 2022, Vol. 42
2022, Vol. 42文章信息
- 刘文倩, 李家湘, 龚俊伟, 赵丽娟, 项文化, 程幸, 王维嘉, 张意灵
- LIU Wenqian, LI Jiaxiang, GONG Junwei, ZHAO Lijuan, XIANG Wenhua, CHENG Xing, WANG Weijia, ZHANG Yiling
- 柯-青冈常绿阔叶林优势树种叶片性状变异及适应策略
- Variation in leaf functional traits and adaptation strategies of dominant tree species in a Lithocarpus glaber-Cyclobalanopsis glauca evergreen broad-leaved forest
- 生态学报. 2022, 42(17): 7256-7265
- Acta Ecologica Sinica. 2022, 42(17): 7256-7265
- http://dx.doi.org/10.5846/stxb202103100659
-
文章历史
- 收稿日期: 2021-03-10
- 网络出版日期: 2022-04-03





2. 中南林业科技大学生命科学与技术学院, 长沙 410004
2. College of Science and Technology, Central South University of Forestry and Technology, Changsha 410004, China
植物功能性状是植物在长期进化过程中与环境相互适应形成的, 在形态、生理、物候等方面可度量和描述的植物性状[1-2]。作为植物光合、呼吸、蒸腾作用的最重要器官[3], 叶片性状能够反映植物对环境变化的响应及适应策略[4]。叶面积、比叶面积、叶厚度、叶片干物质含量等结构性状和叶片碳、氮和磷含量等化学性状可表征植物碳同化、防御和养分周转能力, 可指示植物在不同环境下的资源利用效率和生存适应策略, 已成为揭示植物生态适应策略和群落对全球变化响应的重要途径[5]。Wright等对全球175个地区2548种植物叶片性状研究表明, 叶片的形态结构、化学组分和生理过程之间存在相互联系、互相依存的内在关系, 提出著名的叶片经济型谱(leaf economics spectrum, LES), 认为“缓慢投资-收益”型物种的资源利用成本高、叶片寿命长、氮含量低且光合速率慢, 而“快速投资-收益”型物种的构建成本低、寿命短、氮含量高且光合速率快[6-7]。LES能够解释不同类型植物根据其功能需求在不同生境条件下的资源权衡配置, 体现了叶片结构、化学和生理性状之间的权衡关系[8], 已广泛应用于叶片功能性状与环境间的关系[9-11]及群落构建机制研究等诸多方面[12-13], 但对于其在局域群落中的适用性, 仍存在争议[8]。
植物功能性状差异将物种适应策略与群落生态系统功能和过程有机联系, 其变异的大小因种类而异, 并与环境梯度密切相关[14-15]。区域尺度下, 环境梯度引起群落物种组成差异, 性状的种间变异可以很好地解释不同群落环境条件下的物种共存机制[16]。但在局域尺度, 环境异质性不高, 物种通过增加个体间表型差异来增强对有限资源的竞争力, 性状的种内变异对于物种共存则更为重要[14-16]。有研究发现性状的种内变异解释了30%以上的群落性状变异[17], 不同群落内功能性状的种间和种内差异贡献相差甚远;Laforest-Lapointe等[18]基于全球性状数据库的分析表明, 种内性状变异是影响气候等环境变化的决定因素, 其作用随生境的贫乏程度而增强。个体是群落构建的基本单位, 功能多样性概念的出发点也强调个体水平差异的重要性[19]。同时, 局域群落将环境因子和功能性状置于同一系统中, 定量群落内植物功能性状的种内、种间变异, 将有助于群落功能性状分异的潜在来源和解释植物对环境变化的适应性机制。
常绿阔叶林是我国亚热带湿润地区特有的生态系统, 在生物多样性保护和维持碳平衡方面具有重要作用[20-22]。由于长期而频繁的人类干扰, 植被退化严重, 生态问题凸显, 基于地带性植被典型群落结构和生态过程的研究已成为植被恢复和重建的重要手段[23]。柯(Lithocarpus glaber)-青冈(Cyclobalanopsis glauca)林是亚热带最常见常绿阔叶林类型之一, 主要分布于我国亚热带海拔600 m以下的山地、丘陵区, 处于演替后期阶段, 结构稳定, 生态系统功能较高, 代表着区域内植被的恢复方向[24]。本研究利用长沙县大山冲林场1 hm2的柯-青冈常绿阔叶林固定监测样地, 系统调查乔木层优势树种叶片结构和化学性状, 旨在回答: (1)群落中树木叶片功能性状变异来源是什么?(2)叶片功能性状之间存在怎样的权衡关系?结果可为理解森林群落物种的环境适应策略、预测群落动态变化和制定植被恢复措施提供科学依据。
1 研究区域概况研究样地位于湖南省长沙县大山冲国有林场(113°17′46″-113°19′8″ E, 28°23′58″-28°24′58″ N), 低山丘陵地貌, 与幕阜山余脉西缘接壤, 海拔55-217.40 m, 坡度20-30°;属亚热带东南季风湿润气候区, 年均气温16.50-17.60 ℃, 年日照1300-1800 h, 年均降水量1420.00 mm, 全年无霜期216-269 d;土壤以板岩和页岩等变质岩发育而成的酸性黏土质红壤为主[22]。地带性植被为亚热带常绿阔叶林, 经过60-70 a的封山育林, 该林场现存植被多为次生林和人工林, 其中柯-青冈林为研究区内残存且保存最为完好的常绿阔叶林类型[25]。
2 调查和研究方法 2.1 叶片样品采集依托2009年建立的柯-青冈常绿阔叶林1 hm2固定样地, 对胸径大于等于1.00 cm的所有个体挂牌监测, 选择重要值排前6位的优势树种为对象:柯、青冈、马尾松(Pinus massoniana)、红淡比(Cleyera japonica)、杉木(Cunninghamia laceolata)和南酸枣(Choerospondias axillaris)(表 1)。叶片取样参考Pérez-Harguindeguy等[26]的方法, 将样地划分为100个10 m×10 m的样方, 以样方为单位对优势种采样, 每种确定1-3株标准木, 随机选取树冠外层东、西、南、北方位上4个无病虫害的末端当年枝, 用沾水滤纸包好及时放入自封袋贴好标签带回实验室, 摘取10-20片健康完整叶片用于测定叶片性状。6个优势种共确定278株标准木, 采集了4448个叶片。
| 物种Species | 缩写 Abbreviation |
生活型 Life form |
重要值 Important value/% |
平均胸径±标准误 Average DBH±SE/cm |
平均树高±标准误 Average tree height±SE/m |
采样株数 Sample individuals |
| 柯Lithocarpus glaber | LG | 常绿阔叶 | 21.78 | 7.16±0.23 | 6.75±0.17 | 88 |
| 青冈Cyclobalanopsis glauca | CG | 常绿阔叶 | 12.02 | 7.00±0.44 | 6.29±0.27 | 75 |
| 马尾松Pinus massoniana | PM | 常绿针叶 | 10.60 | 21.32±1.73 | 14.48±0.51 | 15 |
| 红淡比Cleyera japonica | CJ | 常绿阔叶 | 9.33 | 4.43±0.14 | 5.03±0.12 | 60 |
| 南酸枣Choerospondias axillaris | CA | 落叶阔叶 | 6.39 | 22.15±1.02 | 15.74±0.66 | 14 |
| 杉木Cunninghamia lanceolata | CL | 常绿针叶 | 4.45 | 8.50±0.61 | 8.19±0.53 | 26 |
| DBH:胸径Diameter at Breast Height;SE:标准误差Standard Error | ||||||
本研究选择8个叶片性状指标进行分析, 包括叶片厚度(LT, mm)、叶片面积(LA, mm2)、叶片干物质含量(LDMC, mg/g)、比叶面积(SLA, cm2/g)等结构性状和叶片碳含量(LC, mg/g)、叶片氮含量(LN, mg/g)、叶片磷含量(LP, mg/g)、叶片碳氮比(LC ∶ LN)等化学性状, 对这些性状进行描述性统计(表 2)。
| 功能性状Functional traits | PM | CL | LG | CG | CJ | CA |
| LA/(mm2) | 187.67±42.63d(0.39) | 243.13±54.42d(0.20) | 2789.30±96.30b(0.30) | 3268.89±104.10a(0.24) | 1896.39±85.46c(0.31) | 2554.41±277.78bc(0.22) |
| LT/(mm) | 0.44±0.13a(0.36) | 0.25±0.06bc(0.39) | 0.26±0.07bc(0.14) | 0.27±0.02b(0.52) | 0.27±0.01bc(0.14) | 0.16±0.02c(0.22) |
| SLA/(cm2/g) | 15.57±5.94b(0.23) | 40.76±14.03d(0.32) | 116.47±18.5bc(0.16) | 104.53±17.29bc(0.17) | 132.84±23.73b(0.18) | 195.99±106.80a(0.55) |
| LDMC/(mg/g) | 608.75±211.84ab(0.25) | 601.90±55.53ab(0.47) | 705.48±22.59a(0.28) | 691.83±20.27a(0.22) | 479.32±24.66b(0.36) | 499.53±58.02ab(0.23) |
| LC/(mg/g) | 475.48±5.05a (0.02) | 472.52±2.87a (0.03) | 479.20±1.15a (0.03) | 471.96±1.96a (0.03) | 447.16±3.49b (0.05) | 461.06±7.67ab (0.03) |
| LN/(mg/g) | 13.23±0.74d (0.11) | 15.09±0.53cd (0.18) | 16.66±0.22b (0.11) | 16.18±0.23bc (0.11) | 14.56±0.28d (0.13) | 21.86±2.28a (0.21) |
| LP/(mg/g) | 1.05±0.19ab (0.32) | 1.19±0.13a (0.31) | 0.74±0.03b (0.31) | 0.77±0.02b (0.22) | 0.66±0.02b (0.24) | 1.22±0.41a (0.59) |
| LC∶LN | 36.33±2.28a (0.13) | 32.24±1.11a (0.18) | 29.06±0.34c (0.10) | 29.44±0.35bc (0.09) | 31.07±0.47ab (0.10) | 21.82±2.34d (0.21) |
| 不同字母代表不同树种间差异显著(P < 0.05);括号内数字代表性状的种内变异系数;LA:叶面积leaf area;LT:叶片厚度leaf thicknes;SLA:比叶面积specific leaf area;LDMC:叶片干物质含量leaf dry matter content;LN:LC:叶碳含量leaf carbon content;叶氮含量leaf nitrogen content;LP:叶磷含量leaf phosphorus content;LC ∶ LN:碳氮比leaf carbon-nitrogen ratio | ||||||
阔叶树利用游标卡尺(精确至0.01 mm)测定靠近主脉的前、中、末端的厚度值5个, 取平均值作为叶片厚度;针叶树则利用游标卡尺测定针叶中部的垂直厚度[27]。随后, 平铺叶片并保证叶片完全展开, 用扫描仪(HP Officejet ProX576dw MFP)扫描获取平面图像。利用Adobe Photoshop CS 6软件[28]对阔叶树种每个叶片所占像素点进行统计, 取平均值作为叶面积;针叶树种测定叶长(LL)、中间部叶宽(LW), 利用公式 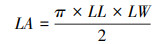
将测定完叶片面积和干物质含量的样品先在105℃下杀青5 min, 再85℃烘干至恒重, 研磨并过100目筛。采用重铬酸钾加热法测定叶片碳含量, 凯氏定氮法测定叶片氮含量, 钼锑抗比色法测定叶片磷含量[29]。所有元素测定均同时做2组平行试验与2个空白, 以减少试验误差, 随后取平均值作为该样方该物种叶片碳、氮和磷含量测定值, 同时计算LC ∶ LN值。
2.3 数据分析统计分析时, 对性状值进行log转化使其满足正态分布或接近正态。首先基于描述性统计分析6个优势种叶片性状的种内变异, 利用LSD多重比较分析叶片功能性状的种间和生活型(常绿阔叶、落叶阔叶和常绿针叶)间差异[19]。然后, 利用混合效应模型[30]解析群落叶片功能性状变异的主要来源, 将生活型设为固定效应, 种间及种内变异作为随机效应, 量化生活型间、种间及种内功能变异的贡献率。最后, 在不区分树种和生活型的情况下, 使用Pearson′s相关分析检验性状间的相关性;利用主成分分析(PCA)揭示性状间的协变关系, 通常第一主成分(PC1)和第二主成分(PC2)能解释较高比例的方差, 分析计算了PC1和PC2载荷量与每个性状的相关系数, 用以分析单个性状和植物经济策略的变异模式。所有数据分析通过R 3.5.2语言“Lme4”、“piecewiseSEM”、“vegan”包进行[31-32]。
3 结果与分析 3.1 叶片性状的变异群落优势种叶片功能性状种内、种间差异显著, 变异中等(CV : 0.02-0.59), 结构性状变异程度普遍高于化学性状(表 2)。其中, 马尾松拥有最大的LT和LC ∶ LN, 以及最小的LN、LA、SLA;南酸枣则反之, LT、LC ∶ LN最小, LN、LP、SLA最大;柯的LC最大, 红淡比最小, 与马尾松、杉木、青冈、柯之间差异显著;LDMC除在红淡比中最低外, 其他树种间无显著差异(P < 0.05)。在生活型水平上, 针叶树种的LA、SLA显著低于阔叶树种, 且常绿阔叶树种的SLA低于落叶阔叶树种;常绿树种的LC和LDMC最小, 而落叶树种拥有最大的SLA、LN和LP, 但LT和LC ∶ LN最小。
3.2 叶片性状变异的来源混合效应模型分析可知, 生活型间、种间和种内变异对群落叶片功能性状变异的贡献不同(图 1)。生活型对多数性状的贡献率较大, 其中LDMC、LC ∶ LN分别有93.11%和91.76%的变异来源于生活型差异, 而种间与种内变异影响极小;LA、LC、LN和LP来源于生活型的变异亦均在70.00%以上。同时, 6个优势种的8个植物叶片性状的种间变异贡献变化幅度较大(0.00%-80.00%), 结构性状的种间变异(LDMC除外)均高于化学性状变异, 而种内变异对LP的贡献率较高(23.66%)。

|
| 图 1 生活型、种间及种内变异对群落叶片功能性状的相对贡献 Fig. 1 Relative contribution of life form, interspecific and intraspecific variation to various leaf functional traits variation LA:叶面积;LT:叶片厚度;SLA:比叶面积;LDMC:叶片干物质含量;LN:叶氮含量;LC:叶碳含量;LP:叶磷含量;LC ∶ LN:碳氮比 |
多数叶片功能性状间存在显著相关关系(表 3)。SLA与LDMC、LT显著负相关, 而LA与LDMC、LN、LC呈显著正相关。LC与SLA呈显著负相关(P < 0.01), 与LDMC和LN呈显著正相关(P < 0.001)。LP除与LN呈显著正相关(P < 0.05), 与LA显著负相关(P < 0.001)外, 与其他性状无显著关系;LC ∶ LN与LA、SLA、LN和LC呈显著负相关(P < 0.001), 与LT显著正相关(P < 0.001)。LN、LC和LA之间呈显著正相关, 与LC ∶ LN显著负相关。
| 性状Traits | LA/ (mm2) |
LT/ mm |
SLA/ (cm2/g) |
LDMC/ (mg/g) |
LC/ (mg/g) |
LN/ (mg/g) |
LP/ (mg/g) |
| LT/mm | -0.04 | ||||||
| SLA/(cm2/g) | 0.24*** | -0.23*** | |||||
| LDMC/(mg/g) | 0.15* | 0.01 | -0.24*** | ||||
| LC/(mg/g) | 0.19* | -0.07 | -0.24*** | 0.33*** | |||
| LN/(mg/g) | 0.25*** | -0.20** | -0.24*** | 0.06 | 0.58*** | ||
| LP/(mg/g) | -0.15*** | -0.13 | -0.13 | 0.03 | 0.05 | 0.18* | |
| LC∶LN | -0.24*** | 0.19*** | -0.32*** | 0.05 | -0.32*** | -0.94*** | -0.13 |
| *, P < 0.05;* *, P < 0.01;* * *, P < 0.001 | |||||||
主成分PC1和PC2可解释8个叶片功能性状54.69%的变异。其中第一主成分与LN、LT、LC ∶ LN相关性较高;而第二主成分则与LP、LDMC相关性较高;LC、SLA和LA与两个主成分都存在显著相关性(图 2)。落叶阔叶树种南酸枣位于第一象限, 有着较高的LN、SLA、LP和较低的LT、LC ∶ LN、LDMC;沿着对角线依次为常绿阔叶树种柯和青冈;最左下端为常绿针叶树种杉木和马尾松, 具有较高的LMDC、LT、LP、LC ∶ LN和较低的LA、SLA;而常绿小乔木红淡比聚集于第二象限, 具有较高的SLA、LT和LC ∶ LN和较低的LC、LN、LDMC(图 2)。

|
| 图 2 柯-青冈常绿阔叶林叶片功能性状的主成分分析 Fig. 2 Principal component analysis of leaf functional traits species in L. glaber-C. glauca evergreen broad-leaved forest |
亚热带柯-青冈常绿阔叶林群落6个优势树种的8个叶片功能性状平均值均在全球叶片性状值变化范围内[33-34]。性状变异程度中等, 生活型(平均65.62%, 最大93.11%)、种间(平均23.31%, 最大79.60%)和种内变异(平均9.10%, 最大23.66%)均有显著贡献。结果与众多学者的观点一致[8], 植物功能性状的分异主要来源于功能型和种间变异, 种内变异的作用也不容忽视, 说明精确度量功能性状的多源变异对深入认识群落构建过程的意义重大[35-36]。与其它植被类型相比较[37-38], 该群落的种内变异贡献率较低, 可能与研究区位于低山、丘陵地区, 群落内地形、土壤等环境梯度变化较小[24]有关, 结果支持“性状空间变异分割假说”, 即在局域尺度上因环境异质性和个体数量的限制, 种内变异的重要性较小[39]。
植物叶片功能性状受遗传因素和环境因素共同决定, 通过种内变异呈现生态策略的趋异性, 从而产生生态位的分化减小竞争强度[38]。柯-青冈常绿阔叶林群落中, 优势树种叶片化学性状(LC、LN、LP、LC ∶ LN)的种内变异高于结构性状(LA、SLA、LDMC), 而种间变异则表现出相反趋势(表 2), 说明叶片化学性状变异在应对局域环境变化上具有较高可塑性。赵丽娟[24]研究发现, 柯-青冈监测样地内土壤和地形因子具有中等程度的空间变异, 对群落物种组成和空间分布影响显著。众所周知, 土壤中氮、磷可获得性是叶片氮、磷含量主要影响因素[37], 优势种为了适应较为复杂的环境条件, 通过调整化学性状可塑性以增加其沿环境梯度的分布空间。相比较而言, 结构性状与生活型、物种差异有着密切关联且较为稳定, 种间变异较大, 可能与遗传因素有关[34]。众多研究发现[39], 不同系统发育阶段的植物叶片大小、叶片寿命、叶氮含量以及光合能力上存在很大差异, 本研究中的6个优势种分属于壳斗科(Fagaceae)、山茶科(Theaceae)、漆树科(Anacardiaceae)、松科(Pinaceae)和杉科(Taxodiaceae), 其显著差异的系统发育背景[41]可能通过功能性状协调而有助于植物在多变环境中生存。
植物生活型是叶片功能性状的主要决定因子之一, 以往大量研究已发现不同生活型植物叶片功能性状存在显著差异[37, 40-41]。本研究中, 常绿阔叶树种的种内变异小于针叶树种和落叶阔叶树种(表 2), 可能由于针叶树种和落叶阔叶树种为最先定居该区域的先锋树种, 如散生于样地中的马尾松、南酸枣等, 种群正处于衰退阶段, 为实现最大限度地存活, 通过增加LA、SLA、LP等性状的可塑性以应对群落内光资源和土壤养分的限制[8]。同时, 与落叶树种相比, 常绿树种LDMC、LT、LA和LC种间变异较大, 可能不同生活型叶片构建成本以及季相变化有关;针叶树种拥有最低的LA及较高的SLA、LC和LC ∶ LN, 与唐青青等[40]的研究结果一致。同时, 除LT和SLA外, 其余性状的变异都主要来源于生活型差异(图 1), 说明不同生活型植物的物种特性对养分利用策略和干物质存储的影响较大[42]。
4.2 叶片功能性状权衡关系研究结果表明柯-青冈常绿阔叶林叶片结构性状与化学性状间存在相关关系(表 3)。SLA、LDMC、LN等都反映植物对环境的适应策略[6, 8, 43], SLA、LN显著正相关, 说明SLA越高, 光合能力越强, 光合产物积累越多, 养分循环越快;然而, LT和LDMC是与保护、防御相关的功能性状, SLA与两者均呈显著负相关, 而LMDC与LT之间无显著相关性, 反映出柯-青冈常绿阔叶林SLA主要受叶片厚度和干物质含量的影响。叶片防御结构的构建消耗巨大, 植物需长时间的光合作用形成LC以补偿构建消耗, 表现为LC随着LDMC、LN的增加而升高, 结果与许多研究相一致[44-45]。同时, 光照是影响SLA的重要因子[46], 水分则是影响LDMC的关键[47], 而本研究发现LN和LC ∶ LN与SLA的相关性大于LDMC, 因此推断柯-青冈群落内光资源可获得性较水分梯度对植物生长有更重要的影响。
同时, 植物器官的养分含量是植物与环境相互作用的结果, 可以反映在一定环境条件下植物吸收营养元素的能力[43]。研究发现LN随LA的增加而显著增加, 而LP则呈降低趋势, 且LP与其他结构性状间没有显著相关关系(表 3)。这种截然不同的关系表明植物在氮、磷利用策略上差异显著, 与环境中氮、磷来源和丰富度紧密相关。植物器官中氮主要来源于大气和土壤中的硝酸盐, 而磷主要来源于土壤矿化, 研究区为亚热带丘陵红壤区, N沉降已使得生态系统达到N饱和状态[48-49], 强烈的淋溶使得区域内磷可获得性处于较低水平, 磷成为了区域内植物生长的限制因子[37, 50]。因此, 在柯-青冈群落中, LP具有较高的种内变异(表 2), 与LN通过显著正相关关系来维持化学内稳态, 并降低LA来抵御不利环境;与SLA、LT、LMDC等无显著相关关系可能因较高的种内变异贡献率削弱了种间性状的相关性[51], 该结果与Keenan和Niinemets在全球尺度上[52]和荀彦涵等在局域尺度上[8]的研究结果不一致, 可能与研究的树种不同有关。
4.3 亚热带树种叶片功能性状的适应策略叶片经济型谱将植物排列在特定的生态轴上, 一端代表着比叶面积大、光合和呼吸速率快且叶片寿命短的快速-投资收益型策略, 另一端代表着与上述特征相反的缓慢-投资收益型策略[6]。本研究发现, 性状变异第一主成分指示叶经济型谱轴, 与LT、LA、SLA、LN、LC、LC ∶ LN有着较强的相关性, 主要反映物种生态策略从资源保守型到资源获取型的转变(图 2)。树种在叶经济型谱轴两侧的差异性分布, 说明柯-青冈常绿阔叶林内物种生态策略的显著分化。
针叶树和阔叶树聚集在PCA排序图的不同区域, 尤其在LA、SLA、LN、LP等性状上表现出差异显著(图 2), 反映了两类树种在环境适应和资源利用策略上的内在差异。针叶树种马尾松和杉木集中分布于第三象限, 两者表现出类似的叶片性状特征。其中马尾松在群落中具有LT(0.44±0.13)最大、LC ∶ LN(36.33±2.28)最高, 而LA(187.67±42.63)和SLA(15.57±5.94)最小、LN最低的性状特征, 这与马尾松为先锋树种, 最先入驻该群落, 生于相对瘠薄的山脊线, 将叶肉内大部分物质用于储存和防御, 或增加叶肉细胞密度, 采取相对保守的生存策略。而阔叶树种中, 落叶树种南酸枣, 叶片寿命短, 通常将干物质更多地投入到植物本身和叶面积的增长, 具有最小的LT(0.16±0.02)、LC ∶ LN(21.82±2.34)和最高的SLA(195.99±106.80)、LN(21.86±2.28)和LP(1.22±0.41), 属于“快速-投资-收益”型物种;红淡比具有最小的LDMC(479.32±24.66)、LC(447.16±3.49)、LN(14.56±0.28)、LP(0.66±0.02), 最大的LC ∶ LN(31.07±0.47), 说明由于其为常绿小乔木, 树冠很难到达林冠外层(表 1), 但耐荫, 能够通过提高LC ∶ LN即光合效能来适应林内光资源的不足, 属于“缓慢-投资-收益”型树种。常绿阔叶乔木青冈和柯位于林冠, 光照强烈, 将更多资源倾向于保卫组织建立或叶肉密度增强, 以抵御高温对叶片损伤和叶片失水, 提高碳氮固持效率, 因此具有较高的LT、LDMC、LA和低的LP、SLA, 属于中间偏“缓慢投资-收益”型树种。总之, 群落中落叶树种的SLA显著大于常绿树种, 阔叶树种的SLA显著大于针叶树种, 这与中国森林[53]和亚热带常绿落叶阔叶混交林叶片功能性状的研究结果一致[21], 群落中不同树种尤其是不同生活型的树种采用独特的生长策略以适应外界环境变化和利用有限的资源[54]。
同时, 不仅针叶树种马尾松和杉木表现出类似的叶片性状, 亲缘关系近的常绿阔叶树种青冈与柯的叶片性状尤其是化学性状LN、LP、LC ∶ LN亦表现出相同趋势, 且在叶经济型谱轴上的分布表现出一定程度的重叠, 均采取较高LT、LDMC、LA和低LP、SLA生存策略, 表明在长期进化过程中, 亲缘关系较近的物种会通过调节自身养分含量及其比值, 形成类似的化学计量权衡策略来适应群落环境, 在性状进化上表现出保守性和性状间的协同性。而Zhao等[55]的研究发现青冈和柯在样地呈现出差异的空间聚集, 其中青冈集中分布于阴坡最陡峭处, 而柯则分布于向阳坡、土层相对较厚的地段, 两者间在空间分布上存在显著的生态位分化。可见, 这种生态策略及空间分布的差异性降低了种间竞争强度, 从而有利于两个性状相似的优势种共存。
4.4 结论柯-青冈群落优势树种叶片功能性状存在显著的生活型间、种间和种内变异, 结构性状的可塑性较化学性状保守, 总体上呈现中等程度变异格局, 符合“性状空间变异分割假说”。本研究证实了生活型间和种间变异是群落叶片性状变化的重要来源, 但种内变异的贡献也不可忽视。同时, 叶经济型谱在亚热带常绿阔叶林局域群落中同样适用, 不同树种通过调整叶片性状间权衡关系来适应环境变化, 从而实现群落物种共存。
| [1] |
刘晓娟, 马克平. 植物功能性状研究进展. 中国科学: 生命科学, 2015, 45(4): 325-339. |
| [2] |
赖媛, 张玲玲, 郭英荣, 雷平, 任文丹, 刘占锋, 旷远文. 武夷山不同海拔杜鹃和黄山松叶片功能性状的变化. 生态学杂志, 2021, 40(7): 1988-1996. |
| [3] |
冯家宝, 范顺祥, 侯煜飞, 剧永望, 马天蕊, 高英月, 张志东. 河北省森林草原区草本植物叶功能性状种间和种内变异. 东北林业大学学报, 2021, 49(1): 23-28. |
| [4] |
Green D S, Kruger E L. Light-mediated constraints on leaf function correlate with leaf structure among deciduous and evergreen tree species. Tree Physiology, 2001, 21(18): 1341-1346. DOI:10.1093/treephys/21.18.1341 |
| [5] |
盘远方, 陈兴彬, 姜勇, 梁士楚, 陆志任, 黄宇欣, 倪鸣源, 覃彩丽, 刘润红. 桂林岩溶石山灌丛植物叶功能性状和土壤因子对坡向的响应. 生态学报, 2018, 38(5): 1581-1589. |
| [6] |
Wright I J, Reich P B, Westoby M, Ackerly D D, Baruch Z, Bongers F, Cavender-Bares J, Chapin T, Cornelissen J H C, Diemer M, Flexas J, Garnier E, Groom P K, Gulias J, Hikosaka K, Lamont B B, Lee T, Lee W, Lusk C, Midgley J J, Navas M L, Niinemets V, Oleksyn J, Osada N, Poorter H, Poot P, Prior L, Pyankov V I, Roumet C, Thomas S C, Tjoelker M G, Veneklaas E J, Villar R. The worldwide leaf economics spectrum. Nature, 2004, 428(6985): 821-827. DOI:10.1038/nature02403 |
| [7] |
Reich P B. The world-wide 'fast-slow' plant economics spectrum: a traits manifesto. Journal of Ecology, 2014, 102(2): 275-301. DOI:10.1111/1365-2745.12211 |
| [8] |
荀彦涵, 邸雪颖, 金光泽. 典型阔叶红松林主要树种叶性状的垂直变异及经济策略. 植物生态学报, 2020, 44(7): 730-741. |
| [9] |
Maire V, Wright I J, Prentice I C, Batjes N H, Bhaskar R, van Bodegom P M, Cornwell W K, Ellsworth D, Niinemets V, Ordonez A, Reich P B, Santiago L S. Global effects of soil and climate on leaf photosynthetic traits and rates. Global Ecology and Biogeography, 2015, 24(6): 706-717. DOI:10.1111/geb.12296 |
| [10] |
Vergara-Gómez D, Williams-Linera G, Casanoves F. Leaf functional traits vary within and across tree species in tropical cloud forest on rock outcrop versus volcanic soil. Journal of Vegetation Science, 2020, 31(1): 129-138. DOI:10.1111/jvs.12826 |
| [11] |
Liu X R, Chen H X, Sun T Y, Li D Y, Wang X, Mo W Y, Wang R L, Zhang S X. Variation in woody leaf anatomical traits along the altitudinal gradient in Taibai Mountain, China. Global Ecology and Conservation, 2021, 26. |
| [12] |
Wang X, Song N P, Yang X G, Chen L, Qu W J, Wang L. Inferring community assembly mechanisms from functional and phylogenetic diversity: the relative contribution of environmental filtering decreases along a sand desertification gradient in a desert steppe community. Land Degradation & Development, 2021, 32(7): 2360-2370. |
| [13] |
Zhang J X, Swenson N G, Liu J M, Liu M T, Qiao X J, Jiang M X. A phylogenetic and trait-based analysis of community assembly in a subtropical forest in central China. Ecology and Evolution, 2020, 10(15): 8091-8104. DOI:10.1002/ece3.6465 |
| [14] |
Reich P B, Wright I J, Cavender-Bares J, Craine J M, Oleksyn J, Westoby M, Walters M B. The evolution of plant functional variation: traits, spectra, and strategies. International Journal of Plant Sciences, 2003, 164(S3): S143-S164. DOI:10.1086/374368 |
| [15] |
李耀琪, 王志恒. 植物叶片形态的生态功能、地理分布与成因. 植物生态学报, 2021, 45(10): 1154-1172. |
| [16] |
Díaz S, Kattge J, Cornelissen J H C, Wright I J, Lavorel S, Dray S, Reu B, Kleyer M, Wirth C, Colin Prentice I, Garnier E, BÖnisch G, Westoby M, Poorter H, Reich P B, Moles A T, Dickie J, Gillison A N, Zanne A E, Chave J, Joseph Wright S, Sheremet'ev S N, Jactel H, Baraloto C, Cerabolini B, Pierce S, Shipley B, Kirkup D, Casanoves F, Joswig J S, Günther A, Falczuk V, Rüger N, Mahecha M D, Gorné L D. The global spectrum of plant form and function. Nature, 2016, 529(7585): 167-171. DOI:10.1038/nature16489 |
| [17] |
周刘丽. 浙江天童木本植物功能性状种内变异的大小、来源和空间结构[D]. 上海: 华东师范大学, 2016.
|
| [18] |
Laforest-Lapointe I, Martínez-Vilalta J, Retana J. Intraspecific variability in functional traits matters: case study of Scots pine. Oecologia, 2014, 175(4): 1337-1348. DOI:10.1007/s00442-014-2967-x |
| [19] |
许驭丹, 董世魁, 李帅, 沈豪. 植物群落构建的生态过滤机制研究进展. 生态学报, 2019, 39(7): 2267-2281. |
| [20] |
吴征镒. 中国植被. 北京: 科学出版社, 1980.
|
| [21] |
宋永昌. 植被生态学. 上海: 华东师范大学出版社, 2001.
|
| [22] |
陈灵芝, 陈伟烈, 韩兴国. 中国退化生态系统研究. 北京: 中国科学技术出版, 1995.
|
| [23] |
Ledo A, Burslem D F R P, Condés S, Montes F. Micro-scale habitat associations of woody plants in a neotropical cloud forest. Journal of Vegetation Science, 2013, 24(6): 1086-1097. DOI:10.1111/jvs.12023 |
| [24] |
赵丽娟. 中亚热带石栎-青冈群落结构特征及影响因子研究[D]. 长沙: 中南林业科技大学, 2014.
|
| [25] |
赵丽娟, 项文化, 李家湘, 邓湘雯, 刘聪. 中亚热带石栎-青冈群落物种组成、结构及区系特征. 林业科学, 2013, 49(12): 10-17. |
| [26] |
Pérez-Harguindeguy N, Díaz S, Garnier E, Lavorel S, Poorter H, Jaureguiberry P, Bret-Harte M S, Cornwell W K, Craine J M, Gurvich D E, Urcelay C, Veneklaas E J, Reich P B, Poorter L, Wright I J, Ray P, Enrico L, Pausas J G, de Vos A C, Buchmann N, Funes G, Quétier F, Hodgson J G, Thompson K, Morgan H D, ter Steege H, Sack L, Blonder B, Poschlod P, Vaieretti M V, Conti G, Staver A C, Aquino S, Cornelissen J H C. New handbook for standardised measurement of plant functional traits worldwide. Australian Journal of Botany, 2013, 61(3): 167. DOI:10.1071/BT12225 |
| [27] |
肖强, 叶文景, 朱珠, 陈瑶, 郑海雷. 利用数码相机和Photoshop软件非破坏性测定叶面积的简便方法. 生态学杂志, 2005, 24(6): 711-714. DOI:10.3321/j.issn:1000-4890.2005.06.026 |
| [28] |
Bolker B M, Brooks M E, Clark C J, Geange S W, Poulsen J R, Stevens M H H, White J S S. Generalized linear mixed models: a practical guide for ecology and evolution. Trends in Ecology & Evolution, 2009, 24(3): 127-135. |
| [29] |
Borcard D, Gillet F, Legender P. 数量生态学: R语言的应用(第二版). 赖江山, 译. 北京: 高等教育出版社, 2020.
|
| [30] |
赵广帅, 刘珉, 石培礼, 宗宁, 张鑫, 张宪洲. 羌塘高原降水梯度植物叶片、根系性状变异和生态适应对策. 生态学报, 2020, 40(1): 295-309. |
| [31] |
陈文, 王桔红, 马瑞君, 齐威, 刘坤, 张丽娜, 陈学林. 粤东89种常见植物叶功能性状变异特征. 生态学杂志, 2016, 35(8): 2101-2109. |
| [32] |
Albert C H, Thuiller W, Yoccoz N G, Soudant A, Boucher F, Saccone P, Lavorel S. Intraspecific functional variability: extent, structure and sources of variation. Journal of Ecology, 2010, 98(3): 604-613. DOI:10.1111/j.1365-2745.2010.01651.x |
| [33] |
de Bello F, Price J N, Münkemüller T, Liira J, Zobel M, Thuiller W, Gerhold P, GÖtzenberger L, Lavergne S, Lepš J, Zobel K, Pärtel M. Functional species pool framework to test for biotic effects on community assembly. Ecology, 2012, 93(10): 2263-2273. DOI:10.1890/11-1394.1 |
| [34] |
Ding Y, Zang R G, Letcher S G, Liu S R, He F L. Disturbance regime changes the trait distribution, phylogenetic structure and community assembly of tropical rain forests. Oikos, 2012, 121(8): 1263-1270. DOI:10.1111/j.1600-0706.2011.19992.x |
| [35] |
康勇, 熊梦辉, 黄瑾, 龙文兴, 杨小波, 臧润国, 王茜茜, 林灯. 海南岛霸王岭热带云雾林木本植物功能性状的分异规律. 生态学报, 2017, 37(5): 1572-1582. |
| [36] |
Albert C H, Grassein F, Schurr F M, Vieilledent G, Violle C. When and how should intraspecific variability be considered in trait-based plant ecology? Perspectives in Plant Ecology, Evolution and Systematics, 2011, 13(3): 217-225.
|
| [37] |
李家湘, 徐文婷, 熊高明, 王杨, 赵常明, 卢志军, 李跃林, 谢宗强. 中国南方灌丛优势木本植物叶的氮、磷含量及其影响因素. 植物生态学报, 2017, 41(1): 31-42. |
| [38] |
Felsenstein J. Confidence limits on phylogenies: an approach using the bootstrap. Evolution, 1985, 39(4): 783-791. DOI:10.1111/j.1558-5646.1985.tb00420.x |
| [39] |
张佳鑫. 基于叶片功能性状的群落构建机制研究--以湖南八大公山25hm-2样地为例猶[D]. 武汉: 中国科学院大学(中国科学院武汉植物园), 2020.
|
| [40] |
唐青青, 黄永涛, 丁易, 臧润国. 亚热带常绿落叶阔叶混交林植物功能性状的种间和种内变异. 生物多样性, 2016, 24(3): 262-270. |
| [41] |
Díaz S, Kattge J, Cornelissen J H C, Wright I J, Lavorel S, Dray S, Reu B, Kleyer M, Wirth C, Colin Prentice I, Garnier E, Bönisch G, Westoby M, Poorter H, Reich P B, Moles A T, Dickie J, Gillison A N, Zanne A E, Chave J, Joseph Wright S, Sheremet'ev S N, Jactel H, Baraloto C, Cerabolini B, Pierce S, Shipley B, Kirkup D, Casanoves F, Joswig J S, Günther A, Falczuk V, Rüger N, Mahecha M D, Gorné L D. The global spectrum of plant form and function. Nature, 2016, 529(7585): 167-171. |
| [42] |
袁泉, 曹嘉瑜, 刘建峰, 徐德宇, 樊海东, 谭斌, 徐丹妮, 刘立斌, 叶铎, 倪健. 生长型分类方案不同导致森林生态系统植物功能性状的统计偏差. 生态学报, 2021, 41(3): 1106-1115. |
| [43] |
Violle C, Navas M L, Vile D, Kazakou E, Fortunel C, Hummel I, Garnier E. Let the concept of trait be functional!. Oikos, 2007, 116(5): 882-892. |
| [44] |
张林, 罗天祥. 植物叶寿命及其相关叶性状的生态学研究进展. 植物生态学报, 2004, 28(6): 844-852. |
| [45] |
Schulze E D, Kelliher F M, Körner C, Lloyd J, Leuning R. Relationships among maximum stomatal conductance, ecosystem surface conductance, carbon assimilation rate, and plant nitrogen nutrition: a global ecology scaling exercise. Annual Review of Ecology and Systematics, 1994, 25: 629-662. |
| [46] |
Wyka T P, Oleksyn J, Zytkowiak R, Karolewski P, Jagodziński A M, Reich P B. Responses of leaf structure and photosynthetic properties to intra-canopy light gradients: a common garden test with four broadleaf deciduous angiosperm and seven evergreen conifer tree species. Oecologia, 2012, 170(1): 11-24. |
| [47] |
Saura-Mas S, Shipley B, Lloret F. Relationship between post-fire regeneration and leaf economics spectrum in Mediterranean woody species. Functional Ecology, 2009, 23(1): 103-110. |
| [48] |
郑丹楠, 王雪松, 谢绍东, 段雷, 陈东升. 2010年中国大气氮沉降特征分析. 中国环境科学, 2014, 34(5): 1089-1097. |
| [49] |
Yu G, Jia Y, He N, Zhu J, Chen Z, Wang Q, Piao S, Liu X, He H, Guo X, Wen Z, Li P, Ding G, Goulding K. Stabilization of atmospheric nitrogen deposition in China over the past decade. Nature Geoscience, 2019, 12(6): 424-429. |
| [50] |
汪涛, 杨元合, 马文红. 中国土壤磷库的大小、分布及其影响因素. 北京大学学报: 自然科学版, 2008, 44(6): 945-952. |
| [51] |
Reich P B, Wright I J, Cavender-Bares J, Craine J M, Oleksyn J, Westoby M, Walters M B. The evolution of plant functional variation: traits, spectra, and strategies. International Journal of Plant Sciences, 2003, 164(S3). |
| [52] |
Keenan T F, Niinemets V. Global leaf trait estimates biased due to plasticity in the shade. Nature Plants, 2017, 3: 16201. |
| [53] |
王瑞丽, 于贵瑞, 何念鹏, 王秋凤, 赵宁, 徐志伟. 中国森林叶片功能属性的纬度格局及其影响因素. 地理学报, 2015, 70(11): 1735-1746. |
| [54] |
刘可佳, 何念鹏, 侯继华. 中国温带典型森林植物比叶面积的空间格局及其影响因素. 生态学报, 2022, 42(3): 872-883. |
| [55] |
Zhao L J, Xiang W H, Li J X, Lei P F, Deng X W, Fang X, Peng C H. Effects of topographic and soil factors on woody species assembly in a Chinese subtropical evergreen broadleaved forest. Forests, 2015, 6(12): 650-669. |