 2022, Vol. 42
2022, Vol. 42文章信息
- 孙渝雯, 马赞文, 陶贞, 张乾柱, 唐文魁, 吴迪, 钟庆祥, 王振刚, 丁健
- SUN Yuwen, MA Zanwen, TAO Zhen, ZHANG Qianzhu, TANG Wenkui, WU Di, ZHONG Qingxiang, WANG Zhengang, DING Jian
- 海南岛西南部土壤生物硅分布的时空差异及其驱动机制
- Spatio-temporal variation of soil biogenic silicon distribution and its driving mechanism in the southwestern Hainan Island
- 生态学报. 2022, 42(17): 7092-7104
- Acta Ecologica Sinica. 2022, 42(17): 7092-7104
- http://dx.doi.org/10.5846/stxb202108012090
-
文章历史
- 收稿日期: 2021-08-01





2. 台山市新宁中学, 台山 529200;
3. 长江科学院重庆分院, 重庆 400026;
4. 深圳市城市规划设计研究院有限公司, 深圳 518028
2. Taishan Xinning Middle School, Taishan 529200, China;
3. Chongqing Branch, Changjiang River Scientific Research Institute, Chongqing 400026, China;
4. Urban Planning and Design Institute of Shenzhen, Shenzhen 518028, China
元素硅(Si), 尤其是溶解态硅(DSi), 在地球表层的迁移-转化影响海洋硅藻生产力[1]和陆地-海洋底部硅酸盐矿物风化[2—3], 从而被认为是全球碳循环的重要调节器, 影响地球环境变化[4]。作为地壳中第二丰富的元素, Si是植物生长的有益元素之一[5], 能够减弱许多生物和非生物因素对植物生长的限制作用。例如, 植物叶片较高的Si含量可以增强植物对食草动物和病原体的抵抗性[6—7], 提高植物抵抗磷胁迫的能力[8—9]、抗旱性[10]和对重金属的耐受性[11], 进而提高陆生植物的生长性能和生长速度, 促进陆地生态系统的碳汇功能[12]。植物体内的Si通过调控凋落物表面化学成分, 包括养分含量和营养成分化学计量特征以及碳化合物的性质影响凋落物的分解速率[13—14], 最终影响生源要素(碳、氮、磷)的生物地球化学循环。可见元素Si的生物地球化学循环对维持地球生态系统功能和产品质量至关重要[15—16]。
碳(C)、氮(N)、磷(P)元素是土壤养分的重要组成部分, 对提高土壤酶和微生物活性以及促进植物群落发育等方面有积极作用, 与生态系统能量循环、土地生产力密切相关, 引起普遍关注[17—22]。然而人们对Si和C、N、P循环的协同效应研究较少[23]。
在陆地生态系统中, Si主要以溶解态Si(DSi)、非晶质Si(主要是指植物生长过程形成的植物岩或成矿性硅结核)、隐晶质Si(例如次生石英)和晶质Si(包括原生硅酸盐矿物(云母、长石)、石英和次生硅酸盐矿物(粘土矿物))等形式存在于岩石(风化层)、土壤、植物体以及河流和湖泊中[24—27]。土壤中Si含量变化于0.5%—47%之间[28], 其中DSi主要由硅酸盐矿物风化过程慢速水解释放的单硅酸Si (H4SiO4, 原生DSi)[29]和植物生长过程形成的非晶质Si((SiO2·nH2O), Biogenic Silicon, BSi)[30]返回土壤较快溶解形成H4SiO4[31—32]两部分组成。研究发现, 随凋落物返回表层土壤的BSi的溶解度是硅酸盐矿物溶解度的2倍[33]。随着土壤发育原生DSi不断被淋失, BSi溶解对土壤DSi的贡献逐渐增加[4]并形成土壤活性硅酸盐库[31];而且DSi迁移速率也受到植物生物过程的影响[29, 31, 33], 特别在湿热地区这一现象尤其明显。例如, 刚果河流域热带雨林土壤BSi溶解对河流DSi输出的贡献率大约为75%[33], 夏威夷热带雨林约为90%[34];而温带针叶林中土壤BSi溶解对陆地生态系统DSi输出的贡献率仅为12%[35]。因此, 化学风化过程与生物过程分别对陆地生态系统DSi循环的贡献大小一直存在争议[4]。为解决这一问题, 厘清陆地生态系统岩石/矿物化学风化速率和土壤BSi含量变化尤为重要。
本研究选择化学风化过程与生物过程旺盛、岩性以花岗岩为主的海南岛西南部昌化江流域代表性植物群落, 分别于干季、湿季采集土壤样品, 测量土壤理化指标和BSi含量, 揭示热带季风区土壤BSi的时空分布特征及其主要影响因素, 为南海生态系统碳-硅循环研究提供基础数据。
1 研究区概况昌化江位于海南岛西南部(18°32′—19°21′N; 108°36′—109°44′E), 为海南岛第二大河, 发源于海南岛中部的黎母山空禾岭, 呈东北-西南-西北流向, 最终注入南海。研究区受东亚季风和南亚季风的影响, 季风气候显著。多年月平均气温变化于20.54—26.68℃, 其中1月份平均气温最低(17.70℃), 7月最高(27.93℃)。干季(11月—次年4月)、湿季(5月—10月)降水量差别显著, 湿季降水量高达1260.40 mm, 月平均降水210.07 mm;干季降水量仅248.15 mm, 月平均降水41.36 mm。多年月平均降水量125.42 mm。
昌化江流域岩石以花岗岩为主, 占流域总面积的66.84%;变质岩、碎屑岩占22.13%;灰岩夹层占7.20%, 主要分布在中游地区;其余为第四纪松散沉积物, 主要分布在河口地区。研究区地貌类型包括山地、丘陵、台地和平原四种。其中山地约占流域总面积的57.66%, 主要分布在流域上游和中游地区;台地主要分布在中游河谷两侧及下游地区, 约占流域总面积的29.37%;丘陵约占流域总面积的10.91%;平原主要分布在昌化江下游, 仅占流域总面积的2.06%。
研究区土壤分布自分水岭到干流河谷, 呈现黄壤-红壤-红褐土逐渐过渡的规律, 下游为沉积砂土。研究区植被覆盖率(大约67.7%)较高, 且不同地区植物群落组成差别较大:山地地区主要分布热带季雨林和人工经济林(包括橡胶林、桉树林、芒果林等);农作物(主要有香蕉、甘蔗和水稻等)主要分布在地势较平缓的台地和平原地区。
2 样品采集与方法 2.1 样品采集分别于2014年和2016年1月(干季)、2014年和2015年8月(湿季)选择海南岛西南部3种典型植物群落(自然林(热带季雨林)、经济林(橡胶林、桉树林、芒果林)、农作物(香蕉地和甘蔗地))6个样地采集土壤样品, 共采集土壤样品66个(采样点位置和信息见图 1、表 1)。由于地貌部位和岩石风化程度的影响, 研究区不同群落土壤厚度差异较大(表 1)。根据实地土壤厚度, 采样至风化层。按照土壤剖面特征分为表层和底层。其中, 自然林和甘蔗地0—20 cm为表层、20—40 cm为底层;橡胶林、芒果林、香蕉地0—10 cm为表层、10—20 cm为底层;桉树林土层较薄且没有明显颜色、质地变化, 未进行层次划分。

|
| 图 1 研究区植被类型和采样点位置 Fig. 1 Vegetation types and distribution of sampling sites in the study area BWL:季雨林Monsoon forest;XJL:橡胶林Rubber forest;ASL:桉树林Eucalyptus forest;MGL:芒果林Mango forest;BND:香蕉地Banana;GZD:甘蔗地Sugarcane |
| 样地名称 Sampling sites |
样地缩写 Sample name |
经纬度 Location |
植被类型 Vegetation type |
优势种 Dominant species |
土壤类型 Soil type |
土壤质地 Soil texture |
土层厚度 Deepth/cm |
母岩 Parent rock |
| 季雨林 Monsoon forest |
BWL | 19°05′03.83″N; 109°07′15.36″E |
常绿阔叶林 | 乌墨黄毛榕 | 黄壤 | 壤土 | 40 | 花岗岩 |
| 橡胶林 Rubber forest |
XJL | 19°15′31.39″N; 109°05′33.19″E |
经济林 | 橡胶 | 红褐土 | 粘壤土 | 20 | 花岗岩 |
| 桉树林 Eucalyptus forest |
DYA | 18°45′57.42″N; 109°09′18.74″E |
经济林 | 大叶桉 | 红褐土 | 砂土 | 10 | 砂岩夹变质岩 |
| 芒果林 Mango forest |
MGL | 18°58′10.27″N; 108°56′07.83″E |
经济林 | 芒果树 | 红褐土 | 砂土 | 20 | 花岗岩 |
| 香蕉地 Banana |
BND | 19°13′53.37″N; 109°11′19.01″E |
农作物 | 香蕉树 | 红壤 | 砂质粘壤土 | 20 | 花岗岩 |
| 甘蔗地 Sugarcane |
GZD | 19°11′56.32″N; 108°46′55.65″E |
农作物 | 甘蔗 | 红褐土 | 砂土 | 40 | 第四纪沉积物 |
每个样地按V字型采集3个剖面, 将样品装入PE自封袋中并排尽空气, 密封避光保存带回实验室处理、分析。
2.2 分析方法 2.2.1 土壤理化指标测量土壤pH值采用pH计(水土比1∶ 2.5)测定;土壤容重和含水率分别采用环刀法和烘干法(105℃)测定;将土壤样品去除有机质和碳酸盐后使用Mastersizer 2000激光粒度仪测定土壤粒度;经过预处理的土壤样品使用ZSX Primus X射线荧光光谱仪测定土壤样品常量元素含量(相对误差<1%);经过盐酸(HCl)(10%)预处理的土壤样品使用EL CHNS-O型元素分析仪测定土壤有机碳(SOC)(相对误差<2%)和总氮(TN)含量(相对误差<5%)。
2.2.2 土壤生物硅含量测量采用热碱消化连续提取法萃取土壤BSi并测定其含量[36]。称取5 g土壤样品置于50 mL聚丙烯离心管中, 加入过量H2O2(15%)和HCl(10%)去除有机质和碳酸盐;然后加入40 mL Na2CO3(2 mol/L), 85℃恒温水浴5 h, 提取上清液待测。用硅钼蓝比色法测定提取液中硅酸盐浓度:取20 g钼酸铵加水稀释至200 mL, 加入100 mL硫酸(20%)制成显色试剂, 选用国家海洋局第二海洋研究所生产的Si标准溶液(GSB04-1752-2004), 使用UV-3000分光光度计测量土壤BSi的含量。
2.2.3 化学计量特征值计算化学蚀变指数(Chemical Index of Alteration, CIA)[37]和A-CN-K三角图[38—39]常用于指示岩石/矿物化学风化程度。其中CIA值计算公式为:
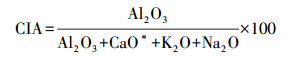
|
(1) |
式中, CaO*为硅酸盐矿物中CaO的摩尔含量。硅酸盐矿物中CaO与Na2O含量通常以1∶1的比例存在, 当CaO>Na2O时, 认为CaO*=Na2O, 反之, 则认为CaO*=CaO[40]。一般认为, 50<CIA<65, 指示寒冷干燥气候条件下低等的化学风化程度;65<CIA<85表示温暖、湿润条件下中等强度的化学风化;CIA>85表示炎热、潮湿的热带、亚热带条件下强烈的化学风化[39]。根据研究区土壤化学元素组成和公式(1)计算研究区CIA值。
C/N比值是表征土壤有机质质量和分解程度的一个重要指标[41]。土壤C/N比值通常随土地利用变化而变化[42]。
2.3 数据处理运用SPSS Statistics19.0对实验数据进行统计分析, 所有数据以(平均值±标准误差)表示。不同群落土壤Si、土壤BSi含量差异应用单因素方差分析(One-way-ANOVA)中的Duncan法进行差异显著性检验;对土壤BSi含量与土壤理化指标、化学蚀变指数(CIA)、生物量等要素进行Pearson相关分析(Pearson Correlation Coefficient), 采用主成分分析(Principal Component Analysis, PCA)识别影响研究区土壤BSi变化的主要环境因子, 显著性水平设为0.05。
文中植被净初级生产力(NPP)数据源自美国地质调查局提供的MOD17A3H数据集, 时间范围为2014年—2016年, 空间分辨率为500m。
文中图件运用Origin 2021、Excel 2019和Adobe Illustrator 2018软件绘制。
3 结果 3.1 研究区土壤理化特征研究区不同群落土壤理化性质存在差异(表 2)。研究区土壤pH值变化于4.36—5.61之间, 平均为5.11, 而且干季与湿季变化不明显。不同群落土壤容重无显著差异(P = 0.68>0.05)。土壤含水率变化于4.31%—18.02%之间, 最低值出现在甘蔗地(GZD), 最高值出现在香蕉地(BND)。根据美国制土壤粒级划分标准:粘粒(<2 μm)、粉粒(2—50 μm)和砂粒(>50 μm), 研究区土壤粘粒、粉粒和砂粒各粒级组分平均含量分别为10.06%、22.24%、67.73%;其中, 橡胶林(XJL)土壤粘粒含量最高(26.82%), 芒果林(MGL)土壤砂粒含量最高(89.87%)。季雨林(BWL)、桉树林(DYA)、芒果林(MGL)、甘蔗地(GZD)砂粒含量均超过60%, 橡胶林(XJL)、香蕉地(BND)粘粒和粉粒含量均超过50%。
| 样地名称 Sampling sites |
pH | 容重 Volumetric weight/(g/cm3) |
含水率/% Moisture content |
粒度Granularity/% | ||
| 粘粒 Clay |
粉粒 Silt |
砂粒 Sand |
||||
| BWL | 4.36±0.38 | 1.33±0.12 | 16.98±4.04 | 3.26±1.89 | 25.71±11.24 | 71.03±13.05 |
| XJL | 4.84±0.26 | 1.65±0.12 | 16.46±2.54 | 26.82±6.74 | 41.83±12.34 | 31.36±17.14 |
| DYA | 5.18±0.09 | 1.75±0.12 | 11.51±0.20 | 5.28±1.97 | 6.89±2.61 | 87.83±4.58 |
| MGL | 5.61±0.34 | 1.57±0.05 | 14.31±2.10 | 4.58±2.04 | 5.55±2.96 | 89.87±4.97 |
| BND | 5.28±0.75 | 1.48±0.10 | 18.02±1.62 | 18.13±12.16 | 29.60±16.36 | 52.27±24.9 |
| GZD | 5.41±0.21 | 1.63±0.14 | 4.31±0.81 | 2.35±0.29 | 23.88±8.09 | 74.04±8.09 |
研究区表土CIA值介于79.61—88.25之间, 平均值为84.14, 远高于上陆壳(Upper Continental Crust, UCC)的平均值47.92[38], 揭示研究区硅酸盐岩化学风化处于较强的中等强度化学风化程度。其中, DYA样地风化程度最高(88.25), XJL样地次之(84.49), MGL样地风化程度最弱(79.61)。
A-CN-K三角图可用来分析土壤矿物化学风化过程中元素迁移趋势[38, 43]。研究区土壤数据点基本平行于A-K线, 趋近顶点A, 且基本分布在代表典型大陆风化趋势的UCC-PAAS连线的左上侧(图 2)。揭示研究区土壤中斜长石等硅酸盐矿物几乎风化殆尽, 云母、伊利石和钾长石等含K矿物经历了更强的风化;次生粘土矿物以高岭石为主, 正处于脱硅富铝化的较高阶段[38, 44]。

|
| 图 2 A-CN-K三角图 Fig. 2 A-CN-K triangular diagram |
研究区土壤全Si含量(TSi, 以SiO2表示)变化于51.92%—84.60%之间, 平均为68.81%, 略高于上陆壳的相应值(66.00%)[38], 这与研究区岩石组成以硅酸盐岩和砂岩为主有关。研究区不同群落土壤TSi含量和BSi含量存在差异(图 3), 其中GZD群落土壤(母质为第四纪松散沉积物)TSi含量最大(84.60%±6.13%), BWL群落土壤(母质为花岗岩风化壳)最小(51.92%±2.16%)。不同群落表层(0—10 cm)土壤平均BSi含量从高到低依次为BND((2.37±0.71)mg/g)>BWL((1.86±1.34)mg/g)>XJL((1.42±0.81)mg/g)>DYA((1.22±0.28)mg/g)>MGL((0.98±0.71)mg/g)>GZD((0.62±0.74)mg/g)。

|
| 图 3 研究区不同群落土壤总硅和生物硅含量 Fig. 3 Soil TSi and BSi content in different communities in the study area 图中大/小写字母代表不同群落之间BSi/Si平均值差异的显著性水平(Duncan检验, P<0.05), 相同字母表示不同群落之间没有显著差异 |
研究区土壤BSi含量存在季节变化(图 4), 其中MGL、XJL、GZD土壤BSi含量季节变化显著(P<0.05)。研究区干季土壤BSi含量变化于0.13—3.85 mg/g之间, 平均为1.53 mg/g;湿季土壤BSi含量变化于0.34—3.56 mg/g之间, 平均为1.28 mg/g。研究区尺度上, 干季土壤BSi含量稍大于湿季的相应值;群落尺度上, 森林群落土壤BSi含量呈现干季大于湿季, 草本群落(BND和GZD)土壤BSi含量则出现相反的季节变化:湿季大于干季(图 4)。可见, 研究区土壤BSi含量季节变化主要受森林群落季节变化的影响。

|
| 图 4 研究区不同群落土壤生物硅含量季节变化 Fig. 4 Seasonal variation of soil BSi content in different communities |
研究区土壤BSi含量随土壤深度增加而减少(图 5), 这一现象一致于“土壤养分的表层聚集效应”[45—46]。

|
| 图 5 不同群落土壤生物硅含量的时空变化 Fig. 5 Spatial and temporal variation of soil BSi content in different communities |
土壤是陆地生态系统重要的碳库和营养源, 其碳储量是大气碳库的3.3倍、生物碳库的4.5倍[47]。土壤中C、N、P、Si等元素是植物生长的养分基础, 其含量及化学计量特征是鉴别土壤质量的重要指标, 并能在一定程度上揭示土壤养分的转化过程和限制性元素[18, 48]。由于成土过程和人类活动的地域差异, 使得土壤C、N、P、Si含量及其化学计量特征存在时空差异。研究区土壤有机碳(SOC)含量干季变化于4.1—11.13 mg/g之间, 平均值为6.90 mg/g, 最大值出现在XJL样地, 最小值出现在GZD样地;湿季SOC含量变化于3.43—13.77 mg/g之间, 平均值为6.69 mg/g, 最大值出现在MGL样地, 最小值出现在BND样地。土壤总氮(TN)干季变化于0.3—1.3 mg/g之间, 最大值出现在XJL样地, 最小值出现在GZD样地;湿季变化于0.27—1.13 mg/g之间, 平均值为0.62 mg/g, 最大值出现在DYA样地, 最小值出现在GZD样地。研究区不同群落土壤总磷(TP, 以P2O5计)变化于0.21—0.62 mg/g之间, 平均值为0.34 mg/g, 最大值出现在GZD样地, 最小值出现在DYA样地。
土壤C: N: P受区域水热条件和成土过程的影响, 空间异质性较大[18]。研究区不同群落土壤C/N干季变化于7.33—14.50之间, 平均值为10.45;湿季变化于7.05—14.09之间, 平均值为11.16, 干、湿季最大值均出现在GZD样地, 最小值均出现在XJL样地。研究区土壤C/N(平均值为11.03)接近中国土壤生态计量平均水平(11.9)[49]。土壤C/P变化于6.35—41.15之间, 平均值为22.03;土壤N/P变化于0.48—4.61之间, 平均值为2.32。不管是C/P还是N/P, 其中最高值均出现在XJL样地, 最低值均出现在BND样地。研究区不同群落土壤C/P、N/P均低于中国土壤生态计量平均水平(61、5.2)[49]。研究区土壤属于N限制型。为保证海南岛香蕉产量和质量, 香蕉地应追施氮肥。
4.2 土壤BSi含量变化的主要影响因素高等植物生长时其根系吸收土壤溶液中DSi, 并以无定形Si(SiO2·nH2O)的形式驻留在植物组织(尤其是叶片)中, 形成植物硅酸体(Phyolith, BSi), 然后又随凋落物返还给土壤表层, 经土壤微生物分解释放出BSi[26—27]。可见, 土壤BSi含量变化受制于其输入和输出过程及其在土壤中的滞留与保存效率。不同植物体内BSi含量差异影响土壤BSi含量, 例如相比于其他湿地植物, 芦苇净初级生产力和BSi含量均较高, 芦苇群落土壤BSi积累较多[23]。BSi在土壤中的滞留与保存受土壤DSi含量、粘土矿物吸附影响;输出包括BSi溶解形成DSi被植物吸收、随地表径流和地下水迁移以及埋藏、新生矿物吸附等[24]。
研究区表层土壤BSi含量与环境因子之间的关系不尽一致(图 6)。研究区表层土壤BSi含量与CIA值、全硅(TSi)含量、容重(ρ)、C/N呈极显著/显著负相关关系, 与土壤粘粒、SOC、TN呈极显著/显著正相关关系, 与NPP有正相关趋势(P=0.122, 图 6)。揭示研究区土壤BSi含量受研究区化学风化程度、TSi含量、土壤容重、粘粒含量、SOC含量、总氮、C/N、NPP等环境因子影响, 即研究区土壤BSi含量变化受非生物因子和生物因子的耦合控制。本研究结果一致于其他人对土壤DSi变化机制的研究结果[50]。

|
| 图 6 研究区土壤生物硅含量与群落环境要素的关系 Fig. 6 Relationships between BSi content and environmental factors |
在土壤发育初级阶段, Si的生物地球化学循环以原生矿物风化释放的DSi(原生DSi)迁移、转化为主[51], 此时BSi在土壤中得以积累保存;随着化学风化程度的增加、土壤脱硅富铝化程度加强, 土壤中原生DSi由于逐渐淋失和被植物吸收而减少, 土壤中部分BSi溶解并参与到Si的循环过程[4], 只有较稳定的BSi能够长期保存在土壤中, 土壤BSi含量趋于稳定[52]。研究区土壤BSi含量与CIA值和TSi含量呈极显著负相关关系(P<0.01), 揭示研究区高温多雨的自然环境利于土壤原生矿物风化且原生DSi淋失。为满足植物生长对DSi的需要, 部分土壤BSi溶解转换为DSi、淋失或者被植物吸收再转化为BSi。土壤BSi在土壤Si循环过程中的作用逐渐加大。研究表明, 热带雨林活植物体吸收的DSi中92.6%来自土壤BSi分解[33]。另外, 土壤BSi含量与TSi含量呈极显著负相关关系(P<0.01), 说明研究区土壤颗粒Si以物理风化源Si为主, 这与研究区土壤质地(砂粒占67.73%)一致。
研究区表层土壤BSi含量与容重呈显著负相关关系(P<0.05), 一致于河口湿地土壤有效硅与土壤容重呈负相关关系的结论[53]。由于容重较大的土壤易出现板结, 限制植物根系生长以及根系密度[54], 地下生物量较小;同时土壤板结降低土壤孔隙度和水分传导速率[55], 不利于地上生物量的形成进而影响土壤BSi的形成和积累。研究区土壤BSi含量与粘粒显著正相关(P<0.05), 是由于土壤吸附元素的潜力很大程度上取决于矿物表面的吸附能力[56], 细颗粒(粘粒和粉粒)具有较大比表面积和表面吸附效率[57], 因此粘粒含量较高的香蕉地土壤具有较高的BSi含量。
SOC、TN是土壤有机质含量和生物活性的重要指标[41]。研究区不同群落土壤BSi含量与SOC、TN呈极显著/显著正相关关系(P<0.01, P<0.05)、与NPP有正相关趋势, 表明研究区土壤BSi含量受制于群落生物生产力及其活动强度的影响。
土壤氮有助于微生物的形成、活动, N含量越高土壤有机质分解速率越快[58]。低的C/N值有利于微生物分解、提高SOC矿化速率[59], 向土壤释放更多的BSi。研究区土壤BSi含量与C/N呈极显著负相关关系(P<0.01), 揭示研究区土壤BSi含量变化受土壤微生物活动控制。
研究区土壤BSi含量变化几乎不受土壤pH值影响(图 6)。由于土壤中的单硅酸可吸附在OH基团上成为吸附态硅酸。因此, 土壤酸碱度影响粘粒对硅酸的吸附[60]。当土壤pH值为6—8时, 土壤粘粒对硅酸的吸附作用随着pH值增加而增强[61];土壤pH>8时, pH对BSi溶解度的影响显著;土壤pH<8时, pH对BSi溶解度的影响较小[62]。研究区不同群落土壤pH值变化于4.36—5.61之间, 研究区土壤pH对BSi含量变化影响较小。
为了识别海南岛西南部典型群落土壤BSi含量的主要影响因子, 对影响土壤BSi含量的环境因素(容重、含水率、SOC、TN、C/N、CIA)进行主成分分析(抽取特征值大于1的成分, 表 3)。结果表明:影响自然林土壤BSi的主要因素是SOC、C/N和CIA;影响经济林土壤BSi的主要因素是TN、CIA;影响农作物土壤BSi的主要因素是含水率、C/N。
| 群落类型 Coenotype |
成份 Components |
初始特征值Initial eigenvalue | 环境因子 Varibles |
主成份Principal components | |||
| 特征值 Eigenvalue |
贡献率 Contribution rate/% |
累积% Accumulative contribution/% |
Z 1 | Z 2 | |||
| 热带季雨林 | 容重 | 3.618 | 60.300 | 60.300 | 容重 | -0.424 | 0.718 |
| Tropical monsoon forest | 含水率 | 1.530 | 25.503 | 85.804 | 含水率 | 0.750 | 0.602 |
| SOC | 0.704 | 11.728 | 97.531 | SOC | 0.974 | -0.082 | |
| TN | 0.139 | 2.312 | 99.843 | TN | 0.964 | -0.226 | |
| C/N | 0.009 | 0.157 | 100.000 | C/N | 0.974 | -0.020 | |
| CIA | 0.000 | 0.000 | 100.000 | CIA | 0.220 | 0.770 | |
| 经济林 | 容重 | 2.719 | 45.320 | 45.320 | 容重 | -0.612 | 0.645 |
| Economic forest | 含水率 | 1.646 | 27.438 | 72.758 | 含水率 | 0.815 | -0.265 |
| SOC | 0.873 | 14.553 | 87.311 | SOC | 0.725 | 0.555 | |
| TN | 0.438 | 7.297 | 94.608 | TN | 0.853 | 0.502 | |
| C/N | 0.316 | 5.263 | 99.871 | C/N | -0.555 | -0.083 | |
| CIA | 0.008 | 0.129 | 100.000 | CIA | -0.346 | 0.771 | |
| 农作物 | 容重 | 4.029 | 67.153 | 67.153 | 容重 | -0.679 | -0.414 |
| Crops | 含水率 | 1.276 | 21.272 | 88.425 | 含水率 | 0.979 | -0.071 |
| SOC | 0.491 | 8.188 | 96.613 | SOC | 0.809 | 0.510 | |
| TN | 0.170 | 2.840 | 99.453 | TN | 0.937 | 0.293 | |
| C/N | 0.032 | 0.533 | 99.986 | C/N | -0.725 | 0.638 | |
| CIA | 0.001 | 0.014 | 100.000 | CIA | 0.742 | -0.588 | |
| SOC:土壤有机碳Soil organic carbon;TN:总氮Total nitrogen;C/N: 碳氮比C/N ratio;CIA:化学蚀变指数Chemical index of alteration | |||||||
季风气候区水热条件的季节变化影响植物生长过程和微生物分解活动[63], 致使土壤BSi含量存在季节变化。高温多雨的湿季, 一方面森林群落大多数植物处于生长旺盛期, 以吸收土壤养分形成有机质为主, 凋落物较少, 返还土壤的BSi较少;另一方面湿热环境促使土壤BSi溶解, 转换为DSi重新被植物吸收或者淋失;再者湿季海南岛降雨强度较大, 冲刷力强, 土壤BSi随雨水、径流淋失。温暖的干季, 主要受水分条件的限制, 植物生长减缓, 吸收土壤DSi的量减少;另一方面植物叶片凋落较多, 导致土壤BSi含量增多。然而农作物(香蕉和甘蔗)土壤BSi含量出现反相变化趋势, 这与香蕉、甘蔗的生态习性有关。8—9月(湿季)是香蕉和甘蔗的成熟期, 凋落物较干季(生长期)增多, 随凋落物返回土壤的BSi量亦增加;同时成熟期植物对土壤Si的吸收减少, BSi得以较大程度地保存在土壤中。除此之外, 香蕉和甘蔗属于草本植物, 叶片中所含BSi较多、木质素等难分解有机质较少[64], 凋落物进入土壤后分解较快并释放较多BSi进入土壤。因此, 研究区农业土壤BSi含量的季节变化与森林群落相反。
比较发现, 研究区土壤BSi含量(1.43 mg/g)低于热带雨林土壤BSi的平均含量(2.5 mg/g)[33], 亦低于我国陆地生态系统土壤BSi平均含量(23.60mg/g)[65]。这是由于土壤BSi含量变化除了受群落类型及其生物多样性影响[66]外, 还受研究区水文气象条件制约。研究区初级生产力显著低于热带雨林;实地观察发现XJL、DYA、MGL等经济林下其他群落不发育, 返还土壤的BSi较少;加之研究区全年高温多雨径流侵蚀较强, 土壤微生物活动活跃, 有利于BSi溶解、被吸收和淋失。
全球尺度上, 北半球土壤BSi含量存在纬度地带性差异(图 7):中高纬度地区土壤BSi含量平均值(5.72 mg/g[57, 67—71]) 显著高于低纬度地区的相应值(1.96 mg/g[33])。这是由于中高纬度地区土壤温度较低, 有机质分解速率缓慢;加之高纬度地区全年以固态降水为主, 地表较平缓, 人类活动较少, 地表侵蚀作用较弱, 有利于BSi在土壤中的积累[28]。低纬度地区, 尤其是研究区水热同期的季风气候有利于表层土壤微生物代谢活动, 致使凋落物快速分解、释放BSi进入土壤并溶解;其次, 低纬度地区全年降水较多且降雨强度较大, 地表径流机械侵蚀较强, 不利于BSi在土壤中积累。研究区地表径流机械侵蚀速率达82 m/Ma[72], 使得较多的土壤BSi发生迁移、转换, 最终以DSi的形式随地表径流向南海输入6.39×108 mol Si/a(包括原生DSi)[73], 在一定程度上保持南海生态系统的营养成分结构, 确保南海生态系统良性循环。

(1) 海南岛西南部不同植物群落土壤BSi含量从大到小依次为:香蕉地((2.38±0.72) mg/g)>热带季雨林((1.86±1.34)mg/g)>橡胶林((1.42±0.81)mg/g)>桉树林((1.22±0.28)mg/g)>芒果林((0.98±0.71)mg/g)>甘蔗地((0.62±0.74)mg/g), 研究区土壤BSi含量存在随群落变化的季节变化:森林群落土壤BSi含量干季大于湿季, 农业草本群落(香蕉地和甘蔗地)土壤BSi含量则出现湿季大于干季。研究区土壤BSi含量季节变化主要受森林群落季节变化制约。
(2) 主成分分析表明研究区土壤BSi含量变化主要受化学风化程度、有机碳、总氮和C/N等因素影响, 揭示研究区土壤BSi含量变化的生物和非生物耦合驱动机制。
(3) 比较发现, 海南岛西南部土壤BSi的平均含量为1.43 mg/g, 低于热带雨林土壤BSi的平均含量(2.5 mg/g)。这是由于研究区水热同期的季风气候有利于表层土壤微生物代谢活动, 致使凋落物快速分解、释放BSi进入土壤并溶解;加之研究区山地较多, 地表侵蚀作用较强, 使得较多的土壤BSi发生迁移-转换, 最终以DSi的形式随地表径流注入南海, 在一定程度上保持南海生态系统的营养成分结构, 确保南海生态系统良性循环。
致谢: 感谢郑伟和周梅参加野外采样工作。| [1] |
Tréguer P J, Sutton J N, Brzezinski M, Charette M A, Devries T, Dutkiewicz S, Ehlert C, Hawkings J, Leynaert A, Liu S M, Monferrer N L, López-Acosta M, Maldonado M, Rahman S, Ran L H, Rouxel O. Reviews and syntheses: the biogeochemical cycle of silicon in the modern ocean. Biogeosciences, 2021, 18(4): 1269-1289. DOI:10.5194/bg-18-1269-2021 |
| [2] |
Conley D J, Carey J C. Silica cycling over geologic time. Nature Geoscience, 2015, 8(6): 431-432. DOI:10.1038/ngeo2454 |
| [3] |
Krissansen-Totton J, Catling D C. A coupled carbon-silicon cycle model over Earth history: reverse weathering as a possible explanation of a warm mid-Proterozoic climate. Earth and Planetary Science Letters, 2020, 537: 116181. DOI:10.1016/j.epsl.2020.116181 |
| [4] |
De Tombeur F, Turner B L, Laliberté E, Lambers H, Mahy G, Faucon M P, Zemunik G, Cornelis J T. Plants sustain the terrestrial silicon cycle during ecosystem retrogression. Science, 2020, 369(6508): 1245-1248. DOI:10.1126/science.abc0393 |
| [5] |
Katz O. Silicon content is a plant functional trait: implications in a changing world. Flora, 2019, 254: 88-94. DOI:10.1016/j.flora.2018.08.007 |
| [6] |
Quigley K M, Griffith D M, Donati G L, Anderson T M. Soil nutrients and precipitation are major drivers of global patterns of grass leaf silicification. Ecology, 2020, 101(6): e03006. |
| [7] |
Hartley S E, DeGabriel J L. The ecology of herbivore-induced silicon defences in grasses. Functional Ecology, 2016, 30(8): 1311-1322. DOI:10.1111/1365-2435.12706 |
| [8] |
Kostic L, Nikolic N, Bosnic D, Samardzic J, Nikolic M. Silicon increases phosphorus (P) uptake by wheat under low P acid soil conditions. Plant and Soil, 2017, 419(1): 447-455. |
| [9] |
Cooke J, Leishman M R. Consistent alleviation of abiotic stress with silicon addition: a meta-analysis. Functional Ecology, 2016, 30(8): 1340-1357. DOI:10.1111/1365-2435.12713 |
| [10] |
Ahmed M, Qadeer U, Ahmed Z I, Hassan F U. Improvement of wheat (Triticum aestivum) drought tolerance by seed priming with silicon. Archives of Agronomy and Soil Science, 2016, 62(3): 299-315. DOI:10.1080/03650340.2015.1048235 |
| [11] |
Imtiaz M, Rizwan M S, Mushtaq M A, Ashraf M, Shahzad S M, Yousaf B, Saeed D A, Rizwan M, Nawaz M A, Mehmood S, Tu S X. Silicon occurrence, uptake, transport and mechanisms of heavy metals, minerals and salinity enhanced tolerance in plants with future prospects: a review. Journal of Environmental Management, 2016, 183: 521-529. DOI:10.1016/j.jenvman.2016.09.009 |
| [12] |
Cooke J, Leishman M R. Silicon concentration and leaf longevity: is silicon a player in the leaf dry mass spectrum?. Functional Ecology, 2011, 25(6): 1181-1188. DOI:10.1111/j.1365-2435.2011.01880.x |
| [13] |
Schaller J, Struyf E. Silicon controls microbial decay and nutrient release of grass litter during aquatic decomposition. Hydrobiologia, 2013, 709(1): 201-212. DOI:10.1007/s10750-013-1449-1 |
| [14] |
Marxen A, Klotzbücher T, Jahn R, Kaiser K, Nguyen V S, Schmidt A, Schädler M, Vetterlein D. Interaction between silicon cycling and straw decomposition in a silicon deficient rice production system. Plant and Soil, 2016, 398(1): 153-163. |
| [15] |
Conley D J. Terrestrial ecosystems and the global biogeochemical silica cycle. Global Biogeochemical Cycles, 2002, 16(4): 1121. |
| [16] |
Song Z L, Liu C Q, Müller K, Yang X M, Wu Y T, Wang H L. Silicon regulation of soil organic carbon stabilization and its potential to mitigate climate change. Earth-Science Reviews, 2018, 185: 463-475. DOI:10.1016/j.earscirev.2018.06.020 |
| [17] |
Heuck C, Weig A, Spohn M. Soil microbial biomass C: N: P stoichiometry and microbial use of organic phosphorus. Soil Biology and Biochemistry, 2015, 85: 119-129. DOI:10.1016/j.soilbio.2015.02.029 |
| [18] |
王绍强, 于贵瑞. 生态系统碳氮磷元素的生态化学计量学特征. 生态学报, 2008, 28(8): 3937-3947. DOI:10.3321/j.issn:1000-0933.2008.08.054 |
| [19] |
Fan H B, Wu J P, Liu W F, Yuan Y H, Hu L, Cai Q K. Linkages of plant and soil C: N: P stoichiometry and their relationships to forest growth in subtropical plantations. Plant and Soil, 2015, 392(1): 127-138. |
| [20] |
Han W X, Fang J Y, Guo D L, Zhang Y. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China. New Phytologist, 2005, 168(2): 377-385. DOI:10.1111/j.1469-8137.2005.01530.x |
| [21] |
李红林, 贡璐, 朱美玲, 刘曾媛, 解丽娜, 洪毅. 塔里木盆地北缘绿洲土壤化学计量特征. 土壤学报, 2015, 52(6): 1345-1355. |
| [22] |
Zechmeister-Boltenstern S, Keiblinger K M, Mooshammer M, Peñuelas J, Richter A, Sardans J, Wanek W. The application of ecological stoichiometry to plant-microbial-soil organic matter transformations. Ecological Monographs, 2015, 85(2): 133-155. DOI:10.1890/14-0777.1 |
| [23] |
Xia S P, Song Z L, Van Zwieten L, Guo L D, Yu C X, Hartley I P, Wang H L. Silicon accumulation controls carbon cycle in wetlands through modifying nutrients stoichiometry and lignin synthesis of Phragmites australis. Environmental and Experimental Botany, 2020, 175: 104058. DOI:10.1016/j.envexpbot.2020.104058 |
| [24] |
陶贞, 张超, 高全洲, 李元. 陆地硅的生物地球化学循环研究进展. 地球科学进展, 2012, 27(7): 725-732. |
| [25] |
Meunier J D, Colin F, Alarcon C. Biogenic silica storage in soils. Geology, 1999, 27(9): 835-838. DOI:10.1130/0091-7613(1999)027<0835:BSSIS>2.3.CO;2 |
| [26] |
Clarke J. The occurrence and significance of biogenic opal in the regolith. Earth-Science Reviews, 2003, 60(3/4): 175-194. |
| [27] |
Sommer M, Kaczorek D, Kuzyakov Y, Breuer J. Silicon pools and fluxes in soils and landscapes—a review. Journal of Plant Nutrition and Soil Science, 2006, 169(4): 582-582. DOI:10.1002/jpln.200690016 |
| [28] |
McKeague J A, Cline M G. Silica in soils. Advances in Agronomy, 1963, 15: 339-396. |
| [29] |
Cornelis J T, Delvaux B. Soil processes drive the biological silicon feedback loop. Functional Ecology, 2016, 30(8): 1298-1310. DOI:10.1111/1365-2435.12704 |
| [30] |
Puppe D, Höhn A, Kaczorek D, Wanner M, Wehrhan M, Sommer M. How big is the influence of biogenic silicon pools on short-term changes in water-soluble silicon in soils? Implications from a study of a 10-year-old soil-plant system. Biogeosciences, 2017, 14(22): 5239-5252. DOI:10.5194/bg-14-5239-2017 |
| [31] |
Fraysse F, Pokrovsky O S, Schott J, Meunier J D. Surface chemistry and reactivity of plant phytoliths in aqueous solutions. Chemical Geology, 2009, 258(3/4): 197-206. |
| [32] |
Li Z C, Song Z L, Singh B P, Wang H L. The impact of crop residue biochars on silicon and nutrient cycles in croplands. Science of the Total Environment, 2019, 659: 673-680. DOI:10.1016/j.scitotenv.2018.12.381 |
| [33] |
Alexandre A, Meunier J D, Colin F, Koud J M. Plant impact on the biogeochemical cycle of silicon and related weathering processes. Geochimica et Cosmochimica Acta, 1997, 61(3): 677-682. DOI:10.1016/S0016-7037(97)00001-X |
| [34] |
Derry L A, Kurtz A C, Ziegler K, Chadwick O A. Biological control of terrestrial silica cycling and export fluxes to watersheds. Nature, 2005, 433(7027): 728-731. DOI:10.1038/nature03299 |
| [35] |
Bartoli F. The biogeochemical cycle of silicon in two temperate forest ecosystems. Environmental Biogeochemistry, 1983, 35: 469-476. |
| [36] |
DeMaster D J. The supply and accumulation of silica in the marine environment. Geochimica et Cosmochimica Acta, 1981, 45(10): 1715-1732. DOI:10.1016/0016-7037(81)90006-5 |
| [37] |
邵菁清, 杨守业. 化学蚀变指数(CIA)反映长江流域的硅酸盐岩化学风化与季风气候?. 科学通报, 2012, 57(11): 933-942. |
| [38] |
陈俊, 王鹤年. 地球化学. 北京: 科学出版社, 2004: 35-63.
|
| [39] |
Nesbitt H W, Young G M. Early Proterozoic climates and plate motions inferred from major element chemistry of lutites. Nature, 1982, 299(5885): 715-717. DOI:10.1038/299715a0 |
| [40] |
McLennan S M. Weathering and global denudation. The Journal of Geology, 1993, 101(2): 295-303. DOI:10.1086/648222 |
| [41] |
Batjes N H. Total carbon and nitrogen in the soils of the world. European Journal of Soil Science, 1996, 47(2): 151-163. DOI:10.1111/j.1365-2389.1996.tb01386.x |
| [42] |
Wang W, Sardans J, Zeng C, Zhong C, Li Y, Peñuelas J. Responses of soil nutrient concentrations and stoichiometry to different human land uses in a subtropical tidal wetland. Geoderma, 2014, 232-234: 459-470. DOI:10.1016/j.geoderma.2014.06.004 |
| [43] |
Nesbitt H W, Young G M. Prediction of some weathering trends of plutonic and volcanic rocks based on thermodynamic and kinetic considerations. Geochimica et Cosmochimica Acta, 1984, 48(7): 1523-1534. DOI:10.1016/0016-7037(84)90408-3 |
| [44] |
李建武, 张甘霖, 李德成, 张鑫慧. 强烈风化条件下玄武岩发育土壤的元素地球化学特征. 地球与环境, 2012, 40(4): 491-498. |
| [45] |
Yang Y, Liu B R, An S S. Ecological stoichiometry in leaves, roots, litters and soil among different plant communities in a desertified region of Northern China. CATENA, 2018, 166: 328-338. DOI:10.1016/j.catena.2018.04.018 |
| [46] |
吴鹏, 崔迎春, 赵文君, 舒德远, 侯贻菊, 丁访军, 杨文斌. 喀斯特森林植被自然恢复过程中土壤化学计量特征. 北京林业大学学报, 2019, 41(3): 80-92. |
| [47] |
Lal R. Soil carbon sequestration impacts on global climate change and food security. Science, 2004, 304(5677): 1623-1627. DOI:10.1126/science.1097396 |
| [48] |
黄磊, 张永娥, 邵芳丽, 余新晓. 冀北山地天然次生林土壤生态化学计量特征及影响因素. 生态学报, 2021, 41(15): 6267-6279. |
| [49] |
Tian H Q, Chen G S, Zhang C, Melillo J M, Hall C A S. Pattern and variation of C: N: P ratios in China's soils: a synthesis of observational data. Biogeochemistry, 2010, 98(1): 139-151. |
| [50] |
Yang X M, Song Z L, Van Zwieten L, Sun X L, Yu C X, Wang W Q, Liu C Q, Wang H L. Spatial distribution of plant-available silicon and its controlling factors in paddy fields of China. Geoderma, 2021, 401: 115215. DOI:10.1016/j.geoderma.2021.115215 |
| [51] |
何跃, 张甘霖. 热带地区玄武岩发育土壤中的生物硅及其发生学意义. 土壤学报, 2010, 47(3): 385-392. |
| [52] |
张瑾, 李辉信, 李德成, 张甘霖. 雷琼地区玄武岩发育时间序列土壤中植硅体特征及其发生学意义. 土壤学报, 2011, 48(3): 453-460. |
| [53] |
高会, 翟水晶, 孙志高, 何涛, 田莉萍, 胡星云. 互花米草入侵影响下闽江河口湿地土壤有效硅的时空变化特征. 生态学报, 2018, 38(17): 6136-6142. |
| [54] |
才晓玲, 李志洪. 土壤容重和施肥条件对玉米根系生长的影响. 云南农业大学学报, 2009, 24(3): 470-473. DOI:10.3969/j.issn.1004-390X.2009.03.029 |
| [55] |
Singh J, Salaria A, Kaul A. Impact of soil compaction on soil physical properties and root growth: a review. International Journal of Food, Agriculture and Veterinary Sciences, 2015, 5(1): 23-32. |
| [56] |
Torn M S, Trumbore S E, Chadwick O A, Vitousek P M, Hendricks D M. Mineral control of soil organic carbon storage and turnover. Nature, 1997, 389(6647): 170-173. DOI:10.1038/38260 |
| [57] |
Spielvogel S, Prietzel J, Kögel-Knabner I. Soil organic matter stabilization in acidic forest soils is preferential and soil type-specific. European Journal of Soil Science, 2008, 59(4): 674-692. DOI:10.1111/j.1365-2389.2008.01030.x |
| [58] |
Xia S P, Song Z L, Li Q, Guo L D, Yu C X, Singh B P, Fu X L, Chen C M, Wang Y D, Wang H L. Distribution, sources, and decomposition of soil organic matter along a salinity gradient in estuarine wetlands characterized by C: N ratio, δ13C-δ15N, and lignin biomarker. Global Change Biology, 2021, 27(2): 417-434. DOI:10.1111/gcb.15403 |
| [59] |
Xia S P, Wang W Q, Song Z L, Kuzyakov Y, Guo L D, Van Zwieten L, Li Q, Hartley I P, Yang Y H, Wang Y D, Quine T A, Liu C Q, Wang H L. Spartina alterniflora invasion controls organic carbon stocks in coastal marsh and mangrove soils across tropics and subtropics. Global Change Biology, 2021, 27(8): 1627-1644. DOI:10.1111/gcb.15516 |
| [60] |
邱思婷, 米慧珊, 高会, 翟水晶. 闽江河口湿地不同植被带土壤全硅的含量及分布特征. 生态学报, 2020, 40(22): 8306-8314. |
| [61] |
李家书, 谢振翅, 胡定金, 吕忠贵, 王富华, 杨维举. 湖北省土壤有效硅含量分布. 热带亚热带土壤科学, 1997, 6(3): 176-181. |
| [62] |
Van Cappellen P. Biomineralization and global biogeochemical cycles. Reviews in Mineralogy and Geochemistry, 2003, 54(1): 357-381. DOI:10.2113/0540357 |
| [63] |
王惠, 马振民, 代力民. 森林生态系统硅素循环研究进展. 生态学报, 2007, 27(7): 3010-3017. DOI:10.3321/j.issn:1000-0933.2007.07.043 |
| [64] |
胡木兰, 左丽, 介冬梅, 高桂在, 孟笑笑, 张涛, 陈念康, 巴合依拉·哈连, 胡含霖. 东北地区草本植物和木本植物植硅体的形态特征鉴别分析. 微体古生物学报, 2018, 35(2): 122-139. |
| [65] |
刘丽君, 黄张婷, 孟赐福, 姜培坤. 中国不同生态系统土壤硅的研究进展. 土壤学报, 2021, 58(1): 31-41. |
| [66] |
Struyf E, Conley D J. Silica: an essential nutrient in wetland biogeochemistry. Frontiers in Ecology and the Environment, 2009, 7(2): 88-94. DOI:10.1890/070126 |
| [67] |
刘森, 冉祥滨, 臧家业, 王能飞. 北极新奥尔松地区土壤中生物硅含量及形态特征. 极地研究, 2015, 27(3): 237-245. |
| [68] |
Clymans W, Struyf E, Govers G, Vandevenne F, Conley D J. Anthropogenic impact on biogenic Si pools in temperate soils. Biogeosciences Discussions, 2011, 8(3): 4391-4419. |
| [69] |
Struyf E, Dausse A, Van Damme S, Bal K, Gribsholt B, Boschker H T S, Middelburg J J, Meire P. Tidal marshes and biogenic silica recycling at the land-sea interface. Limnology and Oceanography, 2006, 51(2): 838-846. DOI:10.4319/lo.2006.51.2.0838 |
| [70] |
刘森, 冉祥滨, 车宏, 马永星, 臧家业. 黄河口湿地土壤中生物硅的分布与植硅体的形态特征. 土壤, 2014, 46(5): 886-893. |
| [71] |
Norris A R, Hackney C T. Silica content of a mesohaline tidal marsh in North Carolina. Estuarine, Coastal and Shelf Science, 1999, 49(4): 597-605. DOI:10.1006/ecss.1999.0506 |
| [72] |
Zhang Q Z, Tao Z, Ma Z W, Gao Q Z, Deng H J, Xu P, Ding J, Wang Z G, Lin Y W. Hydro-ecological controls on riverine organic carbon dynamics in the tropical monsoon region. Scientific Reports, 2019, 9: 11871. DOI:10.1038/s41598-019-48208-y |
| [73] |
Zhang Q Z, Tao Z, Ma Z W, Tang W K, Gao Q Z, Xu P, Ding J, Liu Z F, Lin Y W, Su D, Zheng W. Influences of anthropogenic activities on dissolved silica migration in a granite-hosted basin, Hainan Island, China. Quaternary International, 2017, 440: 99-110. DOI:10.1016/j.quaint.2016.09.037 |