 2022, Vol. 42
2022, Vol. 42文章信息
- 陈翔, 刘树林, 彭飞, 王涛
- CHEN Xiang, LIU Shulin, PENG Fei, WANG Tao
- 中分辨率遥感像元尺度生物土壤结皮覆盖与植被及土壤间的交互关系
- Interactions among biological soil crust, vegetation, and soil on medium resolution pixel scale of remote sensing
- 生态学报. 2022, 42(18): 7336-7348
- Acta Ecologica Sinica. 2022, 42(18): 7336-7348
- http://dx.doi.org/10.5846/stxb202107242000
-
文章历史
- 收稿日期: 2021-07-24
- 网络出版日期: 2022-04-20

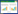




2. 甘肃省绿洲资源环境与可持续发展重点实验室, 兰州 730070;
3. 中国科学院西北生态环境资源研究院, 沙漠与沙漠化重点实验室, 兰州 730000
2. Key Laboratory of Resource Environment and Sustainable Development of Oasis, Gansu Province, Lanzhou 730070, China;
3. Key Laboratory of Desert and Desertification, Northwest Institute of Eco-Environment and Resources, Chinese Academy of Sciences, Lanzhou 730000, China
生物土壤结皮(Biological Soil Crust, BSC)是由细菌界的蓝细菌、真菌界的地衣以及植物界的苔类、藓类及藻类为优势的微型生物与沙土颗粒缠绕在一起, 所形成的皮壳状结构[1—2]。由于大多数荒漠生态系统受水分的限制, 地表无法支撑大面积且连续分布的高等植被群落, 植被群落斑块状的分布为BSC的发育提供了适宜的环境, 使其覆盖度占地表活体覆盖面积的40%以上, 在有些地区甚至已达到70%以上[2]。作为干旱半干旱地区健康生态系统的重要组成部分, BSC具有固碳[3—5]、固氮[6—7]、减少土壤流失[8]、改善土壤条件[2]等多种生态功能, 这些生态功能与BSC的组成及覆盖度密切相关[1—2, 9]。因此, 探索BSC覆盖度的环境影响因子及其之间的关系研究是沙区生态恢复和沙化土地治理的关键[2, 10], 也为更好地评估和保护脆弱生态系统奠定基础[11—12]。
目前已有一些研究探索了影响BSC拓殖与发展的植被及土壤性质[2, 13—15]。Bowker[14]认为, BSC覆盖度与维管植物生长间的关系复杂且相互影响。李新荣等[2, 9]对BSC的物种组成及生物量在景观尺度上与环境因子的相关性进行了系统深入的研究。Kidron等[16]发现植被通过遮荫、影响土壤水分含量来间接地影响BSC群落的丰富度、物种组成和分布格局。Danin和Ganor[17]研究发现, BSC中藓类种的丰富度与多年生植物覆盖度和土壤有机质呈正相关关系。Li等[18]和Su等[6]的结果均发现, 苔藓主导BSC的多样性与覆盖度和一年生植物覆盖度负相关, 藻类主导BSC与一年生植物覆盖度则呈正相关关系。Flechtner[19]的研究结果显示, 地衣主导BSC的物种多样性和覆盖度均与土壤黏粒、粉粒和碳酸钙积累呈正相关关系。何芳兰等[20]研究发现, 民勤绿洲边缘沙丘BSC形成及发育对0—1cm层土壤有机质有显著影响, 藻类、地衣、藓类是提高土壤养分含量的主要因子。Li等[15]认为沙坡头地区不同固沙年代植被区表土层化学元素含量的变化与BSC形成有关。然而, 这些研究方法多采用简单的统计分析、线性拟合、传统回归模型等方法, 大多侧重于孤立地分析BSC与植被、土壤间的单向因果关系, 未能从整体上阐明三者间的交互关系。
由此, 一些学者尝试利用模型来描述BSC分布与植被、土壤间的关系[21—22]。Read等[20]利用增强回归树(boosted regression tree, BRT)方法提出不同土壤质地中BSC的分布模型。Bowker等[14]提出地衣与苔藓主导BSC分布的概念模型并发现BSC的分布显著受土壤养分和质地的影响。Li等[22]基于微空间尺度上BSC分布格局与土壤环境因子间的关系, 提出BSC分布与微地形、土壤质地和降尘间的关系概念模型。此类研究多为小尺度上的定性描述, 对遥感像元尺度上影响BSC覆盖度的植被及土壤性质缺乏定量系统性研究, 尤其以一个区域为代表的综合分析研究尚少见报道。此外, 李新荣等[9]认为, 植被、土壤与BSC形成和发展之间的相互作用很复杂, 且影响BSC的环境因素有明显的尺度差异性。因此, 从遥感像元尺度上综合分析毛乌素沙地生物土壤结皮覆盖度与植被、土壤性质间的交互关系对遥感大尺度探测BSC具有重要意义。
本研究以毛乌素沙地为研究区, 基于实地样方调查BSC覆盖度与植被覆盖度, 以及实测土壤有机质、粒度、pH值、盐度数据, 运用结构方程模型(Structural Equation Modeling, SEM)构建多个因变量路径分析。在系统分析BSC覆盖度与植被、土壤性质间交互关系的基础上, 找出影响BSC覆盖度的关键因子和路径, 为评估、保护和修复BSC、制定有效的荒漠自然生态系统恢复政策措施提供科学依据。
1 研究区概况研究区位于中国北部毛乌素沙地(37°28′—39°49′ N, 106°57′—110°37′ E), 面积约4×104 km2, 海拔875—1685 m(图 1)。在行政区划上, 毛乌素沙地包括内蒙古自治区鄂尔多斯南部、陕西省榆林市榆阳区北部及宁夏回族自治区盐池县东北部。年均气温6.0—8.5℃[23], 降水主要集中在7—9月(特别是8月), 占全年总降水量的60%—75%。年潜在蒸发量为2300 mm, 是年均降水量的6倍。冬、春和秋季盛行西北风, 夏季盛行东南风[23]。土壤类型为松散风沙土, 且土地贫瘠, 易受风蚀影响[24]。土壤pH值大多介于7.7—9.2之间(图 2), 多为碱性土壤, 并长期处于干燥状态。毛乌素沙地大部分地区位于栗钙土干草原地带, 向西北过渡为棕钙土半荒漠地带, 向东南过渡为黄土高原温暖带黑垆土地带, 分布上表现为由东北至西南向的水平地带性变化。草原地带的土壤以沙土为主, 地势高处也有少量黄绵土分布。毛乌素沙地的土壤总体上形成年限短, 成土过程较弱且土层薄;在机械组成上以沙粒为主(图 2);土壤肥力较差, 土壤有机质含量较低;土壤钙质含量较高[25]。植被覆盖度总体由东向西呈减少趋势, 大部分区域植被覆盖度在30%以下, 沙地腹地依然明显存在极低植被覆盖的流沙区[26]。毛乌素沙地80%以上为沙质草地, 优势种为油蒿[27]。苔藓主导的BSC在油蒿群落中广泛存在, 是沙丘固定的重要标志之一[27]。其他自然植被类型, 包括草原、草甸和灌木等也分布于毛乌素沙地。农田主要分布于河流沿岸或散布于沙地、草地、人工林和灌木周边[28]。毛乌素沙地位于典型草原和沙漠过渡地带, 对气候变化和土地利用变化非常敏感, 属半干旱大陆性气候[28]。本研究于2018年8月至2019年9月的植被生长期, 在毛乌素沙地进行5次实地样方调查, 随机选取250个30 m×30 m样方(图 1)。

|
| 图 1 毛乌素沙地及采样点位置 Fig. 1 Mu Us Sandy Land and filed work sites |

|
| 图 2 毛乌素沙地生物土壤结皮、植被覆盖度及供试土壤条件分布 Fig. 2 Distribution of biological soil crust, vegetation coverage and tested soil conditions in Mu Us Sandy Land BSC:生物土壤结皮Biological soil crust |
BSC覆盖度及植被覆盖度数据均由无人机配合样方调查获取, 样方调查时间分别为2018年8月16日至2018年8月28日、2019年5月9日至2019年5月17日、2019年7月6日至2019年7月7日、2019年8月5日、2019年9月15日至2019年9月22日。样方调查选取景观均一的地块使用粗绳围成30 m×30 m的样方(图 3), 手持1 m×1 m铁丝方框(图 3), 沿S型路线走过样方, 以铁丝方框为单位计数BSC与植被面积, 推算出BSC与植被的覆盖度[29]。另外, 由无人机(Phantom 4 Pro, DJI)配合, 先飞至80 m高度拍摄完整样方照片, 再下降至8 m高度沿S型路线拍摄样方内局部照片, 并用数码相机(D3000, Nikon)拍摄样方内景观照片(图 3), 用Adobe Photoshop CC 2019来核对并校正BSC覆盖度与植被覆盖度[30]。图 3展示了毛乌素沙地内一处以苔藓为主导的BSC斑块, 厚度约1 cm, 生长于油蒿周围。

|
| 图 3 无人机配合的样方调查 Fig. 3 Unmanned aerial vehicle cooperation in quadrat survey |
在每个30 m×30 m的样方内, 随机采集3个0—10 cm表层土壤样品[31], 共计750个土样。每个土壤样品重约500 g左右, 放入塑料自封袋后带回实验室。在实验室将采回的土样放在塑料布上, 摊成薄薄的一层, 在室内通风阴干[31]。自然风干后, 拣去动植物残体。然后, 将每个样方内3个土壤样品混合均匀后随机分成4份, 分别用于测定土壤有机质、pH值、盐度和土壤粒度指标。
本研究采用重铬酸钾容量法(外加热法)测量土壤有机质[31];采用激光粒度仪(Mastersizer 3000)对土壤进行粒度分析;采用酸度计(上海雷磁精密酸度计PHSJ-4F)检测土壤pH值[31];采用电导率仪(上海雷磁DDSJ-308F型电导率仪)检测土壤盐度[31]。其中, 粒度分析后用图解法计算得到平均粒径Mz[32], 公式如下:
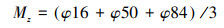
|
(1) |
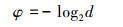
|
(2) |
式中, φ16、φ50、φ84分别为粒度分布相应的分位数, d为颗粒直径, Mz单位为φ, 因此, 平均粒径Mz越大, 说明整体颗粒越细。另外, 根据美国农业部(United States Department of Agriculture, USDA)土壤粒径分类系统7级标准[33], 测量了土壤颗粒物密度:< 2 μm(黏粒)、2—50 μm(粉粒)、50—100 μm(极细砂)、100—250 μm(细砂)、250—500 μm(中砂)、500—1000 μm(粗砂)和1000—2000 μm(极粗砂), 本文将黏粒和粉粒占比之和定义为细颗粒占比, 将粗砂与极粗砂之和定义为粗颗粒占比。最后, 计算每个样方3个重复的平均值进行后续分析。以上数据处理均在Excel 2016中完成。
2.2 研究方法 2.2.1 Pearson相关分析采用IBM SPSS Statistics 26对BSC覆盖度、植被覆盖度、土壤pH值、盐度、有机质、平均粒径、细颗粒占比和粗颗粒占比进行Pearson相关分析。
2.2.2 结构方程模型(Structural Equation Modeling, SEM)路径分析SEM是一种验证性的方法, 须有经验法则支持并在理论引导的前提下构建假设模型图, 即一种呈现客观状态的数学模型[34]。本研究选取SEM中的观察变量路径分析(Path Analysis with Observed Variables, PA-OV)模型, 该分析方法是一种没有包含任何潜在变量的结构方程模型, 模型中所有变量均为测量指标变量[34]。另外, 由于假设BSC覆盖度、植被覆盖度与土壤有机质间存在因果关系, 因此选取路径分析中的非递归模型(Nonrecursive Model)[34—36]。非递归模型特点在于残差间假设有相关存在, 该模型内的因果关系箭头是两个方向相反的单向箭头, 第一个变量直接影响到第二个变量, 而第二个变量也直接影响到第一个变量, 互为因果关系[34]。
为选取SEM的估计方法, 首先对所有数据进行正态性评估(表 1), 在SEM分析中, 如果样本数据的偏度系数>3、峰度系数>8, 则可能偏离正态分布, 尤其当峰度系数>20时, 表示数据变量峰度与正态峰差异极大[37]。本研究中, 盐度数据明显偏离正态分布, 且数据样本量为250, 不属于大样本(样本量>1000), 故选用适合小样本且非正态分布的一般化最小平方法(Generalized Least Squares, GLS)进行参数估计[34]。
| 变量 Variable |
最小值 Minimum value |
最大值 Maximum value |
偏度值 Skewness value |
峰度值 Kurtosis |
| BSC覆盖度BSC coverage | 0.000 | 0.800 | 1.217 | 0.042 |
| 植被覆盖度Vegetation coverage | 0.000 | 0.950 | 0.585 | -0.247 |
| pH值pH value | 6.803 | 10.320 | -0.024 | 1.697 |
| 盐度Salinity | 0.010 | 1.410 | 5.871 | 37.758 |
| 有机质Organic matter | 0.050 | 4.537 | 2.534 | 12.060 |
| 平均粒径Average particle size | 1.630 | 5.227 | 0.904 | 1.640 |
| 粗颗粒占比Proportion of coarse particles | 0.000 | 22.193 | 1.709 | 3.401 |
| 细颗粒占比Proportion of fine particles | 0.000 | 65.500 | 2.036 | 6.478 |
| BSC:生物土壤结皮Biological soil crust | ||||
模型适配度的指标主要选取卡方值(χ2)、卡方自由度比(χ2/df)、渐进残差均方和平方根(Root Mean Square Error of Approximation, RMSEA)、良适性适配指标(Goodness-of-Fit Index, GFI)、调整后良适性适配指标(Adjusted Goodness-of-Fit Index, AGFI)以及临界样本数(Critical N, CN), 计算公示如下[34]:

|
(3) |

|
(4) |

|
(5) |
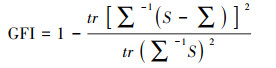
|
(6) |
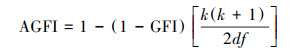
|
(7) |
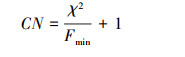
|
(8) |
式中, n为样本数, tr(…)为矩阵内对角线元素之和, S为实际样本数据导出的协方差矩阵, 

为判断模型是否简约, 选取5种模型简约适配度的指标[34]:简约适配度指数(Parsimony Goodness of Fit Index, PGFI)、Akaike讯息效标(Akaike Information Criteria, AIC)、Browne-Cudeck效标(Browne-Cudeck Criterion, BCC)、Bayes讯息效标(Bayes Information Criterion, BIC)和一致性Akaike讯息效标(Consistent Akaike Information Criteria, CAIC)。
假设BSC覆盖度与植被覆盖度、土壤pH值、盐度、有机质、平均粒径、细颗粒占比和粗颗粒占比间存在图 4所示因果关系, 利用模型界定搜寻遍历32768个模型, 最终选出修正后最优模型架构。以上所有操作均在IBM SPSS AMOS 21.0完成。

|
| 图 4 生物土壤结皮覆盖度与植被覆盖度、土壤性质的关系假设模型 Fig. 4 Hypothesized model of the relationships of BSC coverage, vegetation coverage, and soil properties C1—C10表示共变关系(相关系数), V1—V8为变量的方差, W1—W15为路径系数(Path Coefficients, PC); 模型修正后取消的路径未标注名称 |
由表 2可知, 在毛乌素沙地, BSC覆盖度和植被覆盖度、有机质、平均粒径以及细颗粒占比呈极显著正相关(P < 0.01), 与粗颗粒占比呈极显著负相关(P < 0.01)。植被覆盖度与有机质、平均粒径、细颗粒占比呈极显著正相关(P < 0.01), 与pH值呈显著正相关(P < 0.05), 与粗颗粒占比呈显著负相关(P < 0.05)。pH值与盐度、平均粒径、细颗粒占比呈极显著正相关(P < 0.01), 与有机质呈显著正相关(P < 0.05), 与粗颗粒占比呈极显著负相关(P < 0.01)。盐度与有机质、平均粒径和细颗粒占比呈极显著正相关(P < 0.01)。有机质与平均粒径和细颗粒占比均呈极显著正相关(P < 0.01)。平均粒径与粗颗粒占比呈极显著负相关(P < 0.01), 与细颗粒占比呈极显著正相关(P < 0.01)。粗颗粒占比与细颗粒占比呈极显著负相关(P < 0.01)。
| 环境因子 Environmental factors |
BSC 覆盖度 |
植被 覆盖度 |
pH | 盐度 | 有机质 | 平均粒径 | 粗颗粒 占比 |
细颗粒 占比 |
| BSC覆盖度BSC coverage | 1 | |||||||
| 植被覆盖度Vegetation coverage | 0.313*** | 1 | ||||||
| pH | 0.084 | 0.132* | 1 | |||||
| 盐度Salinity | -0.111 | 0.121 | 0.497*** | 1 | ||||
| 有机质Organic matter | 0.194** | 0.461*** | 0.157* | 0.304*** | 1 | |||
| 平均粒径Average particle size | 0.475*** | 0.367*** | 0.348*** | 0.406*** | 0.512*** | 1 | ||
| 粗颗粒占比Proportion of coarse particles | -0.204** | -0.147* | -0.179** | -0.096 | -0.083 | -0.402*** | 1 | |
| 细颗粒占比Proportion of fine particles | 0.396*** | 0.336*** | 0.322*** | 0.511*** | 0.575*** | 0.930*** | -0.229*** | 1 |
| 采用每个样点3个重复观察值的平均数进行相关分析(n=250); *表示相关性达显著水平(P<0.05), **表示相关性达极显著水平(P<0.01), ***表示相关性达极显著水平(P<0.001) | ||||||||
经过模型界定搜寻并修正后, 理论模型(Default Model)达到适配水平, 稳定指标(Stability Index)值为0.124, 介于±1之间, 表示模型稳定。由表 3可知, 显著性概率值P=0.832>0.05, 接受虚无假设, 理论模型与样本模型适配。卡方值自由度比值为0.367 < 2, RMR值< 0.05, RMSEA值< 0.08, GFI>0.9, AGFI>0.9, CN值在显著水平P=0.05及P=0.01上均≥200, 说明适配度指标均达到模型适配标准。理论模型的AIC、BCC、BIC和CAIC4个指标值均分别大于饱和模型和独立模型的4个指标值, 因此, 模型的精简程度达到标准。
| 模型适配参数 Model adaptation parameters |
理论模型 Default model |
模型适配参数 Model adaptation parameters |
理论模型 Default model |
|
| 自由参数数目Number of free parameters | 32 | 渐进残差均方和平方根 Root mean square error of approximation |
0.000 | |
| 卡方值χ2 | 1.470 | 适配度指数Goodness-of-fit index | 0.999 | |
| df | 4 | 调整后适配度指数 Adjusted goodness-of-fit index |
0.987 | |
| P | 0.832 | 临界样本数Critical N, α=0.05 | 1608 | |
| 卡方/自由度χ2/Degree of freedom | 0.367 | 临界样本数Critical N, α=0.01 | 2250 | |
| 残差均方和平方根Root mean square residual | 0.002 | 简约适配度指数 Parsimony goodness of fit index |
0.111 |
SEM路径分析结果显示(图 5), BSC被pH值、盐度、平均粒径、细颗粒占比、粗颗粒占比、有机质以及植被覆盖度解释的变异量为24%(R2=0.24);植被覆盖度被BSC覆盖度、平均粒径和盐度解释的变异量为13%(R2=0.13);土壤有机质被BSC覆盖度、植被覆盖度、细颗粒占比以及粗颗粒占比解释的变异量为42%(R2=0.42)。外因变量(pH值、盐度、平均粒径、细颗粒占比和粗颗粒占比)间均有相关关系(表 2, 图 5)。BSC覆盖度被所有外因变量所影响(图 5)。其中, 平均粒径对BSC覆盖度直接正向影响最大, 路径系数(Path Coefficient, PC)=0.67(P>0.05), 盐度对BSC覆盖度负向影响显著(PC=-0.41;P < 0.05)。粗颗粒占比和pH值对BSC覆盖度有极小的直接正向影响。在影响植被覆盖度的外因变量中, BSC覆盖度对植被盖度有较大的直接正向影响(PC=0.43;P>0.05), 平均粒径和盐度对植被覆盖度的直接正向影响相等且均不显著(PC=0.12;P>0.05);而植被覆盖度对BSC覆盖度的直接影响为负值(PC=-0.22, P>0.05)。最后, 土壤有机质均被细颗粒占比(PC=0.55;P < 0.05)和植被覆盖度(PC=0.32;P < 0.05)正向显著影响, 而被BSC覆盖度负向不显著影响(PC=-0.16;P>0.05)。

|
| 图 5 BSC覆盖度与植被覆盖度、土壤性质间的SEM结果 Fig. 5 SEM results among BSC coverage, vegetation coverage, and soil properties SEM:结构方程模型Structural equation modeling;RMSEA:渐进残差均方和平方根Root mean square error of approximation;AGFI:调整后良适性适配指标Adjusted goodness-of-fit index;GFI:良适性适配指标Goodness-of-fit index; 双向箭头代表共变关系;单箭头起始点为自变量(外因变量), 方向所指的变量为依变量(内因变量), 表示外因变量直、间接影响内因变量;路径系数(Path Coefficient, PC)≥0.20为深灰色单向箭头, PC < 0.20为浅灰色单向箭头;虚线单、双箭头表示PC/相关关系为负, 实线单、双箭头表示PC/相关关系为正 |
图 5中细颗粒占比对BSC覆盖度的直接影响较小且为负向(PC=-0.02, P>0.05), 但表 4显示, 细颗粒占比对BSC覆盖度有间接影响(间接效果值=0.052), 即细颗粒占比正向影响有机质, 有机质又正向影响BSC覆盖度, 因此, 细颗粒占比对BSC覆盖度的总体影响为正(总效果值=0.030)。同理, 平均粒径对植被覆盖度的间接正向影响较大(间接效果值=0.251), 总效果值达到0.366。盐度对BSC覆盖度的直接负向影响较大(图 5), 但通过对植被覆盖度的正向影响从而正向影响有机质, 最后极小地正向间接影响了BSC覆盖度(间接效果值=0.016)。BSC覆盖度通过正向直接影响植被覆盖度而间接正向影响了有机质(间接效果值=0.137), 然而综合其对有机质的直接负向影响, 总影响为负向(总效果值=-0.026)。
| 环境因子 Environmental factors |
有机质 Organic matter |
植被覆盖度 Vegetation coverage |
BSC覆盖度 BSC coverage |
有机质 Organic matter |
植被覆盖度 Vegetation coverage |
BSC覆盖度 BSC coverage |
|
| 间接效果值 Indirect effects |
总效果值 Total effects |
||||||
| 粗颗粒占比Proportion of coarse particles | 0.000 | 0.006 | 0.004 | 0.055 | 0.006 | 0.015 | |
| 细颗粒占比Proportion of fine particles | -0.001 | 0.013 | 0.052 | 0.547 | 0.013 | 0.030 | |
| 平均粒径Average particle size | 0.019 | 0.251 | -0.080 | 0.019 | 0.366 | 0.590 | |
| 盐度Salinity | 0.050 | -0.169 | 0.016 | 0.050 | -0.048 | -0.398 | |
| pH | -0.002 | 0.031 | -0.007 | -0.002 | 0.031 | 0.072 | |
| 有机质Organic matter | -0.003 | 0.039 | -0.009 | -0.003 | 0.039 | 0.092 | |
| 植被覆盖度Vegetation coverage | 0.005 | -0.075 | 0.049 | 0.321 | -0.075 | -0.176 | |
| BSC覆盖度BSC coverage | 0.137 | -0.038 | -0.090 | -0.026 | 0.387 | -0.090 | |
生物土壤结皮在荒漠地区广泛分布, 是固沙和促进沙地生态恢复过程中的重要角色[9]。本研究认为, BSC覆盖度与植被覆盖度相互影响, 这进一步证实了Harper和Pendleton[38], 以及Bochet等[39]的研究结果。其中, BSC覆盖度正向影响植被覆盖度, 原因可能是由于BSC的固碳、固氮能力可以为植物提供所需的养分, 从而有利于植被的生长[38]。王刚和梁学功[40]研究发现, 随着沙层表面BSC厚度逐渐增加, 一年生植物将大量侵入并占据优势地位。在野外工作时也观察到同样的现象, 在BSC生长较厚的地块, 多生长灰绿藜、小画眉草、雾冰藜、狗尾草等一年生草本植物, 这些一年生草本植物显著提高了植被盖度。究其原因主要是由于BSC越厚, 对降水的阻碍作用越大, 使得深根系的灌木得不到降水补给而死亡, 且BSC有较好的持水性, 当被少量降水湿润后, 易为一年生草本植物种子萌发创造条件[2]。然而, Li等[18]和Su等[6]研究结果发现, 苔藓主导BSC的多样性与盖度和一年生植物盖度呈负相关关系, 藻类主导的BSC则与一年生植物呈正相关关系, 他们认为BSC未受干扰时覆盖了地表, 使得一年生植物的种子不易进入土壤。本研究中未发现此规律, 可能与BSC的厚度范围有关, 李新荣等[41]研究结果显示, BSC厚度为0.3-2.0 cm时与物种多样性呈正相关关系, 该厚度范围与毛乌素沙地BSC厚度范围契合。在毛乌素沙地, 生态工程建设和生态系统管理过程中, 决策者易将注意力集中于高大的固沙灌木与草本植物, 而忽略了BSC在生态系统恢复中的功能[2, 9], 未来, 决策者应注重BSC对植被的正向促进作用。与此同时, 虽然植被覆盖度对BSC覆盖度呈直接负向影响, 但植被的生长显著正向影响了土壤有机质, 且维管植物过滤和吸收了部分到达地表的太阳辐射[39], 为BSC的生长发育造就出土壤含水量较高、温度较低且养分富集的有利微生态环境, 这些影响均是正向的、间接的影响。Eldridge等[42]的研究认为, BSC的分布与维管植物间存在负相关关系, 随着植被盖度的增加, 植被与BSC对水分、养分和光照等环境资源的竞争加剧, 从而共同限制了它们的生长。在进行野外实地采样时, 也发现在少数植被覆盖度极高的地方, BSC反而有所退化, 而在极少数BSC非常丰富的地方(覆盖度>80%), 伴随其生长的油蒿反而枯死, 这可能与BSC影响土壤水分的再分配有关[43]。然而, 本文结果并未反映出这一现象, 可能是由于整体数据样本中该情况极其罕见, 属于极小概率事件, 无法在大样本数据中体现出来。
本研究中, 土壤平均粒径正向影响BSC覆盖度与植被覆盖度, 由公式(1)、(2)可知, 平均粒径Mz值越大, 土壤颗粒越细。Lan等[44]认为, 细颗粒多的土壤更易吸收水分, 从而宜于BSC和植被的生长。West[45]发现, BSC破损会造成土壤中细颗粒和有机质的损失而影响植被生长。何芳兰等[20]研究发现, BSC发育初期到形成藻类结皮阶段的过程中, 0—1 cm层土壤细沙粒含量有效提高, 而粗砂粒含量则降低。本研究中, 细颗粒占比对BSC覆盖度的正向影响主要通过有机质对BSC覆盖度造成正向的间接影响。另外, 粗颗粒占比虽然与BSC呈极显著负相关关系, 但其对BSC的直接、间接影响均为正向, 这进一步验证了Rong等[46]的研究结果, 说明风沙土中黏粒和粉粒含量的提高均有利于BSC的拓殖和发展, 但粗颗粒占比超过一定范围后BSC将无法存活。
本研究结果发现pH值、盐度与BSC的相关性不显著。Li等[47]在腾格里沙漠东南缘的研究表明, 流动沙丘固定后, 随着土壤pH值的增大, 蓝藻的物种多样性呈上升趋势。但SEM结果显示, pH值对BSC覆盖度总影响效果值为0.072, 而盐度对BSC覆盖度有显著的直接负向作用效应, 即盐度不利于BSC的生长发育。在野外采样时同样发现盐碱地区几乎没有BSC存在, 说明pH值在一定酸碱度范围内增大时, 会正向影响BSC覆盖度, 而当超过一定范围时, BSC则无法生存, 由此推断BSC可作为土壤盐渍化监测的科学依据。
此外, 对BSC覆盖度与植被覆盖度、土壤有机质含量、土壤平均粒径、土壤细颗粒占比、土壤粗颗粒占比及土壤盐度间进行线性拟合发现(图 6), 相关性显著单变量间线性拟合均达显著水平, 但拟合存在区间差异性, R2均较小, 散点图趋势不规律, 进一步表示BSC覆盖度受综合因素影响, 无法用单一变量进行说明。李新荣等[9]也认为, 高等植物、土壤与BSC形成和发展之间的相互作用很复杂, 且BSC所处不同演替阶段时, 其影响因子由于研究尺度不同而各异。认为, 利用结构方程模型分析之后, 下一步研究可考虑利用机器学习中的关联规则算法等其他强调综合分析的方法对BSC覆盖度与土壤性质的多级强关联规则进行挖掘。

|
| 图 6 生物土壤结皮覆盖度与其显著相关单变量间的直线方程拟合 Fig. 6 Linear equation fitting between biological soil crust coverage and its significant correlation factors 散点图中圆点为灰色半透明, 点的颜色越深表示数量越多的点重合在一起 |
综上所述, 在毛乌素沙地, BSC覆盖度对植被覆盖度有较大的正向影响;植被覆盖度对BSC覆盖度有直接负向影响和间接正向影响。土壤平均粒径和细颗粒占比均可正向影响BSC的覆盖度。盐度对BSC覆盖度有显著的负向直接影响, 但pH值对BSC覆盖度有极小的正向影响。此外, BSC覆盖度受各环境因子综合影响, 无法用单一变量说明。本研究的不足之处在于, 未将表层土的土壤含水量、温度及降水量考虑进模型内, 这也将是未来研究的主要内容。本研究可在脆弱生态系统保护与修复过程中为决策者提供重要的理论基础及机理解释。
| [1] |
Belnap J, Lange O L. Biological Soil Crusts: Structure, Function, and Management. Berlin, Germany: Springer, 2003, 3-32. |
| [2] |
李新荣, 回嵘, 赵洋. 中国荒漠生物土壤结皮生态生理学研究. 北京: 高等教育出版社, 2016: 5-10.
|
| [3] |
Li X R, Zhang P, Su Y G, Jia R L. Carbon fixation by biological soil crusts following revegetation of sand dunes in arid desert regions of China: a four-year field study. CATENA, 2012, 97: 119-126. DOI:10.1016/j.catena.2012.05.009 |
| [4] |
赵河聚, 成龙, 贾晓红, 吴波, 李元寿, 岳艳鹏, 周虹, 赵雪彬. 高寒沙区生物土壤结皮覆盖土壤碳释放动态. 生态学报, 2020, 40(18): 6396-6404. |
| [5] |
李云飞, 都军, 张雪, 谢婷, 李小军. 腾格里沙漠东南缘不同类型生物土壤结皮对土壤有机碳矿化的影响. 生态学报, 2020, 40(5): 1580. |
| [6] |
Su Y G, Zhao X, Li A X, Li X R, Huang G. Nitrogen fixation in biological soil crusts from the Tengger desert, northern China. European Journal of Soil Biology, 2011, 47(3): 182-187. DOI:10.1016/j.ejsobi.2011.04.001 |
| [7] |
Belnap J. Nitrogen fixation in biological soil crusts from southeast Utah, USA. Biology and Fertility of Soils, 2002, 35(2): 128-135. DOI:10.1007/s00374-002-0452-x |
| [8] |
李守中, 肖洪浪, 宋耀选, 李金贵, 刘立超. 腾格里沙漠人工固沙植被区生物土壤结皮对降水的拦截作用. 中国沙漠, 2002, 22(6): 612-616. DOI:10.3321/j.issn:1000-694X.2002.06.016 |
| [9] |
李新荣. 荒漠生物土壤结皮生态与水文学研究. 北京: 高等教育出版社, 2012: 257-373.
|
| [10] |
李新荣, 张元明, 赵允格. 生物土壤结皮研究: 进展、前沿与展望. 地球科学进展, 2009, 24(1): 11-24. DOI:10.3321/j.issn:1001-8166.2009.01.002 |
| [11] |
Rodríguez-Caballero E, Escribano P, Cantón Y. Advanced image processing methods as a tool to map and quantify different types of biological soil crust. ISPRS Journal of Photogrammetry and Remote Sensing, 2014, 90: 59-67. DOI:10.1016/j.isprsjprs.2014.02.002 |
| [12] |
杨晓晖, 张克斌, 赵云杰. 生物土壤结皮——荒漠化地区研究的热点问题. 生态学报, 2001, 21(3): 474-480. |
| [13] |
Li X R, Xiao H L, Zhang J G, Wang X P. Long-term ecosystem effects of sand-binding vegetation in the Tengger desert, Northern China. Restoration Ecology, 2004, 12(3): 376-390. DOI:10.1111/j.1061-2971.2004.00313.x |
| [14] |
Bowker M A, Belnap J, Davidson D W, Goldstein H. Correlates of biological soil crust abundance across a continuum of spatial scales: support for a hierarchical conceptual model. Journal of Applied Ecology, 2006, 43(1): 152-163. DOI:10.1111/j.1365-2664.2006.01122.x |
| [15] |
Li X R. Influence of variation of soil spatial heterogeneity on vegetation restoration. Science in China Series D: Earth Sciences, 2005, 48(11): 2020-2031. DOI:10.1360/04yd0139 |
| [16] |
Kidron G J, Benenson I. Biocrusts serve as biomarkers for the upper 30 cm soil water content. Journal of Hydrology, 2014, 509: 398-405. DOI:10.1016/j.jhydrol.2013.11.041 |
| [17] |
Danin A, Ganor E. Trapping of Airborne Dust By Mosses in the Negev Desert, Israel. Earth Surface Processes and Landforms, 1991, 16(2): 153-162. DOI:10.1002/esp.3290160206 |
| [18] |
Li X R, Jia X H, Long L Q, Zerbe S. Effects of biological soil crusts on seed bank, germination and establishment of two annual plant species in the Tengger Desert (N China). Plant and Soil, 2005, 277(1/2): 375-385. |
| [19] |
Flechtner V R. North American Desert microbiotic soil crust communities//Seckbach J, ed. Cellular Origin, Life in Extreme Habitats and Astrobiology: Netherlands: Springer, 2007: 539-551.
|
| [20] |
何芳兰, 郭春秀, 吴昊, 刘左军, 徐文, 金红喜. 民勤绿洲边缘沙丘生物土壤结皮发育对浅层土壤质地、养分含量及微生物数量的影响. 生态学报, 2017, 37(18): 6064-6073. |
| [21] |
Read C F, Duncan D H, Vesk P A, Elith J. Biological soil crust distribution is related to patterns of fragmentation and landuse in a dryland agricultural landscape of southern Australia. Landscape Ecology, 2008, 23(9): 1093-1105. DOI:10.1007/s10980-008-9270-3 |
| [22] |
Li X R, He M Z, Zerbe S, Li X J, Liu L C. Micro‐geomorphology determines community structure of biological soil crusts at small scales. Earth Surface Processes and Landforms, 2010, 35(8): 932-940. DOI:10.1002/esp.1963 |
| [23] |
Zhang J H, Wu B, Li Y H, Yang W B, Lei Y K, Han H Y, He J. Biological soil crust distribution in Artemisia ordosica communities along a grazing pressure gradient in Mu Us Sandy Land, Northern China. Journal of Arid Land, 2013, 5(2): 172-179. DOI:10.1007/s40333-013-0148-0 |
| [24] |
张新时. 毛乌素沙地的生态背景及其草地建设的原则与优化模式. 植物生态学报, 1994, 18(1): 1-16. DOI:10.3321/j.issn:1005-264X.1994.01.008 |
| [25] |
何彤慧. 毛乌素沙地历史时期环境变化研究[D]. 兰州: 兰州大学, 2008.
|
| [26] |
郭紫晨, 刘树林, 康文平, 陈翔, 张雪琴. 2000—2015年毛乌素沙区植被覆盖度变化趋势. 中国沙漠, 2018, 38(5): 1099-1107. |
| [27] |
Cheng X L, An S Q, Liu S R, Li G Q. Micro-scale spatial heterogeneity and the loss of carbon, nitrogen and phosphorus in degraded grassland in Ordos Plateau, Northwestern China. Plant and Soil, 2004, 259(1/2): 29-37. DOI:10.1023/B:PLSO.0000020948.66471.2b |
| [28] |
Wu B, Ci L J. Landscape change and desertification development in the Mu Us Sandland, Northern China. Journal of Arid Environments, 2002, 50(3): 429-444. DOI:10.1006/jare.2001.0847 |
| [29] |
Chen X, Wang T, Liu S L, Peng F, Tsunekawa A, Kang W P, Guo Z C, Feng K. A new application of random forest algorithm to estimate coverage of moss-dominated biological soil crusts in semi-arid Mu Us Sandy Land, China. Remote Sensing, 2019, 11(11): 1286. DOI:10.3390/rs11111286 |
| [30] |
吉增宝. 用数码照片和Photoshop计算植被覆盖度的简易方法. 水土保持应用技术, 2015(5): 10. DOI:10.3969/j.issn.1673-5366.2015.05.04 |
| [31] |
鲍士旦. 土壤农化分析 (第三版). 北京: 中国农业出版社, 2000: 14-38, 178-200.
|
| [32] |
董治宝, 苏志珠, 钱广强, 罗万银, 张正偲, 吴晋峰. 库姆塔格沙漠风沙地貌. 北京: 科学出版社, 2011: 67-80.
|
| [33] |
Feng X, Qu J J, Tan L H, Fan Q B, Niu Q H. Fractal features of sandy soil particle-size distributions during the rangeland desertification process on the eastern Qinghai-Tibet Plateau. Journal of Soils and Sediments, 2020, 20(1): 472-485. DOI:10.1007/s11368-019-02392-6 |
| [34] |
吴明隆. 结构方程模型——AMOS的操作与应用 (第二版). 重庆: 重庆大学出版社, 2010: 1-68, 263-299.
|
| [35] |
Moore K A, Hofferth S L. Factors affecting early family formation: A path model. Population and Environment, 1980, 3(1): 73-98. DOI:10.1007/BF01253071 |
| [36] |
Mathieu J E. A cross-level nonrecursive model of the antecedents of organizational commitment and satisfaction. Journal of Applied Psychology, 1991, 76(5): 607-618. DOI:10.1037/0021-9010.76.5.607 |
| [37] |
Kline R B. Principles and Practice of Structural Equation Modeling. New York: Guilford Press, 1998.
|
| [38] |
Harper K T, Pendleton R L. Cyanobacteria and cyanolichens: can they enhance availability of essential minerals for higher plants?. Great Basin Naturalist, 1993, 53(1): 59-72. |
| [39] |
Bochet E, Rubio J L, Poesen J. Modified topsoil islands within patchy Mediterranean vegetation in Se Spain. CATENA, 1999, 38(1): 23-44. |
| [40] |
王刚, 梁学功. 沙坡头人工固沙区的种子库动态. 植物学报, 1995, 37(3): 231-237. |
| [41] |
李新荣, 张景光, 刘立超, 陈怀顺, 石庆辉. 我国干旱沙漠地区人工植被与环境演变过程中植物多样性的研究. 植物生态学报, 2000, 24(3): 257-261. |
| [42] |
Eldridge D J, Tozer M E. Environmental factors relating to the distribution of terricolous bryophytes and lichens in Semi-arid Eastern Australia. The Bryologist, 1997, 100(1): 28-39. |
| [43] |
Li X R, Kong D S, Tan H J, Wang X P. Changes in soil and vegetation following stabilisation of dunes in the southeastern fringe of the Tengger Desert, China. Plant and Soil, 2007, 300(1/2): 221-231. |
| [44] |
Lan S B, Wu L, Zhang D L, Hu C X. Analysis of environmental factors determining development and succession in biological soil crusts. Science of the Total Environment, 2015, 538: 492-499. |
| [45] |
West N E. Structure and function of microphytic soil crusts in wildland ecosystems of arid to semi-arid regions. Advances in Ecological Research, 1990, 20: 179-223. |
| [46] |
Rong L J, Li X R, Liu L C, Gao Y H, Li X J. Responses of biological soil crusts to sand burial in a revegetated area of the Tengger Desert, Northern China. Soil Biology and Biochemistry, 2008, 40(11): 2827-2834. |
| [47] |
Li X R, Zhou H Y, Wang X P, Zhu Y G, O'Conner P J. The effects of sand stabilization and revegetation on cryptogam species diversity and soil fertility in the Tengger Desert, Northern China. Plant and Soil, 2003, 251(2): 237-245. |