 2022, Vol. 42
2022, Vol. 42文章信息
- 梁高都, 田义超, 吴彬, 梁铭忠, 张文主, 杨小雄, 陆汝成, 张利国, 顾观海
- LIANG Gaodou, TIAN Yichao, WU Bin, LIANG Mingzhong, ZHANG Wenzhu, YANG Xiaoxiong, LU Rucheng, ZHANG Liguo, GU Guanhai
- 广西北部湾典型海岛红树林的结构特征、空间格局及种间种内关联性
- Structural characteristics, spatial patterns and interspecific and intraspecific associations of mangroves in typical islands of Guangxi Beibu Gulf
- 生态学报. 2022, 42(17): 7244-7255
- Acta Ecologica Sinica. 2022, 42(17): 7244-7255
- http://dx.doi.org/10.5846/stxb202106071508
-
文章历史
- 收稿日期: 2021-06-07
- 网络出版日期: 2021-12-29





2. 北部湾大学资源与环境学院, 钦州 535000;
3. 北部湾大学海洋学院, 钦州 535000
2. College of Resources and Environment, Beibu Gulf University, Qinzhou 535000, China;
3. College of Oceanography, Beibu Gulf University, Qinzhou 535000, China
植物种群结构和空间分布格局是植物生态学的重要内容[1]。种群是群落的基本组成部分, 种群结构的变化对群落特征产生直接影响, 能客观地表征群落的发展和演替趋势[2-3]。种群空间分布格局则是一定尺度上生境(气候、水文、地质和土壤等)、空间(物种和群落的相对位置)和生物(生态位、生活史特征、种间种内竞争、协同、干扰等力等)三大因素共同作用的结果[4-6]。因此, 解释植物种群结构与空间分布格局对研究单个或多个种群在群落内部的镶嵌结构和空间分布格局, 认识种群生物学特征、潜在的生态学过程及种群相互作用具有重要意义[7-8]。
空间点格局是生态学研究植物种群空间分布的重要方法, 通过零模型和点过程模型来模拟不同尺度对植物种群有重要影响的生态学过程[9-10]。种群空间格局能够揭示种群个体在某一地理分布区二维空间上的格局类型, 展现空间分布格局的聚集强度及其尺度效应, 并且可以探究种间、种内的空间关联性[11-14]。植物种群的空间分布格局和空间关联性是植物种群格局研究的两个主要内容, 可以反映出植物种群的生态调控能力和环境响应机制, 体现了种间、种内和环境间的交互性关系及其在群落中的地位和作用[15-16]。
海岛红树林与滨海红树林、海湾红树林、海岸红树林等相比, 因其位置的特殊性、面积的有限性以及空间的隔离性, 海岛红树林容易受到风暴潮、干旱等自然灾害的干扰, 且难以通过自身调节能力恢复, 具有明显的独立性和脆弱性特征[17-19]。但由于海岛生态系统受到人为干扰少, 能够较好地保留了原生红树林群落自然演替的生态学过程, 是植物生态学研究理想对象[20]。目前针对海岛红树林的研究主要集中在生态化学计量、群落结构、种群特征与环境因子协同关系等方面的研究, 而对红树林种群的空间点格局研究关注较少, 且缺乏海岛红树林生态适应对策的探讨[21-24]。鉴于此, 本文采用点格局分析方法, 小尺度范围内定量分析海岛红树林的群落结构、空间分布格局及其关联性, 以期为海岛红树林的保护与生态系统的综合管理提供依据。
1 研究区域与方法 1.1 研究区概况岛屿群落(20°58′-22°50′ N, 107°29′-110°20′ E)位于广西北部湾茅尾海红树林自然保护区的核心区域, 三面临海, 一面接陆, 地势自西北向东南倾斜。该区域属于南亚热带季风气候, 处在著名的亚洲东南部季风区内, 水热资源丰富。太阳辐射量4546.03 MJ/m2, 光合有效辐射量2373.48 MJ/m2, 年均日照时数1783.92 h, 年均气温22 ℃, 全年积温8046.30 ℃·d, 年均降雨量2105.20 mm, 年平均水温21.60 ℃。沿岸潮差较大, 平均潮差2.51 m, 最大潮差5.52 m, 属于强潮岸段。岛屿周围分布着大量的滩涂湿地, 是红树林集中生长的地区之一, 主要的红树林群落类型有桐花树(Aegiceras corniculatum)群落、秋茄(Kandelia obovata)+桐花树群落、白骨壤(Avicennia marina)+桐花树群落和白骨壤群落。
1.2 样地设置与调查在对广西茅尾海红树林自然保护区进行全面踏查的基础上, 于2020年7-8月在红树林分布相对集中的龟仙岛和背风墩岛屿周围的滨海滩涂湿地, 依据红树林群落的遥感分类结果, 综合考虑红树林生长状况、分布位置以及潮位等因素, 选取典型的红树林群落, 样方面积和调查方法参考《湿地生态系统定位观测技术规范》[25], 自内滩向外滩布设了1个40 m×40 m固定样地, 将样地划分成16个10 m×10 m的样方, 对样方内所有红树植物开展每木调查, 测量并记录每一棵红树植物的物种名称、树枝数、基径、树高、冠幅、覆盖度、相对坐标以及生长状态等基础数据。
1.3 高度级与大小级结构本研究借鉴胡刚等[2]对红树林群落的高度级和大小级的分级方法, 并结合海岛红树林群落的生活史特性, 以1.0 m为一个高度级将桐花树、秋茄和白骨壤的高度(H)划分为5级:Ⅰ级(H<1.0 m)、Ⅱ级(1.0 m≤H<2.0 m)、Ⅲ级(2.0 m≤H<3.0 m)、Ⅳ级(3.0 m≤H<4.0 m)、Ⅴ级(H≥4.0 m)。大小级根据桐花树、秋茄和白骨壤的基径(D)进行划分, 以基径<1 cm的植株, 按其高度划分为2级:Ⅰ级(0 m<H<0.5 m)、Ⅱ级(H≥0.5 m), 对于基径≥1 cm的植株, 基径每增加3 cm划分1级, Ⅲ级(1 cm≤D<4 cm)、Ⅳ级(4 cm≤D<7 cm)、Ⅴ级(7 cm≤D<10 cm)、Ⅵ级(10 cm≤D<13 cm)、Ⅶ级(13 cm≤D<16 cm)、Ⅷ级(D≥16 cm), 合计划分为8级。
常用高度级或大小级代替龄级分析种群的结构和动态特征, 采用“空间代替时间法”, 以高度或基径代替年龄作为结构划分标准[26]。首先对高度和基径进行相关性的回归分析, 验证以高度或基径代替年龄进行种群结构分析的可靠性, 并结合红树林群落的自身生长特性, 桐花树、秋茄以高度级划分为3个龄级:龄级Ⅰ(幼苗、幼树与小树, 高度级Ⅰ、Ⅱ级)、龄级Ⅱ(中树, 高度级Ⅲ级)、龄级Ⅲ(大树与老树, 高度级Ⅳ、Ⅴ级), 白骨壤以大小级划分为3个龄级, 龄级Ⅰ(幼苗、幼树与小树, 大小级Ⅰ、Ⅱ、Ⅲ、Ⅳ级)、龄级Ⅱ(中树, 大小级Ⅴ级)、龄级Ⅲ(大树与老树, 大小级Ⅵ、Ⅶ、Ⅷ级), 以确保不同龄级的株数。
1.4 研究方法 1.4.1 最近邻指数分析最近邻指数(Nearest neighbor indicator, NNI)通过点的最近邻距离来判断点的分布模式, 比较观察值的最近邻的点对的平均距离与随机分布模式下最近邻的点对的平均距离, 用两者比值来判断观察值与随机分布的偏离[27]。计算公式可表示为:
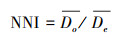
|
(1) |
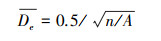
|
(2) |
式中:Do为观察平均最近邻距离, De为期望平均最近邻距离;n为样点数, A为研究区面积。当NNI<1时, 样点整体呈聚集分布, 且值越小越聚集, NNI>1时为离散分布, NNI=1时为随机分布。
1.4.2 点格局分析点格局分析主要包括概括性统计量的选择、零模型选择与模拟、模型的比较与检验等主要步骤, 是通过比较观测格局与零假设模拟产生的格局, 判断研究对象在不同尺度上的格局过程, 并根据生态学理论, 建立分布格局与生态学过程之间的联系[28-29]。点格局分析可以分为单变量、双变量以及多变量格局分析, 可以通过不同零假设模型来探索群落与种群分布格局、种间与种间关系以及生态过程在不同尺度上的响应。
本文采用配对相关函数g(r)(Pair correlation function)分析红树林群落的空间格局及空间关联性, 配对相关函数g(r)由Ripley K函数推演而来, 是基于圆环内的点进行计算, 不存在累积效应, 被认为是最好的二阶统计量, 可以较容易的扩展到双变量形式[30-31]。
单变量K(r)函数和g(r)函数的公式如下:
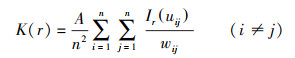
|
(3) |
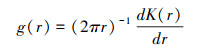
|
(4) |
双变量K12(r)函数和g12(r)函数的公式如下:
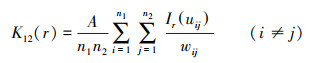
|
(5) |
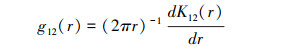
|
(6) |
式中:r为空间尺度, A为研究区面积, n是研究对象的样本数(n1与n2表示不同变量的样本数量), uij是i和j之间的距离, 当uij≤r, Ir uij =1;若Ir uij >r, 则Ir uij =0, wij为以i为圆心, uij为半径的圆位于A中的弧长与圆周长之比。
1.4.3 零模型选择零模型在点过程模型起到析因、求证、多种模拟推断和拟合等作用, 根据不同的研究目的和问题假设要选择正确合适的零模型是十分重要的[32]。本文中单变量函数g(r)选取了完全空间随机模型(Complete spatial randomness, CSR)、异质泊松模型(Heterogeneous Poisson model, HP), 其中完全空间随机模型是检验某一格局的空间分布模型, 而异质泊松模型可以排除生境异质性对某一格局的影响[31, 33]。双变量g12(r)函数则采用环形转变模型(Toroidal shift, TS), 是通过模拟某一格局的随机变动对另一格局的影响, 检验两种格局的空间关联性[34]。通过Monte-Carlo法检验零模型偏离观测值情况, 利用模拟的最大值和最小值构建上下两条包迹线, 即置信区间若观测值处于两条包迹线之间, 零假设成立, 否则拒绝零假设。
采用Programita[35]软件进行点格局分析, Origin 2018软件进行绘图, 选择配对相关函数g(r), 样方的空间尺度为0-40m, 环宽设置为3m, 分析步长为0.2m, 通过Monte-Carlo法模拟199次, 确定95%的置信区间。其他数据处理采用Excel 2016软件完成。
2 结果与分析 2.1 红树林种群的结构特征红树林群落生长和发育情况良好。调查样地内出现了桐花树、秋茄和白骨壤3种红树植物。桐花树相对矮小, 但树枝的数量较多;生长紧促, 冠幅较小, 但树叶茂密, 覆盖度高。秋茄呈翠绿色, 树干笔直, 植株较高, 但基径较小;树枝数量较多, 但冠幅较小, 覆盖度较低。白骨壤植株较高, 但树枝的分支数量较少;基径较粗, 冠幅较大, 树叶较小, 覆盖度较低。桐花树、秋茄和白骨壤之间差异显著(表 1)。
| 红树林群落Mangrove populations | 株高 Plant height/m |
基径 Base diameter/cm |
树支数 Tree branch |
冠幅 Crown width/m |
覆盖度 Coverage/% |
| 桐花树Aegiceras corniculatum | 2.08±0.36a | 4.06±1.39a | 5.57±2.59a | 1.10±0.53a | 98±3.03a |
| 秋茄Kandelia obovata | 2.76±0.66b | 5.66±2.02b | 3.25±3.37a | 1.31±0.52b | 64±0.39b |
| 白骨壤Avicennia marina | 3.37±0.65bc | 9.83±5.66bc | 1.63±1.43b | 1.98±1.09bc | 62±0.32c |
| 表内数据为平均值±标准误差, a、b、c表示不同树种间经过单因素方差分析, 通过固定差距检验(HSD)检验后的具有显著性差异(P<0.05) | |||||
高度级和大小级结构表明群落处于不同的演替周期(图 1)。桐花树高度级中位于Ⅱ、Ⅲ级的个体较多, 分别占总数的45.45%和51.03%;大小级中位于Ⅲ级的个体数最多, 占总数的58.67%。秋茄高度级中位于Ⅲ级的个体数占总数的45.99%, 大小级中位于Ⅳ、Ⅴ级的个体数分别占总数的52.19%、25.55%。白骨壤高度级中位于Ⅳ、Ⅴ级的个体数分别占总数的58.33%、21.97%, 大小级中位于Ⅳ、Ⅴ级的个体数分别占总数的32.58%、20.45%。桐花树中树较多, 且能维持一定的小树, 树种更新良好;秋茄中树、成年树较为丰富, 老树较少, 种群相对稳定;白骨壤个体数相对较少, 且小树较少成年树、老树较多, 种群更新受阻, 表现出衰退趋势。

|
| 图 1 高度级和大小级分布图 Fig. 1 Heightness class and Size class distribution map |
基径代替年龄分析种群结构的方法被广泛应用, 以往也存在用高度代替年龄的种群结构研究[26], 而原生红树林的树龄难以辨认且生境复杂, 其大小级或高度级结构是否能够较为精确地描述种群结构有待争议。基于此, 通过对高度和基径进行回归分析加以验证(图 2)。采用3种可能的曲线模型, 桐花树3种模型的相关系数R分别为0.472、0.439、0.418, 且均显示为中等相关(P<0.001), 秋茄3种模型的相关系数R分别为0.494、0.500、0.505, 后两种模型表现为强相关(P<0.001), 白骨壤则为0.562、0.547、0.569, 均显示为强相关(P<0.001), 表明三种红树林种群均存在一定的曲线相关性, 高度级或大小级能够较好地代替桐花树、秋茄、白骨壤的龄级结构分析。

|
| 图 2 3种可能的基径和树高的关系模型图 Fig. 2 Three possible diameter-height models |
桐花树是红树林群落的建群种。通过样方调查数据, 分别计算桐花树、白骨壤、秋茄的相对多度、相对频度、相对显著度和重要值(表 2)。桐花树的相对多度、相对频度、相对显著度均较大, 重要值达到59.43, 是红树林群落的先锋树种;白骨壤的相对多度较小, 但相对频度、相对显著度较大, 重要值也较大, 是仅次于桐花树的建群种;秋茄的相对频度、相对显著度最小, 重要值低于桐花树和白骨壤。不同红树植物群落的重要值充分体现各物种的不同优势地位。
| 红树林种群Mangrove populations | 相对多度 Relative abundance |
相对频度 Relative frequency |
相对显著度 Relative significance |
重要值 Important value |
| 桐花树Aegiceras corniculatum | 76.33 | 54.70 | 47.26 | 59.43 |
| 秋茄Kandelia obovata | 15.98 | 2.56 | 19.43 | 12.66 |
| 白骨壤Avicennia marina | 7.70 | 42.74 | 33.31 | 27.91 |
采用最近邻指数分析不同红树林种群的空间分布模式(表 3、图 3)。结果显示:红树林群落整体呈现空间集聚分布, 不同种群的红树林分布模式有所差异。桐花树、白骨壤的最近邻指数小于1, 其中桐花树的Z检验值小于-2.58(即置信区间为99%), 白骨壤的Z检验值小于-1.65(即置信区间为90%), 均为聚集分布模式。秋茄的最近邻指数大于1, 表现为随机分布。从红树林群落的集聚程度来看:桐花树>整体>白骨壤。
| 红树林种群Mangrove populations | 平均最近距离 Average closest distance/m |
期望最近距离 Expected closest distance/m |
最邻近指数 Nearest neighbor index |
Z检验值 Z test value |
置信区间 Confidence interval/% |
| 整体Overall | 0.44 | 0.48 | 0.91 | -7.37 | 99 |
| 桐花树Aegiceras corniculatum | 0.49 | 0.55 | 0.88 | -8.00 | 99 |
| 秋茄Kandelia obovata | 1.83 | 1.74 | 1.05 | 1.10 | - |
| 白骨壤Avicennia marina | 1.14 | 1.22 | 0.94 | -1.74 | 90 |

|
| 图 3 不同红树林种群个体分布点图 Fig. 3 Individual distribution points of different mangrove populations |
基于完全随机模型(CSR)的单变量g(r)函数分析表明(图 4), 红树林群落随着尺度的变化出现聚集、随机和均匀分布3种类型。桐花树种群在0.3-0.7 m尺度上是强烈聚集分布, 1.5-1.7 m和2.1-2.3 m尺度上属于均匀分布, 在其他尺度上基本表现为随机分布, 尽管在某些尺度上略有偏离;秋茄种群除了在0.1-0.5 m尺度上为集聚分布, 所有尺度上均为随机分布;白骨壤种群在4.3-5.3 m、6.7-7.1 m、9.5-10.3 m、13.7-14.5 m等尺度均出现集聚分布。通过异质泊松模型(HP)(图 5)排除生境异质性的影响, 桐花树种群在0.1-0.7 m、4.9-5.1 m为强烈集聚分布, 在1.3-2.7 m尺度上为均匀分布, 其他多为随机分布;秋茄种群在0.1-0.5 m上为集聚分布, 其余尺度上为随机分布;白骨壤种群0.1-0.7 m为轻微集聚分布、而4.9-5.1 m尺度上为强烈聚集分布, 在5.9-6.5 m上为均匀分布。

|
| 图 4 基于完全空间随机模型的点格局分析 Fig. 4 Analysis of Point Pattern based on CSR 实线代表观测值,虚线代表置信区间 |

|
| 图 5 基于异质泊松模型的点格局分析 Fig. 5 Analysis of Point Pattern based on HP 实线代表观测值,虚线代表置信区间 |
基于环形转换模型的双变量g12(r)函数分析不同种群的空间关联性, 结果表明(图 6):桐花树种群与秋茄种群、白骨壤种群之间随着尺度增加表现为空间无关联性-负关联性-无关联性的演变特征, 秋茄种群与白骨壤种群之间为空间无关联性。桐花树种群和秋茄种群在0.9-1.9 m为空间负关联性, 大于1.9 m后均为空间无关联性;桐花树种群和白骨壤种群在0.9-2.1 m和5.7-6.1 m为空间负关联性, 而秋茄种群和白骨壤种群则是整个尺度都为空间无关联性。

|
| 图 6 不同种群红树林的空间关联性分析 Fig. 6 Analysis of Spatial Correlation of Mangroves of Different Population 实线代表观测值,虚线代表置信区间 |
通过环形转换模型的双变量g12(r)函数分析不同龄级的3种红树林种群的空间关联性, 结果显示(图 7):桐花树的龄级Ⅰ-龄级Ⅱ(0.3-6.9 m、9.3-9.5 m尺度)、龄级Ⅰ-龄级Ⅲ(0.9-1.9 m、3.5-3.7 m尺度)之间存在空间负关联性, 而龄级Ⅱ-龄级Ⅲ(0.1-0.5 m尺度)则是存在空间正关联性;秋茄种群的不同龄级之间则在全尺度中均不存在空间关联性, 尽管在某些尺度上略有偏离;白骨壤则在不同龄级之间均存在一定程度的空间正关联性。

|
| 图 7 不同龄级的3种红树林种群的空间关联性分析 Fig. 7 Analysis of Spatial Correlation among Three Mangroves of Different Ages 实线代表观测值,虚线代表置信区间 |
红树植物种群结构与红树植物群落的演替阶段密切相关, 分析种群结构有助于揭示种群的生存状态和更新阶段, 推断种群演替的动态过程及其潜在的生态学机制[5]。桐花树种群小树和中树的个体数目比例较高, 种群结构金字塔型, 种群处于增长状态。秋茄种群中树个体数目最多, 成年树次之, 老树较少, 种群结构呈钟型, 种群数量处于稳定状态, 种群密度在一段时间内将保持稳定。白骨壤种群中幼树个体数目较少, 老树比例较大, 死亡率大于出生率, 种群结构呈纺锤型, 属于衰退型种群。桐花树主要分布于潮间带的低潮带和中潮带, 是红树林群落演替中、前期阶段中出现的树种;白骨壤主要分布于潮间带的低潮带, 是红树林演替早期阶段的先锋树种[36];秋茄多分布于潮间带的中潮带, 属于演替中、后期阶段的常见树种。红树植物种群结构表明岛屿群落红树植物种群正处于群落演替的中、前期阶段[2, 37]。
红树植物种群的更新状况与其所处的生境条件和种群生物学特征密切相关。海岛红树林易受到风暴潮、海浪等自然因素的干扰, 白骨壤作为海岛红树林群落的先锋树种, 其隐胎生繁殖特性、发达的根系和高大的树形使其能够快速占据海岛周围贫瘠的沙质裸地, 环境适应能力强, 且广泛分布于潮间带的低潮滩, 容易形成郁闭的潮间带先锋群落[38]。白骨壤群落形成后, 发挥了防风消浪和促淤保滩的生态服务功能, 加速了滩涂的淤积, 同时, 土壤有机质含量和盐度不断增加, 为适应淤泥质生境的其它红树植物生长创造了立地条件。随着土壤盐度和光照环境的改变致使白骨壤幼苗难以繁殖, 更新受阻[37]。桐花树是紫金牛科常绿灌木, 属于隐胎生繁殖, 种子的繁殖和自播能力较强, 是仅次于白骨壤的红树林先锋树种之一, 而秋茄具有显胎生繁殖特性, 因其快速适应淤泥质生境而逐步取得生长优势, 于是白骨壤群落受到具有萌发和更新能力较强的桐花树和秋茄相继入侵, 白骨壤由之前的大面积分布逐步演变为点状分布, 桐花树广泛分布于白骨壤周围, 秋茄则由零星分布演变为块状分布。因此, 广西北部湾茅尾海红树林从现有的白骨壤和白骨壤+桐花树群丛逐步演替为以秋茄为优势种的群丛, 如桐花树+秋茄群丛、秋茄群丛等[39]。
3.2 红树植物种群空间分布格局红树植物空间点格局是种群对周围环境长期适应和选择的结果。种群特征、扩散限制、密度依赖、生境异质性、人类活动干扰等生物与非生物因素和过程是影响物种在不同空间尺度上格局形成和变化的潜在生态学机制[40]。广西北部湾茅尾海红树植物种群在整体上呈现出空间集聚的分布格局, 其中, 桐花树种群表现出强烈的空间聚集分布特征。深圳福田的秋茄、桐花树和白骨壤[41], 珠海淇澳岛的无瓣海桑(Sonneratia apetala)、秋茄、卤蕨(Acrostichum aureum)和老鼠簕(Acanthus ilicifolius)[42], 广州南沙的无瓣海桑、黄槿(Hibiscus tiliaceus)和木榄(Bruguiera gymnorrhiza)[43], 湛江高桥的桐花树、白骨壤、红海榄(Rhizophora stylosa)和秋茄[44], 海南东寨港的角果木(Ceriops tagal)[45], 福建福鼎的秋茄[46]等红树林群落均呈现出空间集聚分布的特征。综上所述, 区域尺度上的聚集分布应该是我国沿海地区红树植物种群空间分布格局的主要特征。
植物种群在大尺度上空间分布格局可能是由物种分布的异质性以及土壤、水分、养分、盐度与地形等非生物环境的异质性造成的, 而在小尺度上, 种群空间分布格局可能是由于不同程度的种内或种间竞争、种子扩散限制、密度效应等生物因素所致[47]。桐花树种群在小尺度上表现为强烈集聚分布特征, 聚集强度随尺度的增加而有所下降。秋茄种群整体表现为随机分布, 但在小尺度上表现出轻微集聚分布特征。白骨壤种群在大尺度下表现为集聚分布。在剔除生境异质性的影响后, 桐花树、秋茄和白骨壤在不同尺度上都表现出更为显著的空间聚集分布特征。桐花树为隐胎生植物, 无性系繁殖特性有助于形成聚集格局, 轻且易于漂流的胎生苗有利于种群形成随机分布;秋茄种群尚处于发育早期, 基本上表现为随机分布特征;白骨壤种内竞争作用增强而造成密度制约性的自疏作用, 导致种群的个体数量递减, 降低了聚集程度。岛屿周围滩涂区域的地形起伏较大, 潮带分异和海水周期性淹没, 降低了红树植物种群分布的空间集聚度[48]。
3.3 红树植物的空间关联性植物种群内部的关联性是表示同一物种不同龄级在空间分布上的相互关联, 反映了种群内个体间的相互作用[49]。随着空间尺度的变化, 桐花树种群内部表现出空间正关联性、无关联性和负关联性, 幼树、小树与中树和成年树、老树之间存在空间负关联性, 中树与成年树、老树存在空间正关联性;秋茄种群的不同龄级之间均表现出空间无关联性;白骨壤种群则在不同龄级之间均出现空间正关联性。在资源限制的条件下, 种群内部相关竞争, 种群内不同龄级植株个体之间的关系呈现出负关联[14]。随着尺度的增大, 种群形成适宜自身生长的相互有利的环境, 个体之间产生了正向的关系, 产生强烈的“庇护和被庇护”效应, 增加种间竞争力, 抵御外来物种入侵, 使种群在群落中处于优势地位, 维持种群的生存和发展[47]。
不同植物种群之间的关联性是植物资源利用方式、种间相互作用和环境因素需求的表现, 反映了物种间的竞争关系和群落的环境资源配置情况, 种间关联性存在着较强的空间尺度效应[50]。桐花树和秋茄、白骨壤在小尺度上表现出无关联, 桐花树植株个体较小, 与秋茄、白骨壤之间不存在强烈的资源竞争, 相互之间竞争影响较小, 个体之间都处于一种稳定状态。桐花树和秋茄、白骨壤在中尺度上表现出负关联, 这可能是由于在较大尺度上不同种群各自都形成了较大的植物斑块, 对土壤水分、养分、光照等资源的获取起到了一定的限制作用, 种群之间表现出竞争关系。在大尺度上, 桐花树、秋茄、白骨壤并不互相依赖, 它们相对独立地共享着生境资源。
4 结论通过分析广西北部湾茅尾海自然保护区红树林岛屿群落的结构特征、空间格局及种间种内关联性, 红树林群落中桐花树、秋茄和白骨壤种群分别处于增长、稳定和衰退状态, 红树林种群处于群落演替的中、前期阶段。红树植物种群整体上呈现出空间集聚分布格局, 其中, 桐花树种群表现出强烈的空间聚集分布特征。桐花树种内个体之间随着尺度的变化出现空间无关联性、负关联性和正关联性, 秋茄种内关系表现出空间无联性, 白骨壤种内关系则出现空间正关联性。桐花树和白骨壤、秋茄种群之间呈现出一定的空间负关联性, 表现出种间的竞争关系。红树林人工种植和恢复需要参照红树林群落的自然演替规律, 研制出近似天然的人工红树林林分结构和功能的优化配置和调控技术, 提升红树林的植物丰富度和林分的混交度, 提高群落的生物多样性、群落稳定性和生态效益。
| [1] |
李明辉, 何风华, 潘存德. 天山云杉天然林不同林层的空间格局和空间关联性. 生态学报, 2011, 31(3): 620-628. |
| [2] |
胡刚, 黎洁, 覃盈盈, 胡宝清, 刘熊, 张忠华. 广西北仑河口红树植物种群结构与动态特征. 生态学报, 2018, 38(9): 3022-3034. |
| [3] |
范繁荣, 潘标志, 马祥庆, 沈琼桃, 谢荣樟. 白桂木的种群结构和空间分布格局研究. 林业科学研究, 2008, 21(2): 176-181. DOI:10.3321/j.issn:1001-1498.2008.02.008 |
| [4] |
孙世贤, 卫智军, 吴新宏, 姜超, 郭利彪. 不同放牧强度季节调控下荒漠草原主要植物种群点格局及空间关联性. 生态学报, 2016, 36(23): 7570-7579. |
| [5] |
Velázquez E, Kazmierczak M, Wiegand T. Spatial patterns of sapling mortality in a moist tropical forest: consistency with total density-dependent effects. Oikos, 2016, 125(6): 872-882. DOI:10.1111/oik.02520 |
| [6] |
宋同清, 彭晚霞, 曾馥平, 王克林, 覃文更, 谭卫宁, 刘璐, 杜虎, 鹿士杨. 木论喀斯特峰丛洼地森林群落空间格局及环境解释. 植物生态学报, 2010, 34(3): 298-308. DOI:10.3773/j.issn.1005-264x.2010.03.007 |
| [7] |
董雪, 杜昕, 孙志虎, 谷会岩, 陈祥伟. 生境梯度影响下的天然红松种群空间格局与种内关联. 生态学报, 2020, 40(15): 5239-5246. |
| [8] |
王鑫厅, 侯亚丽, 刘芳, 常英, 王炜, 梁存柱, 苗百岭. 羊草+大针茅草原退化群落优势种群空间点格局分析. 植物生态学报, 2011, 35(12): 1281-1289. |
| [9] |
张育新, 马克明, 祁建, 冯云, 张洁瑜. 北京东灵山海拔梯度上辽东栎种群结构和空间分布. 生态学报, 2009, 29(6): 2789-2796. DOI:10.3321/j.issn:1000-0933.2009.06.004 |
| [10] |
Wiegand T, Gunatilleke S, Gunatilleke N. Species associations in a heterogeneous Sri Lankan dipterocarp forest. The American Naturalist, 2007, 170(4): E77-E95. DOI:10.1086/521240 |
| [11] |
张娅娅, 刘旻霞, 李博文, 夏素娟, 孙瑞弟, 宋佳颖, 李全弟. 不同海拔矮嵩草与火绒草种群分布格局及空间关联性. 生态学杂志, 2020, 39(2): 404-411. |
| [12] |
张金屯. 植物种群空间分布的点格局分析. 植物生态学报, 1998, 22(4): 344-349. DOI:10.3321/j.issn:1005-264X.1998.04.008 |
| [13] |
许爱云, 刘金龙, 米世明, 朱宁宁, 于双, 许冬梅. 宁夏荒漠草原蒙古冰草(Agropyron mongolicum)种群小尺度空间分布及其关联性. 生态学报, 2019, 39(12): 4551-4559. |
| [14] |
拓锋, 刘贤德, 刘润红, 赵维俊, 敬文茂, 马剑, 武秀荣, 赵晶忠, 马雪娥. 祁连山大野口流域青海云杉种群空间格局及其关联性. 植物生态学报, 2020, 44(11): 1172-1183. DOI:10.17521/cjpe.2020.0177 |
| [15] |
赵成章, 高福元, 盛亚萍, 董小刚, 周伟. 狼毒种群小尺度空间分布格局及空间关联性研究. 干旱区地理, 2011, 34(3): 492-498. |
| [16] |
党晶晶, 赵成章, 任珩, 杨泉, 查高德. 高寒草地狼毒与阴山扁蓿豆种群的空间格局. 生态学杂志, 2013, 32(2): 292-298. |
| [17] |
池源, 石洪华, 王恩康, 郭振, 丰爱平, 麻德明. 庙岛群岛北五岛景观格局特征及其生态效应. 生态学报, 2017, 37(4): 1270-1285. |
| [18] |
王晓丽, 王嫒, 石洪华, 郑伟, 周然. 海岛陆地生态系统固碳估算方法. 生态学报, 2014, 34(1): 88-96. DOI:10.3969/j.issn.1673-1182.2014.01.019 |
| [19] |
刘超, 崔旺来, 朱正涛, 叶芳, 俞仙炯. 海岛生态保护红线划定技术方法. 生态学报, 2018, 38(23): 8564-8573. |
| [20] |
宁世江, 邓泽龙, 蒋运生. 广西海岛红树林资源的调查研究. 广西植物, 1995, 15(2): 139-145. |
| [21] |
金川, 王金旺, 郑坚, 陈秋夏, 李俊清, 卢翔. 异速生长法计算秋茄红树林生物量. 生态学报, 2012, 32(11): 3414-3422. |
| [22] |
张晓君, 管伟, 廖宝文, 朱宁华, 李琳, 何奕雄. 珠海人工红树林与天然红树林群落特征比较研究. 生态科学, 2014, 33(2): 321-326. |
| [23] |
李皓宇, 彭逸生, 刘嘉健, 王树功, 陈桂珠. 粤东沿海红树林物种组成与群落特征. 生态学报, 2016, 36(1): 252-260. |
| [24] |
成家隆, 吕瑜良, 陈玉军, 廖宝文, 丁冬静, 邓智泓, 魏军发. 水东湾红树林不同生长位置生长差异性研究. 生态科学, 2015, 34(6): 30-35. |
| [25] |
LY/T 2898-2017, 湿地生态系统定位观测技术规范[S]. 北京: 中国标准出版社, 2008.
|
| [26] |
李文良, 张小平, 郝朝运, 吴建勋, 王磊, 王静, 张姗姗, 程东升, 黄海明. 湘鄂皖连香树种群的年龄结构和点格局分析. 生态学报, 2009, 29(6): 3221-3230. DOI:10.3321/j.issn:1000-0933.2009.06.054 |
| [27] |
Clark P J, Evans F C. Distance to nearest neighbor as a measure of spatial relationships in populations. Ecology, 1954, 35(4): 445-453. DOI:10.2307/1931034 |
| [28] |
马志波, 肖文发, 黄清麟, 庄崇洋. 生态学中的点格局研究概况及其在国内的应用. 生态学报, 2017, 37(19): 6624-6632. |
| [29] |
Diggle P J. Statistical analysis of spatial and spatio-temporal point patterns. 3rd ed. Boca Raton: CRC Press, 2013.
|
| [30] |
Stoyan D, Stoyan H. Fractals, Random Shapes and Point Fields: Methods of Geometrical Statistics. New York: John Wiley & Sons, 1994. |
| [31] |
Illian J, Penttinen A, Stoyan H, Stoyan D. Statistical Analysis and Modelling of Spatial Point Patterns. Chichester, UK: John Wiley & Sons, 2008. |
| [32] |
王鑫厅, 侯亚丽, 梁存柱, 王炜, 刘芳. 基于不同零模型的点格局分析. 生物多样性, 2012, 20(2): 151-158. |
| [33] |
Rajala T, Illian J. A family of spatial biodiversity measures based on graphs. Environmental and Ecological Statistics, 2012, 19(4): 545-572. DOI:10.1007/s10651-012-0200-9 |
| [34] |
Wiegand T, Moloney K A. Handbook of Spatial Point-Pattern Analysis in Ecology. Boca Raton, FL, USA: CRC Press, 2014. |
| [35] |
Wiegand T, Moloney K A. Rings, circles, and null-models for point pattern analysis in ecology. Oikos, 2004, 104(2): 209-229. DOI:10.1111/j.0030-1299.2004.12497.x |
| [36] |
王友绍. 红树林生态系统评价与修复技术. 北京: 科学出版社, 2013.
|
| [37] |
梁士楚, 刘镜法, 梁铭忠. 北仑河口国家级自然保护区红树植物群落研究. 广西师范大学学报: 自然科学版, 2004, 22(2): 70-76. |
| [38] |
缪绅裕, 陈桂珠, 李海生. 红树林植物桐花树和白骨壤及其湿地系统. 广州: 中山大学出版社, 2007.
|
| [39] |
范航清, 黎广钊, 周浩郎. 广西北部湾典型海洋生态系统---现状与挑战. 北京: 科学出版社, 2015.
|
| [40] |
Condit R, Ashton P S, Baker P, Bunyavejchewin S, Gunatilleke S, Gunatilleke N, Hubbell S P, Foster R B, Itoh A, LaFrankie J V, Lee H S, Losos E, Manokaran N, Sukumar R, Yamakura T. Spatial patterns in the distribution of tropical tree species. Science, 2000, 288(5470): 1414-1418. DOI:10.1126/science.288.5470.1414 |
| [41] |
李明顺, 蓝崇钰, 陈桂珠, 李世华, 陈相如, 刘治平, 谭凤仪, 黄玉山. 深圳福田红树林的群落学研究Ⅱ. 多样性与种群格局. 生态科学, 1994, 12(1): 82-86. |
| [42] |
邱霓, 徐颂军, 邱彭华, 杨文槐, 杨秀, 杨倩. 珠海淇澳岛红树林群落分布与景观格局. 林业科学, 2019, 55(1): 1-10. |
| [43] |
邱霓, 徐颂军, 邱彭华, 宋焱, 牛安逸, 许观嫦. 南沙湿地公园红树林物种多样性与空间分布格局. 生态环境学报, 2017, 26(1): 27-35. |
| [44] |
缪绅裕, 陈桂珠, 陈正桃, 吴中亨. 广东湛江保护区红树林种群的生物量及其分布格局. 广西植物, 1998, 18(1): 19-23. |
| [45] |
张育霞, 吕晓波, 杨小波, 李东海, 李英英, 罗文启, 熊梦辉, 邢莎莎, 朱静, 林泽钦, 岑举人, 郭照彬. 海南东寨港不同潮间带角果木种群结构及分布格局. 热带生物学报, 2015, 6(4): 388-396. |
| [46] |
何东进, 肖石红, 李晓景, 王韧, 蔡金标, 游巍斌. 中国红树林北缘区秋茄树种群空间分布格局和结构特征分析. 植物资源与环境学报, 2014, 23(2): 41-47. |
| [47] |
黄小, 朱江, 姚兰, 艾训儒, 王进, 吴漫玲, 朱强, 陈绍林. 水杉原生种群结构及空间分布格局. 生物多样性, 2020, 28(4): 463-473. |
| [48] |
Snedarker S C, Snedarker J G. 红树林生态系统研究方法. 郑德璋, 郑松发, 廖宝文, 译. 广州: 广东科技出版社, 1994.
|
| [49] |
樊登星, 余新晓. 北京山区栓皮栎林优势种群点格局分析. 生态学报, 2016, 36(2): 318-325. |
| [50] |
梁爽, 许涵, 林家怡, 李意德, 林明献. 尖峰岭热带山地雨林优势树种白颜树空间分布格局. 植物生态学报, 2014, 38(12): 1273-1282. |