 2022, Vol. 42
2022, Vol. 42文章信息
- 丁茂, 郑标, 王丽, 张金铭, 周守标, 王智
- DING Mao, ZHENG Biao, WANG Li, ZHANG Jinming, ZHOU Shoubiao, WANG Zhi
- 鹞落坪国家级自然保护区草本植物多样性及空间分布
- Diversity and spatial distribution of herbaceous species with deciduous broad-leaved forest in the Yaoluoping National Nature Reserve
- 生态学报. 2022, 42(19): 8015-8030
- Acta Ecologica Sinica. 2022, 42(19): 8015-8030
- http://dx.doi.org/10.5846/stxb201912272809
-
文章历史
- 收稿日期: 2019-12-27





2. 安徽省水土污染治理与修复工程实验室, 芜湖 241003;
3. 生态环境部南京环境科学研究所, 南京 210042
2. Anhui Province Engineering Laboratory of Water and Soil Pollution Control and Remediation, Wuhu 241003, China;
3. Nanjing Institute of Environmental Sciences, Ministry of Ecology and Environment, Nanjing 210042, China
林下草本植物是森林群落中的重要组成部分和关键要素。研究表明[1—2], 林下草本植物在森林中发挥着重要的生态学作用, 其可为生物提供栖息地和食物资源, 增加森林物种丰富度;可过滤和拦截地表径流, 减少林地水土流失, 增加土壤层的氮(N)、磷(P)等营养元素和有机质, 使土壤结构和肥力维持高水平状态。此外, 林下草本植物还能促进森林生态系统有机碳积累、提高碳汇, 在调节局部小气候及群落演替等方面也有明显作用。
森林空间格局是指种群在森林群落内的分布格局及在空间上的配置状况, 是植物种群的基本特征, 研究主要集中于种群的空间分布格局和种间关联性两个方面[3]。种群的空间分布格局是指在一定水平空间范围内某一种群的分布状况, 它反映了种群的生物生态学特性, 种群内和种群间的关系, 以及种群与环境相互作用的结果[4]。研究种群的空间分布格局能够更深入地认识种群个体的生长状况与结构特征、了解种群分布规律、掌握群落演替程度、预测整个森林及生态系统变化趋势。种间关联性主要是指在一定空间范围内种群内不同物种之间存在的联系或差异, 研究种间联结性可以更好地了解物种的特征、物种在群落中的分布情况、物种间的相互关系以及物种与环境的相互作用, 是揭示群落类型、结构和群落演替的重要手段[5—7]。
近年来, 鹞落坪自然保护区开展了森林群落空间结构、植物区系和物种组成[8—9]方面的研究, 但研究空间与时间尺度较小, 缺少大型规范的综合性观测和研究平台。调查表明, 鹞落坪自然保护区在海拔1000 m以上是典型的落叶阔叶林, 是研究落叶阔叶林组成、结构与功能及其变化规律的理想区域。本研究以鹞落坪自然保护区落叶阔叶林11.56 km2大样地内林下草本植物为调查对象, 通过对大样地内草本层植物调查与统计, 分析草本植物多样性、空间分布格局及优势种的种间联结性和相关性;分析重要值前10位草本植物的种群空间分布格局、种间联结性和相关性;研究草本层植物的空间分布与环境之间关联性。该研究为进一步探讨鹞落坪落叶阔叶林生物多样性和物种共存机制提供参考;为自然保护区森林生态系统保护管理与资源合理利用提供理论依据[10—11]。
1 研究区概况 1.1 地理位置与区划如图 1所示, 鹞落坪自然保护区位于安徽省岳西县境内大别山东南处(30°57′—31°06′N, 116°02′—116°11′E), 是安徽、湖北和河南三省交界处, 与桐柏山脉相连, 属秦岭余脉。鹞落坪自然保护区主要保护对象为大别山地区具有典型代表性的森林植被和国家重点保护与珍稀濒危动植物资源, 也是大别山地区的重要水源涵养林保护区。保护区总面积123 km2, 划分为三个功能区, 其中核心区面积21.2 km2(包括石佛寺-大川岭核心区、吊罐井核心区、麒麟沟核心区和多枝尖核心区), 缓冲区面积28.4 km2, 实验区面积73.4 km2。

|
| 图 1 研究区示意图 Fig. 1 The diagram of the study area |
鹞落坪自然保护区呈现南高北洼之势, 因受造山运动和外力作用的影响, 使得保护区内群峰林立、山体宽厚, 发育着瀑布、陡崖、悬流等景观。在气候上属于亚热带季风气候, 受副热带高压与江淮气候的影响, 降水充沛、空气湿润、气温偏低;境内温差较大, 最高处与最底处年平均气温相差5.8℃, 全年平均气温为12.7℃, 全年降水量在1400—1800 mm之间。
鹞落坪自然保护区溪流纵横交错, 是大别山地区重要的水源涵养地。保护区地带性植被属于北亚热带常绿落叶阔叶混交林带, 沿海拔梯度向上植被类型呈现向暖温带落叶阔叶林变化趋势[12]。研究表明[8—9], 保护区内蕴藏着丰富的植物资源, 共有维管植物2000多种, 大型真菌200多种, 其中珍稀濒危植物有30余种;近年来新发表的植物新种数十种, 如美丽鼠尾草(Salvia meiliensis)、黄山石豆兰(Bulbophyllum huangshanense)、大别山冬青(Ilex dabieshanensis)、四棱过路黄(Lysimachia jinzhaiensis)、大别山排草(Lysimachia dabieshanensis)、天马过路黄(Lysimachia tianmaensis)等。
2 研究方法 2.1 样地建设与调查方法如图 2所示, 大样地(30°57′46.11—30°57′47.81N, 116°04′41.33—116°04′41.08E)大小为340 m×340 m, 总面积为11.56 km2, 共划分为289个20 m×20 m样方, 在每个样方的四角分别设置样桩(包括水泥桩和钢管桩)。样桩起始点位于样地东北角, 以每个样桩为起点, 分别向4个方向拉出5 m×5 m的亚样方, 在每个亚样方中划分出25个1 m×1 m的小样方对草本进行调查, 将调查后的小样方草本数据列入相应的样方中(20 m×20 m)。

|
| 图 2 鹞落坪样地永久样方示意图 Fig. 2 Schematic diagram of permanent sample of Y aoluping sample plot |
调查采用实验室研发的动态监测样地野外植物物种多样性的调查工具系统(已获软件著作权), 记录大样地内草本的种名、株数、平均高度、最高高度和盖度等指标。
2.2 植物区系分析大样地内草本植物参照《中国植物志》在线数据库进行鉴定与分类。植物区系成分分析以吴征镒[13—14]种子植物科、属分布区类型的划分为依据, 分析大样地内草本植物科、属的分布区类型。
2.3 物种多样性指数分析(1) 重要值计算[15]
重要值=(相对频度+相对多度+相对盖度)/3。
其中, 相对频度=(某种的频度/所有种的总频度)×100%;相对多度=(某种的株数/所有种的株数)×100%;相对盖度=(某个物种的盖度/所有物种的总盖度)×100%。
(2) 物种多样性指数计算[16]
采用Simpson指数(DS)、Shannon-Wiener指数(H)和Pielou均匀度指数(J), 如表 1所示。
| 多样性指数 Diversity index |
计算方式 Calculation formula |
备注 Remarks |
| 辛普森指数(DS) Simpson (DS) |
 |
式中, Pi是一个个体属于第i种的概率;S为样地内出现的物种数 |
| 香浓-维尔指数(H) Shannon-Wiener (H) |
 |
|
| Pielou均匀度指数(J) Pielou (J) |
 |
采用7种分析方法[17—18], 如表 2所示。综合分析样地内草本优势种的空间分布格局, 在聚集强度分析时, 当7个指标中有4个及以上判定结果一致, 则认为该结果为种群的聚集强度分析结果, 使用Excel软件进行统计计算, 具体指标计算如下:
| 计算指标 Calculation index |
计算方式 Calculation formula |
备注 Remarks |
| 扩散系数(C) Diffusion coefficient (C) |
 |
X为各种群多度的均值, S2为各种群多度的方差。C>1时为聚集分布;C=1时为随机分布;当C<1时为均匀分布。 |
| 平均拥挤度(m*) Average congestion degree (m*) |
 |
m*>1时为聚集分布, m*=1时为随机分布, m*<1时为均匀分布。 |
| 负二项参数(K) Negative binomial parameter (K) |
 |
当K值趋于无穷大时, 则逼近Poisson分布;K<0时为聚集分布;K=0时为均匀分布。 |
| 丛生指标(I) Cluster index (I) |
 |
当I>0时为聚集分布;I=0时为随机分布;I<0时为均匀分布。 |
| 聚块性指标(PI) Agglomeration index (PI) |
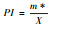 |
当PI>1时为聚集分布;PI=1时为随机分布;PI<1时为均匀分布。 |
| Cassie指标(CA) Cassie index (CA) |
 |
当CA>0时为聚集分布;CA=0时为随机分布;CA<0时为均匀分布。 |
| Green指标(GI) Green index (GI) |
 |
n为基本样方数, 当GI>0时为聚集分布;GI = 0时为随机分布;GI<0时为均匀分布 |
如表 3所示, 建立2×2联列表, 使用SPSS软件进行统计计算, 采用χ2检验、联结系数AC方法对种间关联程度与联结性进行测定。
| 物种A Species A | 物种B Species B | 合计Total | |
| 出现Appear | 不出现Disappear | ||
| 出现 Appear | a | b | a + b |
| 不出现 Disappear | c | d | c + d |
| 合计 Total | a + c | b + c | N = a + b + c + d |
| a为2个物种同时出现的样方数,b、c为2个物种单独出现的样方数,d为2个物种均未出现的样方数 | |||
(1) χ2检验
由于取样的非连续性, 对于非连续性数据的χ2用Yates的连续校正公式计算[19]。
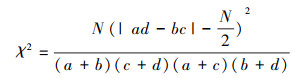
|
式中, N为小样方总数, a为2个物种同时出现的样方数,b、c为2个物种单独出现的样方数,d为2个物种均未出现的样方数。χ2检验以1为自由度, 95%的显著性水平下的卡方值为3.841, 99%的显著性水平下的卡方值为6.635。拟假定置信区间在99%水平下显著为极显著, 在95%水平下显著为显著。
(2) 联结系数AC
联结系数AC主要用来测定种间关联程度。
① ad≥bc时, 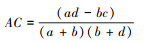
② ad<bc且d≥a时, 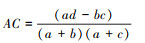
③ ad<bc且d<a时, 
联结系数AC的值域为[-1, 1], AC值趋近于-1时, 表明种对间的负关联程度更强;AC值趋近于1时, 表明种对间的正关联程度更强;AC值为0时, 则表明种对间处于完全独立的状态。
2.4.3 种间相关性分析运用Pearson相关系数检验和Spearman秩相关系数检验测定种间的关联强度[20], 将数据定量化去分析联结强度的大小以及种间关联性的差异。
Pearson相关系数计算:

|
式中, rs(i, j)是物种i与k之间在j样方中的Pearson相关系数, N为总样方数, xij和xkj分别为物种i与物种k在样方j中的多度值, 它们分别组成两个向量xi与xk;xi和xk分别是物种i和物种k在所有样方中的平均多度值。rik值的范围为[-1, 1], 所得值为正即为正相关, 所得值为负即为负相关[20]。
Spearman秩相关系数计算:
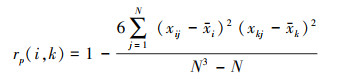
|
式中, rp(i, j)分别是种i与种k之间在j样方中的Spearman秩相关系数。N为总样方数, xij和xkj分别为种i和种k在j样方中的多度值的秩[20]。
3 结果分析 3.1 草本植物区系组成经调查与统计, 大样地内共有草本植物52科134属195种(将具有草本性状特征的木质藤本和亚灌木一并列为草本植物)。由于实际分类精确问题, 将莎草科(Cyperaceae)薹草属(Carex sp.)植物中除青绿薹草(Carex breviculmis)、垂穗薹草(Carex brachyathera)和宽叶薹草(Carex siderosticta)之外的物种统一标记为薹草属(Carex sp.)。
3.1.1 科的分析(1) 大小科的分析
如表 4所示, 在样地内52科草本植物中, 含10种及以上的科有5个, 共67种, 分别占样地草本植物总科数和总种数的9.62%和34.36%, 在样地植物区系中占主导作用;含2—9种的科有24个, 共105种, 分别占样地草本植物总科数和总种数的46.15%和53.85%, 其中含6—9种的科数与种数占11.54%和24.10%, 含2—5种的科数与种数占34.62%和29.74%;含1种的科有23个, 共23种, 分别占样地草本植物总科数和总种数的44.23%和11.79%。在前十种草本优势种中, 堇菜科和禾本科各有1种, 茜草科和十字花科各有2种, 莎草科有4种。
| 种数 No. of species |
科名(属数/种数) No. of families |
种数 No. of families |
| ≥10 | 百合科(12/18);唇形科(6/10);堇菜科(1/10);菊科(13/17);毛莨科(8/12) | 67 |
| 6—9 | 禾本科(8/9);石竹科(6/9);伞形科(6/8);天南星科(4/8);蓼科(4/7);茜草科(3/6) | 47 |
| 2—5 | 兰科(5/5);莎草科(2/5);十字花科(2/5);报春花科(1/5);景天科(2/4);荨麻科(3/4);罂粟科(2/4);葫芦科(2/3);龙胆科(3/3);葡萄科(2/3);蔷薇科(3/3);虎耳草科(2/2);桔梗科(2/2);马兜铃科(2/2);猕猴桃科(1/2);木通科(2/2);玄参科(2/2);紫草科(2/2) | 58 |
| 1 | 23科(1/1)(科名略) | 23 |
| 合计Total | 195 |
(2) 科的分布区类型
如表 5所示, 在样地内52个科的草本植物中, 世界分布类型28科, 占样地内总科数的53.85%;泛热带分布类型10科, 占样地内总科数的19.23%, 北温带分布类型8科, 占样地内总科数的15.38%。表明样地内草本植物区系除世界分布科外, 科的分布具有泛热带和北温带双重性质。
| 分布区类型 Areal type |
科数 No. of families |
百分比 Percentage/% |
| 1 世界分布 Cosmopolitan | 28 | 53.85 |
| 2 泛热带分布 Pantropic | 10 | 19.23 |
| 3 东亚(热带、亚热带)及热带南美间断分布 Trop. & Subtr. E. Asia & (S.) trop. Amer. disjuncted | 1 | 1.92 |
| 4 热带亚洲至热带大洋洲分布 Tropical Asia to Tropical Australasia | 1 | 1.92 |
| 5 北温带分布 N.Temp. | 8 | 15.38 |
| 6 东亚及北美间断分布 E. Asia & N. Amer. disjuncted | 2 | 3.85 |
| 7 旧世界温带分布 Old world Temperate | 1 | 1.92 |
| 8 东亚分布 E. Asia | 1 | 1.92 |
| 合计 Total | 52 | 100 |
(1) 大小属的分析
如表 6所示, 大样地内草本植物的134属中, 含5种以上的属有1个, 占总属的0.75%;含3—5种的属有5个, 占总属的3.73%;含1—3种的属有128个, 占总属的95.52%。结果表明, 样地内植物属分散, 植物组成复杂。
| 属内含种量 No. of species in genera |
属数 No. of genera |
占总属数/% %/of total genera |
种数 No. of species |
占总种数/% %/of total species |
| >5 | 1 | 0.75 | 10 | 5.13 |
| 3—5 | 5 | 3.73 | 21 | 10.77 |
| <3 | 128 | 95.52 | 164 | 84.10 |
| 总计 Total | 134 | 100 | 195 | 100 |
(2) 属的分布区类型
大样地内草本植物属的分布区类型划分, 如表 7所示。世界分布的属共有25属, 占总属数的18.66%, 主要包括珍珠菜属(Lysimachia)、车前属(Plantago)、鼠尾草属(Salvia)、香科科属(Teucrium)、灯心草属(Juncus)等。
| 分布类型 Areal type |
属数 No. of genera |
百分比 Percentage/% |
| 1.世界分布 Cosmopolitan | 25 | 18.66 |
| 2.泛热带分布 Pantropic | 11 | 8.21 |
| 3.旧世界热带分布 Old World Tropics | 1 | 0.75 |
| 4.热带亚洲至热带大洋洲分布 Tropical Asia to Tropical Australasia | 3 | 2.24 |
| 5.热带亚洲至热带非洲分布 Trop. Asia to Trop. Africa | 4 | 2.99 |
| 6.热带亚洲分布 Trop. Asia | 6 | 4.48 |
| 7.北温带分布 N. Temp. | 39 | 29.10 |
| 8.东亚和北美洲间断分布 E. Asia & N. Amer. disjuncted | 9 | 6.72 |
| 9.旧世界温带分布 Old World Temperate | 12 | 8.95 |
| 10.温带亚洲分布 Temp. Asia | 1 | 0.75 |
| 11.东亚分布 E. Asia | 20 | 14.93 |
| 12.中国特有分布 Endemic to China | 3 | 2.24 |
| 合计 Total | 134 | 100 |
热带分布的属共有25属, 其中泛热带分布的属有11属, 占总属数的8.21%, 主要包括求米草属(Oplismenus)、泽兰属(Eupatorium)、虾脊兰属(Calanthe)等。旧世界热带分布的属仅有1属, 即楼梯草属(Elatostema)。热带亚洲至热带大洋洲分布的属有3属, 包括淡竹叶属(Lophatherum)、兰属(Cymbidium)和通泉草属(Mazus)。热带亚洲至热带非洲分布的属有4属, 包括芒属(Miscanthus)、赤瓟属(Thladiantha)、大丁草属(Leibnitzia)和魔芋属(Amorphophallus)。热带亚洲分布的属有6属, 主要有香茶菜属(Isodon)、绞股蓝属(Gynostemma)、翅果菊属(Pterocypsela)等。
温带分布的属共有81属, 占总属数的60.44%, 在样地中占据主要地位。其中北温带分布的属有39属, 主要包括百合属(Lilium)、黄精属(Polygonatum)、藜芦属(Veratrum)等。东亚和北美洲间断分布的属有9属, 主要有黄精叶钩吻属(Croomia)、鹿药属(Smilacina)、黄水枝属(Tiarella)等。旧世界温带分布的属有12属, 包括重楼属(Paris)、野芝麻属(Lamium)、苜蓿属(Medicago)等。温带亚洲分布的属仅有1属, 即孩儿参属(Pseudostellaria)。东亚分布的属有20属, 包括大百合属(Cardiocrinum)、开口箭属(Campylandra)、沿阶草属(Ophiopogon)等。
中国特有分布的属共有3属, 包括大血藤属(Sargentodoxa)、血水草属(Eomecon)和车前紫草属(Sinojohnstonia)。
大样地内草本植物区系地理组成成分复杂, 涵盖众多分布区类型, 其中温带成分较多, 占总属数的60.44%, 表明该区落叶阔叶林群落温带特性明显, 但与东灵山[21]等北方地区典型的落叶阔叶林的温带成分属占总属数的93.75%相比还存在较大差别;热带植物成分其次占总属数的18.66%, 表明样地内草本植物对温带生长环境具有明显的亲缘性。
3.2 重要值和物种α多样性分析 3.2.1 重要值重要值是测定物种在植物群落中相对重要性的一个指标, 它能反映一个物种在植物群落中的优势程度。经统计, 大样地内草本植物共计222694株, 按重要值由大到小的顺序排列出前10种草本优势植物, 如表 8所示。前10种的草本植物重要值占总重要值的58.57%, 其中青绿薹草、垂穗薹草与宽叶薹草各占26.07%、8.82%与4.49%, 明显高于其它草本优势种, 表明样地内的生态环境最适合薹草属植物生长。
| 植物名称 Species |
相对频度/% Relative frequency |
相对多度/% Relative dominance |
相对盖度/% Relative abundance |
重要值/% Important value |
| 青绿薹草 Carex breviculmis | 5.32 | 42.21 | 30.68 | 26.07 |
| 垂穗薹草 Carex brachyathera | 3.49 | 11.80 | 11.16 | 8.82 |
| 宽叶薹草 Carex siderosticta | 4.40 | 3.49 | 5.56 | 4.49 |
| 柯孟披碱草 Elymus kamoji | 2.40 | 3.66 | 5.23 | 3.77 |
| 鸡腿堇菜 Viola acuminata | 4.00 | 3.47 | 2.32 | 3.26 |
| 六叶律 Galium hoffmeisteri | 2.94 | 3.63 | 2.15 | 2.90 |
| 光头山碎米荠 Cardamine engleriana | 1.36 | 3.33 | 3.35 | 2.68 |
| 大叶碎米荠Cardamine macrophylla | 1.65 | 1.68 | 4.70 | 2.67 |
| 薹草属 Carex sp. | 2.22 | 1.91 | 1.88 | 2.01 |
| 四叶葎 Galium bungei | 2.25 | 2.58 | 0.89 | 1.91 |
物种多样性是群落组成的重要特征, 不仅反映了群落组成物种丰富度和种间均匀分配程度, 同时也反映了不同自然地理条件与群落的相互关系以及群落的稳定性与动态[22]。
如表 9所示, Simpson指数(DS)是对多样性反面集中性的度量, 集中性越高表明多样性越低。反映一个群落中物种优势程度的重要指标, 数值范围为0—1, 数值越接近1则表明多样性越高。大样地内草本植物物种的Simpson指数(DS)为0.83, 表明样地内草本植物多样性很高。
| 多样性指数 Diversity index |
多样性指数 Diversity index |
|||
| Simpson 指数 Simpson index(DS) | 0.83 | Pielou 均匀度指数 Pielou evenness index(J) | 0.69 | |
| Shannon-Wiener 指数 Shannon-Wiener index(H) | 3.63 |
Shannon-Wiener指数(H)可以预测从群落中随机抽出一个个体所属种的平均不定度。当群落中种的数目增加和已存在种的个体数量分布愈来愈均匀时, 不定度明显增加, 多样性也就愈大, 所得数值一般在1.5—3.5之间[23]。数值能反映一个群落中物种或个体的数量特征、物种在生境上的差异、群落的结构特征以及群落的演替状况。样地内草本物种的Shannon-Wiener指数(H)为3.63, 表明调查的大样地内草本植物具有很高的多样性。
Pielou均匀度指数(J)是考虑群落中种的个体均匀度, 进一步对多样性测定进行补充和完善, 反映的是个体完全均匀分布的多样性。Pielou均匀度指数(J)的范围为0—1, 0代表只有一个样方内有物种分布, 1代表所有样方都存在物种且数量相同, 样地内草本物种的Pielou均匀度指数为0.69, 表明大样地内草本植物种类整体分布较均匀。
如图 3所示, 各样方的Shannon-Wiener指数(H)在0.18—2.14之间变动, 大样地内北部阳坡物种多样性较低, 南部阴坡、中部半阳坡的物种多样性较高, 因为有溪流经过的区域, 土壤水分较多, 有利于草本植物生长发育。由于受光照时间、强度的不同使得不同坡向的温度、湿度与水分等生境条件产生了差异, 从而造成大样地内不同区域草本植物丰富度和多样性的差异, 这与郑世群等[24]对福建戴云山保护区研究结果相似。

|
| 图 3 各样方草本Shannon-Wiener指数 Fig. 3 The Shannon-Wiener index of herbs in each plot |
大样地草本植物物种的Shannon-Wiener指数(H)和Pielou均匀度指数(J)明显高于西双版纳热带森林样地[25]和古田山亚热带常绿阔叶林样地[26]的林下草本植物, 如表 10所示。由于西双版纳热带森林和古田山亚热带常绿阔叶林群落均为发育较成熟的森林群落, 乔木层发育良好, 林下郁闭度较高, 草本层一年四季都处于光照较弱的荫闭环境, 仅仅适合少量耐荫植物生长;而大样地内乔木层处于群落演替中期阶段[27], 冠幅相对较小且树种间间距相对较大, 给林下草本层植物生长创造了良好的生长环境;此外, 大样地为落叶阔叶林, 具有明显的季相变化, 在乔木层发叶之前, 林下阳光充足生长着大量早春植物和一年生或多年生草本植物。本研究的调查时间为5月份, 正是早春植物和一年生或多年生草本植物生长与繁殖期[28], 因此林下草本植物较为丰富, 物种丰富度与多样性较高。
| 群落类型 Community type |
Shannon-Wiener指数(H) Shannon-Wiener (H) |
Pielou均匀度指数(J) P ielou (J) |
| 鹞落坪落叶阔叶林 Yaoluoping deciduous broad-leaved forest |
3.63 | 0.69 |
| 西双版纳热带森林 Xishuangbanna tropical forest |
2.75 | 0.49 |
| 古田山亚热带常绿阔叶林 Gutianshan sub-tropical evergreen broad-leaved forest |
1.39 | 0.32 |
种群的空间分布格局是了解森林群落动态特征及物种分布的重要指标。采用种群多度的方差/均值扩散系数(C)来测定格局类型, 采用平均拥挤度(m*)、负二项参数(K)、丛生指标(I)、聚块性指数(PI)、Cassie指标(CA)和Green指数(GI)[29]综合分析了样地内重要值前10位的优势草本植物空间分布格局。10个优势种群均表现为聚集分布, 如表 11所示。从大样地优势草本植物种的聚集程度来看, 不同草本植物对温度、湿度和阳光等环境因子的适应性各不相同。在森林群落演替过程中, 物种聚集程度与生态系统的稳定性存在一定的联系, 物种聚集程度越高, 说明该物种对群落内的环境越适应, 对群落资源的利用更加合理, 群落更加稳定。
| 种名 Species |
扩散系数 Diffusion coefficient (C) |
平均拥挤度 Average congestion degree (m*) |
负二项参数 Negative binomial parameter (K) |
丛生指标 Cluster index (I) |
聚块性指标 Agglomeration index (PI) |
Cassie指标 Cassie index (CA) |
Green指标 Green index (GI) |
分布格局 Distribution pattern |
| 青绿薹草 Carex breviculmis | 422.31 | 137786.52 | 0.77 | 421.31 | 423.61 | 1.30 | 475.83 | 聚集分布 |
| 垂穗薹草 Carex brachyathera | 281.81 | 25906.45 | 0.32 | 280.81 | 284.90 | 3.09 | 88.66 | 聚集分布 |
| 宽叶薹草 Carex siderosticta | 50.53 | 1409.03 | 0.54 | 49.53 | 52.37 | 1.84 | 4.62 | 聚集分布 |
| 柯孟披碱草 Elymus kamoji | 133.87 | 3910.29 | 0.21 | 132.87 | 138.57 | 4.71 | 13.01 | 聚集分布 |
| 鸡腿堇菜 Viola acuminata | 65.22 | 1807.45 | 0.42 | 64.22 | 67.62 | 2.40 | 5.96 | 聚集分布 |
| 六叶律 Galium hoffmeisteri | 92.41 | 2673.40 | 0.31 | 91.41 | 95.68 | 3.27 | 8.86 | 聚集分布 |
| 光头山碎米荠 Cardamine engleriana |
211.56 | 5645.34 | 0.12 | 210.56 | 219.76 | 8.20 | 18.78 | 聚集分布 |
| 大叶碎米荠 Cardamine macrophylla |
96.41 | 1341.11 | 0.14 | 95.41 | 103.80 | 7.38 | 4.28 | 聚集分布 |
| 薹草属 Carex sp. | 96.32 | 1514.38 | 0.15 | 95.32 | 102.78 | 6.47 | 4.87 | 聚集分布 |
| 四叶葎 Galium bungei | 116.27 | 2429.43 | 0.17 | 115.27 | 122.06 | 5.79 | 7.96 | 聚集分布 |
由图 4可以看出, 大样地内群落前10个优势种45个物种对中, 呈现极显著相关的有20对占总物种对的44.44%, 呈现显著相关的有7对占总物种对的15.56%, 不显著相关的有18对占总物种对的40.00%, 表明群落中草本种群种间联结性较弱, 草本层植物发育不成熟。呈现正相关的有26对占总物种对的57.78%, 其中显著相关与极显著相关的种对有19对, 不显著正相关的有7对。大样地中六叶律与四叶葎表现出极显著的正相关, 两者同属茜草科(Rubiaceae)拉拉藤属(Galium), 生物学特征与生长习性极为相似;呈现负相关的有19对占总物种对的42.22%, 其中显著负相关与极显著负相关的种对有8对, 不显著负相关的有11对, 如柯孟披碱草与鸡腿堇菜呈现极显著的负相关, 主要是因为两者对温度、空气湿度以及土壤水分等环境的适应性不同。

|
| 图 4 样地内优势种群种间关联的χ2半矩阵图 Fig. 4 Semi-matrix of interspecific correction χ2-test of association of dominant population in the polt 1.青绿薹草(Carex breviculmis);2.垂穗薹草(Carex brachyathera);3.宽叶薹草(Carex siderosticta);4.柯孟披碱草(Elymus kamoji);5.鸡腿堇菜(Viola acuminata);6.六叶律(Galium hoffmeisteri);7.光头山碎米荠(Cardamine engleriana);8.大叶碎米荠(Cardamine macrophylla);9.薹草属(Carex sp.);10.四叶葎(Galium bungei) |
在对种间联结性分析时, 运用χ2检验仅能客观准确地得出种间联结性是否显著, 却无法判断出种间联结性的大小, 因此需要用联结系数AC综合判断种间关联性。
大样地内群落10个草本优势种联结系数AC半矩阵图, 如图 5所示。AC≥0.40的种对数有4对占总种对数的8.89%, 分别为垂穗薹草与宽叶薹草、垂穗薹草与鸡腿堇菜、六叶律与四叶葎和光头山碎米荠与大叶碎米荠, 说明各种对之间正联结程度较高, 对生境的需求相似;0.20≤AC<0.40的种对数有6对占总种对数的13.33%, 说明种对间呈现不显著正联结;-0.20≤AC<0.20的种对数有29对占总种对数的64.45%, 说明种对间联结松散, 分布趋于独立;有2对(宽叶薹草与薹草属、宽叶薹草与四叶葎)AC值为0, 说明它们在大样地内分布完全独立, 相互排斥性极强;-0.40≤AC<-0.20的种对数有6对占总种对数的13.33%, 说明种对间呈现不显著负相关。结合χ2检验值与联结系数AC来看, 正联结的种对虽多于负联结的种对, 但是关联程度集中在较低的水平, 表明群落中草本种群种间联结性较弱, 草本层植物发育不成熟。

|
| 图 5 样地内优势种群种间联结半矩阵图 Fig. 5 Semi-matrix of interspecific association of dominant population in the polt AC: 联结系数 Association coefficient |
利用χ2检验来判断种间关联性, 结合Pearson相关系数、Spearman秩相关系数检验种间的相关性, 能更准确全面地了解大样地内草本优势种群间的相互关联性[30]。
种对的Pearson相关系数与Spearman秩相关系数是反映两个物种间线性关系的重要指标[31]。如图 6所示, 正相关种对数多于负相关的种对数, 且极显著正相关的种对数量较多, 表明种对间具有相似的生态适应性和生态位, 对于抵御外部环境干扰的能力较差, 整体关联性不强。这种较强的正相关性, 也说明群落的成熟程度较低, 各物种间很少以相互独立关系求得资源的充分利用, 从而无法保持物种间稳定发展, 这也与前文χ2检验所述一致。

|
| 图 6 样地内草本优势种群种间Pearson相关系数和Spearman秩相关系数半矩阵图 Fig. 6 Semi-matrix of Pearson′s correlation coefficients and Spearman′s rank correlation coefficients of main population in the polt |
大样地内优势种群的Pearson相关检验, 如表 12所示。前10个草本优势种的45个物种种对中, 呈现正相关的种对为25对, 占总种对数的55.56%, 其中呈极显著正相关的对数为12, 占总种对数的26.67%;呈显著正相关的种对数为4, 占总种对数的8.89%。种对间呈现负相关的种对数为20对, 其中呈现极显著负相关与显著负相关的种对数各为3, 各占总种对数的6.67%。呈现不显著的总种对数共有23对, 占总种对数的51.11%, 可以看出优势种种间关联性不强, 种间整体松散状态, 这与优势种群χ2检验结果一致。
| 检验方法 Test methods |
正联结(相关)Positive association(correlation) | 负联结(相关)Negative association(correlation) | ||||||
| 极显著 Distinctly significant (P<0.01) |
显著 Significant (P≤0.05) |
不显著 Not significant (P>0.05) |
总数 Sum |
极显著 Distinctly significant (P<0.01) |
显著 Significant (P≤0.05) |
不显著 Not significant (P>0.05) |
总数 Sum |
|
| χ2检验 χ2-test |
16(35.56) | 3(6.67) | 7(15.56) | 26(57.78) | 4(8.89) | 4(8.89) | 11(24.44) | 19(42.22) |
| Pearson 相关系数 Pearson′s correlation coefficient |
12(26.67) | 4(8.89) | 9(20.00) | 25(55.56) | 3(6.67) | 3(6.67) | 14(31.11) | 20(44.44) |
| Spearman 秩相关系数 Spearman′s rank correlation coefficient |
17(37.78) | 1(2.22) | 9(20.00) | 27(60.00) | 9(20.00) | 2(4.44) | 7(15.56) | 18(40.00) |
大样地内优势种群的Spearman秩相关检验, 如表 12所示。前10个草本优势种的45个物种种对中, 呈现正相关的种对为27对, 占总种对数的60.00%, 其中呈极显著正相关的对数为17, 占总种对数的37.78%;呈显著正相关的种对数为1, 占总种对数的2.22%。种对间呈现负相关的种对数为18对, 其中呈现极显著负相关的种对数为9, 占总种对数的20.00%;呈现显著负相关的种对数为2, 占总种对数的4.44%。呈现不显著的总种对数为16对, 占总种对数的35.56%, 可以看出优势种种间关联性不强, 种间整体松散状态, 这与优势种群χ2检验结果一致。
4 讨论与结论森林是地球上最重要的陆地生态系统, 由具有丰富的生物多样性、生态复杂性和地方特有性, 不同森林类型和规模均在区域发挥着重要的环境功能[32]。在森林生态系统中, 各种生命形式、动植物物种多样性都是各种生境和生态系统的重要指标[33]。森林空间格局是指种群在森林群落内的分布格局及在空间上的配置状况, 是植物种群的基本特征。森林空间格局的研究主要集中于种群的空间分布格局和种间关联性两个方面[3]。种群的空间分布格局是指在一定水平空间范围内某一种群的分布状况, 它不仅仅反映了种群的生物生态学特性, 还反映了种群及群落的动态演替特征, 以及种群与环境相互作用的过程, 对揭示种群的恢复机理具有重要意义[34]。优势种群的群落结构和动态能反映群落的稳定性, 种间关联性可以揭示群落物种间的联结强度与相关程度, 同时物种间的关联关系也受生境、区域气候变化以及群落演替进程的影响[35—37]。
本研究对鹞落坪自然保护区落叶阔叶林11.56 km2大样地内林下草本植物进行的系统调查中共记录到52科134属195种222694株。与典型北方落叶阔叶林下的草本植物区系相比, 大样地内草本区系成分复杂, 虽有热带区系成分, 但总体表现为温带区系特征。从重要值来看, 大样地内青绿薹草、垂穗薹草与宽叶薹草占据明显优势, 表明薹草属植物更适合生存于落叶阔叶林下的环境条件。采用七种种群分布格局分析的方法综合分析大样地内优势种的分布格局, 10个优势种群均表现为聚集分布, 说明该落叶阔叶林处于演替中期阶段, 尚未达到稳定成熟的落叶阔叶林。大样地内的Simpson指数(DS)、Shannon-Wiener指数(H)和Pielou均匀度指数(J)分别为0.83、3.63、0.69, 表明落叶阔叶林下草本植物多样性较高, 但各样方中物种多样性具有较大差异。多样性高的样方多集中在两类区域, 其一为有河流穿过的大样地中部与西南角, 原因可能是有河流经过的地方不利于乔木及灌木生长, 致使草本植物生长受到的竞争较小;其二为大样地南部阴坡, 大样地内南部阴坡的草本植物多样性高于北部阳坡, 这与王子婷等[38]在黄土丘陵区研究不同坡向草本群落分布特征的结果相一致。
种间联结性是在一定空间范围内不同物种间的关联程度以及物种对生境的适应状况, 在不同的群落演替阶段种间的关联程度有着不同的变化[39]。在χ2检验基础上, 采用联结系数AC定性分析种间的联结性强度, 结合Pearson相关系数与Spearman秩相关系数定量分析种间的相关性程度, 综合结果表明大样地内的草本植物整体关联性较差, 种间竞争较强, 物种间少有独立的关系对资源进行充分利用。通过对大样地内的草本植物种间联结性检验结果分析表明, Pearson相关系数检验的适用范围是针对于两个变量之间是连续且线性关系、总体是正态分布或者是接近正态的单峰分布的数据;而Spearman秩相关系数检验对两个变量的总体分布形态、样本容量的大小没有要求;因此Spearman秩相关系数检验的灵敏度较Pearson相关系数检验更高, 应用范围较广, 能弥补Pearson相关系数检验的不足, 这与大多数的研究结果一致[40—42]。
一般来讲, 群落稳定性越强, 种间联结越趋向于正联结;反之, 群落越不稳定, 种间排斥性就越大, 种间联结越趋向于负相关[43]。大样地林下草本植物中正相关与正联结的种对数多于负相关负联结的种对数, 但关联强度与相关程度大多集中在相对较低的水平, 表明草本植物种间以及与环境之间关系尚未达到相对的动态平衡状态, 群落处于演替中期阶段, 尚未达到稳定成熟的落叶阔叶林, 草本分析与乔木分析该落叶阔叶林发育阶段的结论保持一致[27]。
草本植物多样性在维管植物多样性中占比很大, 且对各种全球变化驱动因素较为敏感, 包括大气沉降和森林经营等因素[44]。鹞落坪自然保护区落叶阔叶林11.56 km2大样地内林下草本植物组成丰富, 覆盖度较高, 是该地区植物群落的重要组成部分。草本植物能有效地调节水循环、改善水质、拦截蓄水, 增加大别山地区的水源涵养量, 在雨期水土流失的预防和提高土壤有机质含量方面起到重要作用[45]。研究该地区林下草本植物空间分布对该落叶阔叶林生物多样性恢复、土壤结构改良、林地小气候改善等具有重要意义, 为大样地深入研究提供理论依据。
| [1] |
陈煜, 许金石, 张丽霞, 郭垚鑫, 柴永福, 王茂, 张晨光, 岳明. 太白山森林群落和林下草本物种变化的环境解释. 西北植物学报, 2016, 36(4): 784-795. |
| [2] |
Elliott K J, Vose J M, Knoepp J D, Clinton B D, Kloeppel B D. Functional role of the herbaceous layer in eastern deciduous forest ecosystems. Ecosystems, 2015, 18(2): 221-236. DOI:10.1007/s10021-014-9825-x |
| [3] |
吴初平, 袁位高, 盛卫星, 黄玉洁, 陈庆标, 沈爱华, 朱锦茹, 江波. 浙江省典型天然次生林主要树种空间分布格局及其关联性. 生态学报, 2018, 38(2): 537-549. |
| [4] |
Hui G Y, Li L, Zhao Z H, Dang P X. Comparison of methods in analysis of the tree spatial distribution pattern. Acta Ecologica Sinica, 2007, 27(11): 4717-4728. DOI:10.1016/S1872-2032(08)60008-6 |
| [5] |
Maestre F T, Rodríguez F, Bautista S, Cortina J, Bellot J. Spatial associations and patterns of perennial vegetation in a semi-arid steppe: a multivariate geostatistics approach. Plant Ecology, 2005, 179(2): 133-147. DOI:10.1007/s11258-004-4641-6 |
| [6] |
祝燕, 白帆, 刘海丰, 李文超, 李亮, 李广起, 王顺忠, 桑卫国. 北京暖温带次生林种群分布格局与种间空间关联性. 生物多样性, 2011, 19(2): 252-259. |
| [7] |
范娟, 赵秀海, 汪金松, 张春雨, 何俊, 夏富才. 江西九连山亚热带常绿阔叶林优势种空间分布格局. 生态学报, 2012, 32(9): 2729-2737. |
| [8] |
李珍, 周忠泽, 汪文革, 沈三宝. 安徽鹞落坪自然保护区维管植物区系分析. 生物学杂志, 2008, 25(6): 26-30. |
| [9] |
王志高, 张中信, 汪文革, 储俊. 安徽岳西县鹞落坪国家级自然保护区森林群落结构的初步分析. 植物生态学报, 2016, 40(6): 615-619. |
| [10] |
Wyatt J L, Silman M R. Centuries-old logging legacy on spatial and temporal patterns in understory herb communities. Forest Ecology and Management, 2010, 260(1): 116-124. DOI:10.1016/j.foreco.2010.04.010 |
| [11] |
Small C J, McCarthy B C. Spatial and temporal variability of herbaceous vegetation in an eastern deciduous forest. Plant Ecology, 2003, 164(1): 37-48. DOI:10.1023/A:1021209528643 |
| [12] |
谢中稳, 吴国芳. 安徽省鹞落坪自然保护区的植被类型及其分布. 华东师范大学学报: 自然科学版, 1995(3): 93-102. |
| [13] |
吴征镒. 中国种子植物属的分布区类型. 云南植物研究, 1991, 13(S4): 1-139. |
| [14] |
吴征镒, 周浙昆, 李德铢, 彭华, 孙航. 世界种子植物科的分布区类型系统. 云南植物研究, 2003, 25(3): 245-257. |
| [15] |
孙儒泳, 李博, 诸葛阳, 尚玉昌. 普通生态学. 北京: 高等教育出版社, 1993.
|
| [16] |
钱迎倩, 马克平. 生物多样性研究的原理与方法. 北京: 中国科学技术出版社, 1994.
|
| [17] |
刘俊霞, 袁登胜, 袁秀英. 花棒群落物种多样性及其种群空间分布格局的研究. 西北林学院学报, 2008, 23(5): 55-59. |
| [18] |
范玉龙, 胡楠, 丁圣彦, 卢训令. 伏牛山国家级自然保护区森林生态系统草本优势种空间分布格局. 河南农业大学学报, 2014, 48(5): 590-595. |
| [19] |
张金屯. 数量生态学. 北京: 科学出版社, 2004.
|
| [20] |
刘润红, 姜勇, 常斌, 李娇凤, 荣春艳, 梁士楚, 杨瑞岸, 刘星童, 曾惠帆, 苏秀丽, 袁海莹, 傅桂焕, 吴燕慧. 漓江河岸带枫杨群落主要木本植物种间联结与相关分析. 生态学报, 2018, 38(19): 6881-6893. |
| [21] |
刘晓. 北京东灵山亚高山草甸维管植物区系与多样性研究[D]. 北京: 北京林业大学, 2011.
|
| [22] |
王伯荪. 植物群落学. 北京: 高等教育出版社, 1987.
|
| [23] |
Magurran A E. Ecological Diversity and Its Measurement. Dordrecht: Springer, 1988.
|
| [24] |
郑世群. 福建戴云山国家级自然保护区植物多样性及评价研究[D]. 福州: 福建农林大学, 2013.
|
| [25] |
李强. 西双版纳热带森林林窗空间格局与林下植物多样性相关性研究[D]. 昆明: 云南大学, 2019.
|
| [26] |
田锴. 古田山亚热带常绿阔叶林草本植物群落结构和多样性格局研究[D]. 金华: 浙江师范大学, 2013.
|
| [27] |
王丽, 常锦利, 周守标, 王晓英, 张佳期, 闫少凯, 张金铭, 陈鑫, 赵昕, 王智. 鹞落坪国家级自然保护区乔木物种多样性与种间联结. 生态学报, 2019, 39(1): 309-319. |
| [28] |
Elliott K J. Herbaceous species composition and richness of mesophytic cove forests in the southern Appalachians: synthesis and knowledge gaps. The Journal of the Torrey Botanical Society, 2014, 141(1): 39-71. |
| [29] |
郭树江, 杨自辉, 王多泽, 李得禄, 李爱德, 詹科杰, 王强强. 民勤绿洲一荒漠过渡带植物物种多样性及其优势种群空间分布格局研究. 水土保持研究, 2011, 18(3): 92-96. |
| [30] |
简敏菲, 刘琪璟, 朱笃, 游海. 九连山常绿阔叶林乔木优势种群的种间关联性分析. 植物生态学报, 2009, 33(4): 672-680. |
| [31] |
张金屯. 植被数量生态学方法. 北京: 中国科学技术出版社, l995.
|
| [32] |
Pérez M R, Fernández C G, Sayer J A. Los servicios ambientales de los bosques. Ecosistemas, 2007, 16(3): 81-90. |
| [33] |
Kutschker A, Brand C, Miserendino M L. Evaluación de la calidad de los bosques de ribera en ríos del NO del Chubut sometidos a distintos usos de la tierra. Ecología Austral, 2009, 19(1): 19-34. |
| [34] |
Borchsenius F, Nielsen P K, Lawesson J E. Vegetation structure and diversity of an ancient temperate deciduous forest in SW Denmark. Plant Ecology, 2004, 175(1): 121-135. |
| [35] |
张明霞, 王得祥, 康冰, 张岗岗, 刘璞, 杜焰玲, 于飞. 秦岭华山松天然次生林优势种群的种间联结性. 林业科学, 2015, 51(1): 12-21. |
| [36] |
申文辉, 李志辉, 欧芷阳, 何琴飞, 谭长强, 彭玉华, 谭一波. 桂西南蚬木群落乔木层优势种的种间关联性分析. 南方农业学报, 2016, 47(2): 256-262. |
| [37] |
燕亚媛, 金一兰, 刘思齐, 丛日慧, 陈俊俊, 牛建明, 丁勇. 内蒙古草原优势种群种间关联特征分析. 内蒙古大学学报: 自然科学版, 2016, 47(6): 617-624. |
| [38] |
王子婷, 杨磊, 李广, 柴春山, 张洋东, 刘冬皓. 黄土丘陵区坡面柠条(Caragana korshinskii)林地草本植物分布特征. 中国沙漠, 2021, 41(02): 120-128. |
| [39] |
叶权平, 张文辉, 于世川, 薛文艳. 桥山林区麻栎群落主要乔木种群的种间联结性. 生态学报, 2018, 38(9): 3165-3174. |
| [40] |
邓莉萍, 白雪娇, 李露露, 牛沙沙, 韩美娜, 秦胜金, 周永斌. 辽东山区次生林优势木本植物种间联结与相关分析. 生态学杂志, 2015, 34(6): 1473-1479. |
| [41] |
高永恒, 曾晓阳, 周国英, 王根绪. 长江源区高寒湿地植物群落主要种群种间关系分析. 湿地科学, 2011, 9(1): 1-7. |
| [42] |
杨兆静, 张钦弟, 李豪, 白玉宏, 毕润成. 山西五鹿山自然保护区暴马丁香群落木本植物种间联结性分析. 植物科学学报, 2013, 31(1): 42-48. |
| [43] |
王乃江, 张文辉, 陆元昌, 范少辉, 王勇. 陕西子午岭森林植物群落种间联结性. 生态学报, 2010, 30(1): 67-78. |
| [44] |
Gilliam F S. The ecological significance of the herbaceous layer in temperate forest ecosystems. BioScience, 2007, 57(10): 845-858. |
| [45] |
张艳华, 聂绍荃, 王志西. 林隙对草本植物的影响. 植物研究, 1999(1): 94-99. |