 2022, Vol. 42
2022, Vol. 42文章信息
- 赵洪贤, 张洋军, 徐铭泽, 卫腾宙, 毛军, 雒宇, 贾昕, 查天山
- ZHAO Hongxian, ZHANG Yangjun, XU Mingze, WEI Tengzhou, MAO Jun, LUO Yu, JIA Xin, ZHA Tianshan
- 油蒿叶片氮分配对其最大净光合速率季节变异的影响
- Effects of leaf nitrogen allocation on seasonal variation in maximum net photosynthetic rate in Artemisia ordosica
- 生态学报. 2022, 42(17): 7156-7166
- Acta Ecologica Sinica. 2022, 42(17): 7156-7166
- http://dx.doi.org/10.5846/stxb202107061799
-
文章历史
- 收稿日期: 2021-07-06
- 修订日期: 2021-12-29





2. 宁夏盐池毛乌素沙地生态系统国家定位观测研究站, 盐池 751500;
3. 北京林业大学水土保持国家林业局重点实验室, 北京 100083;
4. 甘肃省林业科学研究院, 兰州 730020
2. Yanchi Ecology Research Station of Mu Us Desert, Yanchi 751500, China;
3. Key Laboratory of State Forestry Administration on Soil and Water Conservation, Beijing Forestry University, Beijing 100083, China;
4. Gansu Research Academy of Forestry Science and Technology, Lanzhou 730020, China
光合作用是碳循环的驱动力, 植物叶片的光合作用模型可以反映植物对环境的适应能力[1—2]。叶片最大净光合速率(Amax)衡量光合潜力, 通过研究植物叶片光合参数的动态变化, 可以分析植物对其所处环境的响应与其叶片光合作用机构运转状况。
叶片光合能力受生物与非生物因素的限制, 这些限制因素多是由植物的个体发育和外界环境因素造成[3]。如对于荒漠植物, 空气温度(Ta)和光合有效辐射(PAR)可以通过影响植物叶片气孔的开度, 影响CO2的吸收, 从而影响植物的最大净光合速率(Amax)[4], 也有研究报道Ta和叶片比叶面积(SLA)是影响油蒿最大羧化效率(Vcmax)和最大电子传递速率(Jmax)的主要影响因素[5], 根据前人研究发现, 对于半干旱区植物, 植物对水的可获得性是限制其生长的重要环境因子[6], 土壤水分是重要的限制因素, 水分胁迫会对植物叶片的Vcmax有明显的抑制作用[7]。
氮是植物光合作用过程中重要的营养元素, 最大净光合速率与叶氮含量之间的关系被广泛认可, 它的供应直接影响植物的光合作用[8]。叶绿素是植物进行光合作用的主要色素, 其含量Cc对植物的光合速率有较大的影响, 它的含量也会影响氮在叶片中的分配[9]。为满足不同生长阶段植物对氮的需求, 植物对光合作用和构建细胞壁的氮分配权衡可能存在其不同的生长阶段[10], 叶片中的氮素会随着植物体的需求分配给光合系统和非光合系统[11—12]。光合系统包括一系列相互作用的子系统, 分配到的氮按照一定的比例分配给羧化系统(Pc)、生物力学系统(Pb)、捕光系统(PL)。非光合系统组分中分配的氮被用来构建植物细胞的细胞壁等防御机构[13—14]。有研究提出在同一地区, 有些物种为进行更快更高效的光合, 会将更多的氮分配到叶绿体中, 以争取更多的养分[15—16];也有研究认为当植物处于环境胁迫条件下时, 会更倾向于投资非光合机构从而具有较小的氮利用效率和较低的光合能力[13]。因此氮素在植物叶片中的分配对植物的光合作用有着极大的影响。根据已有研究发现, 叶片氮素在植物叶片不同机构中分配的季节动态会受环境因素和植物自身生理特性的影响[4, 7, 17], 如有研究认为在低氮水平下, 植物倾向于将氮素更多的用于光合系统, 以提高其光合能力[18]。
最大电子传递速率与最大羧化效率的比值Jmax/Vcmax可以表明氮在植物光合系统中的分配情况[19];Vcmax和Jmax可以描述叶片的光合能力, 是决定碳同化速率的关键[20—22], 光合作用发生在叶绿体中, 涉及光依赖和非光依赖的生化反应, 前者受到Jmax的限制[22];Vcmax与光合作用的关键酶Rubisco直接相关, Rubisco与植物叶片的光合速率相关[22]。而Vcmax和Jmax是估算氮在叶片光合机构中的分配比例的重要参数[16], 因此, 本研究推测植物最大净光合速率的季节动态与植物叶氮含量在叶片不同机构中的分配有关。
目前, 关于荒漠植物光合响应曲线参数季节动态及其调控机理的研究, 多集中在环境因子或植物叶片性状对植物光合速率的影响[23—25], 也有人研究了叶氮含量对光合特性的影响[5, 26], 而将最大净光合速率与叶氮分配的季节动态结合研究的报道非常有限。在土壤肥力较低的荒漠地区, 植物叶片的最大净光合速率与氮素在叶片各个系统间分配比例的关系是什么, 在不同的物候时期, 植物如何调节对叶片氮素的利用策略来满足自身在不同时期生长的需求, 仍需进一步的研究。干旱半干旱地区是全球变暖过程中最敏感脆弱的地区之一, 其在我国的分布范围占国土总面积的48.8%[4, 27], 对此区域内的荒漠植物生理机制的研究十分必要。因此, 本研究以荒漠典型灌木油蒿(Artemisia ordosica)为研究对象, 通过对光合响应曲线的全生长期原位观测和叶片氮含量的测定, 分析油蒿最大净光合速率的季节动态, 阐明叶氮分配的季节变化对油蒿最大净光合速率季节动态的影响。
1 材料和方法 1.1 研究区概况实验在宁夏盐池毛乌素沙地生态系统国家定位观测研究站(37°42′31″N, 107°13′47″ E)进行。气候类型属于典型中温带大陆性季风气候, 降水主要集中在7—9月, 年降水量约287 mm, 蒸发量约是降水量的7倍[4]。年平均气温7.7 ℃, 昼夜温差较大, 日照充足。研究区内优势植物种为油蒿。根据研究站之前观察结果, 油蒿展叶期在一年的第100—150天(4月11日至6月1日), 完全展叶期在第151—240天(6月2日至8月29日), 落叶期在第241—290天(8月30日至10月18日)[28]。生长季内, 试验样地土壤全氮含量均值为0.17g/kg。
1.2 研究方法实验于2018年5—10月进行, 结合手动测量与通量塔连续观测的气象数据进行分析研究。在距通量塔1公里处的100m×100 m油蒿样地内, 随机选择5株受光均匀的成年油蒿植株作为观测样株。
从2018年5月3日到9月24日, 以10d为一个周期进行观测, 选择周期内晴朗无风的天气, 使用LI-6400XT便携式光合测定仪(LI-COR, USA)和LI-6400的红蓝光源叶室(LI-COR, USA)在8:00—11:30测量油蒿的光响应曲线和CO2响应曲线, 流速设置为500 μmol/s。测量光响应曲线时, 光强梯度设置为2000、1800、1500、1200、1000、800、600、400、200、100、50和0 μmol m-2 s-1, 在每个光强梯度下适应3 min, 仪器自动将样品室与参比室进行匹配后记录其光合速率;测量CO2响应曲线时, 设定光强为1800 μmol m-2 s-1, 利用LI-6400的注入系统和外接CO2小钢瓶提供并控制CO2浓度, CO2浓度梯度设置为400、200、100、50、400、600、800、1000、1200、1500、1800、2000 μmol CO2/mol, 在每个浓度下适应3 min后, 仪器自动测量其光合速率并记录。将被测叶片带回实验室, 称量其叶片鲜重, 并用ImagJ软件分析计算得到其叶面积, 并对所测得的光合响应曲线数据进行校正。
在各个样株上采集与所测叶片叶位、叶形、叶龄相近的叶片若干, 称取0.2g的新鲜叶片, 分别加入适量碳酸钙、石英砂和95%的乙醇, 用研钵研成均浆, 抽滤后用95%的乙醇定容至25mL, 以95%的乙醇溶液作为空白对照, 在波长649nm和665nm下测定提取液的吸光度, 根据经验公式法, 计算得出油蒿叶片叶绿素含量Cc;其余叶片在105℃下杀青2h, 接着在75℃下持续烘干至恒重, 用研钵制成粉末状, 过100目筛, 每份称取1.5g, 装入自封袋中, 妥善保存至生长季结束, 带回北京委托中国科学研究院植物所测定叶片氮含量。
1.3 数据处理及分析 1.3.1 数据处理根据分光光度计法实验原理中提供的计算公式, 计算油蒿叶片中叶绿素含量Cc(mg/g):
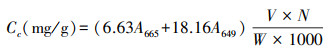
|
式中, A665为叶绿素在665nm波长下的吸光度, A649为叶绿素在649nm波长下的吸光度, V为提取液体积, N为稀释倍数, W为样品鲜质量(g)。
本研究中利用叶子飘等[29—30]提出的双曲线修正模型对所测得的光响应曲线进行拟合计算, 得到Amax:
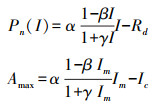
|
式中β为修正系数[30];α是光响应曲线的初始斜率;γ是光响应曲线的初始斜率与植物最大光合速率的比值;Ic是植物的光补偿点;I为光合有效辐射;Rd为植物的暗呼吸速率[31];其中Im为植物的饱和光照强度, 其计算公式为:
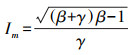
|
CO2响应曲线参数的计算, 是根据1980年Farquhar等提出的FvCB模型, 计算最大羧化效率Vcmax(μmol m -2 s-1)和最大电子传递速率Jmax(μmol m -2 s-1)[32—33]。本研究中, 数据分析使用RStudio 4.0.5软件[34], 利用R包plantecophys进行拟合计算, 公式如下:

|
式中, Ac为Rubisco活动限制下净光合速率, Ci为胞间CO2浓度, Oi为胞间O2浓度, Г*为无暗呼吸时的CO2补偿点, Kc, Ko分别为CO2和O2 Rubisco活性的Michaelis-Menten系数[35—36];J为电子传输速率, θ取值0.85, α取值0.24, 核酮糖-1, 5-双磷酸(RuBP)Aj为RuBP再生受到限制时的净光合速率[36]。
本研究中, 用经验公式法估算油蒿叶片氮素在各组分之间的分配比例[16], 根据Vcmax、Jmax和叶绿素含量Cc估算氮在羧化系统中的分配比例Pc、在生物力能学组分中的比例Pb和在捕光系统中的分配比例PL[16, 37], 计算公式如下:
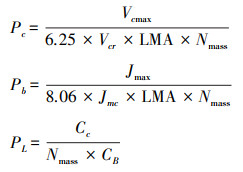
|
式中, Vcr是单位Rubisco的CO2固定活性, 取值为20.78 μmolCO2g-1 Rubisco-1, Jmc是单位Cyt f的最大电子传递速率, 取值为155.65 μmol e-μmol-1cyt fs-1 [14, 16, 37];Nmass为单位干重叶片氮含量(g/g), Cc为叶绿素含量(mmol/g), CB为捕光系统中叶绿素与氮的比值(mmol Chl/g N)[16], 其计算公式为:
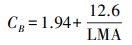
|
式中, LMA为比叶重(g/m2), 为叶片干重与叶面积的比值。氮在叶片光合系统的分配比例Pp与在非光合系统中的分配比例P1-P为:
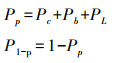
|
基于SPSS软件, 用双变量相关性分析法和回归分析法分析油蒿叶片最大净光合速率Amax与各个生物相关性, 基于R[34], 利用agricolae包, 对叶氮分配相关参数与Amax之间的关系进行通径分析, 并基于通径系数表, 分析各因子对Amax季节动态的作用路径;软件制图采用Origin9.0。
2 结果 2.1 油蒿最大净光合速率的季节动态在不同的物候时期, 由于植物对光强、温度等因子的敏感度不同, 油蒿最大净光合速率(Amax)存在季节动态。在生长季内, 油蒿的Amax在23.42 μmol CO2 m-2 s-1周围波动变化(图 1), 在完全展叶中期(第182天)有最大值29.48 μmol CO2 m-2s-1, 在完全展叶末期有最小值16.85 μmol CO2 m-2 s-1, Amax季节变异系数(Cv)为14%。Vcmax和Jmax具有相同的季节变异趋势(图 1)。

|
| 图 1 毛乌素沙地油蒿光合参数季节动态 Fig. 1 Seasonal variations in photosynthetic parameters in Artemisia ordosica in Mu Us Sandy land 横向参照线为各参数观测期内均值, 纵向参照线划分不同物候期; 横坐标即年积日 |
油蒿叶片氮分配的相关参数Pc、Pb、PL在生长季内呈波动变化, 油蒿叶片氮利用效率(PNUE)在完全展叶中期有最大值11.82 μmolCO2 gN -1 s-1(图 2)。油蒿叶氮分配参数在生长季内呈波动变化, 均在完全展叶中期有最大值(第182天), Pc、Pb、PL分别为0.26、0.09、0.08(图 2), 即Pp在此时有最大值0.43(Pc+Pb+PL), 此时叶片氮素在油蒿光合系统中的分配比例最大。在完全展叶末期(第232天), Pc、Pb、PL有最小值(图 2), 此时油蒿叶氮在非光合系统中的分配比例P1-p有最大值0.79, PNUE也在此时有最小值6.72μmolCO2 gN -1 s-1。

|
| 图 2 2018年油蒿生物因子季节动态 Fig. 2 Seasonal variations in biotic factors in Artemisia ordosica in 2018 横向参照线为各参数观测期内均值, 纵向参照线划分不同物候期。横坐标即年积日 |
对油蒿最大净光合速率Amax与Nmass进行回归分析, 结果显示, Amax与Nmass极显著正相关(P < 0.01)(图 3), 即油蒿最大净光合速率与叶片氮含量相关;因此对Amax与叶片氮分配相关参数Pc、Pb、PL进一步进行回归分析, 结果表明Amax与Pb极显著正相关(P < 0.01), 与PL显著正相关(P < 0.05), 与Pc相关, 与P1-p显著负相关(P < 0.05)(图 4)。

|
| 图 3 2018年毛乌素沙地油蒿叶片最大净光合速率Amax与叶片氮含量Nmass的关系 Fig. 3 Relationship between the maximum net photosynthetic rate Amax and Nmass in Artemisia ordosica in Mu Us sandy land in 2018 |

|
| 图 4 2018年毛乌素沙地油蒿叶片最大净光合速率Amax与叶片氮分配相关参数的关系 Fig. 4 Relationship between the maximum net photosynthetic rate Amax and nitrogen distribution parameters in Artemisia ordosica in Mu Us sandy land in 2018 |
主要分析2018年5—10月油蒿最大净光合速率受氮分配相关参数的调控机制, 但各个因子之间的相关性导致的信息重叠, 因此本研究基于通径分析方法, 对Pc、Pb、PL对Amax的影响进行通径分析, 根据直接、间接通径系数(表 1)分析其对油蒿Amax季节动态的影响路径。
| 自变量 Independent variable |
相关系数 Correlation coefficient |
直接通径系数 Direct path coefficient |
间接通径系数 Indirect path coefficient |
| 氮在捕光系统中分配比例PL Nitrogen distribution in light capture system (PL) |
0.44 | 0.406 | 0.233(PL→Pb→Amax) 0.070(PL→Pc→Amax) |
| 氮在生物力能学组分中的比例Pb Nitrogen distribution in bioenergy system (Pb) |
0.49 | 0.343 | 0.276(Pb→PL→Amax) 0.140(Pb→Pc→Amax) |
| 氮在羧化系统中的比例Pc Nitrogen distribution in carboxylation system (Pc) |
0.36 | 0.185 | 0.261(Pc→Pb→Amax) 0.154(Pc→PL→Amax) |
通径分析结果表明, 氮在捕光系统中分配比例PL的直接通径系数为正值且最大, 其次是氮在生物力能学组分中的比例Pb和氮在羧化系统中的比例Pc均对Amax产生了正向影响。Pc通过Pb间接作用于Amax的间接通径系数为正值。
3 讨论叶片氮素在光合与非光合系统之间的分配权衡已经被广泛讨论。在本研究中, 通过对研究区主要建群种油蒿的光合参数及叶片氮含量的全生长期原位测定, 研究其叶片氮分配对最大净光合速率季节变化的影响。本文的研究结果表明, 油蒿在不同的物候阶段, 为满足其生长的需求, 氮在叶片不同机构中的分配比例不同[14], 植株通过不同的氮素利用策略来维持生长。油蒿叶片Amax在完全展叶中期有最大值, 这与前人研究所得出的油蒿叶片Amax在展叶期和落叶期有较大值有别[5], 说明油蒿Amax的季节动态特征可能存在年际差异。氮素向叶片光合机构的分配是PNUE重要的决定因素之一[11], 在观测期内, Jmax/Vcmax在完全展叶中期有最大值3.86(图 5), 而Jmax与Vcmax的比值可以表示叶片氮素在光合系统中的分配情况[19], 即在该物候期油蒿羧化和RuBP再生速率较强[22], 可能是由于在此期间, 油蒿叶片氮素在光合系统的分配比例Pp有最大值(图 2), 氮素的分配倾向于光合系统, 叶片向Rubisco投入更多的氮以增加光合能力, PNUE也在此物候期有最大值, 同时油蒿叶片具有较小的LMA, 即此时叶片的光合能力最大, 光合氮利用效率最高, 在此物候时期油蒿处于快速生长阶段。在不同物候时期, 叶片氮分配的改变有利于植物在自然生境中的生存。本研究发现, 在完全展叶初期和末期, 油蒿叶片氮素分配更倾向于非光合系统, 表现为在这两个时期Pp均有最小值0.21(第162天、第232天), 此时油蒿叶片光合速率较小, 光合能力较弱, 而比叶重(LMA)较高(图 5), 这说明油蒿在叶片结构生物量中投入了更多的氮, 有利于维持叶片机械韧性, 获得较长的叶片寿命, 提高油蒿抵抗不利环境的能力[12—13]。

|
| 图 5 2018年毛乌素沙地油蒿叶片比叶重LMA和Jmax/Vcmax的季节动态 Fig. 5 Seasonal variations in LMA and Jmax/Vcmax in Artemisia ordosica in Mu Us sandy land in 2018 横向参照线为各参数观测期内均值, 纵向参照线划分不同物候期; 横坐标即年积日 |
氮素对植物叶片光合系统所有的组成部分都至关重要[22], 在本研究中, Nmass与Amax显著相关, 随着单位干重叶片氮含量的增加, Amax显著增加, 即油蒿最大光合能力增大, 这与前人研究结果相似[17]。油蒿叶片氮素在各个光合机构中的分配比例与Amax均正相关, 随着其在光合系统中的分配总比例Pp增加, 油蒿最大净光合速率Amax增大。本研究中, Amax与LMA显著负相关(图 6), 与前人研究的具有高LMA的植株具有较低的光合速率的结论相似[9, 13], 这可能是由于在具有较高LMA的叶片中, 有更多的氮被用于构建细胞壁, 这可能导致分配给Rubisco的氮含量相对减少, 分配到光合系统的氮较低[12—14, 38], 使油蒿光合能力降低, 表现为Amax减小(图 6)。在观测期内油蒿的PNUE明显低于常绿灌木[39], 根据前人对不同物种PNUE差异的生理原因的研究结果表明, 这可能是油蒿以牺牲PNUE来提高其在非光合系统中氮的投入, 从而提高叶片应对干旱胁迫时的抗逆性[11, 14, 39]。

|
| 图 6 2018年毛乌素沙地油蒿叶片最大净光合速率Amax与比叶重LMA、叶绿素含量Cc的关系 Fig. 6 Relationship between the maximum net photosynthetic rate Amax and LMA and Cc in Artemisia ordosica in Mu Us sandy land in 2018 |
本研究发现油蒿叶片氮素在捕光系统中分配比例PL对Amax产生了最大的正向影响, 这可能是由于PL与叶片叶绿素含量Cc密切相关[16], 而本研究中Amax与Cc显著正相关(P < 0.05)(图 6), 因此PL对Amax有最大的直接正向影响。从回归分析结果可知, 氮在生物力能学组分中分配比例Pb对Amax的季节动态变化解释度最大(图 4), 通径分析也表明Pb对Amax有直接的正向影响, Pb对Amax的影响可能是由于当CO2浓度到达一定大小时, 植株叶片的光合作用受最大电子传递效率Jmax的限制[2, 18, 22], 从而体现为正相关关系;回归分析结果显示氮在羧化系统分配比例Pc与Amax的季节动态变化呈不显著正相关(图 4), 但本研究发现Pc主要通过Pb对Amax的季节动态产生了间接的正向影响, 这可能是由于油蒿叶片在不同的物候时期, 会通过调节叶片氮素在羧化系统和生物力能学系统之间的分配比例而影响Amax的大小[19]。油蒿在生长旺盛时期叶片倾向于投资较多的氮进行光合作用, 在生长初期和末期则倾向于投资更多的氮来提高植物叶片的机械韧性, 维持较长的叶片寿命。本文的研究表明了荒漠典型灌木油蒿在不同物候时期通过调控氮在叶片光合和非光合系统之间的分配比例, 通过不同的氮素利用策略, 调节叶片最大净光合速率与叶片机械韧性之间的权衡以适应环境变化。
4 结论(1) 观测期内, 油蒿最大净光合速率(Amax)在完全展叶中期(第182天)有最大值, 在此物候阶段, 油蒿的光合能力最大。
(2) 油蒿Amax与氮素在叶片各个光合机构中的分配比例均正相关, 考虑氮分配相关参数对Amax季节动态的影响时, 氮在捕光系统中分配比例PL与Amax呈显著正相关, PL对Amax的季节动态有最大的直接正向影响, 氮在生物力能学组分中的比例Pb与Amax极显著正相关(R2=0.49, P < 0.01), 氮在羧化系统中分配比例Pc主要通过影响Pb对Amax产生正向作用。
| [1] |
Sitch S, Smith B, Prentice I C, Arneth A, Bondeau A, Cramer W, Kaplan J O, Levis S, Lucht W, Sykes M T, Thonicke K, Venevsky S. Evaluation of ecosystem dynamics, plant geography and terrestrial carbon cycling in the LPJ dynamic global vegetation model. Global Change Biology, 2003, 9(2): 161-185. DOI:10.1046/j.1365-2486.2003.00569.x |
| [2] |
Walker A P, Beckerman A P, Gu L H, Kattge J, Cernusak L A, Domingues T F, Scales J C, Wohlfahrt G, Wullschleger S D, Woodward F I. The relationship of leaf photosynthetic traits-Vcmax and Jmax - to leaf nitrogen, leaf phosphorus, and specific leaf area: a meta-analysis and modeling study. Ecology and Evolution, 2014, 4(16): 3218-3235. DOI:10.1002/ece3.1173 |
| [3] |
Grassi G, Magnani F. Stomatal, mesophyll conductance and biochemical limitations to photosynthesis as affected by drought and leaf ontogeny in ash and oak trees. Plant, Cell & Environment, 2005, 28(7): 834-849. |
| [4] |
李鑫豪, 闫慧娟, 卫腾宙, 周文君, 贾昕, 查天山. 油蒿资源利用效率在生长季的相对变化及对环境因子的响应. 植物生态学报, 2019, 43(10): 889-898. DOI:10.17521/cjpe.2019.0214 |
| [5] |
王子奇, 查天山, 贾昕, 吴雅娟, 张明艳, 穆家伟. 油蒿光合参数季节动态及其与叶氮含量和比叶面积的关系. 生态学杂志, 2017, 36(4): 916-924. |
| [6] |
Nemani R R, Keeling C D, Hashimoto H, Jolly W M, Piper S C, Tucker C J, Myneni R B, Running S W. Climate-driven increases in global terrestrial net primary production from 1982 to 1999. Science, 2003, 300(5625): 1560-1563. DOI:10.1126/science.1082750 |
| [7] |
闫霜, 张黎, 景元书, 何洪林, 于贵瑞. 植物叶片最大羧化速率与叶氮含量关系的变异性. 植物生态学报, 2014, 38(6): 640-652. |
| [8] |
Hou W F, Tränkner M, Lu J W, Yan J Y, Huang S Y, Ren T, Cong R H, Li X K. Interactive effects of nitrogen and potassium on photosynthesis and photosynthetic nitrogen allocation of rice leaves. BMC Plant Biology, 2019, 19(1): 302. DOI:10.1186/s12870-019-1894-8 |
| [9] |
崔西甜, 袁凤辉, 王安志, 关德新, 吴家兵, 金昌杰. 蒙古栎叶片光合作用随叶龄的变化及其与叶片功能性状的关系. 生态学杂志, 2017, 36(11): 3160-3167. |
| [10] |
Tang J C, Sun B D, Cheng R M, Shi Z M, Luo D, Liu S R, Centritto M. Seedling leaves allocate lower fractions of nitrogen to photosynthetic apparatus in nitrogen fixing trees than in non-nitrogen fixing trees in subtropical China. PLoS One, 2019, 14(3): e0208971. DOI:10.1371/journal.pone.0208971 |
| [11] |
Hikosaka K, Hanba Y T, Hirose T, Terashima I. Photosynthetic nitrogen-use efficiency in leaves of woody and herbaceous species. Functional Ecology, 1998, 12(6): 896-905. DOI:10.1046/j.1365-2435.1998.00272.x |
| [12] |
Onoda Y, Wright I J, Evans J R, Hikosaka K, Kitajima K, Niinemets V, Poorter H, Tosens T, Westoby M. Physiological and structural tradeoffs underlying the leaf economics spectrum. New Phytologist, 2017, 214(4): 1447-1463. DOI:10.1111/nph.14496 |
| [13] |
Wright I J, Reich P B, Westoby M, Ackerly D D, Baruch Z, Bongers F, Cavender-Bares J, Chapin T, Cornelissen J H C, Diemer M, Flexas J, Garnier E, Groom P K, Gulias J, Hikosaka K, Lamont B B, Lee T, Lee W, Lusk C, Midgley J J, Navas M L, Niinemets V, Oleksyn J, Osada N, Poorter H, Poot P, Prior L, Pyankov V I, Roumet C, Thomas S C, Tjoelker M G, Veneklaas E J, Villar R. The worldwide leaf economics spectrum. Nature, 2004, 428(6985): 821-827. DOI:10.1038/nature02403 |
| [14] |
刘涛. 氮供应对油菜叶片氮形态分配与光合氮利用效率的影响机制[D]. 武汉: 华中农业大学, 2018.
|
| [15] |
Feng Y L, Auge H, Ebeling S K. Invasive Buddleja davidii allocates more nitrogen to its photosynthetic machinery than five native woody species. Oecologia, 2007, 153(3): 501-510. DOI:10.1007/s00442-007-0759-2 |
| [16] |
史作民, 唐敬超, 程瑞梅, 罗达, 刘世荣. 植物叶片氮分配及其影响因子研究进展. 生态学报, 2015, 35(18): 5909-5919. |
| [17] |
卫腾宙. 油蒿叶光合氮利用效率的季节动态与机制[D]. 北京: 北京林业大学, 2019.
|
| [18] |
陈宗梅. 叶氮在光合机构和抗虫蛋白或遗传物质间分配的权衡[D]. 沈阳: 沈阳农业大学, 2016.
|
| [19] |
李登峰. 金沙江干热河谷4种生态恢复树种生理生态特征的干湿季对比研究[D]. 雅安: 四川农业大学, 2019.
|
| [20] |
Kattge J, Knorr W, Raddatz T, Wirth C. Quantifying photosynthetic capacity and its relationship to leaf nitrogen content for global-scale terrestrial biosphere models. Global Change Biology, 2009, 15(4): 976-991. DOI:10.1111/j.1365-2486.2008.01744.x |
| [21] |
Song G M, Wang Q, Jin J. Leaf photosynthetic capacity of sunlit and shaded mature leaves in a deciduous forest. Forests, 2020, 11(3): 318. DOI:10.3390/f11030318 |
| [22] |
Quebbeman J A, Ramirez J A. Optimal allocation of leaf‐level nitrogen: implications for covariation of Vcmax and Jmax and photosynthetic downregulation. Journal of Geophysical Research: Biogeosciences, 2016, 121(9): 2464-2475. DOI:10.1002/2016JG003473 |
| [23] |
李雪娇, 李伟, 高冠龙. 极端干旱区胡杨净光合速率日变化及主要影响因子. 山西大学学报: 自然科学版: 1-9. [2020-11-13]. https: //doi.org/10.13451/j.sxu.ns.2020091.
|
| [24] |
郑有飞, 倪艳利, 麦博儒, 吴荣军, 冯妍, 孙健, 李健, 徐静馨. 太阳辐射减弱对冬小麦旗叶光合速率的影响. 应用生态学报, 2011, 22(6): 1457-1464. |
| [25] |
张彦敏, 周广胜. 植物叶片最大羧化速率及其对环境因子响应的研究进展. 生态学报, 2012, 32(18): 5907-5917. |
| [26] |
李勇. 氮素营养对水稻光合作用与光合氮素利用率的影响机制研究[D]. 南京: 南京农业大学, 2011.
|
| [27] |
张存杰, 廖要明, 段居琦, 宋艳玲, 黄大鹏, 王胜. 我国干湿气候区划研究进展. 气候变化研究进展, 2016, 12(4): 261-267. |
| [28] |
李媛. 半干旱区油蒿光合生理参数季节动态[D]. 北京: 北京林业大学, 2015.
|
| [29] |
叶子飘, 段世华, 安婷, 康华靖. C4作物电子传递速率对CO2响应模型的构建及应用. 植物生态学报, 2018, 42(10): 1000-1008. DOI:10.17521/cjpe.2018.0129 |
| [30] |
叶子飘, 于强. 一个光合作用光响应新模型与传统模型的比较. 沈阳农业大学学报, 2007, 38(6): 771-775. DOI:10.3969/j.issn.1000-1700.2007.06.001 |
| [31] |
叶子飘. 光合作用对光和CO2响应模型的研究进展. 植物生态学报, 2010, 34(6): 727-740. DOI:10.3773/j.issn.1005-264x.2010.06.012 |
| [32] |
Duursma R A. Plantecophys - An R package for analysing and modelling leaf gas exchange data. PLoS One, 2015, 10(11): e0143346. DOI:10.1371/journal.pone.0143346 |
| [33] |
唐星林, 曹永慧, 顾连宏, 周本智. 基于FvCB模型的叶片光合生理对环境因子的响应研究进展. 生态学报, 2017, 37(19): 6633-6645. |
| [34] |
R Core Team. R: A Language and Environment for Statistical Computing. Vienna, Austria: R Foundation for Statistical Computing, 2020.
|
| [35] |
Farquhar G D, von Caemmerer S, Berry J A. A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta, 1980, 149(1): 78-90. DOI:10.1007/BF00386231 |
| [36] |
Medlyn B E, Dreyer E, Ellsworth D, Forstreuter M, Harley P C, Kirschbaum M U F, Roux X L, Montpied P, Strassemeyer J, Walcroft A, Wang K, Loustau D. Temperature response of parameters of a biochemically based model of photosynthesis. Ⅱ. A review of experimental data. Plant, Cell & Environment, 2002, 25(9): 1167-1179. |
| [37] |
Niinemets V, Tenhunen J D. A model separating leaf structural and physiological effects on carbon gain along light gradients for the shade-tolerant species Acer saccharum. Plant, Cell & Environment, 1997, 20(7): 845-866. |
| [38] |
Luo X Z, Keenan T F, Chen J M, Croft H, Prentice I C, Smith N G, Walker A P, Wang Han, Wang R, Xu C G, Zhang Y. Global variation in the fraction of leaf nitrogen allocated to photosynthesis. Nature Communications, 2021, 12: 4866. DOI:10.1038/s41467-021-25163-9 |
| [39] |
Takashima T, Hikosaka K, Hirose T. Photosynthesis or persistence: nitrogen allocation in leaves of evergreen and deciduous Quercus species. Plant, Cell & Environment, 2004, 27(8): 1047-1054. |