 2022, Vol. 42
2022, Vol. 42文章信息
- 李佳, 薛亚东, 吴波, 李迪强
- LI Jia, XUE Yadong, WU Bo, LI Diqiang
- 气候变化背景下秦岭地区羚牛生境脆弱性评估及适应性保护对策
- Assessing vulnerability of suitable golden takins (Budorcas taxicolor bedfordi) habitat and adaptive conservation strategies to climate change in Qinling Mountains, China
- 生态学报. 2022, 42(18): 7484-7494
- Acta Ecologica Sinica. 2022, 42(18): 7484-7494
- http://dx.doi.org/10.5846/stxb202108062159
-
文章历史
- 收稿日期: 2021-08-06

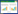




2. 中国林业科学研究院森林生态环境与自然保护研究所/国家林业和草原局生物多样性保护重点实验室, 北京 100091
2. Ecology and Nature Conservation Institute, Chinese Academy of Forestry, and Key Laboratory of Biodiversity Conservation of National Forestry and Grassland Administration, Beijing 100091, China
全球气候变暖给生物多样性带来的影响引起人们的普遍关注[1]。大量观测事实表明, 全球气候变暖已经对动物、植物、生物多样性、乃至整个系统产生深刻影响, 且这些影响在未来将会更加剧烈[2—3]。如果全球气候变暖得不到有效遏制, 未来温度升高2℃(升高2℃被UNFCCC定义为“危险”温度[4]), 预测全球将会有35%的物种面临灭绝风险[5—7], 无疑将会给未来生物多样性保护带来严峻挑战。脆弱性是指物种受气候变化影响的程度, 脆弱性评估是研究气候变化影响生物多样性的重要内容, 是适应和减缓未来气候变化影响的关键和基础[8]。国际上IPCC、IUCN、欧盟等组织, 以及美国等国家正在开展相关研究, 试图通过脆弱性评估工作, 提出物种适应未来气候变化的科学对策[9—11]。目前, 脆弱性评估是比较新的研究领域, 现有文献中将评估方法归纳为三类:生物气候包络模型评估、机理性生态位模型和脆弱性指数评估[8, 12—13]。基于空间生态位理论发展起来的生物气候包络模型是评估物种脆弱性使用最普遍的方法, 通过模型模拟技术获取准确的物种当前潜在分布范围, 并能预测未来分布变化[14—15]。物种是否脆弱主要通过比较当前和未来分布区的变化, 如果物种未来分布区与当前分布区重叠较少, 或者未来分布区大范围缩减, 则表明气候变化对物种影响较大[16—17]。
羚牛(Budorcas taxicolor)主要分布在我国四川、陕西、甘肃、云南和西藏等地区, 以及印度东北部和不丹、缅甸北部等区域[18—19], 是森林生态系统中体型最大的物种之一。羚牛主要栖息在高山、亚高山针叶林和针阔混交林生境, 具有季节性沿海拔梯度垂直迁徙的生活习性[20]。自20世纪80年代以来, 研究人员开展了一系列羚牛生态学研究, 涉及生境、食性、繁殖习性、活动节律、季节性迁徙、以及家域等方面[21—26]。近几十年来, 由于人类活动范围的不断扩大, 以及对森林资源的过度利用, 羚牛分布范围急剧减少, 适宜栖息地从大面积的连续斑块缩减成破碎化的独立小斑块, 种群数量呈现明显下降趋势, 羚牛被IUCN列为“易危”物种[27], 也是我国国家I级重点保护野生动物。中国政府采取颁布野生动物保护法、建立自然保护区、天然林保护工程、野生动物监测项目等措施来保护羚牛种群[28—29]。经过多年共同努力, 许多曾经对羚牛种群生存产生严重威胁的影响因素(如森林采伐、偷猎、非法贸易等)得到有效缓解, 部分地区野外种群数量开始缓慢增长[30]。当前, 人类活动以及由此造成的全球气候变暖是威胁羚牛种群的主要因素, 可能会给许多保护措施带来极大的挑战, 迫切需要采取适应性保护对策来减缓气候变化给羚牛种群带来的不利影响。
全球气候变化背景下, 秦岭地区生物多样性保护工作面临着挑战, 尤其是栖息在该区域的羚牛、大熊猫(Ailuropoda melanoleuca)、川金丝猴(Rhinopithecus roxellana)等濒危物种对气候变化非常敏感, 气候变化正影响着这些物种是无庸置疑的[31—34]。我国政府高度重视秦岭生态保护与建设, 在秦岭地区设立国家公园, 主要保护该区域生物多样性和自然生态系统原真性、完整性, 对推进秦岭生物多样性科学保护具有极其重要的意义。因此, 本研究选择秦岭地区羚牛作为研究对象, 预测气候变化背景下羚牛当前和未来适宜生境变化, 评估脆弱程度, 以及当前已建立保护区体系面对未来气候变化的保护成效性;同时, 鉴定潜在气候避护所, 构建生态廊道, 进而为提出羚牛适应性保护对策和国家公园建设提供科学依据。
1 研究地点秦岭山系位于陕西省南部(图 1;东经106°30′—108°05′, 北纬32°40′—34°35′), 是我国长江和黄河域的分水岭, 也是我国重要南北自然地理界线。山势起伏较大, 北坡陡峭, 南坡较缓, 山地多处于海拔1000 m以上, 最高峰太白山海拔3767 m, 为陕西省第一高峰。秦岭山系受亚热带湿润季风气候和暖温带半湿润季风性气候的双重影响, 气候具有明显垂直分带, 中高海拔山区寒冷且湿润, 年均温度6—8℃, 年均降水量800—1200 mm;低山、河谷年均温度11—14℃, 年均降水量800 m, 南坡比北坡年均温度更高。秦岭山系具有典型亚热带与暖温带过渡特征生态系统和明显垂直带谱, 为许多古老生物种提供了避难所, 也是许多珍惜濒危物种唯一或重要栖息地[35]。秦岭山系是羚牛分布的最北界, 主要分布在佛坪、太白、宁陕县、洋县等1500—3600 m的落叶阔叶林、针阔混交林、亚高山针叶林及灌丛草甸中。受益于大熊猫保护, 羚牛种群获得了较好的保护, 部分区域种群数量出现缓慢增长[30]。目前, 秦岭山系已建有佛坪、长青、太白山、等国家级自然保护区, 老县城、黄柏塬、皇冠山等省自然保护区, 共计19个自然保护区[36]。

|
| 图 1 秦岭山系羚牛分布点 Fig. 1 Distribution of golden takin in Qinling Mountains |
秦岭地区羚牛分布点位数据来源包括:(1)红外相机监测和野外巡护数据(381个点位, 2014—2017年);(2)保护区总体规划(94个点位, 2007—2013年)、管理计划(185个点位, 2001—2010年)、科学考察报告中野生动物分布图(3个点位, 2007年);(3)公开发表的文献资料(175个点位, 2009—2012年;图 1)。对于没有矢量化的点位, 利用ArcGIS 10.4(ESRI Inc., Redlands, CA)进行地理校正并数字化物种位点, 共收到838个羚牛点位。由于收到的点位不是系统调查结果, 因此我们对所收集到的点位进行删除, 随机删除同一1 km×1 km栅格内的多个羚牛分布点位, 每个栅格仅保留一个分布点[37—38]。
2.2 环境变量数据来源本研究采用世界气候数据库Worldclim(http://www.worldclim.org)的当前(1950—2000年)和未来气候情景下2050s年(2041—2060年)两期气候数据, 空间分辨率为30″(约1 km)。未来气候数据采用典型浓度路径RCP 4.5排放情景[39], 在该排放情景下, 到2050s年, 全球平均温度将升高0.9—2.0℃, 符合《巴黎气候大会协定》制定的将全球气温升幅控制在2℃以内的目标[4]。其它环境变量数据包括:河流密度、铁路密度、道路密度、居民点密度, 均来源于Openstreet map矢量数据(https://www.openstreetmap.org/), 海拔数据来源于Worldclim数据库30″分辨率的高程数据, 坡度和坡向数据利用ArcGIS 10.4(ESRI Inc., Redlands, CA)空间分析由海拔数据转化而来。由于2050s年的环境变量无法获取, 我们假设当前环境变量在未来保持不变[40]。
利用ArcGIS 10.4将上述所有环境变量数据统一边界, 并采用同一投影坐标系(Asia North Albers Equal Area Conic), 统一栅格空间分辨率为30″尺度。为消除环境变量之间存在的共线性, 采用相关性检验, 相关系数绝对值大于0.6, 则认为两个环境变量存在较强的共线性[41], 只保留1个变量用来构建羚牛分布模型(由SMDtoolbox插件完成[42])。
2.3 适宜生境模型近年来, 基于最大熵理论发展起来的MaxEnt生态模型, 被广泛用于预测物种分布研究。Maxent模型仅需物种分布点位和研究区域的相关环境信息, 就能预测物种分布[43]。因此, 本研究利用Maxent 3.3.3 K模型来预测气候变化背景下秦岭地区羚牛当前和未来(2050s年)两期适宜生境的变化。由于野外调查样点取样存在偏差(如多数物种点位仅在调查样线两侧, 河流、峡谷、高山等区域取样较少), 仅有物种有的分布信息, 缺少物种无的分布信息, 可能会对模型模拟精度产生影响, 需利用同样存在偏差的背景点作为“伪—未发现点”来构建模型, 从而提高模型精度[44]。本研究基于秦岭羚牛分布位点生成10 km缓冲区, 10 km缓冲区外随机生成10000个背景点作为“伪—未发现点”来构建羚牛分布模型。
首先, 将羚牛分布信息和环境变量数据输入Maxent软件界面;然后, 随机选取75%羚牛分布信息作为训练数据来建立模型, 剩余25%分布信息作为检验数据来验证模型;其次, 利用刀切法来检验环境变量用于构建模型的重要贡献性;最后, 模型模拟重复10次, 采用重采样法评估模型表现[45], 并创建环境变化反应曲线, 其余参数设置为默认值。采用受试者工作特征曲线下的面积(AUC)对模型的精度进行评估, AUC值评价标准:失败(0.5—0.6);较差(0.6—0.7);一般(0.7—0.8);满意(0.8—0.9);优秀(0.9—1.00)。
模型输出结果为物种出现概率栅格图, 输出值位于0和1之间, 值越接近1表示潜在分布的可能性越高[45]。本研究采用灵敏度和特异度之和最大值作为阈值[33], 将出现概率高于阈值的栅格作为羚牛潜在适宜生境。基于羚牛家域面积(平均57 km2)[46], 对于面积小于57 km2生境小斑块, 将作为噪点被清除。利用A Mann-Whitney U检验羚牛当前和未来2050s年适宜生境平均海拔变化差异, 用SPSS 19.0(IBM Inc., USA)进行数据分析。
2.4 保护空缺分析保护空缺分析是寻找保护体系之外具有较高保护价值区域的过程, 为保护规划提供指导依据[47]。本研究将秦岭地区羚牛当前和2050s年潜在适宜生境与已建立起来的保护区边界进行叠加, 识别现有保护区体系之外具有较高保护价值的适宜生境, 评估保护区面对未来气候变化的保护成效。
2.5 脆弱性评估本研究通过对比羚牛当前和2050s年适宜生境的变化, 评估气候变化背景下物种脆弱程度, 评估标准如下:(1)脆弱生境:当前适宜生境到2050s年转化为不适宜生境的区域;(2)不变适宜生境:当前和2050s年适宜生境重叠的区域(将成为羚牛躲避未来气候变化的庇护所);(3)新增适宜生境:当前不适宜生境到2050s年转化为适宜生境的区域;(4)不适宜生境:当前和2050s年不适宜生境重叠的区域。
同时, 本研究利用适宜生境变化指数评估气候变化对羚牛的影响[32, 48]:(1)适宜生境变化率(AC);(2)当前适宜生境丧失率(SHc);(3)未来适宜生境增加率(SHf)。公式如下:
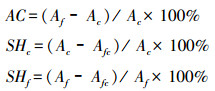
|
式中, Ac:羚牛当前适宜生境总面积;Af:2050s年羚牛适宜生境总面积;Afc:当前和2050s年羚牛适宜生境重叠区域的总面积。
2.6 生态廊道构建本研究将模型输出的羚牛出现概率栅格图进行求导, 作为构建羚牛生态廊道的阻力图层, 阻力值转化成1至100, 计算公式如下[49]:
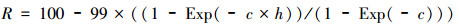
|
式中, R表示阻力图层值, c为阈值(即表示灵敏度和特异度之和最大值), h模型模拟输出栅格图的概率值。借助ArcGIS 10.4平台中Linkage Mapper toolbox(ArcGIS 10.0—10.6 Version)(https://circuitscape.org/linkagemapper/)插件中的最小费用距离(Calculate Least-Cost Corridors and Paths)[50], 构建秦岭地区羚牛适宜生境斑块(面积大于57 km2)之间潜在生态廊道。
3 结果 3.1 物种分布模型最终, 共计559个分布位点和10个环境变量用于构建秦岭地区羚牛分布模型。模型平均训练数据集和验证集的AUC值分别为(0.950±0.001)和(0.943±0.001), 表明模型预测效果优秀。环境因子对模型综合贡献的排序重要性, 从高到低依次为:降水季节性变化(37.5%)、最冷季平气温(28%), 最湿月降水(22.1%)、居民点密度(3.2%)、道路密度(2.8%)、河流密度(1.8%)、铁路密度(1.6%)、年均日较差(1.6%)、坡向(0.9%)和坡度(0.2%)。灵敏度和特异度之和最大值(P=0.154)对应的羚牛分布概率值作为阈值, 不适宜生境(P≤0.154)和适宜生境(P>0.154)。
3.2 适宜生境变化当前气候条件下, 秦岭地区羚牛适宜生境总面积为6473 km2, 主要分布在周至县、城固县、太白县、佛坪县、留坝县、洋县和宁陕县(图 2);到2050s年, 预测秦岭地区羚牛适宜生境总面积为4217 km2, 减少34.85%(AC), 主要分布在太白县、佛坪县、洋县和周至县交界处, 以及宁陕县。未来气候变化将导致秦岭地区羚牛适宜生境向高海拔地区转移, 到2050s年羚牛适宜生境平均海拔(2134±417)m显著(Z=-27.65, P=0.000)高于当前平均海拔(1922±426)m, 转移约210 m。

|
| 图 2 秦岭山系羚牛当前和2050s年适宜生境分布 Fig. 2 Current and the 2050s suitable habitat for golden takin in Qinling Mountains |
保护区空缺分析结果表明, 在当前气候条件下, 秦岭地区已建立的保护区覆盖49.82%羚牛适宜生境, 尚有3248 km2的适宜生境处于保护区之外;到2050s年, 保护区覆盖43.87%羚牛适宜生境, 尚有2367 km2的适宜生境处于保护区之外。到2050s年, 预测板桥(AC=-100%)、娘娘山(AC=-100%)、桑园(AC=-100%)等16个保护区的羚牛适宜生境将会出现不同程度的丧失(表 1), 而平河梁、鹰嘴石和牛背梁保护区的羚牛适宜生境则有所增加(图 2)。
| 自然保护区 Nature reserve |
适宜生境面积 Suitable habitat area/km2 |
适宜生境面积变化率 Percentage of area change /% |
|
| 当前Current | 2050s | ||
| 1-板桥自然保护区Banqiao nature reserve | 198.42 | 0.00 | -100.00 |
| 2-长青自然保护区Changqing nature reserve | 309.45 | 71.06 | -77.04 |
| 3-佛坪自然保护区Foping nature reserve | 298.75 | 138.10 | -53.78 |
| 4-观音山自然保护区Guanyinshan nature reserve | 145.56 | 79.42 | -45.44 |
| 5-汉中-朱鹮自然保护区Hanzhongzhuhuan nature reserve | 10.12 | 0.00 | -100.00 |
| 6-黄柏塬自然保护区Huangbaiyuan nature reserve | 188.93 | 105.24 | -44.30 |
| 7-皇冠山自然保护区Huangguanshan nature reserve | 152.95 | 89.36 | -41.58 |
| 8-老县城自然保护区Laoxiancheng nature reserve | 120.57 | 84.55 | -29.87 |
| 9-摩天岭自然保护区Motianling nature reserve | 35.37 | 0.00 | -100.00 |
| 10-娘娘山自然保护区Niangniangshan nature reserve | 99.21 | 0.00 | -100.00 |
| 11-牛背梁自然保护区Niubeiliang nature reserve | 57.79 | 91.40 | 58.15 |
| 12-盘龙自然保护区Panlong nature reserve | 107.81 | 0.00 | -100.00 |
| 13-平河梁自然保护区Pingheliang nature reserve | 28.77 | 85.67 | 197.81 |
| 14-桑园自然保护区Sangyuan nature reserve | 103.87 | 0.00 | -100.00 |
| 15-太白-牛尾河自然保护区Taibainiuweihe nature reserve | 52.74 | 2.95 | -94.41 |
| 16-太白山自然保护区Taibaishan nature reserve | 537.49 | 504.21 | -6.19 |
| 17-天华山自然保护区Tianhuashan nature reserve | 290.19 | 192.16 | -33.78 |
| 18-鹰嘴石自然保护区Yingzuishi nature reserve | 0.00 | 37.25 | 100.00 |
| 19-周至自然保护区Zhouzhi nature reserve | 487.01 | 368.44 | -24.35 |
模型预测结果表明, 在2050s年气候条件下, 当前分布在太白县、佛坪县、洋县和宁陕县等区域的3490 km2羚牛适宜生境将会成为生境脆弱区域, 丧失53.92%(SHc);共有2983 km2当前和2050s年保持不变适宜生境, 将成为羚牛躲避未来气候变化的庇护所, 主要分布在周至县、太白县和眉县三县交界处, 周至县、佛坪县和宁陕县三县交界处, 以及户县;同时, 2050s年新增1234 km2羚牛适宜生境, 增加29.26%(SHf), 主要分布在周至县北部、宁陕县南部、长安县, 以及秦岭山系东北部的华县和潼关县(图 3)。

|
| 图 3 秦岭山系羚牛适宜生境脆弱评估, 保护空缺和生境廊道 Fig. 3 Vulnerability analysis suitable habitat, protection gaps and habitat corridors of golden takins in Qinling Mountains |
Linkage Mapper利用最小费用距离构建13条潜在生态廊道, 连接秦岭山系羚牛当前适宜生境斑块, 生态廊道总长度288.21 km, 平均长度(22.17±19.64) km。同时, Linkage Mapper构建19条潜在生态廊道, 连接秦岭地区羚牛2050s年适宜生境斑块, 生态廊道总长度586.68 km, 平均长度(30.88±30.11)km。
本研究根据Linkage Mapper构建的当前和未来潜在生态廊道, 提出5条秦岭地区羚牛迁徙潜在廊道(图 3;C1—C5), 提升羚牛适宜栖息地之间的整体连接度。C1廊道有助于栖息在的周至县的羚牛种群和秦岭中部(佛坪县、洋县、太白县)的羚牛种群相互交流;C2—C4廊道有助于栖息在平和梁、皇冠山、牛背梁等保护区的羚牛局域种群逃离未来不适宜气候条件, 扩散到秦岭中部的核心栖息地;C5迁移廊道有助于羚牛种群利用未来新增的适宜生境。
4 讨论本研究较好反映了全球气候变化对秦岭地区濒危物种分布的可能影响, 具重要的典型性、代表性, 为重新审视该区域已建立保护区体系、选址和管理在应对全球气候变化的保护成效提供科学依据。当前, 秦岭地区多数濒危物种分布的区域已纳入国家生态功能功和主体功能区, 在这些区域内开展项目开发利用的可能性较小。未来气候变化可能是导致秦岭地区濒危物种栖息地降级和破碎化的主要威胁因素, 将给当前制定的保护措施带来巨大挑战[32—33, 51]。因此, 开展气候变化背景下秦岭地区濒危物种脆弱性评估工作, 有助于减少气候变化带来的不利影响, 预测濒危物种栖息地未来演替方向, 提前制定适应性(如生态廊道)保护对策, 为羚牛等濒危物种保留必要的生存空间, 最终为人类能够有效应对未来气候变化和生物多样性保护提供强有力的依据。
全球气候变化背景下, 即使在适中的温室气体排放情景下(RCP 4.5), 秦岭地区羚牛分布格局发生了较大的变化。模型结果表明, 气候变暖将导致大量的羚牛当前适宜生境到2050s年转变成不适宜生境, 尤其是部分羚牛活动连通性较低区域(如留坝县、洋县和城固), 气候变化将加快这些区域栖息地破碎化进程, 威胁羚牛种群的生存和繁衍。因此, 建议在这些区域实施前瞻性的保护措施(如生境修复)来缓解气候变化带来的不利影响, 同时构建生境廊道, 促进活动这些区域的羚牛种群迁徙到秦岭中部的核心栖息地。模型结果预测宁陕县、柞水县羚牛适宜生境将会增加, 然而人类活动和自然景观(如居民区、道路和河流)阻隔羚牛扩散到这些区域[52—53]。因此, 在这些区域开展以促进生境恢复为目标的森林经营或生态系统保护等项目, 提升羚牛栖息地规模和连接水平, 来援助更多的羚牛种群扩散到未来潜在的适宜生境区(如皇冠山和平和梁已有羚牛较小种群, 有力证明这些区域适宜羚牛生存[54—55])。同时, 模型结果预测在周至县北部、秦岭东北部的华县和潼关县未来将会有羚牛适宜生境增加, 这些新增加的适宜生境距离当前羚牛分布区域较远, 不适宜构建生态廊道来援助羚牛迁徙这些区域, 但可考虑作为羚牛迁地保护、繁殖救护的选址。
长期以来, 保护区是我国生物多样性保护和管理的主体, 在生物多样性保护中发挥极其重要的作用[56]。当前, 我国许多保护区都能感受到气候变化带来的影响, 但多数保护区没有开展系统生物多样性监测, 缺乏主动适应气候变化的对策[57]。秦岭地区保护区在规划和设计时, 主要根据生物多样性自然性、典型性、稀有性等特征, 针对栖息在这些区域的大熊猫、川金丝猴、羚牛等濒危物种建立保护区, 受当时条件的限制, 规划这些保护区未能充分考虑未来气候变化的影响。因此, 面对未来气候变化, 这些保护区的保护成效可能会降低, 甚至丧失。研究结果表明, 秦岭地区保护区体系保护羚牛适宜栖息地不充分, 仅覆盖了49.82%的当前羚牛适宜栖息地, 以及43.87%的未来适宜生境, 尚有大量的适宜栖息地处于保护区之外。尤其是秦岭西南地区的保护区(如娘娘山、板桥、盘龙等), 预测未来气候变化将导致这些保护区内的羚牛适宜生境几乎全部丧失, 失去对羚牛的有效保护功能。当前, 需加强位于宁陕县南部(皇冠山和平河梁)和柞水县(牛背梁)的保护区应对未来气候变化的适应能力建设, 因为这些区域已有羚牛种群活动[54—55], 但与秦岭地区整体保护区体系相分隔, 且预测未来这些保护区内及周边羚牛适宜生境将会增加。
脆弱性评估有助于找出气候变化导致哪些区域适宜生境发生变化, 为制定适应气候变化保护对策提供建议[58]。适应性保护对策应该注重生境脆弱区域, 因为气候变化给这些区域带来较大影响, 适宜生境将会丧失且破碎化, 对栖息在这些区域的羚牛种群构成严重威胁。同时, 应该关注适宜生境没有发生变化的区域, 这些区域是羚牛躲避未来气候变化的庇护所[59], 有助于维持种群在此区域长期生存。
5 保护对策秦岭地区是全球生物多样性最具代表性的区域之一, 保护好秦岭是我国对世界生物多样性保护的贡献和责任。《生物多样性保护战略与行动计划(2011—2030年)》将气候变化背景下濒危物种脆弱性评估工作, 列为优先行动计划[31]。然而, 目前我国脆弱性评估工作仍处于探讨、介绍的层面, 对评估方法的认识存在不足。本研究开展秦岭地区羚牛生境脆弱性评估工作, 提出羚牛适应未来气候变化的保护性对策, 以期为该区域羚牛保护和国家公园建设提供指导依据。基于研究成果, 就羚牛应对未来气候变化的保护工作, 提出以下几点建议:
1) 考虑将适宜生境纳入国家公园
羚牛作为大熊猫重要伴生物种, 其适宜生境也是大熊猫生存的重要栖息地。当前秦岭地区保护区保护羚牛栖息地不充分, 在周至县、宁陕县存在大量的羚牛适宜栖息地处于保护空缺状态(图 3), 需进一步完善保护区体系分布。国家公园试点为重新审视秦岭地区已建立保护区体系提供了前所未有的机遇。因此, 我们建议除将当前已建立起的保护区范围纳入国家公园外, 考虑将羚牛适宜生境保护空缺区域一同纳入国家公园范围, 提升国家公园生态系统完整性。
2) 建立生态廊道
栖息地破碎使秦岭地区羚牛形成了多个局域种群, 提前规划羚牛迁徙到庇护所的生态廊道, 减少迁移阻障, 有助促进种群相互交流和扩散。生态廊道构建及选址(图 3;C1—C5)见本文3.5部分。
3) 增强物种适应气候变化能力
减少非气候因素带来的影响(如外来人类活动干扰、环境污染、入侵物种、疾病等), 有助于增强羚牛适应气候变化能力。如春秋季节是羚牛垂直迁徙季节, 也是人类活动较多的季节(如采药、放牧、挖竹笋等), 提前预判人类活动集中区域, 加大巡护力度。
4) 加强野生动物监测
在制定总体规划时, 多数保护区没有考虑未来气候变化给野生动物带来影响。同样, 科研人员、管理人员也没有完全明白野生动物如何响应气候变化, 该采取怎样的措施来缓解气候变化带来的不利影响。因此, 建议国家公园建立长期系统的野生动物监测计划, 全面收集气候变化影响野生动物的数据。
6 总结本研究以秦岭地区羚牛作为研究对象, 分析气候变化对其栖息地当前和2050s年分布格局的影响, 讨论已建立的保护区体系面对未来气候变化的保护成效及其空缺。期待国家公园规划时考虑气候变化带来的影响, 制定长期的动态的生物多样监测计划。
| [1] |
Garcia R A, Cabeza M, Rahbek C, Araújo M B. Multiple dimensions of climate change and their implications for biodiversity. Science, 2014, 344(6183): 1247579. DOI:10.1126/science.1247579 |
| [2] |
Bellard C, Bertelsmeier C, Leadley P, Thuiller W, Courchamp F. Impacts of climate change on the future of biodiversity. Ecology Letters, 2012, 15(4): 365-377. DOI:10.1111/j.1461-0248.2011.01736.x |
| [3] |
Weiskopf S R, Rubenstein M A, Crozier L G, Gaichas S, Griffis R, Halofsky J E, Hyde K J W, Morelli T L, Morisette J T, Muñoz R C, Pershing A J, Peterson D L, Poudel R, Staudinger M D, Sutton-Grier A E, Thompson L, Vose J, Weltzin J F, Whyte K P. Climate change effects on biodiversity, ecosystems, ecosystem services, and natural resource management in the United States. Science of the Total Environment, 2020, 733: 137782. DOI:10.1016/j.scitotenv.2020.137782 |
| [4] |
United Nations Framework Convention on Climate Change (UNFCCC). Adoption of the Paris agreement. Report No. FCCC/CP/2015/L. 9/Rev. 1.2015. http://unfccc.int/resource/docs/2015/cop21/eng/l09r01.pdf.
|
| [5] |
Thomas C D, Cameron A, Green R E, Bakkenes M, Beaumont L J, Collingham Y C, Erasmus B F N, de Siqueira M F, Grainger A, Hannah L, Hughes L, Huntley B, van Jaarsveld A S, Midgley G F, Miles L, Ortega-Huerta M A, Peterson A T, Phillips O L, Williams S E. Extinction risk from climate change. Nature, 2004, 427(6970): 145-148. DOI:10.1038/nature02121 |
| [6] |
Urban M C. Climate change. Accelerating extinction risk from climate change. Science, 2015, 348(6234): 571-573. DOI:10.1126/science.aaa4984 |
| [7] |
Pereira H M, Leadley P W, Proença V, Alkemade R, Scharlemann J P W, Fernandez-Manjarrés J F, Araújo M B, Balvanera P, Biggs R, Cheung W W L, Chini L, Cooper H D, Gilman E L, Guénette S, Hurtt G C, Huntington H P, Mace G M, Oberdorff T, Revenga C, Rodrigues P, Scholes R J, Sumaila U R, Walpole M. Scenarios for global biodiversity in the 21st century. Science, 2010, 330(6010): 1496-1501. DOI:10.1126/science.1196624 |
| [8] |
李佳, 刘芳, 张宇, 薛亚东, 李迪强. 气候变化背景下野生动物脆弱性评估方法研究进展. 生态学报, 2017, 37(20): 6656-6667. |
| [9] |
Levinsky I, Skov F, Svenning J C, Rahbek C. Potential impacts of climate change on the distributions and diversity patterns of European mammals. Biodiversity and Conservation, 2007, 16(13): 3803-3816. DOI:10.1007/s10531-007-9181-7 |
| [10] |
Foden W B, Butchart S H M, Stuart S N, Vié J C, Akçakaya H R, Angulo A, DeVantier L M, Gutsche A, Turak E, Cao L, Donner S D, Katariya V, Bernard R, Holland R A, Hughes A F, O'Hanlon S E, Garnett S T, Şekercio Ç H, Mace G M. Identifying the world's most climate change vulnerable species: a systematic trait-based assessment of all birds, amphibians and corals. PLoS One, 2013, 8(6): e65427. DOI:10.1371/journal.pone.0065427 |
| [11] |
Foden W B, Young B E, Akçakaya H R, Garcia R A, Hoffmann A A, Stein B A, Thomas C D, Wheatley C J, Bickford D, Carr J A, Hole D G, Martin T G, Pacifici M, Pearce-Higgins J W, Platts P J, Visconti P, Watson J E M, Huntley B. Climate change vulnerability assessment of species. WIREs Climate Change, 2019, 10(1): e551. |
| [12] |
Rowland E L, Davison J E, Graumlich L J. Approaches to evaluating climate change impacts on species: a guide to initiating the adaptation planning process. Environmental Management, 2011, 47(3): 322-337. |
| [13] |
Pacifici M, Foden W B, Visconti P, Watson J E M, Butchart S H M, Kovacs K M, Scheffers B R, Hole D G, Martin T G, Akçakaya H R, Corlett R T, Huntley B, Bickford D, Carr J A, Hoffmann A A, Midgley G F, Pearce-Kelly P, Pearson R G, Williams S E, Willis S G, Young B, Rondinini C. Assessing species vulnerability to climate change. Nature Climate Change, 2015, 5(3): 215-224. DOI:10.1038/nclimate2448 |
| [14] |
Arribas P, Abellán P, Velasco J, Bilton D T, Millán A, Sánchez-Fernández D. Evaluating drivers of vulnerability to climate change: a guide for insect conservation strategies. Global Change Biology, 2012, 18(7): 2135-2146. DOI:10.1111/j.1365-2486.2012.02691.x |
| [15] |
Sudo K, Watanabe K, Yotsukura N, Nakaoka M. Predictions of kelp distribution shifts along the northern coast of Japan. Ecological Research, 2020, 35(1): 47-60. DOI:10.1111/1440-1703.12053 |
| [16] |
Guisan A, Thuiller W. Predicting species distribution: offering more than simple habitat models. Ecology Letters, 2005, 8(9): 993-1009. DOI:10.1111/j.1461-0248.2005.00792.x |
| [17] |
Carvalho S B, Brito J C, Crespo E J, Possingham H P. From climate change predictions to actions-conserving vulnerable animal groups in hotspots at a regional scale. Global Change Biology, 2010, 16(12): 3257-3270. DOI:10.1111/j.1365-2486.2010.02212.x |
| [18] |
吴家炎. 中国羚牛分类、分布的研究. 动物学研究, 1986, 7(2): 167-175. |
| [19] |
Sangay T, Rajaratnam R, Vernes K. Current distribution and conservation status of Bhutan Takin Budorcas whitei Lydekker, 1907 (Artiodactyla: Bovidae). Journal of Threatened Taxa, 2016, 8(14): 9630. DOI:10.11609/jott.2925.8.14.9630-9637 |
| [20] |
Wang T J, Skidmore A K, Zeng Z G, Beck P S A, Si Y L, Song Y L, Liu X H, Prins H H T. Migration patterns of two endangered sympatric species from a remote sensing perspective. Photogrammetric Engineering & Remote Sensing, 2010, 76(12): 1343-1352. |
| [21] |
Guan T P, Ge B M, McShea W J, Li S, Song Y L, Stewart C M. Seasonal migration by a large forest ungulate: a study on takin (Budorcas taxicolor) in Sichuan Province, China. European Journal of Wildlife Research, 2013, 59(1): 81-91. DOI:10.1007/s10344-012-0650-2 |
| [22] |
Zeng Z G, Beck P S A, Wang T J, Skidmore A K, Song Y L, Gong H S, Prins H H T. Effects of plant phenology and solar radiation on seasonal movement of golden takin in the Qinling Mountains, China. Journal of Mammalogy, 2010, 91(1): 92-100. DOI:10.1644/08-MAMM-A-390R.1. |
| [23] |
曾治高, 宋延龄. 秦岭羚牛春夏季昼夜活动节律与时间分配. 兽类学报, 2001, 21(1): 7-13. DOI:10.3969/j.issn.1000-1050.2001.01.002 |
| [24] |
曾治高, 宋延龄, 钟文勤, 巩会生, 张坚, 党高第. 秦岭羚牛的食性. 动物学杂志, 2001, 36(3): 36-44. DOI:10.3969/j.issn.0250-3263.2001.03.010 |
| [25] |
王学志, 宋延龄, 曾治高, 巩会生, 赵德怀, 赵纳勋. 秦岭雄性羚牛的发情行为与其社会状态的关系. 兽类学报, 2006, 26(1): 33-37. DOI:10.3969/j.issn.1000-1050.2006.01.006 |
| [26] |
Li J, Xue Y D, Zhang Y, Dong W, Shan G Y, Sun R Q, Hacker C, Wu B, Li D Q. Spatial and temporal activity patterns of Golden takin (Budorcas taxicolor bedfordi) recorded by camera trapping. PeerJ, 2020, 8: e10353. DOI:10.7717/peerj.10353 |
| [27] |
Song Y L, Smith A T, & MacKinnon, J. Budorcas taxicolor. The IUCN Red List of ThreatenedSpecies. e. T3160A9643719. http://dx.doi.org/10.2305/IUCN.UK.2008.RLTS.T3160A9643719.en. 2008. Downloaded on 23 July 2019.
|
| [28] |
Li Y, Viña A, Yang W, Chen X D, Zhang J D, Ouyang Z Y, Liang Z, Liu J G. Effects of conservation policies on forest cover change in giant panda habitat regions, China. Land Use Policy, 2013, 33(1): 42-53. |
| [29] |
Zhang P, Shao G, Zhao G, le Master D C, Parker G R, Dunning J B Jr, Li Q. China' s forest policy for the 21st century. Science, 2000, 288(5474): 2135-2136. DOI:10.1126/science.288.5474.2135 |
| [30] |
袁朝晖, 孙建存. 长青保护区大熊猫、金丝猴、羚牛种群数量监测报告. 陕西师范大学学报: 自然科学版, 2007, 35(S1): 100-103. |
| [31] |
环境保护部. 中国生物多样性保护战略与行动计划: 2011-2030. 北京: 中国环境科学出版社, 2011.
|
| [32] |
Li J, Liu F, Xue Y D, Zhang Y, Li D Q. Assessing vulnerability of giant pandas to climate change in the Qinling Mountains of China. Ecology and Evolution, 2017, 7(11): 4003-4015. DOI:10.1002/ece3.2981 |
| [33] |
Li J, Li D Q, Xue Y D, Wu B, He X J, Liu F. Identifying potential refugia and corridors under climate change: a case study of endangered Sichuan golden monkey (Rhinopithecus roxellana) in Qinling Mountains, China. American Journal of Primatology, 2018, 80(11). |
| [34] |
李佳. 秦岭地区濒危物种对气候变化的响应及脆弱性评估[D]. 北京: 中国林业科学研究院, 2017.
|
| [35] |
赵纳勋, 张希明, 董伟. 秦岭自然观察手册. 北京: 中国大百科全书出版社, 2014.
|
| [36] |
国家林业局. 第四次大熊猫调查报告发布. 国家林业局, 2015.
|
| [37] |
Kramer-Schadt S, Niedballa J, Pilgrim J D, Schröder B, Lindenborn J, Reinfelder V, Stillfried M, Heckmann I, Scharf A K, Augeri D M, Cheyne S M, Hearn A J, Ross J, MacDonald D W, Mathai J, Eaton J, Marshall A J, Semiadi G, Rustam R, Bernard H, Alfred R, Samejima H, Duckworth J W, Breitenmoser-Wuersten C, Belant J L, Hofer H, Wilting A. The importance of correcting for sampling bias in MaxEnt species distribution models. Diversity and Distributions, 2013, 19(11): 1366-1379. DOI:10.1111/ddi.12096 |
| [38] |
Radosavljevic A, Anderson R P. Making better Maxent models of species distributions: complexity, overfitting and evaluation. Journal of Biogeography, 2014, 41(4): 629-643. DOI:10.1111/jbi.12227 |
| [39] |
Baek H J, Lee J, Lee H S, Hyun Y K, Cho C, Kwon W T, Marzin C, Gan S Y, Kim M J, Choi D H, Lee J, Lee J, Boo K O, Kang H S, Byun Y H. Climate change in the 21st century simulated by HadGEM2-AO under representative concentration pathways. Asia-Pacific Journal of Atmospheric Sciences, 2013, 49(5): 603-618. DOI:10.1007/s13143-013-0053-7 |
| [40] |
Stanton J C, Pearson R G, Horning N, Ersts P, Reşit Akçakaya H. Combining static and dynamic variables in species distribution models under climate change. Methods in Ecology and Evolution, 2012, 3(2): 349-357. DOI:10.1111/j.2041-210X.2011.00157.x |
| [41] |
Cord A F, Klein D, Mora F, Dech S. Comparing the suitability of classified land cover data and remote sensing variables for modeling distribution patterns of plants. Ecological Modelling, 2014, 272: 129-140. DOI:10.1016/j.ecolmodel.2013.09.011 |
| [42] |
Brown J L. SDMtoolbox: a python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. Methods in Ecology and Evolution, 2014, 5(7): 694-700. DOI:10.1111/2041-210X.12200 |
| [43] |
Elith J, Phillips S J, Hastie T, Dudík M, Chee Y E, Yates C J. A statistical explanation of MaxEnt for ecologists. Diversity and Distributions, 2011, 17(1): 43-57. DOI:10.1111/j.1472-4642.2010.00725.x |
| [44] |
Phillips S J, Dudík M, Elith J, Graham C H, Lehmann A, Leathwick J, Ferrier S. Sample selection bias and presence-only distribution models: implications for background and pseudo-absence data. Ecological Applications: a Publication of the Ecological Society of America, 2009, 19(1): 181-197. DOI:10.1890/07-2153.1 |
| [45] |
Wisz M S, Hijmans R J, Li J, Peterson A T, Graham C H, Guisan A. Effects of sample size on the performance of species distribution models. Diversity and Distributions, 2008, 14(5): 763-773. DOI:10.1111/j.1472-4642.2008.00482.x |
| [46] |
宋延龄, 曾治高, 张坚, 王学杰, 巩会生, 王宽武. 秦岭羚牛的家域研究. 兽类学报, 2000, 20(4): 241-249. DOI:10.3969/j.issn.1000-1050.2000.04.001 |
| [47] |
Scott J M, Davis F, Csuti B, Noss R, Groves C, Anderson H, Caicco S, Edwards T, Ulliman J, Wright R G. Gap analysis: a geographic approach to protection of biological diversity. Wildlife Monographs, 1993, 123: 3-41. |
| [48] |
Thuiller W, Lavorel S, Araújo M B. Niche properties and geographical extent as predictors of species sensitivity to climate change. Global Ecology and Biogeography, 2005, 14(4): 347-357. DOI:10.1111/j.1466-822X.2005.00162.x |
| [49] |
Keeley A T H, Beier P, Gagnon J W. Estimating landscape resistance from habitat suitability: effects of data source and nonlinearities. Landscape Ecology, 2016, 31(9): 2151-2162. DOI:10.1007/s10980-016-0387-5 |
| [50] |
McRae B H, Kavanagh D M. Linkage Mapper Connectivity Analysis Software. The Nature Conservancy, Seattle WA. 2011. Available at: http://www.circuitscape.org/linkagemapper.
|
| [51] |
Shen G Z, Pimm S L, Feng C Y, Ren G F, Liu Y P, Xu W T, Li J Q, Si X F, Xie Z Q. Climate change challenges the current conservation strategy for the giant panda. Biological Conservation, 2015, 190: 43-50. DOI:10.1016/j.biocon.2015.05.004 |
| [52] |
Fan J T, Li J S, Quan Z J, Wu X P, Hu L L, Yang Q P. Impact of road construction on giant panda's habitat and its carrying capacity in Qinling Mountains. Acta Ecologica Sinica, 2011, 31(3): 145-149. DOI:10.1016/j.chnaes.2011.03.003 |
| [53] |
Fan J T, Li J S, Xia R, Hu L L, Wu X P, Li G. Assessing the impact of climate change on the habitat distribution of the giant panda in the Qinling Mountains of China. Ecological Modelling, 2014, 274: 12-20. DOI:10.1016/j.ecolmodel.2013.11.023 |
| [54] |
陈文贵, 罗磊, 刘小军, 田宁朝, 杨实海. 陕西皇冠山保护区鸟兽区系研究. 陕西林业科技, 2009, 2: 17-25. DOI:10.3969/j.issn.1001-2117.2009.03.005 |
| [55] |
孟祥明. 陕西平河梁省级自然保护区哺乳动物资源分布与保护[D]. 杨凌: 西北农林科技大学, 2008.
|
| [56] |
李迪强. 全国林业系统自然保护区体系规划研究. 北京: 中国大地出版社, 2003.
|
| [57] |
Araújo M B, Alagador D, Cabeza M, Nogués-Bravo D, Thuiller W. Climate change threatens European conservation areas. Ecology Letters, 2011, 14(5): 484-492. DOI:10.1111/j.1461-0248.2011.01610.x |
| [58] |
Pacifici M, Foden W B, Visconti P, Watson J E M, Butchart S H M, Kovacs K M, Scheffers B R, Hole D G, Martin T G, Akçakaya H R, Corlett R T, Huntley B, Bickford D, Carr J A, Hoffmann A A, Midgley G F, Pearce-Kelly P, Pearson R G, Williams S E, Willis S G, Young B, Rondinini C. Assessing species vulnerability to climate change. Nature Climate Change, 2015, 5(3): 215-224. DOI:10.1038/nclimate2448 |
| [59] |
Araújo M B, Alagador D, Cabeza M, Nogués-Bravo D, Thuiller W. Climate change threatens European conservation areas. Ecology Letters, 2011, 14(5): 484-492. DOI:10.1111/j.1461-0248.2011.01610.x |