应这种水淹缺氧状态, 在水淹初期开始形成少量不定根、茎基部开始膨大, 随着水淹时间的延长形成更多的不定根, 茎基部膨大部分形成大量皮孔, 茎增粗[25]。水淹过程中不定根和肥大皮孔的产生, 这是耐淹木本植物对水淹的标志性形态反应之一[39]。有研究表明这些肥大的皮孔有大量的细胞间隙增强了气体扩散, 给被水淹的根系提供氧气, 是茎和不定根在大气和内部组织间进行气体交换的一种重要途径, 以便于植物在水淹胁迫下度过不良的外界环境[19, 25, 39]。同时, 在水淹胁迫过程中不定根的形成通过给植株提供水分、矿物质和激素来维持其生长, 使植株耐受水淹[40]。就不定根的发生和茎基部膨大的皮孔来讲, 中华蚊母树在形态上表现出对水淹有一定的适应能力。由此可见, 前期干旱胁迫促进了中华蚊母树对后期水淹胁迫的适应, 如通过叶片脱落、形成不定根及茎基部膨大等形态学的变化增强植物对水淹胁迫的适应。
3.2 酶促系统对中华蚊母树清除非生物胁迫冗余ROS的影响ROS清除能力是决定植物对逆境适应的关键因素之一[41]。植物受到干旱、水淹等非生物胁迫时, ROS含量升高, 质外体ROS的来源之一是NADPH氧化酶(NOX)[42-43], 胁迫使质外体NOX产生氧化应激反应[44], 将还原辅酶Ⅱ(NADPH)的电子(e-)传递给细胞基质的O2, O2得到e-后转化为O2·[45]。酶促系统是清除ROS的重要途径之一。一方面, 在SOD作用下分解O2·为H2O2[46], 而H2O2可通过Fenten反应, 夺取亚铁离子(Fe2+)的e-, 转化为羟自由基(· OH)[44]。同时, CAT也可清除H2O2, 以此抑制· OH的产生。另一方面, 抗坏血酸-谷胱甘肽(ASA-GSH)循环可有效清除H2O2, 且APX可能是这个循环过程中最有效的酶[47], APX以ASA为底物, 可将SOD歧化生成的H2O2还原为H2O。中华蚊母树幼苗在干旱或水淹单一胁迫下, 中华蚊母树· OH、O2·等ROS水平明显高于对照, 表现出氧化应激反应, 其SOD、CAT、APX等抗氧化系统酶活性均显著高于对照, 表现出一定的抗氧化防御作用机制, 且在复合胁迫下, SOD、CAT、APX酶活性显著高于单一胁迫(图 2), 可有效清除冗余的ROS, 使ROS趋于正常水平。因此, SOD、CAT、APX等酶活性的增强, 可能是中华蚊母树通过抗氧化酶系统清除冗余ROS, 是适应逆境胁迫的主要机制(图 6)。

|
| 图 6 非生物胁迫下, 中华蚊母树ROS可能的产生途径及外源物质对清除ROS的促进作用 Fig. 6 Under abiotic stress, the possible ROS production pathways and the promoting effect of allogenic material on scavenging ROS in the D. chinense PPP:磷酸戊糖途径Pentose phosphate pathway;NADPH:还原型辅酶II, 尼克酰胺腺嘌呤二核苷酸磷酸(还原态)Nicotinamide adenine dinucleotide phosphate(reduced state);NADP+:尼克酰胺腺嘌呤二核苷酸磷酸(氧化态) Nicotinamide adenine dinucleotide phosphate(oxidation state);NOX:NADPH氧化酶NADPH oxidase;ASA-GSH:抗坏血酸-谷胱甘肽Ascorbic-glutathione |
植物受到非生物胁迫, ROS激增, 植物通过SOD等抗氧化酶系统和非酶促系统的小分子(ASA、Pro、GSH)直接或间接参与ROS的清除。Smirnoff等首次报道了Pro作为一种· OH的清除剂[48]。Hossain等采用脯氨酸预处理绿豆后进行盐胁迫, 结果发现AsA和GSH含量增加, GSH/GSSR的比率加大, APX、GR(谷胱甘肽还原酶)、CAT活性增加, 降低了H2O2和MDA的含量[13]。白林利等[49]和Ye等[50]均发现水淹-干旱胁迫后脯氨酸和可溶性糖含量显著增加, 脯氨酸含量的增加被认为是提高植物抗氧化胁迫能力的重要指标。在本研究中, 中华蚊母树遭受干旱和水淹胁迫恢复阶段施加外源Pro, Pro与APX、CAT表现出较强的正相关性, SOD与O2·表现出较强的负相关性, Pro与· OH表现出正相关(图 3)。因此, 可以推断Pro作为一种重要的非酶促系统小分子, 在逆境胁迫下Pro的积累, 可能是清除ROS的主要非酶促适应机制, 酶促与非酶促系统表现出协同性, 有利于ROS有效清除。其主要的酶促和非酶促清除ROS的途径可归纳到图 6。逆境胁迫恢复阶段, 中华蚊母树SOD、CAT、APX等酶促系统与Pro等非酶促系统清除ROS表现出不同的贡献率(图 7), 酶促系统清除ROS占主导作用, 且SOD在清除冗余ROS的过程中是最主要的ROS清除酶(图 7)。

|
| 图 7 干旱与水淹阶段酶促系统及非酶促系统在清除ROS中的贡献率 Fig. 7 Contribution rate of enzymatic and non-enzymatic system on ROS scavenging during drought and flooding stresses |
逆境胁迫解除后的快速生长恢复能力是判断消落带岸生物种幼苗水淹耐受能力的重要指标之一[30]。Stewart等采用ABA处理干旱胁迫小麦叶片, Pro大量积累, 表明外源添加ABA, 可诱导植株内源Pro的积累, 进而清除· OH[15]。外源施加ABA后, 中华蚊母树的Pro显著增加(图 4和图 5), 有利于· OH的清除。施加Pro后, 中华蚊母树幼苗CAT、APX酶活性显著增强(图 4和图 5), 表明外源施加Pro, 中华蚊母树可能通过增强CAT与APX等抗氧化酶系统的活性, 进而清除环境压力产生的冗余ROS。在应对应激胁迫时, 植物体内的可溶性糖含量将大量增加[51], Glu可通过氧化的PPP(磷酸戊糖途径)提高NADPH的产生, 而NADPH是ASA-GSH循环的重要辅助因子[17], 同时Glu的添加与· OH表现出显著的负相关(图 4)。但外源施加Glu, 保护酶活性未显著升高(图 4)。因此, 我们推断外源添加Glu, 可能是通过增强中华蚊母树的ASA-GSH循环, 从而以ASA为底物的APX将H2O2降解为H2O, 同时, 可间接降低· OH的含量(图 6)。因此, ABA、Pro、Glu等外源物质的施加, 可促进中华蚊母树对逆境胁迫产生的冗余ROS的有效清除。
3.5 干旱-水淹交叉胁迫对中华蚊母树逆境胁迫耐受能力的影响适当的交叉逆境胁迫可增强植物对于胁迫的耐受力。薹草属植物瘤囊薹草(Carex schmidtii Meinsh)在水淹和干旱复合胁迫下, SOD等抗氧化酶系统表现出了更高的活性, 同时脯氨酸含量增加及形态学的适应性特征表现出了较单一胁迫对复合胁迫更好的适应性[52]。干旱-水淹交叉胁迫下, 中华蚊母树幼苗水淹阶段SOD、CAT、APX等抗氧化酶防御体系与Pro等抗氧化小分子较单一干旱阶段均显著上调, 且水淹处理下幼苗的O2·与· OH显著低于干旱处理, 同时SOD、CAT、APX等抗氧化酶活性较单一水淹胁迫显著上调, 表现出对水淹胁迫更好的耐受能力。因此, 在干旱-水淹交叉胁迫下前期适度干旱胁迫可促进中华蚊母树对后期水淹的耐受能力。
总之, 植物对于非生物胁迫的适应是多方面的。遭受干旱、水淹胁迫时, O2·、· OH等ROS将大量增加, 中华蚊母树可能是通过SOD、CAT、APX等抗氧化酶系统及Pro等非酶促抗氧化小分子提高抗氧化能力并表现出协同效应, 清除冗余的ROS, 减少ROS增加造成的损伤。逆境胁迫下, 中华蚊母树表现出较好的抗氧化防御, 可减少氧化应激对植物造成的损伤, 同时, 施加ABA、Pro、Glu等外源物质对ROS清除产生影响:外源添加Pro, 中华蚊母树可能是通过增强CAT、APX酶的活性提高抗氧化能力;外源添加ABA, 可能是通过诱导植株Pro的积累, 从而清除· OH;而外源添加Glu, 虽酶促系统活性未提升, 中华蚊母树可能是通过PPP途径, 间接清除H2O2。结果表明, 中华蚊母树植株对于干旱、水淹胁迫具有一定的耐受能力, 并具有清除冗余ROS的有效机制, 同时在形态上表现出适应性特征。逆境胁迫下, 中华蚊母树产生与清除ROS的可能机制如图 6, 并表现出前期适度干旱胁迫可促进中华蚊母树对后期水淹的耐受能力。
| [1] |
Ferreira C S, Piedade M T F, Junk W J, Parolin P. Floodplain and upland populations of Amazonian Himatanthus sucuuba: effects of flooding on germination, seedling growth and mortality. Environmental and Experimental Botany, 2007, 60(3): 477-483. DOI:10.1016/j.envexpbot.2007.01.005 |
| [2] |
Du Y L, Zhao Q, Chen L W, Yao X D, Zhang W, Zhang B, Xie F T. Effect of drought stress on sugar metabolism in leaves and roots of soybean seedlings. Plant Physiology and Biochemistry, 2020, 146: 1-12. DOI:10.1016/j.plaphy.2019.11.003 |
| [3] |
袁兴中, 熊森, 李波, 徐静波, 刘红, 王强. 三峡水库消落带湿地生态友好型利用探讨. 重庆师范大学学报: 自然科学版, 2011, 28(4): 23-25. |
| [4] |
吴佳, 高学杰, 张冬峰, 石英, Giorgi F. 三峡水库气候效应及2006年夏季川渝高温干旱事件的区域气候模拟. 热带气象学报, 2011, 27(1): 44-52. DOI:10.3969/j.issn.1004-4965.2011.01.005 |
| [5] |
李昌晓, 钟章成. 三峡库区消落带土壤水分变化对落羽杉(Taxodium distichum)幼苗根部次生代谢物质含量及根生物量的影响. 生态学报, 2007, 27(11): 4394-4402. DOI:10.3321/j.issn:1000-0933.2007.11.003 |
| [6] |
McCord J M, Fridovich I. Superoxide dismutase: an enzymic function for erythrocuprein (hemocuprein). Journal of Biological Chemistry, 1969, 244(22): 6049-6055. DOI:10.1016/S0021-9258(18)63504-5 |
| [7] |
Gill S S, Tuteja N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiology and Biochemistry, 2010, 48(12): 909-930. DOI:10.1016/j.plaphy.2010.08.016 |
| [8] |
Smirnoff N. The role of active oxygen in the response of plants to water deficit and desiccation. New Phytologist, 1993, 125(1): 27-58. DOI:10.1111/j.1469-8137.1993.tb03863.x |
| [9] |
Marino D, Dunand C, Puppo A, Pauly N. A burst of plant NADPH oxidases. Trends in Plant Science, 2012, 17(1): 9-15. DOI:10.1016/j.tplants.2011.10.001 |
| [10] |
Apel K, Hirt H. Reactive oxygen species: metabolism, oxidative stress, and signal transduction. Annual Review of Plant Biology, 2004, 55: 373-399. DOI:10.1146/annurev.arplant.55.031903.141701 |
| [11] |
Heller J, Tudzynski P. Reactive oxygen species in phytopathogenic fungi: signaling, development, and disease. Annual Review of Phytopathology, 2011, 49(1): 369-390. DOI:10.1146/annurev-phyto-072910-095355 |
| [12] |
Wrzaczek M, Brosché M, Kangasjärvi J. ROS signaling loops-production, perception, regulation. Current Opinion in Plant Biology, 2013, 16(5): 575-582. DOI:10.1016/j.pbi.2013.07.002 |
| [13] |
Hossain M A, Hasanuzzaman M, Fujita M. Coordinate induction of antioxidant defense and glyoxalase system by exogenous proline and glycinebetaine is correlated with salt tolerance in mung bean. Frontiers of Agriculture in China, 2011, 5(1): 1-14. DOI:10.1007/s11703-010-1070-2 |
| [14] |
Wani A S, Ahmad A, Hayat S, Tahir I. Is foliar spray of proline sufficient for mitigation of salt stress in Brassica juncea cultivars?. Environmental Science and Pollution Research, 2016, 23(13): 13413-13423. DOI:10.1007/s11356-016-6533-4 |
| [15] |
Stewart C R, Voetberg G. Relationship between stress-induced ABA and proline accumulations and ABA-induced proline accumulation in excised barley leaves. Plant Physiology, 1985, 79(1): 24-27. DOI:10.1104/pp.79.1.24 |
| [16] |
Jiang M Y, Zhang J H. Effect of abscisic acid on active oxygen species, antioxidative defence system and oxidative damage in leaves of maize seedlings. Plant and Cell Physiology, 2001, 42(11): 1265-1273. DOI:10.1093/pcp/pce162 |
| [17] |
Couée I, Sulmon C, Gouesbet G, El Amrani A. Involvement of soluble sugars in reactive oxygen species balance and responses to oxidative stress in plants. Journal of Experimental Botany, 2006, 57(3): 449-459. DOI:10.1093/jxb/erj027 |
| [18] |
Thomas A L, Guerreiro S M C, Sodek L. Aerenchyma formation and recovery from hypoxia of the flooded root system of nodulated soybean. Annals of Botany, 2005, 96(7): 1191-1198. DOI:10.1093/aob/mci272 |
| [19] |
Voesenek L A C J, Bailey-Serres J. Flood adaptive traits and processes: an overview. New Phytologist, 2015, 206(1): 57-73. DOI:10.1111/nph.13209 |
| [20] |
Ashraf M. Inducing drought tolerance in plants: recent advances. Biotechnology Advances, 2010, 28(1): 169-183. DOI:10.1016/j.biotechadv.2009.11.005 |
| [21] |
Kaur G, Asthir B. Molecular responses to drought stress in plants. Biologia Plantarum, 2017, 61(2): 201-209. DOI:10.1007/s10535-016-0700-9 |
| [22] |
Zhang Z Y, Chang H T, Endress P K. Hamamelidaceae//Wu Z Y, Raven P H, Hong D Y, eds. Flora of China. Beijing: Science Press, 2003: 18-42.
|
| [23] |
王勇, 厉恩华, 吴金清. 三峡库区消涨带维管植物区系的初步研究. 武汉植物学研究, 2002, 20(4): 265-274. DOI:10.3969/j.issn.2095-0837.2002.04.005 |
| [24] |
Xiang L, Li X L, Wang X S, Yang J, Lv K, Xiong Z Q, Chen F Q, Huang C M. Genetic diversity and population structure of Distylium chinense revealed by ISSR and SRAP analysis in the Three Gorges Reservoir Region of the Yangtze River, China. Global Ecology and Conservation, 2020, 21: e00805. DOI:10.1016/j.gecco.2019.e00805 |
| [25] |
Li X L, Li N, Yang J, Ye F Z, Chen F J, Chen F Q. Morphological and photosynthetic responses of riparian plant Distylium chinense seedlings to simulated autumn and winter flooding in Three Gorges Reservoir Region of the Yangtze River, China. Acta ecologica Sinica, 2011, 31(1): 31-39. DOI:10.1016/j.chnaes.2010.11.005 |
| [26] |
Liu Z B, Cheng R M, Xiao W F, Guo Q S, Wang N. Effect of off-season flooding on growth, photosynthesis, carbohydrate partitioning, and nutrient uptake in Distylium chinense. PLoS One, 2014, 9(9): e107636. DOI:10.1371/journal.pone.0107636 |
| [27] |
王书宏, 杜永吉. 外源激素对干旱胁迫下草莓光合特性的影响. 中国农学通报, 2008, 24(12): 367-371. |
| [28] |
唐依萍, 张晓艳, 刘士壮, 萧蓓蕾. 外源脯氨酸对NaCl胁迫下番茄幼苗光合特性的影响. 安徽农业科学, 2015, 43(24): 9-11. DOI:10.3969/j.issn.0517-6611.2015.24.004 |
| [29] |
汪菊香. 外源葡萄糖对盐胁迫下黄瓜幼苗光合作用和离子分布的影响. 安徽农业科学, 2015, 43(1): 7-10. DOI:10.3969/j.issn.0517-6611.2015.01.003 |
| [30] |
Luo F L, Nagel K A, Scharr H, Zeng B, Schurr U, Matsubara S. Recovery dynamics of growth, photosynthesis and carbohydrate accumulation after de-submergence: a comparison between two wetland plants showing escape and quiescence strategies. Annals of Botany, 2011, 107(1): 49-63. DOI:10.1093/aob/mcq212 |
| [31] |
Giannopolitis C N, Ries S K. Superoxide dismutases: I. Occurrence in higher plants. Plant Physiology, 1977, 59(2): 309-314. DOI:10.1104/pp.59.2.309 |
| [32] |
Wang Y S, Tian S P, Xu Y. Effects of high oxygen concentration on pro-and anti-oxidant enzymes in peach fruits during postharvest periods. Food Chemistry, 2005, 91(1): 99-104. DOI:10.1016/j.foodchem.2004.05.053 |
| [33] |
Kanamoto H, Miyake C. Photoinactivation of ascorbate peroxidase in isolated tobacco chloroplasts: Galdieria partita APX maintains electron flux through the water-water cycle in transplastomic tobacco plants//Allen J F, Gantt E, Golbeck J H, Osmond B, eds. Photosynthesis. Energy from the Sun. Dordrecht: Springer, 2008: 613-616.
|
| [34] |
王爱国, 罗广华. 植物的超氧物自由基与羟胺反应的定量关系. 植物生理学通讯, 1990(6): 55-57. |
| [35] |
Bates L S, Waldren R P, Teare I D. Rapid determination of free proline for water-stress studies. Plant and Soil, 1973, 39(1): 205-207. DOI:10.1007/BF00018060 |
| [36] |
Mommer L, Lenssen J P M, Huber H, Visser E J W, De Kroon H. Ecophysiological determinants of plant performance under flooding: a comparative study of seven plant families. Journal of Ecology, 2006, 94(6): 1117-1129. DOI:10.1111/j.1365-2745.2006.01175.x |
| [37] |
Vreugdenhil S J, Kramer K, Pelsma T. Effects of flooding duration, -frequency and -depth on the presence of saplings of six woody species in north-west Europe. Forest Ecology and Management, 2006, 236(1): 47-55. DOI:10.1016/j.foreco.2006.08.329 |
| [38] |
Parolin P. Submergence tolerance vs. escape from submergence: two strategies of seedling establishment in Amazonian floodplains. Environmental and Experimental Botany, 2002, 48(2): 177-186. DOI:10.1016/S0098-8472(02)00036-9 |
| [39] |
Yamamoto F, Sakata T, Terazawa K. Physiological, morphological and anatomical responses of Fraxinus mandshurica seedlings to flooding. Tree Physiology, 1995, 15(11): 713-719. DOI:10.1093/treephys/15.11.713 |
| [40] |
Islam M A, Macdonald S E. Ecophysiological adaptations of black spruce (Picea mariana) and tamarack (Larix laricina) seedlings to flooding. Trees, 2004, 18(1): 35-42. DOI:10.1007/s00468-003-0276-9 |
| [41] |
Taylor C B. Proline and water deficit: ups, downs, ins, and outs. The Plant Cell, 1996, 8(8): 1221-1224. DOI:10.1105/tpc.8.8.1221 |
| [42] |
Kámán-Tóth E, Dankó T, Gullner G, Bozsó Z, Palkovics L, Pogány M. Contribution of cell wall peroxidase-and NADPH oxidase-derived reactive oxygen species to Alternaria brassicicola-induced oxidative burst in Arabidopsis. Molecular Plant Pathology, 2019, 20(4): 485-499. DOI:10.1111/mpp.12769 |
| [43] |
Wieczfinska J, Sitarek P, Skała E, Kowalczyk T, Pawliczak R. Inhibition of NADPH oxidase-derived reactive oxygen species decreases expression of inflammatory cytokines in A549 Cells. Inflammation, 2019, 42(6): 2205-2214. DOI:10.1007/s10753-019-01084-0 |
| [44] |
Lambeth J D. NOX enzymes and the biology of reactive oxygen. Nature Reviews Immunology, 2004, 4(3): 181-189. DOI:10.1038/nri1312 |
| [45] |
Singh R, Singh S, Parihar P, Mishra R K, Tripathi D K, Singh V P, Chauhan D K, Prasad S M. Reactive oxygen species (ROS): beneficial companions of plants' developmental processes. Frontiers in Plant Science, 2016, 7: 1299. |
| [46] |
Gechev T S, Hille J. Hydrogen peroxide as a signal controlling plant programmed cell death. Journal of Cell Biology, 2005, 168(1): 17-20. DOI:10.1083/jcb.200409170 |
| [47] |
Ishikawa T, Yoshimura K, Sakai K, Tamoi M, Takeda T, Shigeoka S. Molecular characterization and physiological role of a glyoxysome-bound ascorbate peroxidase from spinach. Plant and Cell Physiology, 1998, 39(1): 23-34. DOI:10.1093/oxfordjournals.pcp.a029285 |
| [48] |
Smirnoff N, Cumbes Q J. Hydroxyl radical scavenging activity of compatible solutes. Phytochemistry, 1989, 28(4): 1057-1060. |
| [49] |
白林利, 韩文娇, 李昌晓. 前期水淹对水杉树苗响应干旱胁迫的影响. 西北农林科技大学学报: 自然科学版, 2015, 43(5): 42-50. |
| [50] |
Ye T T, Shi H T, Wang Y P, Chan Z L. Contrasting changes caused by drought and submergence stresses in bermudagrass (Cynodon dactylon). Frontiers in Plant Science, 2015, 6: 951. |
| [51] |
Sun L, Li X L, Wang X S, Xiang, L, Yang J, Min Q F, Chen G H, Chen F Q, Huang C M, Wang G X. Growth and respiratory metabolic adaptation strategies of riparian plant Distylium chinense to submergence by the field study and controlled experiments. Plant Physiology and Biochemistry, 2020, 157: 1-12. |
| [52] |
Zhang D J, Qi Q, Wang X H, Tong S Z, Lv X G, An Y, Zhu X Y. Physiological responses of Carex schmidtii Meinsh to alternating flooding-drought conditions in the Momoge wetland, northeast China. Aquatic Botany, 2019, 153: 33-39. |