 2022, Vol. 42
2022, Vol. 42文章信息
- 祖悦晴, 魏妍儿, 张曦文, 于德永
- ZU Yueqing, WEI Yaner, ZHANG Xiwen, YU Deyong
- 生态保护区外斑块对景观连接度的影响评价
- Evaluation of the importance of patches outside protected areas to landscape connectivity
- 生态学报. 2022, 42(17): 6937-6947
- Acta Ecologica Sinica. 2022, 42(17): 6937-6947
- http://dx.doi.org/10.5846/stxb202109052505
-
文章历史
- 收稿日期: 2021-09-05





当今全球范围内正在经历着前所未有规模的城市化过程[1], 这种剧烈的土地利用改变会带来包括土地覆被、气候、水文、生物地球化学循环和生物多样性等一系列的环境变化及环境问题[2—3]。在大部分城市区域, 生物多样性仅存在于城市发展过程中保留的处于孤立和破碎化的生境斑块中[4]。城市化导致的土地利用变化过程会使城市生境斑块损失及破碎化, 影响物种运动、捕食等过程, 改变物种组成和种群结构, 最终降低生物多样性[2—6]。因此, 为了保护和维持城市生物群落及生物多样性, 同时考虑城市发展情况, 需要城市规划者确定城市关键保护区域, 采取有效的措施保护现有的自然或半自然生境。目前, 一些国家采取划定生态保护区的方式来管理和保护城市生境及生物多样性, 在生态保护区中限制城市建设[7], 如中国深圳为保障城市生态系统的完整性和连通性, 根据其划定的生态保护范围确定了基本生态控制线;澳大利亚依据生态重要性、保护程度和生态系统的脆弱性, 在大堡礁保护区中划定了生境保护区[8—9]。然而, 已有研究表明生态保护区并不能完全覆盖某些生物生存活动的必要区域和栖息地[10], Leroux等的研究发现生态保护区外临近的生境斑块在保护和维持物种及其生态过程中也起到至关重要的作用[11—12], 但这部分保护区外的生境斑块并没有给予应有的重视。当这些未受到保护的临近生境受到城市化的影响时, 生境会发生损失, 从而影响物种觅食、迁徙等生态活动, 造成生物多样性损失[13]。因此, 忽略生态保护区外潜在对生物活动具有重要生态价值的生境斑块不利于城市生态保护区内生物多样性的整体保护。
景观连接度可以用于生物多样性保护的研究[4]。景观连接度是指景观促进或阻碍生物体或某种生态过程在源斑块间运动的程度[14]。保护和提高景观连接度是在有限的空间范围内应对生境损失和破碎化, 维持生物多样性的有效途径之一[15]。目前景观连接度的量化方法包括图论法、空间格局指数法、最近邻距离法、扩散率法等, 而图论法可以将生境空间情况与生物迁移相结合, 能很好地预测景观潜在连接度, 且适用于保护区等大尺度的景观连接度研究[16]。Bunn等最早将图论法应用于景观连接度的研究中[17]。基于图论法可以定量评估整体景观连接度, 还可以评价各个生境斑块对于景观的重要性。国内外已有部分研究基于图论量化景观连接度和生物多样性保护、城市景观规划相结合。例如, Zetterberg等以斯德哥尔摩地区为例, 利用图论法确定了生境中的重要结构和廊道, 并基于结果进行景观规划设计[18]。于德永等基于图论模型, 对广东省深圳市核心生境的整体景观连接度进行了评价, 确定了生态保护网络和生态廊道, 并应用到实际保护规划方案中[19]。蒙吉军等基于图论模型量化了黑河中游地区景观连接度的年际变化, 并根据特定距离阈值确定了不同规模生态斑块的区域生态重要性[20]。其中一些研究通过模拟移除某斑块后的景观连接度变化来量化其在生态网络中的重要性[21—22]。例如, 荀斌等运用图论模型, 结合连续移除斑块的模拟方法, 判断深圳市生境斑块重要性并确定了生境变化情景[23]。在判断生境斑块重要性时, 仅仅根据斑块属性确定斑块移除顺序可能与城市发展实际情况不符, 较少有研究综合考虑未来城市发展实际情况, 从而影响生态生境变化情景的制定。因此, 为了在满足未来城市可持续发展的需求, 应考虑该城市的未来土地利用变化, 并根据情景结果确定生境斑块的移除顺序, 从而将生物多样性保护与城市发展相结合[24—25]。
北京作为中国的首都, 在过去几十年经历了快速的城市化过程[26], 但是针对当地生态保护区景观格局的研究较少。因此, 本研究基于图论法, 以北京市延庆区为研究区, 划分生境斑块, 并构建生境网络, 评价和模拟城市化过程导致的生境斑块损失对当地不同森林哺乳动物的影响, 量化和分析该地生态保护区外生境斑块对于生物活动的重要性并识别对维持景观连接度具有重要作用的潜在生境斑块。本文主要回答以下科学问题:(1)在不同的生境变化情境下哪些生境斑块对于维持景观连接度有较大影响?(2)当生境景观格局变化时, 对于不同迁移能力的物种, 景观连接度的变化特征有什么不同?本文可以为该地的生物多样性保护和景观格局优化提供科学依据和方法参考。
1 研究区概况与数据来源 1.1 研究区概况北京市延庆区位于北京市西北部(图 1), 西与河北省接壤, 面积1993.75km2, 介于东经115°44′—116°34′、北纬40°16′—40°47′之间。延庆区属大陆季风气候区, 四季分明, 全区多年平均气温为8.8℃, 最高气温为39℃, 最低气温为-27.3℃[27]。该地植被类型为暖温带落叶阔叶林和温带针叶林。全区平均海拔500m以上, 东北部山地平均海拔1000m左右, 南部山地地势较低。

|
| 图 1 延庆区区位图和延庆区2020年土地利用图 Fig. 1 Location and land use map of Yanqing District |
作为国家级生态示范县和首都生态涵养发展区, 延庆区生物多样性丰富, 是北京西北的重要生态屏障。延庆区内的北京松山国家级自然保护区是以丰富的动植物资源为主要保护对象的综合性自然保护区, 总面积为46.71km2[27]。保护区内有多种大型哺乳动物以及鸟兽, 大型野生动物有中华斑铃、狍、野猪、豹猫、猪獾、狗獾、貉等[27—28], 其中中华斑羚为国家二级保护动物。
1.2 数据来源本文获取研究区相关基础地理数据, 将栅格数据的像元大小统一重采样为30m, 经处理后用于后续划分生境斑块、构建阻力面、模拟城市土地利用变化、计算景观连接度。具体的数据类型和数据来源如表 1。
| 数据类型 Data type |
数据来源 Data source |
数据说明 Data description |
| 地形要素数据 Digital line graphic |
国家基础地理信息中心(http://www.ngcc.cn/ngcc/) | 1:25万;包括水系、公路、铁路、居民地数据 |
| 数字高程模型数据 DEM (Digital Elevation Data) |
地理空间数据云(http://www.gscloud.cn/) | 30m分辨率 |
| 植被净初级生产力 NPP (Net Primary Productivity) |
美国地质调查局USGS(http://glovis.usgs.gov/) | MODIS NPP产品数据;2019年7—8月;500m分辨率 |
| 地表覆盖 Land cover |
国家基础地理信息中心(http://www.ngcc.cn/ngcc/) | 2020、2010版;30m分辨率 |
| 行政区边界 Administrative boundary |
资源环境科学与数据中心(http://www.resdc.cn/) | 矢量数据 |
| 生态保护红线 Ecological redline |
北京市人民政府(http://www.beijing.gov.cn/) | 矢量化“北京市生态保护红线分布范围图” |
考虑到生态过程具有空间连续性, 为避免城市的边缘效应, 根据基础地理数据的可用性和保护物种的迁移距离大小, 在延庆区行政区边界周围建立5km缓冲区[25]。城市内的斑块需满足一定要求才能作为森林哺乳动物的栖息地, 从而被划为生境斑块, 设定生境斑块需符合以下三个条件:地表覆盖为林地或灌木、面积大于0.2km2[23]、远离主要道路1km[29]以上。
现有生态保护区对于维护生物繁衍和生物多样性有重要意义, 其内森林和灌木受政策保障而得到严格保护, 因此将包含北京市生态保护红线划定方案内区域的生境斑块划分为核心生境斑块, 共划分出4个核心生境斑块。生态保护区外临近的生境斑块在生物迁移过程中也可能起到重要作用, 因此将位于北京市生态保护红线划定方案外的生境斑块划分为潜在生境斑块, 共划分出74个潜在生境斑块, 按面积从小到大编号(图 2)。核心生境斑块和潜在生境斑块的质心作为生境网络中的节点, 用于后续景观连接度的计算。

|
| 图 2 生境斑块划分结果图 Fig. 2 Map of the spatial distributions of habitat patches |
本研究选用连接概率(Probability of connectivity, PC)指数评价生境网络的景观连接度。连接概率指数被定义为随机放置在某一区域的生境斑块中的两个动物, 能够通过其它斑块和斑块间连接路径彼此相遇的可能性[30], 该指数在景观尺度上度量连接度。该指数越大则表明生境网络连接度越高, 越利于生物的扩散和生态过程的传播。PC指数受到斑块间空间结构、斑块属性和物种迁移能力的影响, 值介于0—1之间。公式如下:
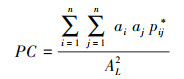
|
(1) |
式中, ai、aj是斑块i和j的质量, 面积越大的斑块能为生物提供更多可利用的资源, 本研究选择斑块面积作为斑块质量[23]。AL是整个区域质量的总和。pij*为斑块i到斑块j所有可能路径连接概率(pij)的乘积中的最大值。pij表示斑块i和j之间直接连接的概率, 其大小为两个斑块之间距离的负指数函数, 公式如下[31]:
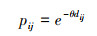
|
(2) |
式中, θ为常量, 是使得物种的迁移距离与迁移概率相匹配的参数, 本文选择迁移概率p=0.5所代表的物种中等扩散距离以确定θ。dij为两个斑块之间的距离, 本研究选用最小耗费路径长度作为斑块间距离, 与欧氏距离相比, 考虑地表阻力的最小耗费路径距离更符合森林物种在斑块间迁移的实际情况。
物种的迁移距离作为其迁移能力的表现, 能够用于判断斑块间是否连接。选择不同物种迁移距离作为参数用于景观连接度指数的计算, 可以评价景观格局变化对不同迁移能力物种的影响。选取延庆区内保护物种并收集其饮食类型与平均体重, 根据Sutherland的公式[32]计算出各物种的估计分散度中位数(表 2), 由此确定物种迁移距离参数为1、8、10km。
| 物种名称 Species names |
饮食类型 Trophic type |
平均体重/kg Average body mass |
公式 Equation |
估计分散度中位数/km Median dispersal distance |
| 中华斑羚 Naemorhedus griseus |
食草动物与杂食动物 | 27 | 1.45(±1.05)×M0.54(±0.01) | 1.45×270.54=8.60 |
| 豹猫 Prionailurus bengalensis |
食肉动物 | 3.4 | 3.45(±1.07)×M0.80(±0.03) | 3.45×3.40.8=9.18 |
| 沟牙鼯鼠 Aeretes melanopterus |
食草动物与杂食动物 | 0.6 | 1.45(±1.05)×M0.54(±0.01) | 1.45×0.60.54=1.10 |
生境斑块对于整个生境网络连接度的贡献程度可用移除某斑块后网络连接度的变化率来表示[31]。变化率值越大, 则斑块在整个网络中越重要、贡献越大。公式如下:
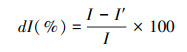
|
(3) |
式中, I为包含某一生境斑块在内的整体网络连接度大小, I′为移除该斑块后整体网络连接度的大小, 本研究中景观连接度大小用PC值表达。
2.3 生境阻力面的构建生境阻力的大小反映了物种迁移的困难程度[33], 生境阻力面的构建是计算最小耗费路径长度的基础。地表覆盖类型、坡度、人类活动[25]等因素均影响物种的迁移过程。较陡的坡度和频繁的人类活动会阻碍物种的运动, 增加生境阻力。植被净初级生产力(Net Primary Productivity, NPP)指绿色植物在单位时间、单位面积内由光合作用所产生的有机质总量减去自身呼吸后的剩余部分[34], 是生物生存和繁衍的物质基础, 本研究选用MODIS的NPP产品数据代表地表覆盖情况(表 1)。对研究区内所有像元, 使用最大最小缩放对NPP、坡度数据层进行1—100的阻力赋值, 使NPP值小、坡度值大的像元阻力值大。最大最小缩放公式如下:
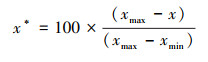
|
(4) |
式中, x*表示缩放后的像元值, x表示缩放前的像元值, xmax表示缩放前研究区内像元最大值, xmin表示缩放前研究区内像元最小值。
水体和河流是大多数森林物种迁移的屏障, 居民地及其周边区域的人类活动频繁且地表覆盖类型不适宜森林动物迁移, 本研究赋予这些地区最高阻力值300。线状交通干线对周围景观具有明显的破碎化和障碍效应, 会阻碍物种的迁移运动过程, 因此依据不同的铁路、公路级别建立不同缓冲区并设定不同阻力值(表 3)[31]。将道路、河流建立缓冲区, 可以防止在栅格化过程中线要素丢失或不连续, 避免出现路径沿对角线跨越道路、河流的情况发生。本研究综合考虑地表覆盖类型、坡度、人类活动等因素, 将NPP数据层、坡度数据层和道路数据层赋予相同的权重, 结合水体、河流和居民地数据层叠加生成生境阻力面。
| 数据层 Data layer |
分级 Classification |
缓冲宽度/m Buffer width |
阻力值 Resistance value |
|
| 叠加数据层 | NPP | 1—100 | ||
| Compound data layer | 坡度 | 1—100 | ||
| 道路 | 铁路 | 1000 | 100 | |
| 一级道路 | 1000 | 100 | ||
| 二级道路 | 800 | 80 | ||
| 三级道路 | 600 | 60 | ||
| 四级道路 | 400 | 40 | ||
| 单独数据层 | 居民地 | 100 | 300 | |
| Single data layer | 河流 | 50 | 300 | |
| 水体 | 300 | |||
| NPP: 植被净初级生产力Net primary productivity | ||||
结合未来土地利用空间格局变化的模拟, 可以研究城市化过程对区域生境网络和景观连接度的影响。本研究选用CLUE-S模型(https://www.environmentalgeography.nl/site/)进行研究区土地利用类型变化的模拟, 该模型能够在小尺度上综合驱动因素和土地利用演变特征, 预测土地利用变化并识别变化的热点地区[35—36]。模型以两个假设为前提条件, 一是模拟区域内土地利用和覆盖明显受到土地需求的驱动;二是研究区域内的土地需求、自然环境和社会经济状况总是与土地利用空间格局保持动态平衡。基于上述假设条件, CLUE-S模型将土地需求结果进行空间分配[37], 实现不同地类之间的相互转化。
模型的输入数据包括模拟范围、驱动因子和各类土地利用类型的需求量。本研究以研究区作为模拟范围, 生态保护红线内区域设置为限制区域, 其土地利用类型不会改变。以2010和2020年的土地利用数据为基础, 考虑移除生境斑块的需求, 线性模拟获取各类土地利用类型的需求量。采用DEM、坡度、距河流距离、距主要道路距离、距居民地距离作为驱动因子, 运用SPSS软件中二元Logistic回归模型计算各地类与驱动因子之间的相关系数。
2.5 生境变化情景的设计城市生境斑块容易受到人类活动干扰和城市化扩展过程的影响, 本研究分析从整个生境网络中移除部分斑块对整体景观连接度的影响。本研究设计了三种生境变化情景[23], 潜在生境斑块在不同生境变化情景中将以不同的顺序从生境网络中移除, 核心生境斑块不移除。情景1, 将潜在生境斑块按面积升序依次单独移除, 每次都基于原始的生境网络单独移除某一个斑块。情景2, 将潜在生境斑块按面积升序连续移除, 每次基于上一次移除斑块后的新生境网络。设计上述单独和连续移除斑块的两个情景用以识别生境网络中能够显著提升景观连接度的生境斑块。情景3, 依据城市土地利用变化模拟的结果判断潜在生境斑块的移除顺序并将其连续移除。将2020年和模拟结果的生境斑块对应, 计算每个斑块面积的变化率ri, 根据变化率由大到小的顺序决定斑块移除先后顺序。其中未来消失的斑块先移除, 面积小的斑块优先移除, 与大斑块合并的小斑块不移除。斑块面积变化率计算公式如下:
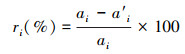
|
(5) |
式中, ri表示斑块i的面积变化率, ai表示2020年斑块i的面积, a′i表示土地利用变化模拟结果中2029年斑块i的面积。该方案模拟了未来生境斑块消失的可能顺序, 连续移除斑块考虑到潜在斑块间的连接作用, 用以识别受到城市化过程影响较大且对网络连接度有较大贡献的生境斑块。
3 结果对于全部三个生境变化情景, PC值所表征的景观连接度随着物种迁移距离的增加而增大(图 3)。因为迁移能力越强的物种越容易通过其他斑块和斑块间连接路径实现迁移过程, 达到两个动物的相遇。物种迁移距离由8km增加至10km时PC值增加0.0048, 由1km增加至8km时PC值增加0.0060, 前者的单位距离PC值增加量远大于后者, 说明对于相同的景观格局变化, 物种迁移距离为8—10km的景观连接度变化更大。

|
| 图 3 三种生境变化情景下生境网络景观连接度变化 Fig. 3 Changes of landscape connectivity of habitat networks under three different habitat change scenarios 景观连接度变化包括连接概率指数PC和网络连接度变化率dI;情景1为斑块依面积由小到大逐个单独移除,情景2为斑块依面积由小到大连续移除,情景3为通过模拟该城市的未来土地利用变化顺序移除斑块 |
dI值随着物种迁移距离的增加而增大。全部三个生境变化情景中, 物种迁移距离为1km时的dI值很小, 对于全部潜在生境斑块dI值均接近零;物种迁移距离为8—10km时, dI值因斑块特征而异(图 3), 且斑块dI值越高其dI值受物种迁移距离变化的影响越大。说明物种迁移距离为1km时潜在生境斑块的移除对景观连接度几乎没有影响, 物种迁移距离为8—10km时关键的潜在生境斑块则对维持景观连接度有重要意义。
除了物种迁移距离为1km时PC值在0.16附近极小幅浮动而看不出显著变化趋势外, 三种生境变化情景下PC值的变化特征差异均较为明显(图 3)。单独移除斑块的情景1表明, 相对核心生境斑块所确定的PC值基数, 每个潜在生境斑块对PC值的贡献量很小, 均小于0.27%, 这与研究区景观格局特征有关。对比生境变化情境1和情景2下PC值的变化, 可知即使单个斑块的损失对于景观连接度影响较小, 但生境斑块的连续损失对景观连接度仍然具有较大影响, 且该影响随着连续损失斑块面积的增大而增大。连续移除斑块的情景2和情景3显示潜在生境斑块移除顺序的差异会影响PC值减小的速度。情景2中PC值随累计移除生境斑块数量增加而平缓降低, PC值最后因移除较大面积潜在斑块而迅速降低;情景3中PC值则在某些斑块被移除后有相对明显的突然降低。
将不同生境变化情景下dI值按降序排列, 结合dI值整体变化规律, 选取前10%的斑块作为对整个生境网络的景观连接度有显著贡献的生境斑块。情景1和情景2中1、68—74号斑块具有显著贡献, 情景3中1、4、59、62、68—72号斑块具有显著贡献, 这些潜在生境斑块需要重点保护(图 4), 其中59、69号斑块只在物种迁移距离为1km时有显著贡献, 1、62号斑块在物种迁移距离为8、10km时才有显著贡献。核心生境斑块对景观连接度的贡献显著高于潜在生境斑块的贡献, 但延庆区内面积占比达53.30%的核心斑块没有被划入当前生态保护红线内, 因此生态保护红线外的核心斑块区域同样需要注意保护。

|
| 图 4 三种生境变化情景下重点保护生境斑块图 Fig. 4 Map of habitat patches requiring key protection under three different habitat change scenarios 左图为情景1和情景2中重点保护斑块, 因为这两种情景下保护斑块相同;右图为情景3中重点保护斑块 |
生境斑块的dI值大小与斑块面积和其所在生境网络的位置有关, 面积大的斑块或在网络中位于关键位置的斑块均对景观连接度有较大贡献。由结果可知, 生境中面积较大的核心生境斑块虽然数量仅占5%, 但维持了生境网络98%左右的景观连接度, 表明大中型斑块对于维护景观连接度水平具有极重要的意义, 这与前人研究结果一致[20, 38]。此外, 选择三种生境变化情景下dI值最高的前10个斑块(表 4)分析它们所具有的特征, 68—74号斑块均为研究区中面积较大的潜在生境斑块, 因此其对景观连接度的贡献也相对较高。dI值排名第一位的1号斑块为研究区内面积最小的潜在生境斑块, 其他部分面积较小的斑块也具有较高的dI值, 原因可能是这些斑块与核心生境斑块的距离较近, 且处于整个生境网络相对中心的位置。这也反映了位于关键位置的小型斑块对于维护生境网络景观连接度的重要性[39]。
| 情景1 Scenarios1 | 情景2 Scenarios2 | 情景3 Scenarios3 | |||||||||||
| 编号 Number |
面积/m2 | 比例/% | dI | 编号 Number |
面积/m2 | 比例/% | dI | 编号 Number |
面积/m2 | 比例/% | dI | ||
| 1 | 203421 | 0.01 | 0.27 | 1 | 203421 | 0.01 | 0.27 | 1 | 203421 | 0.01 | 0.27 | ||
| 72 | 4609860 | 0.23 | 0.17 | 72 | 4609860 | 0.23 | 0.22 | 72 | 4609860 | 0.23 | 0.17 | ||
| 73 | 5356630 | 0.27 | 0.15 | 73 | 5356630 | 0.27 | 0.15 | 4 | 209205 | 0.01 | 0.13 | ||
| 74 | 6286520 | 0.32 | 0.14 | 74 | 6286520 | 0.32 | 0.13 | 70 | 3911620 | 0.20 | 0.11 | ||
| 70 | 3911620 | 0.20 | 0.12 | 68 | 2671000 | 0.13 | 0.11 | 71 | 4176760 | 0.21 | 0.10 | ||
| 71 | 4176760 | 0.21 | 0.11 | 70 | 3911620 | 0.20 | 0.11 | 68 | 2671000 | 0.13 | 0.10 | ||
| 68 | 2671000 | 0.13 | 0.09 | 71 | 4176760 | 0.21 | 0.10 | 62 | 846551 | 0.04 | 0.09 | ||
| 69 | 3191760 | 0.16 | 0.07 | 62 | 846551 | 0.04 | 0.09 | 69 | 3191760 | 0.16 | 0.07 | ||
| 65 | 1011520 | 0.05 | 0.06 | 65 | 1011520 | 0.05 | 0.07 | 65 | 1011520 | 0.05 | 0.06 | ||
| 52 | 595010 | 0.03 | 0.04 | 69 | 3191760 | 0.16 | 0.07 | 53 | 627059 | 0.03 | 0.05 | ||
| dI, 网络连接度变化率;表中展示物种迁移距离为10km时, 三种生境变化情景下dI值最高的前10个斑块的编号、面积、该斑块面积占全部潜在生境斑块面积的比例、dI值 | |||||||||||||
景观格局变化对不同迁移能力物种有明显不同的影响, 应针对生境中保护物种的需要制定相应的保护方案。本研究中物种迁移距离为1km时, 单独和连续移除潜在生境斑块的情景对景观连接度都几乎没有影响, 核心生境斑块的特征决定了其生境景观连接度大小。因此保护迁移能力非常弱的物种时, 应重点保护该物种集中分布的生境斑块, 尤其应避免核心斑块中位于生态保护红线外的区域发生破碎化。对比物种迁移距离为8km和10km时的PC值变化, 迁移能力更强的物种对移除相同生境斑块的响应更明显, 其对景观格局变化更敏感。迁移能力越强, 通过踏脚石进行远距离迁移的可能性越高, 则潜在生境斑块的损失对其影响越大。保护迁移能力较强的物种时, 应保护对景观连接度具有显著贡献的潜在生境斑块, 同时应注意缓解城市道路的障碍效应对物种迁移的阻碍[40]。
4.3 生境特征对生境保护的影响本研究选取的生境中以大中型的核心生境斑块为主, 小型潜在生境斑块质量较小且破碎化程度高。全部潜在生境斑块对整个网络的景观连接度有最大2.15%的影响, 单独斑块对景观连接度有最大0.28%的影响, 因而小型斑块对维护生境的景观连接度仍然具有一定贡献。因此, 本研究通过综合评价不同情境下各生境斑块的相对重要性, 确定了需要着重保护的关键斑块, 即重点保护大中型核心生境斑块和个别对景观连接度有显著贡献的小型潜在生境斑块(图 4)。在本研究区中, 大部分应重点保护的生境斑块还未归入已划定的生态红线范围内, 其中包括核心生境斑块的大部分区域, 未来城市规划者应注意避免城市建筑、道路等的建设对这些区域的侵占、切割和破碎化。
不同生境中斑块的特征可能存在较大的差异, 因此需要采取不同的保护措施。对于生境中大中型生境斑块占据面积较大、位置集中的地区, 应该重点保护大中型生境斑块, 此外还要根据斑块特征, 识别位于生境网络关键位置且易被破坏的重要小型生境斑块[41]。除了关注生态保护区内的核心生境, 还应该关注生态保护区外对于物种迁徙和景观连接度有重要作用的潜在生境斑块及生态廊道[42], 针对不同生境和物种的需要因地制宜地采取保护措施, 以达到生态保护投入和生物多样性保护效率协调统一。
4.4 城市化扩展对景观连接度的影响城市化扩展过程使得适合动植物生存的生境逐渐被建筑物、水泥路面等城市基础设施替代, 导致生境破碎化现象严重并影响到生物体迁移等生态过程[43]。对比情景2和情景3下PC值变化特征, 可知对景观连接度有显著贡献的生境斑块如因城市化扩展过程被较早移除, 景观连接度下降迅速。对比三种不同情境下dI值的变化, 发现情境1和情景2下对景观连接度贡献较大的生境斑块高度重合, 都为1、68—74号斑块;但情境3下对景观连接度贡献较大的生境斑块与前两个情景不同, 为1、4、59、62、68—72号斑块。相较于仅考虑生境斑块特征的情景, 增加考虑城市化扩展情景能更好地识别对生境网络景观连接度具有显著贡献的生境斑块, 这些斑块应给予优先保护, 如情景3识别出的重点保护生境斑块。另外, 对比《延庆分区规划(国土空间规划)(2017—2035年)》中的国土空间规划分区, 68和72号重点保护的潜在生境斑块毗邻规划的城镇建设用地区域, 上述两个生境斑块更易在未来城市扩展过程中破碎化或损失, 应着重保护。
4.5 本研究不足之处本研究选取生境斑块质心到质心的最小耗费路径长度作为PC指数计算的参数, 但对于研究区内面积较大的生境斑块, 生物从邻近生境斑块迁移至大斑块边缘的成本距离远小于迁移至斑块质心的成本距离, 使得计算出的成本距离与实际情况间存在偏差。另一方面, 本研究基于多因素赋值确定生境阻力面, 实践中应基于受保护物种的观测数据构建阻力值方案。由于受数据可获取性等限制, 本文结合文献资料设置阻力值, 存在一定主观性[44], 可以通过构建多条最小耗费路径来划定受保护物种可能的移动区域[45], 以降低经验赋值的不确定性。此外, 国土空间规划政策可以改变城市未来空间形态、改变特定区域土地利用变化的方向, 有研究表明考虑国土空间规划政策的城市空间形态模拟在中长期尺度上具有更高的现实性和精度[46]。进行未来土地利用变化模拟时, 可以增加对研究区国土空间规划的参考。这些研究不足将在以后的研究中进行改进和完善。
5 结论城市化过程导致的城市生境斑块破碎化和损失可能降低生境网络的景观连接度, 不利于物种迁移及生物多样性的保护。本研究关注生态保护区外的生境斑块, 基于图论构建生境网络, 选取景观连接度指数PC和dI, 结合未来土地利用变化模拟设计三种生境变化情景, 基于生境变化情景进行生境网络景观连接度和生境斑块重要性的评价。本研究识别出了对生境网络景观连接度具有显著贡献的各类生境斑块。景观格局变化对不同迁移能力的物种可能产生显著差异性的影响, 本研究中不同生境变化情景下生境网络景观连接度的变化特征差异明显。在实际工作中, 应同时考虑生境斑块本身特征、各类生境斑块对网络景观连接度的贡献和城市化扩展过程的压力制定适宜的生境斑块管理措施, 以最大程度地提高资源利用效率和保护生物多样性。本研究的结果可为城市生物多样性保护和城市土地利用管理和规划提供科学支撑。
| [1] |
Seto K C, Fragkias M, Guneralp B, Reilly M K. A meta-analysis of global urban land expansion. PLoS One, 2011, 6(8): e23777. DOI:10.1371/journal.pone.0023777 |
| [2] |
Grimm N B, Faeth S H, Golubiewski N E, Redman C L, Wu J G, Bai X M, Briggs J M. Global change and the ecology of cities. Science, 2008, 319(5864): 756-760. DOI:10.1126/science.1150195 |
| [3] |
Kalnay E, Cai M. Impact of urbanization and land-use change on climate. Nature, 2003, 425(6953): 528-531. |
| [4] |
Rudd H, Vala J, Schaefer V. Importance of backyard habitat in a comprehensive biodiversity conservation strategy: a connectivity analysis of urban green spaces. Restoration Ecology, 2002, 10(2): 368-375. DOI:10.1046/j.1526-100X.2002.02041.x |
| [5] |
Alberti M, Marzluff J M, Shulenberger E, Bradley G, Ryan C, Zumbrunnen C. Integrating humans into ecology: opportunities and challenges for studying urban ecosystems. BioScience, 2003, 53(12): 1169-1179. DOI:10.1641/0006-3568(2003)053[1169:IHIEOA]2.0.CO;2 |
| [6] |
McKinney M L. Urbanization, biodiversity, and conservation. BioScience, 2002, 52(10): 883-890. DOI:10.1641/0006-3568(2002)052[0883:UBAC]2.0.CO;2 |
| [7] |
DeFries R, Hansen A, Turner B L, Reid R, Liu J G. Land use change around protected areas: management to balance human needs and ecological function. Ecological Applications, 2007, 17(4): 1031-1038. DOI:10.1890/05-1111 |
| [8] |
林勇, 樊景凤, 温泉, 刘述锡, 李滨勇. 生态红线划分的理论和技术. 生态学报, 2016, 36(5): 1244-1252. |
| [9] |
饶胜, 张强, 牟雪洁. 划定生态红线创新生态系统管理. 环境经济, 2012(6): 57-60. |
| [10] |
Newmark W D. Legal and biotic boundaries of western North American national parks: a problem of congruence. Biological Conservation, 1985, 33(3): 197-208. DOI:10.1016/0006-3207(85)90013-8 |
| [11] |
Leroux S J, Kerr J T. Land development in and around protected areas at the wilderness frontier. Conservation Biology, 2013, 27(1): 166-176. DOI:10.1111/j.1523-1739.2012.01953.x |
| [12] |
Hansen A J, DeFries R. Ecological mechanisms linking protected areas to surrounding lands. Ecological Applications, 2007, 17(4): 974-988. DOI:10.1890/05-1098 |
| [13] |
Hanski I. Habitat loss, the dynamics of biodiversity, and a perspective on conservation. AMBIO, 2011, 40(3): 248-255. DOI:10.1007/s13280-011-0147-3 |
| [14] |
Taylor P D, Fahrig L, Henein K, Merriam G. Connectivity is a vital element of landscape structure. Oikos, 1993, 68(3): 571-573. DOI:10.2307/3544927 |
| [15] |
Laita A, Mönkkönen M, Kotiaho J S. Woodland key habitats evaluated as part of a functional reserve network. Biological Conservation, 2010, 143(5): 1212-1227. DOI:10.1016/j.biocon.2010.02.029 |
| [16] |
吴昌广, 周志翔, 王鹏程, 肖文发, 滕明君. 景观连接度的概念、度量及其应用. 生态学报, 2010, 30(7): 1903-1910. |
| [17] |
Bunn A G, Urban D L, Keitt T H. Landscape connectivity: a conservation application of graph theory. Journal of Environmental Management, 2000, 59(4): 265-278. DOI:10.1006/jema.2000.0373 |
| [18] |
Zetterberg A, Mörtberg U M, Balfors B. Making graph theory operational for landscape ecological assessments, planning, and design. Landscape and Urban Planning, 2010, 95(4): 181-191. DOI:10.1016/j.landurbplan.2010.01.002 |
| [19] |
Yu D Y, Xun B, Shi P J, Shao H B, Liu Y P. Ecological restoration planning based on connectivity in an urban area. Ecological Engineering, 2012, 46: 24-33. DOI:10.1016/j.ecoleng.2012.04.033 |
| [20] |
蒙吉军, 王晓东, 尤南山, 朱利凯. 黑河中游生态用地景观连接性动态变化及距离阈值. 应用生态学报, 2016, 27(6): 1715-1726. |
| [21] |
刘伊萌, 杨赛霓, 倪维, 何锦彪. 生态斑块重要性综合评价方法研究——以四川省为例. 生态学报, 2020, 40(11): 3602-3611. |
| [22] |
Bodin Ö, Saura S. Ranking individual habitat patches as connectivity providers: integrating network analysis and patch removal experiments. Ecological Modelling, 2010, 221(19): 2393-2405. DOI:10.1016/j.ecolmodel.2010.06.017 |
| [23] |
Xun B, Yu D Y, Wang X. Prioritizing habitat conservation outside protected areas in rapidly urbanizing landscapes: a patch network approach. Landscape and Urban Planning, 2017, 157: 532-541. DOI:10.1016/j.landurbplan.2016.09.013 |
| [24] |
史芳宁, 刘世梁, 安毅, 孙永秀, 董世魁, 武雪. 城市化背景下景观破碎化及连接度动态变化研究——以昆明市为例. 生态学报, 2020, 40(10): 3303-3314. |
| [25] |
Tarabon S, Calvet C, Delbar V, Dutoit T, Isselin-Nondedeu F. Integrating a landscape connectivity approach into mitigation hierarchy planning by anticipating urban dynamics. Landscape and Urban Planning, 2020, 202: 103871. DOI:10.1016/j.landurbplan.2020.103871 |
| [26] |
王文杰, 申文明, 刘晓曼, 张峰, 潘英姿, 罗海江. 基于遥感的北京市城市化发展与城市热岛效应变化关系研究. 环境科学研究, 2006, 19(2): 44-48. |
| [27] |
沈洁滢, 崔国发, 刘润泽, 曲宏, 黄三祥, 吴记贵, 范雅倩. 北京松山国家级自然保护区旅游线路周边野生动物分布规律. 北京林业大学学报, 2016, 38(7): 71-80. |
| [28] |
吉晟男, 刘雅欣, 赵志平, 陶夏秋, 肖能文, 李春旺. 北京市延庆区哺乳动物多样性、分布及影响因素. 动物学杂志, 2020, 55(1): 9-19. |
| [29] |
He J H, Huang J L, Liu D F, Wang H, Li C. Updating the habitat conservation institution by prioritizing important connectivity and resilience providers outside. Ecological Indicators, 2018, 88: 219-231. DOI:10.1016/j.ecolind.2017.12.067 |
| [30] |
Saura S, Pascual-Hortal L. A new habitat availability index to integrate connectivity in landscape conservation planning: comparison with existing indices and application to a case study. Landscape and Urban Planning, 2007, 83(2-3): 91-103. DOI:10.1016/j.landurbplan.2007.03.005 |
| [31] |
荀斌. 城市绿地生境网络连接度评价及优化研究[D]. 北京: 北京师范大学, 2013.
|
| [32] |
Sutherland G D, Harestad A S, Price K, Lertzman K P. Scaling of natal dispersal distances in terrestrial birds and mammals. Conservation Ecology, 2000, 4(1): 16. DOI:10.5751/ES-00184-040116 |
| [33] |
Avon C, Bergès L. Prioritization of habitat patches for landscape connectivity conservation differs between least-cost and resistance distances. Landscape Ecology, 2016, 31(7): 1551-1565. DOI:10.1007/s10980-015-0336-8 |
| [34] |
Lieth H, Whittaker R H. Primary Productivity of the Biosphere. Berlin, Heidelberg: Springer, 1975, 339-339. |
| [35] |
王秉义. 巢湖流域土地利用演变的生态响应与优化模拟研究[D]. 徐州: 中国矿业大学, 2018.
|
| [36] |
蔡玉梅, 刘彦随, 宇振荣, Verburg P H. 土地利用变化空间模拟的进展——CLUE-S模型及其应用. 地理科学进展, 2004, 23(4): 63-71. |
| [37] |
赵天. 基于CLUE-S模型的南昌市土地利用变化模拟研究[D]. 南昌: 江西农业大学, 2014.
|
| [38] |
古璠, 黄义雄, 陈传明, 程栋梁, 郭佳蕾. 福建省自然保护区生态网络的构建与优化. 应用生态学报, 2017, 28(3): 1013-1020. |
| [39] |
欧维新, 袁薇锦. 基于景观连接度的盐城滨海湿地丹顶鹤生境斑块重要性评价. 资源科学, 2015, 37(4): 823-831. |
| [40] |
Fu W, Liu S L, Degloria S D, Dong S K, Beazley R. Characterizing the "fragmentation-barrier" effect of road networks on landscape connectivity: a case study in Xishuangbanna, Southwest China. Landscape and Urban Planning, 2010, 95(3): 122-129. DOI:10.1016/j.landurbplan.2009.12.009 |
| [41] |
Li W B, Yang P P, Li B W, Liu C, Sun L X, Li J H. Habitat characteristics or protected area size: what is more important for the composition and diversity of mammals in nonprotected areas?. Ecology and Evolution, 2021, 11: 7250-7263. DOI:10.1002/ece3.7540 |
| [42] |
Hofmann D D, Behr D M, McNutt J W, Ozgul A, Cozzi G. Bound within boundaries: do protected areas cover movement corridors of their most mobile, protected species?. Journal of Applied Ecology, 2021, 58(6): 1133-1144. DOI:10.1111/1365-2664.13868 |
| [43] |
Saunders D A, Hobbs R J, Margules C R. Biological consequences of ecosystem fragmentation: a review. Conservation Biology, 1991, 5(1): 18-32. DOI:10.1111/j.1523-1739.1991.tb00384.x |
| [44] |
陈春娣, 贾振毅, 吴胜军, 童笑笑, 周文佐, 陈若漪, 张超林. 基于文献计量法的中国景观连接度应用研究进展. 生态学报, 2017, 37(10): 3243-3255. |
| [45] |
Rayfield B, Fortin M J, Fall A. The sensitivity of least-cost habitat graphs to relative cost surface values. Landscape Ecology, 2010, 25(4): 519-532. DOI:10.1007/s10980-009-9436-7 |
| [46] |
马世发, 艾彬, 念沛豪. 基于主体功能空间引导的城市增长形态模拟. 城市规划, 2019, 43(9): 78-85. |