 2022, Vol. 42
2022, Vol. 42文章信息
- 李善家, 王子濠, 苏培玺, 王辉, 王福祥, 崔莉娟
- LI Shanjia, WANG Zihao, SU Peixi, WANG Hui, WANG Fuxiang, CUI Lijuan
- 荒漠植物性状权衡策略及功能多样性研究进展
- Research progress on the trade-off strategy and functional diversity of desert plants
- 生态学报. 2022, 42(18): 7308-7320
- Acta Ecologica Sinica. 2022, 42(18): 7308-7320
- http://dx.doi.org/10.5846/stxb202107151909
-
文章历史
- 收稿日期: 2021-07-15

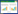




2. 中国科学院西北生态环境资源研究院, 兰州 730000
2. Key Laboratory of Land Surface Process and Climate Change in Cold and Arid Regions, Northwest Institute of Eco-Environment and Resources, Chinese Academy of Sciences, Lanzhou 730000, China
干旱区生态系统蒸发强烈、多风沙暴、高光高温、土壤贫瘠盐渍化, 环境异常严酷, 这一特殊生境孕育了广袤而丰富的旱生、超旱生、盐生和沙生的荒漠植被资源。荒漠植被是由荒漠地区干旱的气候及其特定的自然条件综合作用形成的地带性植被, 指示着干旱地区生态与环境的变化规律, 对环境变迁、气候变化及人为影响的敏感性较强。经过长期的环境适应和自身演化, 荒漠植被在形态结构、生理生态等方面形成了独特的适应特征并表现出相应的功能对策[1]。因此, 研究荒漠植物的适应策略对于植物特殊功能的发现、脆弱生态环境保护、荒漠环境演化历史、抗逆基因提取及资源可持续利用等方面都有着至关重要的意义。
植物功能性状(plant functional traits)是植物体生命周期中获取、利用、保留资源的一系列关键属性, 它可以单独或协同指示植物生长策略对环境变化做出响应, 还可以反映生态系统功能和过程的变化[2—3]。植物的功能性状是动态的, 会随着物种进化而发生适应性分化, 在器官、个体、种群、群落和生态系统尺度上, 已成为解决生态学热点问题的可靠途径。植物功能性状的分类方法众多, 根据植物对环境的相应对策可分为营养性状和繁殖性状, 根据与生态系统的关联可分为响应性状和效应性状[4]。响应性状和效应性状是近年来新提出的植物功能性状分类方法, 它们分别定义了植物响应环境梯度变化和植物影响生态系统功能的物种性状。目前被广泛接受的分类方法还包括软性状和硬性状, 软性状通常指那些可以轻易得到并能够快速测定的植物性状, 如株高、比叶面积等;而硬性状则指那些难以定量的特征, 如光合作用效率、植物耐寒耐阴性等[2]。与软性状相比, 硬性状虽然获得方法困难但却能更准确的反映生态系统功能特征[4]。
本文主要从荒漠植物组织、器官功能性状特征、性状权衡策略、功能多样性组分及选择3个方面梳理了荒漠植物研究的进展脉络:1) 荒漠植物独特的组织、器官功能性状特征体现了植被对环境的适应策略;2) 荒漠植物的性状权衡策略是在特定生境下形成的性状组合, 关键性状已经被发掘并创造性的提出了“经济谱”概念;3) 荒漠植物功能多样性能对荒漠生态系统功能产生影响, 功能多样性的组分可以从不同角度反映群落的生态位占据状况和资源利用程度, 指数的选择要体现在群落内部物种的功能特征之间的差异程度, 同时要考虑这些物种自身在群落内的优势程度。这三个方面是近年来有关荒漠植物研究的重点内容, 开展相关综述性工作具有一定的借鉴和启发意义。
1 荒漠植物独特的组织和器官功能性状特征地球陆地表面约1/3是干旱荒漠生态系统, 孕育出了形态独特、物种多样的荒漠植物。本综述着眼于荒漠植物根、茎、叶等3个器官的功能性状特征, 介绍了荒漠植物功能性状研究在近20余年来的发展概况(表 1)。对荒漠植物的功能性状进行研究, 可以揭露植被对环境变化的响应以及对生态系统功能的影响[25]。研究表明, 基于植物功能性状的研究有助于解决许多生态学的关键性问题[26], 如Cadotte等通过对植物性状多样性的测定和理解, 优化了生态系统保护和恢复策略[27];何念鹏等将传统功能性状特征与宏观生态学联系起来, 创造性的提出了“生态系统特征”的概念等[3]。因此, 对荒漠植物组织、器官功能性状特征的研究就显得尤为重要。
| 组织 Tissue |
采集地点或数据来源 Collection site or data source |
植被类群 Vegetation types |
关键性状 Key traits |
参考文献 References |
| 根Root | 塔里木盆地北缘 | 盐生植物等 | 化学计量特征 | [5] |
| 黄土丘壑沟壑区等 | 灌木、半灌木 | 细根长、细根生物量 | [6] | |
| 宁夏引黄灌区 | 盐生植物 | 比根长、比根面积 | [7] | |
| 乌兰布和沙漠 | 草本植物、灌木 | 根冠比 | [8] | |
| 奇瓦瓦沙漠北缘 | 草本植物、灌木等 | 比根长 | [9] | |
| 莫哈维沙漠 | 灌木 | 根体积 | [10] | |
| Telteca保护区 | 草本植物 | 地上根比、日伸长率、根长密度 | [11] | |
| 茎Stem | 烟台、石家庄、银川、吐鲁番 | 沙生灌木 | 茎比密度、茎水分含量 | [12] |
| 河西走廊中段 | 沙生灌木 | 茎干液流 | [13] | |
| 加利福尼亚 | 小乔木、灌木 | 比茎面积 | [14] | |
| 加利福尼亚及地中海区域 | 乔木、灌木 | 茎型性状 | [15] | |
| 叶Leaf | 塔克拉玛干沙漠南缘 | 灌木 | 相对含水量、水势、失水速率 | [16] |
| 新疆 | 乔木、灌木、草本植物 | 化学计量特征 | [17] | |
| 艾比湖 | 小乔木、灌木、草本植物 | 叶绿素含量 | [18] | |
| 黑河流域 | 半灌木 | 叶水势、相对含水量 | [19] | |
| 晋西北黄土丘陵区 | 半灌木、小半灌木、草本植物 | 叶干物质含量、比叶面积 | [20] | |
| 奇瓦瓦沙漠 | 灌木、草本植物 | 叶面积指数 | [21] | |
| 埃及地中海沿岸 | 灌木、草本植物等 | 叶脉厚度、叶脉容积 | [22] | |
| 沙特阿拉伯吉达地区 | C3植物 | 气孔导度 | [23] | |
| 索诺拉沙漠 | 小乔木、灌木 | 化学计量特征 | [24] |
植物叶片是维持陆地生态系统功能的基本元素, 同时也是维护植物碳水耦合权衡的重要器官[28], 叶片的功能性状对植物个体、群落、生态系统、地球生物化学循环具有“源动力”性的作用[29]。植物叶片在植物进化过程中对环境的变化比较敏感, 具有更强的可塑性, 其结构与功能特性不仅展现了植物对环境的相应对策及植物利用资源的能力[30], 更展现了植物为获取最大化碳收益所采用的生存适应性策略[31]。荒漠植物叶片(或同化枝)表皮的基本特征是:具有丰富的表皮附属物, 包括大量表皮绒毛、角质膜蜡质片层或晶体颗粒、表面结节或疣状突起、气孔器相对凹陷、密度低[32]。荒漠植物为了适应极端干旱和高量盐分的环境, 有些叶片缩小甚至演化成为薄鳞片状或硬刺状;有些叶片肉质化且形状多为不规则柱状体或锥状体;有些叶片发生强烈的分裂现象、出现白色茸毛、气孔深陷等从而减少水分的散失;有些叶片已经退化, 以当年的嫩枝进行光合作用[33]。然而, 在测量叶面积等常用指标时, 存在实际的困难, 即使是非肉质化旱生植物叶片, 由于耐旱的需要, 不同程度上会发生对折、卷曲或被覆大量表皮毛, 这对于光合表面积作为量化植物功能的参比是非常不利的。研究发现, 对于荒漠植物而言, 需要建立以叶体积为基础(volume-based)的标准化性状指标来进行叶性状功能和数量化方法的研究以阐明植物功能性状间对有限资源进行的权衡作用[33]。Wang等人测定了灌木的叶、茎和根性状之间的关系, 得出叶片性状是整个植物性状的有力替代, 植物适应环境的策略可能在很大程度上反映在叶片的功能性状上[34]。因此, 在研究荒漠植物适应性的过程中, 要综合考虑不同叶片性状因子之间的相互关系, 同时要探寻一些关键叶片性状进行描述来反映荒漠植物的环境适应特征。深入探讨荒漠植物叶片形态、功能和与环境的联系, 对研究荒漠植物特殊的生态适应性具有重要理论意义, 同时可以为干旱生态系统的维持及服务功能的稳定发挥提供理论支撑。
茎是植物根系和叶片之间的养分运输桥梁, 同时对植物体起到支持和承载作用[35]。荒漠植物器官茎为了适应极端环境而形成了独特的结构特征[36], 其功能性状往往反映了植物水分利用能力、生长能力、抵抗逆境能力间的权衡[37]。茎往往被周皮包裹(幼茎为不同程度的角质层)以抑制植物体内的蒸腾作用;表皮皮层中普遍含有绿色同化组织, 可显著提高荒漠植物的光合效力;茎的输导组织十分发达以储存和运输水分;茎中有大量薄壁细胞, 有些含有粘液细胞或晶体等特殊物质, 通过提高细胞的渗透压和保水性, 以适应干旱胁迫;茎普遍存在异常结构, 如盐囊泡(一种盐腺的类型)、表皮毛等, 这对于荒漠植物长久在极端环境下的生长发育尤为重要;部分荒漠植物的维管组织具有韧皮纤维, 呈环状排列, 韧皮部与木质部之间存在明显的形成层带, 增强了轴器官的支持力, 可有效缓解恶劣环境对轴器官造成的损伤[36]。生态学家们发现, 抵抗逆境能力强的荒漠植物往往具有高的茎干密度和机械强度[35]。
根是植物在地下进行物质分配和交换的重要营养器官[38], Holdaway等通过对根长和根平均直径等相关性状的研究发现植物地下根系与土壤之间存在物质交换[39], 其功能性状能够显现出根系对营养成分的吸收和运输效率[40]。荒漠植物为了在极端环境下得以生存, 其根的形态结构往往不同于其他植物, 有些荒漠植物根系为了适应极度缺水、土壤盐渍化的生理环境, 根系极发达且具有较强的抗旱能力, 地下深度可达数米[5];有些荒漠植物根系呈现空间网状结构, 分支结构复杂且次级分支多[41];有些荒漠植物根系连接长度较长以汲取更多营养来供给地上部分的正常生长发育[42]。由于荒漠植物的根系结构十分复杂又位于地下深处, 因而不同于其他功能性状能从其外观得到一个基本的认识和了解, 所以学者们通常以根组织密度、比根长、根冠比、根系拓扑结构等指标来体现荒漠植物的根系生长情况。近期的研究结果表明, 根的功能性状之间存在着“根经济谱(RES)”, 如Reich通过分析根氮浓度与根寿命、根呼吸及根形态性状的相关性发现存在根生长与细根持久性之间权衡的RES[43]。Roumet等在研究包括草本植物、矮小灌木和高大灌木等74种植物根结构与功能的关系时, 揭示了根系形态与呼吸和碳收支两个过程之间的联系, 明确了多种共变性状, 并证明根呼吸与剩余根量负相关, 74种植物的数据支持基于碳模式的RES的存在[44]。RES很好的反映了在物种水平上根对营养获取和建成代价之间的权衡[37]。
2 荒漠植物性状权衡策略植物功能性状之间通常存在一种权衡性关系, 这种权衡策略是经过自然的不断筛选而逐渐形成的, 也可以称之为“生态策略”, 它最终可以在不同的生态系统和生物群落间表现为沿基础资源轴排列, 资源轴即代表各种环境因子, 资源轴的一端聚集了资源快速获取的生态策略, 为“快速投资-收益”型物种”型物种;另一端则聚集资源高度保存的生态策略, 为“缓慢投资-收益”型物种, 沿这条主轴分布的连续变化的植物功能性状组合则被称为“经济谱”[45—46]。植物有效地获取所需资源的生态策略, 以及将有限的可利用资源优化配置的方式被称为植物资源权衡策略[47—48]。植物功能性状的重要指示意义在于量化表达了“资源权衡策略”这个抽象的生态学概念[43], 研究植物功能性状间的权衡策略不仅能够了解植物生态策略在不同环境内和环境间的差异, 更可以深入探索生态位分化和物种共存的内在机制, 而且有助于理解生态系统中能量流动与物质循环的发展和变化[6, 49]。以植物功能性状为研究对象的功能多样性和权衡策略的理论正在扮演承上启下——联系宏观和微观的生态学功能与过程、实现尺度互推的重要角色, 最终通过模型模拟建立全面、客观的植物与环境的关系网络(图 1)。

从近些年的研究来看, 对植物功能性状的研究已经从单一或成组性状衍生到植物功能性状之间关系的研究[51], 这种关系一般认为包含其内在联系和权衡策略[52], 而权衡策略则是植物功能性状之间存在的最普遍的联系[26]。对于荒漠植物而言, 其生境多呈营养匮乏、干旱少雨及盐渍化特征[53], 为了生存, 荒漠植物长期的演绎进化形成了独特的适应特征并表现出相应的功能对策[54]。不同类型的植物会根据其功能需求的不同, 在自身的功能性状之间对资源进行权衡分配, 继而达到对资源的有效利用[4]。最近几年, 通过对荒漠植物功能性状之间关系的研究发现, 对荒漠植物“生态策略”的研究有助于:揭示荒漠植物的生长策略及资源分配模式, 探索在荒漠生态系统中植物生理生态过程的内在机制[51];了解荒漠植物在不同环境间和同一环境内的生态策略差异, 进而探求生态位分化和物种共存的内在机理[55];预测全球气候变化对荒漠植物的影响, 探讨荒漠生态系统功能的原理[51]。
在功能多样性及权衡策略研究中, 筛选出与特定生态系统过程相关联的植物功能性状是关键, 只有与资源的获取、利用和保护直接相关的功能性状才被认为是关键性状[43]。Wright等通过收集全世界175个地区2548种植物叶功能性状数据形成了“叶经济谱(LES)”理论, 提炼出了6个重要的性状, 即叶寿命、比叶重、净光合速率、暗呼吸速率、叶氮含量与叶磷含量[56]。Cui等在模拟气候变暖、干旱、CO2升高和氮沉降的210个实验中研究了515个物种的叶片经济性状之间的双变量相关性, 准确描述了植物叶片功能性状间的权衡策略, 并由此揭示了其对全球环境变化的响应在不同的研究尺度上存在的不同格局与机制, 研究结果对全球环境变化的响应规律、陆地生态系统生产力的预测都具有重要指示意义[57]。He等以青藏高原草地上维管植物叶片为研究对象, 发现在该生境下植物叶片有更高的氮浓度和更强的光合作用能力, 叶片性状间关系与LES一致[58]。Yin等人对黄土高原半干旱地区的47种木本植物的研究表明:气孔保卫细胞长度与叶片厚度和比叶重显著正相关, 荒漠植物的经济性状与水力性状会协同调整以适应干旱条件[59]。李镯等以吉兰泰盐湖地区生长的荒漠植物为研究对象发现, 荒漠植物通过较高的干物质质量分数(LDMC)、较低的比叶面积(SLA)和叶片含水量(LWC)来适应荒漠环境胁迫, 这三个指标相结合可作为荒漠植物筛选的主要性状指标[60]。李军军等以芨芨草为实验对象, 通过盆栽实验得出土壤养分添加水平和氮磷比共同影响着芨芨草的叶片养分生态化学计量学特征和养分回收[61]。刘玉冰等对中国干旱半干旱荒漠区117种的植物叶片(或同化枝)表皮微形态进行了研究, 得出荒漠植物通过表皮附属物与表皮结构的相互协调作用来提高荒漠植物对干旱胁迫的抗性[32]。Rodríguez-Gallego等发现78种沿海沙丘植物叶片的SLA与氮、磷含量呈正相关关系[62]。叶功能性状反映叶片寿命、建成代价与光合作用能力间的权衡策略[52, 56]。
Chave等通过对世界尺度植物的茎干性状进行分析, 得出了具有较高茎干密度和机械强度的物种属于“缓慢投资-收益”型物种, 这些植物在干旱胁迫下通常具有较强的抵抗力, 并提出“茎干经济谱”[35]概念。Reich等基于全球范围内287种草本及木本植物的研究发现, 单位质量下茎和根的氮浓度与暗呼吸速率具有强相关性, 但是在相同的氮浓度条件下, 叶片的平均呼吸速率明显低于茎和根, 主要是因为茎和根需要将更多的氮投资到呼吸代谢中, 用于转化非结构性碳水化合物[63]。Ishida等发现小笠原群岛上抗旱物种的木质密度与叶片SLA显著负相关[64]。Riva等对地中海地区38种木本植物的研究发现, 根、茎、叶的形态性状具有显著相关性, 且这种相关性在群落水平上更加紧密[65]。茎干的功能性状反映了植物在水分利用能力、生长能力、抵抗逆境能力间的权衡策略[66]。
近几年来, “根经济谱”概念也被多次提到, Prieto和Roumet等人研究发现比根长能够很好的与其他根系结构性状相联系, 与根干物质含量、根系平均直径、分枝强度正相关, 与根系组织密度呈负相关[44, 67]。马芳等通过测定内蒙古草原中22种优势植物根的形态和构型属性, 得出根直径与侧根长度显著正相关, 而与分支强度负相关的结论, 体现了草本植物根构建成本较低的特点[68]。马泽清等收集了全球369种植物一级根数据, 发现在水分胁迫较强的荒漠中, 根系多样性由于资源供应的季节性波动而出现下降[69]。郑慧玲等利用Arc GIS建立研究区域的数字高程模型, 研究了祁连山荒漠草原不同坡向红砂根系分叉数和分支角度的关系, 结果表明不同坡向红砂根系与连接长度和分支角度之间存在合理的权衡策略, 这种资源配置模式反映了异质生境中植物种群对环境生态的适应机制[70]。Kong等利用全球800多种植物根系性状数据, 首次在全球尺度上对根系经济学谱假说进行了验证, 这对于理解植物根系的结构、功能与进化等具有重要意义[71]。根性状往往代表了物种繁殖能力、养分利用能力、建成代价间的权衡[72]。
Reich试图通过建立植物根-茎-叶系统的碳-养分-水资源的整株资源权衡策略, 但其试用的尺度及植物个体与群落、生态系统功能结构上的耦合机制仍不确定[43]。Thomas等通过建立目前为止最大的冻土带植物性状数据库, 发现极端环境下的苔原植物同样表现出非常相似的资源经济性状, 并且表现出相同两个维度的性状变异, 因此植物性状关系可以概括到全球性状空间的边缘, 为全球植物群落变化的预测提供信息[73]。Dwyer和Laughlin提出了环境压力梯度与性状关联之间的关系, 得出在生境严苛的地区, 植物性状的关联程度会更高[74]。Cornelissen等建立了全球植物功能性状分类体系及其详细的测量方法, 挑选出具有代表性的对环境改变有较强预测能力且能解决大尺度生态问题的28种关键功能性状指标[2], Pérez-Harguindeguy等补充扩展并细化了该体系, 完善了包括植物整体(根、茎、叶)功能性状、地上部分和地下部分功能性状的植物功能性状研究体系[75]。孟婷婷等根据区域生态系统特征补充修订了适合于我国植物功能性状的分类体系[76]。
荒漠植物研究过程中, 如何针对性的选择性状的优化类型与最佳数量来研究群落构建和生态系统功能, 国际目前尚无定论[77]。综合植物功能性状分类体系与指标手册中的标准方法(用于全球范围内植物功能性状间的可比性)[2, 75], 荒漠植物功能性状主要分为形态结构、化学计量、水力特征、生理生态功能性状四种类型。我们认为荒漠植物功能性状权衡策略的研究今后将集中在以下几个方面, 主要包括:在自然环境中优势种植物叶片性状间、叶片与茎枝性状间、叶片与根性状间以及整株植物性状间的权衡策略;优势种荒漠植物地上、地下关键性状相互权衡及联系;优势种荒漠植物在种内、种间水平上对于环境梯度的权衡作用与性状调整;植物资源权衡策略驱使的功能性状的形成和演变可能依赖于不同研究尺度和环境因子的差异;分析植物功能性状在器官、个体、物种、群落间的变异与成因;考虑到植物叶片在荒漠区环境中的功能特征和敏感地位, 主要研究在干旱区降水梯度下叶片性状间的权衡, 整株各部分根-茎-叶-植株性状相互之间交互作用及协同关系需深入研究;环境因子对植物资源获取方式的限制性作用, 土壤结构与营养对权衡策略的影响;最终阐明荒漠植物特有的生态策略和相应的性状关系。
3 荒漠植物功能多样性组分及测度 3.1 荒漠植物功能性状多样性功能多样性作为一种影响群落结构和生态系统功能的成分, 或有机体性状的数值、范围和分布, 强调了物种之间功能性的总体区别或多样性, 表征群落功能性空间变化的特征[78]。功能多样性是生态系统运行与发挥作用的重要因素, 研究功能多样性可以有效地把个体或物种的功能性状与群落和生态系统的功能特征联系在一起[79];能够预测群落中物种的过程变化对生态系统的影响[80—81]。生物多样性对生态系统的影响, 应归功于植物个体的功能性状(数值、大小及范围)以及其相互作用(如直接或间接的竞争、生物自身对环境的适应、生物改善环境的能力等), 而不仅仅是单一物种本身的数量[82—83]。植物的功能特性和它的功能多样性被认为是解释生物多样性与生态系统的功能关联之有效途径, 相对于物种多样性, 功能多样性对环境胁迫或扰动更加敏感, 更能指示生态系统功能的变化[83—84]。对荒漠植物功能性状及功能多样性的研究可增加对干旱区荒漠植物物种竞争共存、群落结构优化、生物多样性维持、生态系统功能改善的新认识, 同时也可以为荒漠生态系统的管理和资源保护以及可持续利用提供理论基础和科学依据。近年来随着对功能多样性认知的发展, 在全球范围内围绕功能多样性主题开展了提供生态理解, 预测环境变化, 评估人类干扰, 优化管理与保护实践, 评价恢复工程等方面的生态学研究工作;在生境梯度水平下主要被应用于山地、草地、灌丛、农田、湿地、冻土、海岸、河岸植物群落研究中[85]。功能多样性研究的核心是选择最适合的功能多样性指数(即组分)来预测和评估生态系统不同功能[86], 然而, 功能多样性最佳表达指数以及功能冗余机理仍然不清楚[87], Mason和Villeger等强调, 衡量功能多样性可能没有一个“最佳”指数, 并倡导功能多样性指数应灵活选择且应该基于具体的科学研究目的和对象[88—89]。量化功能多样性指数对了解荒漠植被生物多样性、生态系统功能及其环境关联性而言十分重要, 并有助于揭示生物地理中的生态系统功能的变化机制[90]。
3.2 荒漠植物功能性状多样性测度荒漠植物功能多样性指数的选择要考虑这些物种自身在群落内的优势程度。根据荒漠植物的关键功能性状量化功能多样性指数, 在选择指数模型时需要考虑:植物关键性状的选择;性状在群落中的比例;性状多样性的统计测定是否有合理的数学特点;不同尺度上植物性状的整合。功能多样性指数能够解释和预测生态系统特征的变异, 并通过统计学检验各项指数的预测能力。只选少数优势种功能多样性指数的计算结果会偏低估计其真实功能多样性, 根据环境梯度优化功能多样性指标, 同时根据性状测定手册, 选择的物种应按照多度大小顺序选择, 涵盖总生物量的80%以上。
衡量功能多样性最为广泛接受和使用的是Mason提出的方法, 功能多样性包含四个相互联系的组分:群落均值加权后的性状值(community-weighted mean, CWM)、功能丰富度(functional richness, FRic)、功能均匀度(functional evenness, FEve), 功能分歧度(functional divergence, FDiv)[88]。功能多样性的不同组分可以从不同角度反映群落的生态位占据状况和资源利用程度[91], 功能丰富性代表现有物种所占据的生态空间规模, 而较低的表示群落中有部分可利用的生态位没有被利用或部分资源未被利用, 反之群落内所有资源都得到了充分利用[92];功能均匀度衡量物种性状平均值在性状空间里分布的是否规律, 该组分较高表示群落性状分布较为规律均匀, 各种资源均被充分利用;功能分歧度描述性状空间内物种簇间的距离或差异程度, 功能分歧度较高的群落可以通过提高资源利用率来增加生态系统的功能[93];特别值得注意的是Rao′s二次熵函数, 它包含了功能丰富度和功能分歧度, 决定于功能空间的占据范围以及物种之间的相似度[94]。这些组分反映了性状在空间分布的互补性, 正确识别这些组分的偏向性可以避免得出错误的结论并剔除一些冗余指数, 单一组分无法准确代表功能多样性, 应该对各组分分别定量计算[95](表 2)。
| 指数 Index |
计算公式 Computational formula |
描述 Description |
含多度 Contains abundance |
参数含义 Parameter meaning |
特征 Characteristic |
| 功能丰富度 Functional richness |
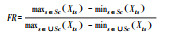 |
个体的功能范围 | 否 | s, 物种;c, 群落;t, 性状;Xts, 物种s的平均性状[88] | 一维度 |
| FRic=包含功能性状空间内所有物种的最小功能体积 | 功能体积 | 否 | 包含功能性状空间内所有物种的最小功能体积[96] | 多维度、物种数需要多余性状数 | |
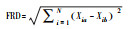 |
聚类树图分支长度总和 | 否 | Xia, Xib是物种a, b的性状i的值[94] | 多维度、很难解释且计算复杂 | |
| 功能均匀度 Functional evenness |
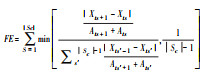 |
性状值均匀度 | 是 | Sc, 群落c中的物种数, Ats, 按照性状t排序的物种丰富度[88] | 一维度 |
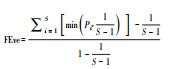 |
聚类最小分支长均匀度 | 是 | Pi′为物种i′相对性状值;S为物种数[89] | 多维度 | |
| 功能分歧度 Functional divergence |
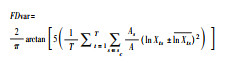 |
性状的对数变异 | 是 | t, 性状;Xts, 物种s的平均性状;Ats, 按照性状t排序的物种丰富度[97] | 一维度;不为零值 |
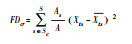 |
性状变异 | 是 | t, 性状;Xts, 物种S的平均性状;Ats, 按照性状t排序的物种丰富度[98] | 一维度 | |
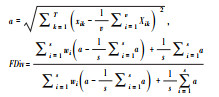 |
距离重心的方差 | 是 | T为所有的功能性状, Xik为物种i在性状轴k上的位置, v为物种i占据的功能空间体积, wi为物种i的相对丰度[89] | 多维度 |
群落水平功能性状值(CWM)是由测定的物种水平的功能性状值, 以物种多度为基础加权平均得到各个性状在群落水平的平均值[3]。根据确定的关键性状值计算CWM以代表功能多样性的不同方面。
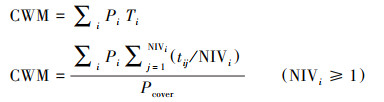
|
式中, Pi是群落中物种i的相对多度, Ti是观测样地中物种i的性状值, NIV是物种i的性状个体值的数量, tij是物种i中F给定性状值j, Pcover是给定群落中所有物种的比例累计相对多度值。物种的相对多度定义为每个物种的生物量以样方中所有物种的生物量总和。
Rao′s二次熵函数指数集中了物种丰富性和物种之间的功能性状差异, 比其他功能多样性指数更适合于表征群落中的种间趋异性, 并更好的将物种丧失与生态系统的功能变化联系在一起。该指数包括两个重要信息:如果一个群落内包含了若干个物种, 那么该群落中每两个物种之间的功能性距离总和可以被视为该群落功能多样性的一个指标, 同时也包含了每个物种在群落内的相对多度的信息。使用R软件进行计算, Rao′s二次熵指数(Rao′s quadratic entropy, FDQ)计算公式如下:
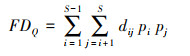
|
式中, S为群落样方内的物种数, pi和pj分别为第i和第j个物种的物种丰富度, 即相对多度, 则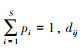
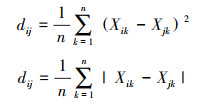
|
式中, n为性状数, Xik为物种i的k性状值。
所有功能多样性指数均可通过R(R Development Core Team 2013)软件vegan包和FD包量化计算[99]。
4 展望随着对荒漠植物特殊功能机制认识的深入, 必须从区域尺度上探讨功能多样性与资源权衡策略的科学问题, 从而阐释群落结构优化与生态系统功能改善的机理, 这是全新的途径和研究思路, 在这一新思路指导下探寻并筛选反映荒漠植物特征的关键性状并用其量化功能多样性进而分析与资源权衡策略的关系是今后研究的重点和难点。
综上所述, 荒漠生态系统是重要的生态屏障, 承载着巨大、多样的生态系统服务功能, 选择这一特殊生境甄选和鉴定关键的生活史性状是亟待努力的方向;探讨不同水平与尺度之间的性状差异与关联的原因及后果是植物功能生态学的重要旨趣;探索植物功能性状分布及多样性格局是理解群落结构及构建最有力的手段之一, 将为物种共存与生物多样性维持运行机制提供深刻的洞见;阐释功能性状组合对有限资源利用的权衡及协变策略是反映荒漠植物适应极端生境、表现特殊功能的必要前提;阐明环境因素与功能性状的关系可谓是生态学中历久弥新的议题。
| [1] |
苏培玺, 严巧娣. 内陆黑河流域植物稳定碳同位素变化及其指示意义. 生态学报, 2008, 28(4): 1616-1624. DOI:10.3321/j.issn:1000-0933.2008.04.032 |
| [2] |
Cornelissen J H C, Lavorel S, Garnier E, Díaz S, Buchmann N, Gurvich D E, Reich P B, Ter Steege H, Morgan H D, Van Der Heijden M G A, Pausas J G, Poorter H. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Australian Journal of Botany, 2003, 51(4): 335-380. DOI:10.1071/BT02124 |
| [3] |
He N P, Liu C C, Piao S L, Sack L, Xu L, Luo Y Q, He J S, Han X G, Zhou G S, Zhou X H, Lin Y, Yu Q, Liu S R, Sun W, Niu S L, Li S G, Zhang J H, Yu G R. Ecosystem traits linking functional traits to macroecology. Trends in Ecology & Evolution, 2019, 34(3): 200-210. |
| [4] |
Funk J L, Cornwell W K. Leaf traits within communities: Context may affect the mapping of traits to function. Ecology, 2013, 94(9): 1893-1897. DOI:10.1890/12-1602.1 |
| [5] |
安申群, 贡璐, 朱美玲, 李红林, 解丽娜, 罗艳. 塔里木盆地北缘典型荒漠植物根系化学计量特征及其与土壤理化因子的关系. 生态学报, 2017, 37(16): 5444-5450. |
| [6] |
单立山, 李毅, 张正中, 种培芳, 王洋, 苏铭, 王珊. 2种典型荒漠植物细根序级结构及功能特征分析. 草地学报, 2017, 25(5): 1014-1019. |
| [7] |
刘涛, 王百田, 曹琪琪, 王文涛, 林晓光, 肖辉杰. 不同覆盖措施对盐碱地紫穗槐细根适应策略的影响. 草业科学, 2020, 37(6): 1098-1106. |
| [8] |
李新乐, 张景波, 董雪, 辛智鸣, 段瑞兵, 罗凤敏, 李永华. 模拟增雨对荒漠植物幼苗生长和根系形态的影响. 生态学报, 2020, 40(10): 3452-3461. |
| [9] |
Gibbens R P, Lenz J M. Root systems of some Chihuahuan Desert plants. Journal of Arid Environments, 2001, 49(2): 221-263. DOI:10.1006/jare.2000.0784 |
| [10] |
Devitt D A, Smith S D. Root channel macropores enhance downward movement of water in a Mojave Desert ecosystem. Journal of Arid Environments, 2002, 50(1): 99-108. DOI:10.1006/jare.2001.0853 |
| [11] |
Riveros C V, Villagra P E, Greco S A. Different root strategies of perennial native grasses under two contrasting water availability conditions: implications for their spatial distribution in desert dunes. Plant Ecology, 2020, 221(7): 633-646. DOI:10.1007/s11258-020-01038-9 |
| [12] |
邓荣华, 高瑞如, 刘后鑫, 赵亚锦, 朱广龙, 魏学智. 自然干旱梯度下的酸枣表型变异. 生态学报, 2016, 36(10): 2954-2961. |
| [13] |
徐世琴, 吉喜斌, 金博文. 典型荒漠植物沙拐枣茎干液流密度动态及其对环境因子的响应. 应用生态学报, 2016, 27(2): 345-353. DOI:10.13287/j.1001-9332.201602.033 |
| [14] |
Ávila‐Lovera E, Zerpa A J, L Santiago L S. Stem photosynthesis and hydraulics are coordinated in desert plant species. New Phytologist, 2017, 216(4): 1119-1129. DOI:10.1111/nph.14737 |
| [15] |
Ávila-Lovera E, Garcillán P P. Phylogenetic signal and climatic niche of stem photosynthesis in the Mediterranean and desert regions of California and Baja California Peninsula. American Journal of Botany, 2020, 108(2): 334-345. |
| [16] |
马天光, 李向义, 林丽莎, 李磊, 李世民, 王波. 遮阴对骆驼刺叶性状和水分生理的影响. 生态学报, 2018, 38(23): 8466-8474. |
| [17] |
何茂松, 罗艳, 彭庆文, 杨思琪, 李凯辉, 韩文轩. 新疆67种荒漠植物叶碳氮磷计量特征及其与气候的关系. 应用生态学报, 2019, 30(7): 2171-2180. DOI:10.13287/j.1001-9332.201907.025 |
| [18] |
张雪妮, 李岩, 何学敏, 吕光辉. 水盐变化对荒漠植物功能多样性与物种多样性关系的影响. 生态学杂志, 2019, 38(8): 2354-2360. DOI:10.13292/j.1000-4890.201908.003 |
| [19] |
李善家, 王辉, 苟伟, 伍国强, 苏培玺. 混生荒漠植物叶片功能性状与其根际微生物多样性的关系. 生态环境学报, 2020, 29(9): 1713-1722. DOI:10.16258/j.cnki.1674-5906.2020.09.001 |
| [20] |
冀雯霞, 赵峰侠, 张瑞, 唐学娟, 闫丽飞, 张锦涛. 晋西北黄土丘陵区土壤和地形因子对草地植被优势种功能性状的影响. 生态学杂志, 2021, 40(3): 749-758. DOI:10.13292/j.1000-4890.202103.016 |
| [21] |
Gibbens R P, Hicks R A, Dugas W A. Structure and function of C3 and C4 Chihuahuan Desert plant communities. Standing crop and leaf area index. Journal of Arid Environments, 1996, 34(1): 47-62. DOI:10.1006/jare.1996.0092 |
| [22] |
Hegazy A, El Amry M. Leaf temperature of desert sand dune plants: perspectives on the adaptability of leaf morphology. African Journal of Ecology, 1998, 36(1): 34-43. DOI:10.1046/j.1365-2028.1998.109-89109.x |
| [23] |
Lawson T, Davey P A, Yates S A, Bechtold U, Baeshen M, Baeshen N, Mutwakil M Z, Sabir J, Baker N R, Mullineaux P M. C3 photosynthesis in the desert plant Rhazya stricta is fully functional at high temperatures and light intensities. New Phytologist, 2014, 201(3): 862-873. DOI:10.1111/nph.12559 |
| [24] |
Castellanos A E, Llano-Sotel J Mo, Machado-Encinas L I, López-Piña J E, Romo-Leon J R, Sardans J, Peñuelas J. Foliar C, N, and P stoichiometry characterize successful plant ecological strategies in the Sonoran Desert. Plant Ecology, 2018, 219(7): 775-788. DOI:10.1007/s11258-018-0833-3 |
| [25] |
Newingham B A, Vanier C H, Charlet T N, Ogle K, Smith S D, Nowak R S. No cumulative effect of 10 years of elevated[CO2] on perennial plant biomass components in the Mojave Desert. Global Change Biology, 2013, 19(7): 2168-2181. DOI:10.1111/gcb.12177 |
| [26] |
刘晓娟, 马克平. 植物功能性状研究进展. 中国科学: 生命科学, 2015, 45(4): 325-339. |
| [27] |
Cadotte M W, Cavender-Bares J, Tilman D, Oakley T H. Using phylogenetic, functional and trait diversity to understand patterns of plant community productivity. PLoS One, 2009, 4(5): e5695. DOI:10.1371/journal.pone.0005695 |
| [28] |
金鹰, 王传宽. 植物叶片水力与经济性状权衡关系的研究进展. 植物生态学报, 2015, 39(10): 1021-1032. DOI:10.17521/cjpe.2015.0099 |
| [29] |
Blonder B, Violle C, Enquist B J. Assessing the causes and scales of the leaf economics spectrum using venation networks in Populus tremuloides. Journal of Ecology, 2013, 101(4): 981-989. DOI:10.1111/1365-2745.12102 |
| [30] |
Westoby M. A leaf-height-seed (LHS) plant ecology strategy scheme. Plant and Soil, 1998, 199(2): 213-227. DOI:10.1023/A:1004327224729 |
| [31] |
Vendramini F, Díaz S, Gurvich D E, Wilson P J, Thompson K, Hodgson J G. Leaf traits as indicators of resource-use strategy in floras with succulent species. New Phytologist, 2002, 154(1): 147-157. DOI:10.1046/j.1469-8137.2002.00357.x |
| [32] |
刘玉冰, 李新荣, 李蒙蒙, 刘丹, 张雯莉. 中国干旱半干旱区荒漠植物叶片(或同化枝)表皮微形态特征. 植物生态学报, 2016, 40(11): 1189-1207. DOI:10.17521/cjpe.2016.0129 |
| [33] |
李善家, 苏培玺, 张海娜, 周紫鹃, 解婷婷. 荒漠植物叶片水分和功能性状特征及其相互关系. 植物生理学报, 2013, 49(2): 153-160. |
| [34] |
Wang M, Wan P C, Guo J C, Xu J S, Chai Y F, Yue M. Relationships among leaf, stem and root traits of the dominant shrubs from four vegetation zones in Shaanxi Province, China. Israel Journal of Ecology and Evolution, 2017, 63(2): 25-32. DOI:10.1163/22244662-06301005 |
| [35] |
Chave J, Coomes D, Jansen S, Lewis S L, Swenson N G, Zanne A E. Towards a worldwide wood economics spectrum. Ecology Letters, 2009, 12(4): 351-366. DOI:10.1111/j.1461-0248.2009.01285.x |
| [36] |
胡云, 燕玲, 李红. 14种荒漠植物茎的解剖结构特征分析. 干旱区资源与环境, 2006, 20(1): 202-208. DOI:10.3969/j.issn.1003-7578.2006.01.040 |
| [37] |
Ma Z Q, Guo D L, Xu X L, Lu M Z, Bardgett R D, Eissenstat D M, McCormack M L, Hedin L O. Evolutionary history resolves global organization of root functional traits. Nature, 2018, 555(7694): 94-97. DOI:10.1038/nature25783 |
| [38] |
徐冰, 程雨曦, 甘慧洁, 周文嘉, 贺金生. 内蒙古锡林河流域典型草原植物叶片与细根性状在种间及种内水平上的关联. 植物生态学报, 2010, 34(1): 29-38. |
| [39] |
Holdaway R J, Richardson S J, Dickie I A, Peltzer D A, Coomes D A. Species- and community-level patterns in fine root traits along a 120000-year soil chronosequence in temperate rain forest. Journal of Ecology, 2011, 99(4): 954-963. DOI:10.1111/j.1365-2745.2011.01821.x |
| [40] |
欧阳园丽, 张参参, 林小凡, 田立新, 顾菡娇, 陈伏生, 卜文圣. 中国亚热带不同菌根树种的根叶形态学性状特征与生长差异: 以江西新岗山为例. 生物多样性, 2021, 29(06): 746-758. |
| [41] |
单立山, 李毅, 任伟, 苏世平, 董秋莲, 耿东梅. 河西走廊中部两种荒漠植物根系构型特征. 应用生态学报, 2013, 24(1): 25-31. |
| [42] |
杨小林, 张希明, 李义玲, 李绍才, 孙海龙. 塔克拉玛干沙漠腹地3种植物根系构型及其生境适应策略. 植物生态学报, 2008, 32(6): 1268-1276. DOI:10.3773/j.issn.1005-264x.2008.06.007 |
| [43] |
Reich P B. The world-wide'fast-slow'plant economics spectrum: a traits manifesto. Journal of Ecology, 2014, 102(2): 275-301. |
| [44] |
Roumet C, Birouste M, Picon-Cochard C, et al. Root structurefunction relationships in 74 species: evidence of a root economics spectrum related to carbon economy. New Phytologist, 2016, 210(3): 815-826. |
| [45] |
Lavorel S, Grigulis K. How fundamental plant functional trait relationships scale-up to trade-offs and synergies in ecosystem services. Journal of Ecology, 2012, 100(1): 128-140. |
| [46] |
Díaz S, Kattge J, Cornelissen J H C, Wright I J, Lavorel S, Dray S, Reu B, Kleyer M, Wirth C, Colin Prentice I, Garnier E, Bönisch G, Westoby M, Poorter H, Reich P B, Moles A T, Dickie J, Gillison A N, Zanne A E, Chave J, Joseph Wright S, Sheremet'ev S N, Jactel H, Baraloto C, Cerabolini B, Pierce S, Shipley B, Kirkup D, Casanoves F, Joswig J S, Günther A, Falczuk V, Rüger N, Mahecha M D, Gorné L D. The global spectrum of plant form and function. Nature, 2016, 529(7585): 167-171. |
| [47] |
Ordoñez J C, Van Bodegom P M, Witte J P M, Wright I J, Reich P B, Aerts R. A global study of relationships between leaf traits, climate and soil measures of nutrient fertility. Global Ecology and Biogeography, 2009, 18(2): 137-149. |
| [48] |
Wright I J, Dong N, Maire V, Prentice I C, Westoby M, Díaz S, Gallagher R V, Jacobs B F, Kooyman R, Law E A, Leishman M R, Niinemets V, Reich P B, Sack L, Villar R, Wang H, Wilf P. Global climatic drivers of leaf size. Science, 2017, 357(6354): 917-921. |
| [49] |
Butler EE, Datta A, Flores-Moreno H, Chen M, Wythers K R, Fazayeli F, Banerjee A, Atkin O K, Kattge J, Amiaud B, Blonder B, Boenisch G, Bond-Lamberty B, Brown K A, Byun C, Campetella G, Cerabolini B E L, Cornelissen J H C, Craine J M, Craven D, Vries F T D, Díaz S, Domingues T F, Forey E, González-Melo A, Gross N, Han W X, Hattingh W N, Hickler T, Jansen S, Kramer K, Kraft N J B, Kurokawa H, Laughlin D C, Meir P, Minden V, Niinemets V, Onoda Y, Peñuelas J, Read Q, Sack L, Schamp B, Soudzilovskaia N A, Spasojevic M J, Sosinski E, Thornton P E, Valladares F, Van Bodegom P M, Williams M, Wirth C, Reich P B. Mapping local and global variability in plant trait distributions. Proceedings of the National Academy of Sciences, 2017, 114(51): E10937-E10946. |
| [50] |
陈莹婷, 许振柱. 植物叶经济谱的研究进展. 植物生态学报, 2014, 38(10): 1135-1153. |
| [51] |
Mouillot D, Villeger S, Scherer-Lorenzen M, Mason N W H. Functional structure of biological communities predicts ecosystem multifunctionality. PLoS One, 2011, 6(3): e17476. |
| [52] |
何芸雨, 郭水良, 王喆. 植物功能性状权衡关系的研究进展. 植物生态学报, 2019, 43(12): 1021-1035. |
| [53] |
张科, 田长彦, 李春俭. 盐土和沙土对新疆常见一年生盐生植物生长和体内矿质组成的影响. 生态学报, 2012, 32(10): 3069-3076. |
| [54] |
Lanta V, Lepš J. Effect of functional group richness and species richness in manipulated productivity-diversity studies: a glasshouse pot experiment. Acta Oecologica, 2006, 29(1): 85-96. |
| [55] |
Rudolf V H W, Rasmussen N L. Ontogenetic functional diversity: size structure of a keystone predator drives functioning of a complex ecosystem. Ecology, 2013, 94(5): 1046-1056. |
| [56] |
Wright I J, Reich P B, Westoby M, Ackerly D D, Baruch Z, Bongers F, Cavender-Bares J, Chapin T, Cornelissen J H C, Diemer M, Flexas J, Garnier E, Groom P K, Gulias J, Hikosaka K, Lamont B B, Lee T, Lee W, Lusk C, Midgley J J, Navas M L, Niinemets V, Oleksyn J, Osada N, Poorter H, Poot P, Prior L, Pyankov V I, Roumet C, Thomas S C, Tjoelker M G, Veneklaas E J, Villar R. The worldwide leaf economics spectrum. Nature, 2004, 428(6985): 821-827. |
| [57] |
Cui E Q, Weng E S, Yan E R, Xia J Y. Robust leaf trait relationships across species under global environmental changes. Nature Communications, 2020, 11(1): 2999. |
| [58] |
He J S, Wang Z H, Wang X P, Schmid B, Zuo W Y, Zhou M, Zheng C Y, Wang M F, Fang J Y. A test of the generality of leaf trait relationships on the Tibetan Plateau. New Phytologist, 2006, 170(4): 835-848. |
| [59] |
Yin QL, Wang L, Lei M L, Dang H, Quan J X, Tian T T, Chai Y F, Yue M. The relationships between leaf economics and hydraulic traits of woody plants depend on water availability. Science of the Total Environment, 2018, 621: 245-252. |
| [60] |
李镯, 党晓宏, 汪季, 冯亚亚, 崔健. 吉兰泰盐湖周边荒漠植物养分、叶片功能性状及适应策略研究. 生态环境学报, 2021, 30(4): 708-717. |
| [61] |
李军军, 李萌茹, 齐兴娥, 王立龙, 徐世健. 芨芨草叶片养分特征对氮磷不同添加水平的响应. 植物生态学报, 2020, 44(10): 1050-1058. |
| [62] |
Rodríguez-Gallego C, Navarro T, Meerts P. A comparative study of leaf trait relationships in coastal dunes in southern Spain. Plant Ecology and Evolution, 2015, 148(1): 57-67. |
| [63] |
Reich P B, Tjoelker M G, Pregitzer K S, Wright I J, Oleksyn J, Machado J L. Scaling of respiration to nitrogen in leaves, stems and roots of higher land plants. Ecology Letters, 2008, 11(8): 793-801. |
| [64] |
Ishida A, Nakano T, Yazaki K, Matsuki S, Koike N, Lauenstein D L, Shimizu M, Yamashita N. Coordination between leaf and stem traits related to leaf carbon gain and hydraulics across 32 drought-tolerant angiosperms. Oecologia, 2008, 156(1): 193-202. |
| [65] |
De La Riva E G, Tosto A, Pérez-Ramos I M, Navarro-Fernández C M, Olmo M, Anten N P R, Marañón T, Villar R. A plant economics spectrum in Mediterranean forests along environmental gradients: Is there coordination among leaf, stem and root traits?. Journal of Vegetation Science, 2016, 27(1): 187-199. |
| [66] |
Wright S J, Kitajima K, Kraft N J B, Reich P B, Wright I J, Bunker D E, Condit R, Dalling J W, Davies S J, Díaz S, Engelbrecht B M J, Harms K E, Hubbell S P, Marks C O, Ruiz-Jaen M C, Salvador C M, Zanne A E. Functional traits and the growth-mortality trade-off in tropical trees. Ecology, 2010, 91(12): 3664-3674. |
| [67] |
Prieto I, Roumet C, Cardinael R, Dupraz C, Jourdan C, Kim J H, Maeght J L, Mao Z, Pierret A, Portillo N, Roupsard O, Thammahacksa C, Stokes A. Root functional parameters along a land-use gradient: evidence of a community-level economics spectrum. Journal of Ecology, 2015, 103(2): 361-373. |
| [68] |
马芳, 曾辉, 李洪波, 马泽清, 郭大立. 内蒙古草原植物根属性的变异格局及环境适应策略. 北京大学学报: 自然科学版, 2019, 55(2): 387-396. |
| [69] |
马泽清, 郭大立, 徐兴良, 卢明镇, Bardgett R D, Eissenstat D M, McCormack M L, Hedin L O. 植物根功能属性的组织方式及进化趋势//2018全国植物生物学大会论文集. 泰安: 中国作物学会, 2018: 45-45.
|
| [70] |
郑慧玲, 赵成章, 徐婷, 段贝贝, 韩玲, 冯威. 红砂根系分叉数和分支角度权衡关系的坡向差异. 植物生态学报, 2015, 39(11): 1062-1070. |
| [71] |
Kong D L, Wang J J, Wu H F, Valverde-Barrantes O J, Wang R L, Zeng H, Kardol P, Zhang H Y, Feng Y L. Nonlinearity of root trait relationships and the root economics spectrum. Nature Communications, 2019, 10(1): 2203. |
| [72] |
Fortunel C, Fine P V A, Baraloto C. Leaf, stem and root tissue strategies across 758 neotropical tree species. Functional Ecology, 2012, 26(5): 1153-1161. |
| [73] |
Thomas H J D, Bjorkman A D, Myers-Smith I H, Elmendorf S C, Kattge J, Diaz S, Vellend M, Blok D, Cornelissen J H C, Forbes B C, Henry G H R, Hollister R D, Normand S, Prevéy J S, Rixen C, Schaepman-Strub G, Wilmking M, Wipf S, Cornwell W K, Beck P S A, Georges D, Goetz S J, Guay K C, Rüger N, Soudzilovskaia N A, Spasojevic M J, Alatalo J M, Alexander H D, Anadon-Rosell A, Angers-Blondin S, te Beest M, Berner L T, Björk R G, Buchwal A, Buras A, Carbognani M, Christie K S, Collier L S, Cooper E J, Elberling B, Eskelinen A, Frei E R, Grau O, Grogan P, Hallinger M, Heijmans M M P D, Kaarlejarvi E, Kulonen A, Lamarque L J, Lantz T C, Lévesque E, Little C J, Michelsen A, Milbau A, Nabe-Nielsen J, Nielsen S S, Ninot J M, Oberbauer S F, Olofsson J, Onipchenko V G, Petraglia A, Rumpf S B, Shetti R, Speed J D M, Suding K N, Tape K D, Tomaselli M, Trant A J, Treier U A, Tremblay M, Venn S E, Vowles T, Weijers S, Wookey P A, Zamin T J, Bahn M, Blonder B, Van Bodegom P M, Bond-Lamberty B, Campetella G, Cerabolini B E L, Chapin Ⅲ F S, Craine J M, Dainese M, Green W A, Jansen S, Kleyer M, Manning P, Niinemets V, Onoda Y, Ozinga W A, Peñuelas J, Poschlod P, Reich P B, Sandel B, Schamp B S, Sheremetiev S N, De Vries F T. Global plant trait relationships extend to the climatic extremes of the tundra biome. Nature Communications, 2020, 11(1): 1351. |
| [74] |
Dwyer J M, Laughlin D C. Constraints on trait combinations explain climatic drivers of biodiversity: the importance of trait covariance in community assembly. Ecology Letters, 2017, 20(7): 872-882. |
| [75] |
Pérez-Harguindeguy N, Díaz S, Garnier E, Lavorel S, Poorter H, Jaureguiberry P, Bret-Harte M S, Cornwell W K, Craine J M, Gurvich D E, Urcelay C, Veneklaas E J, Reich P B, Poorter L, Wright I J, Ray P, Enrico L, Pausas J G, De Vos A C, Buchmann N, Funes G, Quétier F, Hodgson J G, Thompson K, Morgan H D, Ter Steege H, Van Der Heijden M G A, Sack L, Blonder B, Poschlod P, Vaieretti M V, Conti G, Staver A C, Aquino S, Cornelissen J H C. New handbook for standardised measurement of plant functional traits worldwide. Australian Journal of Botany, 2013, 61(3): 167-234. |
| [76] |
孟婷婷, 倪健, 王国宏. 植物功能性状与环境和生态系统功能. 植物生态学报, 2007, 31(1): 150-165. |
| [77] |
Lozanovska I, Ferreira M T, Aguiar F C. Functional diversity assessment in riparian forests-multiple approaches and trends: A review. Ecological Indicators, 2018, 95(1): 781-793. |
| [78] |
Petchey O L, Gaston K J. Functional diversity (FD), species richness and community composition. Ecology Letters, 2002, 5(3): 402-411. |
| [79] |
Wiegand T, Uriarte M, Kraft N J B, Shen G C, Wang X G, He F L. Spatially explicit metrics of species diversity, functional diversity, and phylogenetic diversity: insights into plant community assembly processes. Annual Review of Ecology, Evolution and Systematics, 2017, 48: 329-351. |
| [80] |
Chapin Ⅲ F S, Zavaleta E S, Eviner V T, Naylor R L, Vitousek P M, Reynolds H L, Hooper D U, Lavorel S, Sala O E, Hobbie S E, Mack M C, Díaz S. Consequences of changing biodiversity. Nature, 2000, 405(6783): 234-242. |
| [81] |
Nunes A, Köbel M, Pinho P, Matos P, Bello F D, Correia O, Branquinho C. Which plant traits respond to aridity? A critical step to assess functional diversity in Mediterranean drylands. Agricultural and Forest Meteorology, 2017, 239: 176-184. |
| [82] |
Tilman D. The ecological consequences of changes in biodiversity: a search for general principles. Ecology, 1999, 80(5): 1455-1474. |
| [83] |
Lavorel S, Storkey J, Bardgett R D, Bello F D, Berg M P, Roux X L, Moretti M, Mulder C, Pakeman R J, Díaz S, Harrington R. A novel framework for linking functional diversity of plants with other trophic levels for the quantification of ecosystem services. Journal of Vegetation Science, 2013, 24(5): 942-948. |
| [84] |
Lefcheck J S, Duffy J E. Multitrophic functional diversity predicts ecosystem functioning in experimental assemblages of estuarine consumers. Ecology, 2015, 96(11): 2973-2983. |
| [85] |
Garnier E, Navas M L, Grigulis K. Plant Functional Diversity. Oxford: : Oxford University Press, 2016, 64-88. |
| [86] |
Thomas Clark A, Lehman C, Tilman D. Identifying mechanisms that structure ecological communities by snapping model parameters to empirically observed tradeoffs. Ecology Letters, 2018, 21(4): 494-505. |
| [87] |
Clark C M, Flynn D F B, Butterfield B J, Reich P B. Testing the link between functional diversity and ecosystem functioning in a Minnesota grassland experiment. PLoS One, 2012, 7(12): e52821. |
| [88] |
Mason N W H, Mouillot D, Lee W G, Wilson J B. Functional richness, functional evenness and functional divergence: the primary components of functional diversity. Oikos, 2005, 111(1): 112-118. |
| [89] |
Villéger S, Mason N W H, Mouillot D. New multidimensional functional diversity indices for amultifaceted framework in functional ecology. Ecology, 2008, 89(8): 2290-2301. |
| [90] |
Reichstein M, Bahn M, Mahecha M D, Kattge J, Baldocchi D D. Linking plant and ecosystem functional biogeography. Proceedings of the National Academy of Sciences of the United States of America, 2014, 111(38): 13697-13702. |
| [91] |
Zuo X A, Zhang J, Lv P, Wang S K, Yang Y, Yue X Y, Zhou X, Li Y L, Chen M, Lian J, Qu H, Liu L X, Ma X J. Effects of plant functional diversity induced by grazing and soil properties on above- and belowground biomass in a semiarid grassland. Ecological Indicators, 2018, 93: 555-561. |
| [92] |
Magnago L F S, Edwards D P, Edwards F A, Magrach A, Martins S V, Laurance W F. Functional attributes change but functional richness is unchanged after fragmentation of Brazilian Atlantic forests. Journal of Ecology, 2014, 102(2): 475-485. |
| [93] |
张金屯, 范丽宏. 物种功能多样性及其研究方法. 山地学报, 2011, 29(5): 513-519. |
| [94] |
Mouchet M A, Villéger S, Mason N W H, Mouillot D. Functional diversity measures: an overview of their redundancy and their ability to discriminate community assembly rules. Functional Ecology, 2010, 24(4): 867-876. |
| [95] |
De Bello F, Carmona C P, Lepš J, Szava-Kovats R, Pärtel M. Functional diversity through the mean trait dissimilarity: resolving shortcomings with existing paradigms and algorithms. Oecologia, 2016, 180(4): 933-940. |
| [96] |
Cornwell W K, Schwilk D W, Ackerly D D. A trait-based test for habitat filtering: convex hull volume. Ecology, 2006, 87(6): 1465-1471. |
| [97] |
Mason N W H, Macgillivray K, Steel J B, Wilson J B. An index of functional diversity. Journal of Vegetation Science, 2003, 14(4): 571-578. |
| [98] |
Leps J, De Bello F, Lavorel S, Berman S. Quantifying and interpreting functional diversity of natural communities: Practical considerations matter. Preslia, 2006, 78(4): 481-501. |
| [99] |
Laliberté E, Legendre P. A distance-based framework for measuring functional diversity from multiple traits. Ecology, 2010, 91(1): 299-305. |