 2022, Vol. 42
2022, Vol. 42文章信息
- 赵鹏, 徐先英, 姜生秀, 徐高兴, 柴成武, 张逸君, 马玉莲, 戚福俊, 高德才, 张国平
- ZHAO Peng, XU Xianying, JIANG Shengxiu, XU Gaoxing, CHAI Chengwu, ZHANG Yijun, MA Yulian, QI Fujun, GAO Decai, ZHANG Guoping
- 石羊河下游不同衰退程度多枝柽柳灌丛水分利用格局研究
- Water utilization pattern of Tamarix ramosissima Ledeb. Nebkhas with different decline degrees in the lower reaches of Shiyang River
- 生态学报. 2022, 42(17): 7187-7197
- Acta Ecologica Sinica. 2022, 42(17): 7187-7197
- http://dx.doi.org/10.5846/stxb202101280303
-
文章历史
- 收稿日期: 2021-01-28





2. 中国科学院西北生态环境资源研究院, 沙漠与沙漠化重点实验室, 兰州 730010;
3. 兰州大学西部生态安全省部共建协同创新中心, 兰州 730010;
4. 古浪县海子滩林场, 古浪 733100
2. Key Laboratory of Desert and Desertification, Northwest Institute of Eco-environment and Resources, Chinese Academy of Science, Lanzhou 730010, China;
3. Collaborative Innovation Center for Western Ecological Safety, Lanzhou University, Lanzhou 730010, China;
4. Gulang County Haizitan forest field, Gulang 733100, China
干旱区约占地球陆表面积的40%以上, 所支撑的复杂多样生态系统是全球陆地生态系统的重要组成, 水是干旱区诸多生态系统过程的驱动力和关键的非生物限制因子[1]。植物-土壤水分相互关系是生态水文学研究的核心内容[2]。由于植物生活型、年龄及生境因素的影响, 造成不同或同一物种的水分利用策略产生差异[3]。作为一种干旱应对策略, 植物水分来源与根系分布并非完全一致, 与环境中可利用水的量密切相关[4—5]。降水和地下水是荒漠植物主要水分来源[6], 对不同水源的选择性利用影响着群落的结构和稳定性[7]。氢氧稳定同位素是天然示踪剂, 在水分溯源、水分滞留时间估算与植物水分利用策略等方面具有独特优势[8—9], 被广泛应用于研究新疆莫索湾地区杨树和农作物[10]、毛乌素沙地灌木群落[11]、黄土丘陵区优势造林树种[12]等不同气候区植物水分来源及种间水分关系。研究发现盐生和旱生植物在水分由根系进入植物体的过程中, 氢稳定同位素(D)会发生分馏[13]。因此, 本研究选择氧同位素(18O)为主要示踪指标。
柽柳是中亚干旱荒漠区生态系统中的关键种之一, 在维持生物多样性方面具有重要的生态地位[14]。在不同生境及水文气候条件下, 柽柳属植物主要水分来源明显不同。准格尔盆地东南缘多枝柽柳以利用深层土壤水和地下水为主, 水分来源没有明显季节波动特征[15]。黑河中游柽柳6月主要利用40—60cm土壤水分, 而在8月则主要利用60cm以下深层土壤水或地下水[16]。黑河下游额济纳三角洲多枝柽柳吸收土壤水的层位因地下水位波动、土壤物理特性、河水对土壤水的侧向补给及漫溢补给等存在较大的空间差异, 但其利用河水和地下水的比例未见明显空间差异[17]。塔里木河上游, 柽柳在浅地下水埋深和大于450cm的深地下水埋深条件下均能良好生长, 对不同水分条件的适应能力优于胡杨[18]。塔里木河下游, 为了适应极端干旱, 无论是乔木胡杨、还是灌木柽柳, 水分来源主要是较稳定的深层水源, 且对各水源的利用比例在不同月份波动不大[19]。黄河三角洲贝壳堤岛内柽柳能通过改变自身的主要水分来源适应生境和降水条件的变化, 这种水源变换能力有利于提高其竞争优势[20]。目前, 关于石羊河下游多枝柽柳灌丛水分来源与季节变化研究鲜见报道。为此, 基于稳定氢氧同位素技术, 本文通过研究柽柳灌丛水分来源的时空变化规律, 以期为绿洲边缘退化防护体系的修复与重建提供理论依据。
1 材料与方法 1.1 野外取样按照“空间代替时间”的方法, 结合地上部分的长势、枯枝情况等生物学特征综合判断多枝柽柳灌丛的衰退情况, 共设置沙井子、三角城、外西、刘家地4个样地, 分别代表石羊河下游轻度、中度、重度、极重度衰退程度多枝柽柳灌丛沙堆(图 1、表 1)。植物、土壤、降雨、地下水稳定同位素样品采集参考林光辉主编的《稳定同位素生态学》中稳定同位素样品采集方法[21]。2016年5月至10月, 每月下旬在样地选取3个长势、形态大小基本相同的多枝柽柳灌丛, 在灌丛东、南、西、北不同方向采集枝条木质部样品, 共3个重复。剪取直径约为0.1—0.3cm, 长3—5cm非绿色栓化小枝, 去皮后迅速装入8mL采样瓶, Parafilm封口膜密封, 装入便携式恒温箱冷藏, 带回实验室-18℃冷冻保存。用土钻在各样地以20cm间隔分层取土样, 深度为200cm, 所取土样一分部分装入6个8mL采样瓶, 用Parafilm封口膜密封, 放入恒温箱冷藏, 带回实验室冷冻保存, 测定土壤水分的稳定氢氧同位素值。另一部分装入铝盒内, 测定土壤含水量。降雨样品采用雨量器收集, 每次降雨后记录降雨量和降雨日期, 专用玻璃瓶收集雨水。Parafilm封口膜密封, 3—4℃冷藏保存。地下水样品采自样地附近农田灌溉井, 专用玻璃瓶收集, 封口膜密封, 3—4℃冷藏保存。不同土层间土壤水氢氧稳定同位素的相似性增加了识别独特水源的难度。因此, 依据不同土层土壤含水量及氢氧稳定同位素组成特征, 将土壤划分为0—60cm、60—120cm和120—200cm共3个土层。

|
| 图 1 石羊河下游不同衰退程度多枝柽柳灌丛沙堆 Fig. 1 T. ramosissima nebkhas with different decline degrees in lower reaches of Shiyang River |
| 样地名称 Plot name |
退化程度 Decline degree |
灌丛形态(长×宽×高) Shrub morphology (Length×width×height)/ (m× m×m) |
地下水位 Groundwater level/m |
与农田间距离 Distance to farmland/m |
伴生种 Accompanying species |
| 沙井子 | 轻度 | 7×5×1.8 | 25 | 10 | 雾冰藜(Bassia dasyphylla)、 猪毛菜(Salsola collina)、 沙拐枣(Calligonum mongolicum) |
| 三角城 | 中度 | 12×8×4.8 | 20 | 1500 | 白刺(Nitraria tangutorum)、 红砂(Reaumuria soongarica)、 梭梭(Haloxylon ammodendron) |
| 外西 | 重度 | 5×3×1.5 | 13 | 1200 | 白刺、红砂 |
| 刘家地 | 极重度 | 7×5×2.3 | 28 | 350 | 红砂、猪毛菜、雾冰藜 |
稳定氢氧同位素样品采集具体时间分别为2016年4月22日、23日(降雨后1个月, 3月降雨21.1mm, δ18O为-8.79‰), 5月28日、29日(降雨后5d, 5月20日—5月22日连续降雨12mm, δ18O为-10.27‰), 6月27日、6月28日(降雨后1d, 6月26日降雨1. 1mm, δ18O为-8.64‰), 7月28日、7月29日(降雨后15d, 7月13日降雨7.5mm, δ18O为-4.3‰), 8月31日, 9月1日(降雨后1d, 8月29日降雨4.5mm, δ18O为-2.04‰), 10月9日、10日(降雨后5d, 10月4日降雨4.3mm, δ18O为-1.20‰)。
1.2 样品分析采用烘干称重法进行土壤水分含量测定, 土样带回实验室, 用万分之一电子天平称取鲜重, 置于电热恒温鼓风干燥箱, 在105℃高温烘干12h至恒重, 待冷却后称重其干重。土壤含水量计算公式:
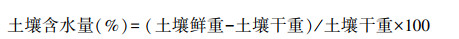
|
氢氧稳定同位素样品预处理与分析测定在荒漠化与风沙灾害防治国家重点实验室培育基地碳水同位素分析室进行。采用低温真空抽提法提取土壤水和植物水。真空蒸馏是提取植物和土壤中水分应用最广泛的方法之一, 操作简单, 且不需要附加的溶剂。真空蒸馏期间, 水分从样品中蒸发出来并凝结在收集管中。在收集管中凝结的同位素遵循雷利(Rayleigh)蒸馏曲线, 水分的抽提必须完全才可获得未分馏水分样品, 抽提率可达98%以上。利用碳水同位素分析实验室Picarro L2120-i水同位素分析仪测定样品氢氧稳定同位素, 测试结果以相对于维也纳标准平均海水(VSMOW)的千分值偏差表示:
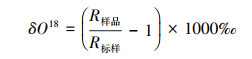
|
式中, R样品和R标样分别为样品和国际通用标准物中元素的轻重同位素之比(18O/16O), 测试精度为:0.5‰ (δD)、0.1‰(δ18O)。
1.3 数据处理数据统计分析与作图采用Origin 9.0软件。运用IsoSource模型计算不同衰退程度多枝柽柳灌丛对各水源的可能利用比例。该模型以同位素质量平衡的多元线性混合方程为基础, 用于计算多个水源时植物对各水源的利用比例[22]。在初步判定两种植物的水分来源后, 将植物木质部水和各潜在水源的δ18O值代入模型, 构建质量平衡公式:
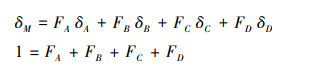
|
式中, FA、FB、FC、FD为各潜在水源所占比例, δA、δB、δC、δD为各水源的δ18O值, δM为植物木质部水的δ18O值。模型为植物的每个潜在水源的利用比例赋值, 比例增加的步长视为来源增量, 如1%或2%, 各水源被利用比例的组合总计为100%, 利用迭代方法计算可能的比例组合。当各水源的δ18O值加权值之和等于植物木质部水的δ18O值, 或小于某一可接受范围(质量平衡公差)时, 视为符合条件的比例组合。模型运行结果中, 植物对各水源的利用比例是一个范围(最大值—最小值)。本研究中, 模型的来源增量设为1%, 质量平衡公差设为0.1‰。
2 结果 2.1 降雨特征2016年4—10月采样期间石羊河下游民勤绿洲总降水量为101.6mm, 占全年降雨量的81.67%, 主要发生在6、7、8月。共发生降雨事件48次, 小于5mm的小降雨事件占95.83%。2016年石羊河下游降水线方程为δD=6.46δ18O-5.11, R2=0.87, 1—5月降水δ18O、δD分别介于为-10.65‰—-7.52‰, -152.5‰—59.51‰, 均值分别为-11.08‰、-104.26‰。6—10月降水δ18O、δD分别介于为-6.85‰—-1.2‰, -44.60‰—7.66‰, 均值分别为-4.57‰、-18.07‰。降水δ18O介于-3.3‰—-11.5‰, 随季节的变化明显(图 2)。由于温度的不同, 降水δ18O夏季、秋季明显高于春季, 表现出明显的季节效应。

|
| 图 2 2016年石羊河下游民勤绿洲降水量、温度、δ18O和δD的季节变化 Fig. 2 Seasonal variations of rainfall, temperature, δ18O and δD of Minqin oasis in 2016 δ18O: 氧同位素δD: 氢同位素 |
不同衰退程度多枝柽柳灌丛季节土壤水分垂直分布各异。由图 3可知, 轻度衰退阶段, 0—60cm土层春季土壤水分逐渐增大, 夏季、秋季增大幅度较小。春季属于萌发期水分消耗少, 多枝柽柳灌丛60—120cm土层土壤水分显著大于夏季、秋季。不同衰退程度多枝柽柳土壤水分表层0—60cm均受降雨及蒸发的影响波动明显,120—200cm土壤水分相对稳定。4月、5月属于萌发期水分消耗少, 土壤水分均大于6、7、8月生长期。生长季多枝柽柳灌丛0—200cm平均土壤水分大小排序为:重度(3.48%)>极重度(2.69%)>中度(1.97%)>轻度(1.87%)。

|
| 图 3 石羊河下游不同衰退程度多枝柽柳灌丛土壤水分(δ18O)垂直变化 Fig. 3 Vertical variation of soil moisture (δ18O) of T. ramosissima nebkhas with different decline degrees in lower reaches of Shiyang River |
由图 4可知, 多枝柽柳灌丛土壤水氢氧同位素值分布在大气降水线之下, 证明降水补给土壤水过程中由于蒸发作用使土壤水中的δ18O、δD富集。不同衰退程度多枝柽柳灌丛土壤水蒸发方程斜率不一样, 蒸发强度排序为重度>中度>轻度>极重度。轻度衰退多枝柽柳灌丛土壤蒸发线为δD=2.92δ18O-45.83, R2=0.84, δ18O、δD变化范围为-7.68‰—25.63‰, -74.97‰—32.90‰, 均值分别为-1.28‰, -49.56‰。中度衰退多枝柽柳灌丛土壤蒸发线为δD=4.6δ18O-46.19, R2=0.90, δ18O、δD变化范围为-23.42‰—7.19‰, -167.13‰—-20.83‰, 均值分别为-0.51‰, -51.55‰。重度衰退多枝柽柳灌丛土壤蒸发线为δD=4.09δ18O-45.53, R2=0.91, δ18O、δD变化范围为-22.68‰—6.13‰, -154.09‰—-18.67‰, 均值分别为-3.80‰, -60.99‰。极重度衰退阶段多枝柽柳灌丛土壤蒸发线为δD=2.54δ18O-48.09, R2=0.68。土壤水δ18O、δD变化范围为-12.15‰—21.04‰, -103.68‰—-21.73‰, 均值分别为-1.41‰, -51.82‰。降水与土壤水δD对比分析表明不同衰退程度多枝柽柳灌丛土壤水δD低于1—5月降水δD均值, 但其δD高于6—10月降水, 证明6—10月降水对土壤水补给贡献较大。

|
| 图 4 石羊河下游不同衰退程度多枝柽柳灌丛土壤水δ18O与δD关系 Fig. 4 Relationship between δ18O and δD in soil water of T. ramosissima nebkhas with different decline degrees in Shiyang River |
多枝柽柳灌丛地下水、木质部水、降水δ18O随季节和退化程度变化规律各异(表 2)。轻度衰退, 地下水δ18O介于-9.1‰—-9.6‰, 变幅0.5‰。中度衰退, 地下水δ18O介于-8.1‰—-10.1‰, 变幅2‰。重度衰退, 地下水δ18O介于-10.5‰—-11.0‰, 变幅0.5‰。极重度衰退, 地下水δ18O介于-8.74‰—-9.18‰, 变幅0.44‰。多枝柽柳木质部水δ18O轻度衰退, 介于-11.43‰—-8.17‰, 中度衰退介于-11.6‰—-8.4‰, 重度衰退介于-10.37‰—-13.98‰, 极重度衰退, 木质部水δ18O介于-9.34‰—-5.52‰, 春季明显高于夏季、秋季。
| 潜在水源 Potential water source |
衰退程度 Severity of decline |
春季 Spring |
夏季 Summer |
秋季 Autumn |
| 浅层土壤水 | 轻度 | 1.02±1.63 | -1.81±1.99 | 5.76±4.42 |
| Shallow soil water | 中度 | -0.25±1.48 | 0.41±1.90 | 4.11±1.52 |
| 重度 | 0.78±1.29 | -0.49±1.36 | 1.02±2.79 | |
| 极重 | 0.41±3.15 | 2.69±4.18 | 0.96±1.21 | |
| 中层土壤水 | 轻度 | 0.26±1.11 | -3.81±0.82 | -2.32±0.99 |
| Middle soil water | 中度 | -3.75±2.11 | -0.75±1.19 | 2.34±0.91 |
| 重度 | -3.25±1.07 | -5.04±1.69 | -4.22±2.82 | |
| 极重 | -1.59±1.68 | -5.54±2.50 | -0.95±1.55 | |
| 深层土壤水 | 轻度 | -3.24±1.20 | -6.29±0.30 | -3.04±2.86 |
| Deep soil water | 中度 | -2.79±1.00 | -3.34±1.81 | 0.08±1.13 |
| 重度 | -6.49±1.41 | -7.82±0.29 | -5.65±2.63 | |
| 极重 | -4.18±1.59 | -7.15±1.02 | -4.92±1.25 | |
| 木质部水 | 轻度 | -8.17±0.60 | -11.11±3.46 | -11.43±2.08 |
| Xylem water | 中度 | -8.40±0.91 | -10.57±1.47 | -11.60±2.81 |
| 重度 | 10.37±1.40 | -13.98±1.48 | -11.12±1.82 | |
| 极重 | -5.52±2.9 | -8.28±1.03 | -9.34±1.45 | |
| 地下水 | 轻度 | -9.6±0.1 | -9.3±0.4 | -9.1±0.1 |
| Groundwater | 中度 | -9.3±0.05 | -10.1±0.2 | -8.1±1.01 |
| 重度 | -10.5±0.5 | -10.6±0.03 | -11.0±0.04 | |
| 极重 | -8.74±0.05 | -8.81±1.00 | -9.18±0.17 | |
| 降雨 | 轻度 | -9.1±3.4 | -3.3±5.3 | -3.6±3.7 |
| Precipitation | 中度 | -9.1±2.3 | -5.5±3.1 | -4.3±3.8 |
| 重度 | -11.5±1.5 | -6.5±0.9 | -4.8±0.02 | |
| 极重 | -9.82±1.90 | -5.52±2.96 | -0.82±1.35 |
由图 5可知, 不同衰退程度多枝柽柳灌丛沙堆土壤水的蒸发线方程为δD=2.9618O-45.9, R2=0.74。蒸发线位于全球大气水线的右下方, 表明降水入渗过程经历了较强的蒸发过程。多枝柽柳木质部水与土壤水、降水、地下水δD-δ18O交叉分布在大气水线以下, 证明土壤水、降水、地下水是不同衰退程度多枝柽柳灌丛的潜在水源。

|
| 图 5 降水、土壤水、植物水与地下水δD与δ18O的线性回归关系 Fig. 5 Regression relationship between δD and δ18O in rainfall, soil water, plants water and groundwater |
不同退化程度多枝柽柳灌丛生长季水分来源差异明显。轻度衰退阶段, 多枝柽柳春季利用地下水46.1%、降水39.3%。夏季利用地下水86.7%, 降水2.6%, 深层土壤水5.8%。秋季利用地下水96.6%, 中层土壤水1.8%。中度衰退阶段, 多枝柽柳春季各水源利用比例分别为地下水46.2%, 降水41.7%, 中层土壤水5%。夏季利用地下水93.6%, 降水2.7%, 深层土壤水1.7%。秋季利用地下水94.5%, 降水2.6%, 深层土壤水1.6%。重度衰退阶段, 多枝柽柳春季利用降水58.5%, 地下水30.2%, 深层土壤水5.8%。夏季地下水85.8%, 深层土壤水6.3%, 降水3.6%。秋季地下水94.9%, 降水1.3%, 深层土壤水2.1%。极重度衰退阶段, 多枝柽柳春季各主要水源利用比例分别为地下水24.4%, 降水23%, 深层土壤水22%。夏季各主要水源利用比例分别为地下水81.8%, 深层土壤水8.8%, 降水4.2%。秋季地下水96.3%, 浅层土壤水0.8%, 深层土壤水2.5%(图 6)。不同衰退程度多枝柽柳灌丛春季对降水的利用比例排序为重度衰退>中度衰退>轻度衰退>极重度衰退, 春季到秋季趋向于利用深层地下水。

|
| 图 6 石羊河下游不同退化程度多枝柽柳灌丛对潜在水源的吸收利用比例 Fig. 6 The absorption and utilization ratio of potential water source of T. ramosissima nebkhas with different degradation degree in Shiyang River |
柽柳科(Tamaricaceae)是一个古老的科, 起源于第三纪, 包括亚洲中部在内的“古地中海”沿岸地区, 我国西北内陆河下游绿洲荒漠过渡带大量分布[23]。柽柳科植物属于利用盐腺向外泌盐的深根性湿生植物[24], 可在低叶水势条件下进行光合作用, 同时具有较低的光补偿点和暗呼吸速率, 气孔随干湿季水分状况对蒸腾速率的适应性调整较灵敏[25]。柽柳林地0—30cm表层含水量受蒸发和降雨影响较大, 林地30—200cm土壤平均含水量较裸沙地低[25]。巴丹吉林沙漠柽柳灌丛土壤水分呈浅层(10—70cm)高、中层(70—150cm)低、深层(150—210cm)高的特点[26], 本研究发现不同衰退程度多枝柽柳灌丛沙堆土壤水分同样存在表层(0—60cm)高的特点, 中下层(60—200cm)土壤水分相对稳定。本研究野外调查发现多枝柽柳通过自我疏枝的方式维持存活, 将有限的水分维持部分枝条的正常生长, 大部分枝条已干枯, 以降低整个灌丛的水分消耗。多枝柽柳适宜生境土壤含水量范围为3.55%—9.4%[27], 本研究生长季多枝柽柳灌丛0—200cm土层平均土壤水分:重度(3.48%)>极重度(2.69%)>中度(1.97%)>轻度(1.87%), 均小于最适生境土壤水分, 处于衰退演替状态。由于主要利用地下水和深层土壤水, 在降水引发的湿润-干旱周期中, 柽柳水分生理参数对降水无响应[28]。与生长正常的柽柳相比, 衰退多枝柽柳灌丛沙堆冠层截留减小, 沙堆裸露面积增大, 降水可直接入渗补给衰退柽柳灌丛。因此, 本研究降水与土壤水氢稳定同位素季节变化特征表明, 6—10月降水对石羊河下游衰退多枝柽柳灌丛土壤水补给贡献较大。
多枝柽柳灌丛的退化是以土壤水分和生物特性为主导因子, 养分、盐分和土壤结构等因子综合影响的结果[30]。我国最早的自然地理书籍《禹贡图注》将石羊河下游的民勤划为雍州区湖泊沼泽平原, 分布有大量沼泽植物。冯绳武推算西汉以前水域面积占民勤县的80%[31]。在清乾隆十四年(1749年)的《镇番县志》中, 民勤境内以柽柳(红柳)命名的地名甚多, 如柳林湖、红柳岗、红柳墩、红柳园, 可反映出当时柽柳的广布性。民国年间, 石羊河下游的大西河、小西河河道断流, 青土湖、白亭湖干涸。20世纪以来, 受干旱气候的影响, 民勤荒漠化急剧扩张, 逐渐形成绿洲边缘较为稳定的柽柳、白刺天然灌丛沙包, 即“柴湾”。植被从水生、沼泽、草甸植物向旱生、强旱生、超旱生方向演替, 物种大量消失, 已演化成现在的沙漠、戈壁与绿洲景观[32]。随地表水锐减, 地下水在民勤沙区农、林业生产及水分平衡中起至关重要的作用[33]。由于民勤盆地每年超采地下水超过3×108m3, 导致了区域性地下水位从20世纪70年代的1—9 m下降到20世纪初的12—28m[34]。随着地下水位下降, 民勤沙井子地区荒漠植物种类和植被类型趋于旱化, 湿生和中生植物相继退化[35]。地下水下降至10m以下, 土壤水分对柽柳的生长起到主导作用。民勤绿洲边缘柽柳荒漠林林带变窄, 盖度降低, 向旱生、超旱生群落发展, 退化明显[36], 以白刺、红砂为主的荒漠植物逐步发展为优势种群[37]。
水资源是石羊河下游多枝柽柳灌丛演替的关键驱动因子。当根系与地下水接触时, 柽柳气孔对水分的控制作用微弱; 地下水降低的干旱年份, 通过气孔导度控制水分散失[38]。作为北美地区的天然入侵种, 柽柳的大量根系通过毛细管作用与地下水接触[39], 不仅能通过深根系获取地下水, 地下水水位下降时也能够吸收利用不饱和区域的土壤水分, 水分竞争力较强[40]。随年内的地下水水位波动, 柽柳选择性利用地下水和浅层(25—60cm)土壤水[41]。宁夏平原北部引黄灌区幼龄多枝柽柳吸收利用浅层不饱和土壤水, 对灌溉反应不明显。20年林龄多枝柽柳对地下水的利用率达18.3%, 不同水分条件下植物的水分利用策略跟林龄密切相关[42]。土壤水是塔克拉玛干沙漠腹地达里雅布依绿洲柽柳幼苗的主要水源, 随着地下水埋深的增加, 柽柳幼苗的吸水深度下移。柽柳幼苗水分利用效率随地下水埋深的增加而略有增加[43]。塔克拉玛干沙漠南缘人工防护林滴灌群落多枝柽柳对中层土壤水(40—100cm)的利用率高于自然群落, 但其主要水分来源仍然是地下水, 说明其根系已经深达地下水层[44]。黄河兰州段河漫滩多枝柽柳在生长季主要水分来源是0—30cm浅层土壤水(28.3%), 对河水的利用率最小(16.6%), 但对各潜在水源的利用比较平均[45]。当夜间空气湿度达到85%时, 在水通道蛋白(PIP2)基因调控下, 柽柳叶片还可以直接吸收利用大气水汽以适应极端干旱环境[46]。小降雨事件不仅有助于抑制蒸发, 还可以通过叶片吸收补充柽柳叶片蒸腾失水[47]。本研究发现石羊河下游不同衰退程度多枝柽柳春季主要利用降水与地下水, 对降水的利用比例排序为重度衰退>中度衰退>轻度衰退>极重度衰退, 夏季和秋季趋向于利用深层地下水。由于民勤绿洲地下水埋深已超过多枝柽柳可吸收利用的深度, 柽柳荒漠林出现大面积的的衰退死亡。研究发现绿洲荒漠过渡带农田-防护林带-荒漠存在水力联系, 农田渗流水是防护林带和荒漠植物种重要的水分来源[48]。研究区残存柽柳灌丛能够利用到的地下水多为绿洲农田灌溉的测渗水。胡杨(Populus euphratica)是地质历史时期残留的荒漠河岸林木, 50年代民勤二坝湖有胡杨片林生长, 随着地下水位的下降, 70年代还有部分胡杨林生长, 但长势已很弱, 时至今日片林已全是枯木。民勤地区是胡杨分布的边缘地带, 现除个别人工栽培抚育管理的林木成活外, 天然分布胡杨已完全绝迹[35]。由于石羊河下游绿洲边缘水文条件难以满足胡杨、多枝柽柳等高大乔灌木生存需求, 由此可以预测若干年后天然分布的多枝柽柳种群也会像胡杨一样逐渐被旱生小灌木、草本所替代。
4 结论(1) 2016年石羊河下游降水线方程为δD=6.46δ18O-5.11, R2=0.87, 以小降雨事件为主, 降雨δ18O的季节效应明显。
(2) 不同衰退程度多枝柽柳灌丛土壤水分在夏季较低, 秋季略为恢复。衰退程度越严重蒸腾耗水越低, 多枝柽柳灌丛0—200cm土层深度平均土壤水分排序为重度(3.48%)>极重度(2.69%)>中度(1.97%)>轻度(1.87%)。6月—10月降水对土壤水补给贡献较大。
(3) 受春季降水蒸发损失小, 冠层截留以及土壤生物结皮的综合影响, 不同衰退程度多枝柽柳春季对降水的平均利用比例达40.63%, 大小排序为重度衰退(58.5%)>中度衰退(41.7%)>轻度衰退(39.3%)>极重度衰退(23%)。春季到秋季衰退多枝柽柳灌丛趋向于利用可靠的深层地下水。
| [1] |
李新荣, 张志山, 王新平, 刘立超, 黄磊. 干旱区土壤植被系统恢复的生态水文学研究进展. 中国沙漠, 2009, 29(5): 845-852. |
| [2] |
王根绪, 钱鞠, 程国栋. 生态水文科学研究的现状与展望. 地球科学进展, 2001, 16(3): 314-323. DOI:10.3321/j.issn:1001-8166.2001.03.004 |
| [3] |
Dawson T E, Pate J S. Seasonal water uptake and movement in root systems of Australian phraeatophytic plants of dimorphic root morphology: a stable isotope investigation. Oecologia, 1996, 107(1): 13-20. DOI:10.1007/BF00582230 |
| [4] |
Ewe S M, Sternberg D L. Seasonal water-use by the invasive exotic, Schinus terebinthifolius, in native and disturbed communities. Oecologia, 2002, 133(4): 441-448. DOI:10.1007/s00442-002-1047-9 |
| [5] |
Ehleringer J R, Dawson T E. Water uptake by plants: perspectives from stable isotope composition. Plant, Cell and Environment, 1992, 15(9): 1073-1082. DOI:10.1111/j.1365-3040.1992.tb01657.x |
| [6] |
Dawson T E, Ehleringer J R. Streamside trees that do not use stream water. Nature, 1991, 350(6316): 335-337. DOI:10.1038/350335a0 |
| [7] |
Ehleringer J R, Phillips S L, Schuster W S F, Sandquist D R. Differential utilization of summer rains by desert plants. Oecologia, 1991, 88(3): 430-434. DOI:10.1007/BF00317589 |
| [8] |
孙龙, 陈利顶, 杨磊. 基于氢氧同位素技术的流域水源涵养研究进展. 生态学报, 2020, 40(24): 8872-8881. |
| [9] |
徐庆, 王婷, 高德强. 碳氢氧稳定同位素在草地生态系统水循环研究中的应用. 林业科学研究, 2019, 32(6): 130-136. |
| [10] |
郭辉, 赵英, 蔡东旭, 兰志龙, 雷加强. 氢氧同位素示踪法探测新疆地区防护林和棉花体系水分来源与竞争. 生态学报, 2019, 39(18): 6642-6650. |
| [11] |
朱雅娟, 崔清国, 杜娟, 许素寒, 刘志兰. 毛乌素沙地三种灌木群落的水分利用过程. 生态学报, 2020, 40(13): 4470-4478. |
| [12] |
吴旭, 牛耀彬, 荀梦瑶, 金俊逸, 李娜艳, 唐亚坤, 陈云明. 黄土丘陵区优势造林树种水分来源对季节性干旱的响应. 生态学报, 2022, 42(10). DOI:10.5846/stxb202106221657 |
| [13] |
Ellsworth P Z, Williams D G. Hydrogen isotope fractionation during water uptake by woody xerophytes. Plant and Soil, 2007, 291(1/2): 93-107. |
| [14] |
尹林克. 中亚荒漠生态系统中的关键种──柽柳(Tamarixspp.). 干旱区研究, 1995, 12(3): 43-47. |
| [15] |
周海, 郑新军, 唐立松, 李彦. 准噶尔盆地东南缘多枝柽柳、白刺和红砂水分来源的异同. 植物生态学报, 2013, 37(7): 665-673. |
| [16] |
龙翔, 孙自永, 周爱国, 余绍文, 曹弘. 黑河中游河岸带柽柳水分来源的18O同位素示踪研究. 干旱区资源与环境, 2014, 28(7): 150-155. |
| [17] |
李亚飞, 于静洁, 陆凯, 王平, 张一驰, 杜朝阳. 额济纳三角洲胡杨和多枝柽柳水分来源解析. 植物生态学报, 2017, 41(5): 519-528. |
| [18] |
周天河, 赵成义, 吴桂林, 蒋少伟, 俞永祥, 王丹丹. 塔里木河上游胡杨(Populus euphratica、柽柳(Tamarix ramosissima)水分来源的稳定同位素示踪. 中国沙漠, 2017, 37(1): 124-131. |
| [19] |
王玉阳, 陈亚鹏, 李卫红, 王日照, 周莹莹, 张建鹏. 塔里木河下游典型荒漠河岸植物水分来源. 中国沙漠, 2017, 37(6): 1150-1157. |
| [20] |
朱金方, 刘京涛, 孙景宽, 赵艳云, 陆兆华, 李俊生. 贝壳堤岛不同生境下柽柳水分来源比较. 生态学杂志, 2017, 36(8): 2367-2374. |
| [21] |
林光辉. 稳定同位素生态学. 北京: 高等教育出版社, 2013: 1-492.
|
| [22] |
曾祥明, 徐宪立, 钟飞霞, 易汝舟, 徐超昊, 张耀华. MixSIAR和IsoSource模型解析植物水分来源的比较研究. 生态学报, 2020, 40(16): 5611-5619. |
| [23] |
张元明, 潘伯荣, 尹林克, 杨维康, 张道远. 柽柳科(Tamaricaceae)植物的研究历史. 西北植物学报, 2001, 21(4): 796-804. DOI:10.3321/j.issn:1000-4025.2001.04.033 |
| [24] |
周三, 韩军丽, 赵可夫. 泌盐盐生植物研究进展. 应用与环境生物学报, 2001, 7(5): 496-501. DOI:10.3321/j.issn:1006-687X.2001.05.020 |
| [25] |
邓雄, 李小明, 张希明, 叶万辉, Andrea Foezki, Michael Runge. 多枝柽柳气体交换特性研究. 生态学报, 2003, 23(1): 180-187. DOI:10.3321/j.issn:1000-0933.2003.01.022 |
| [26] |
刘发民, 金燕, 张小军. 荒漠地区柽柳人工固沙林土壤水分动态研究. 西北植物学报, 2001, 21(5): 937-943. DOI:10.3321/j.issn:1000-4025.2001.05.018 |
| [27] |
张进虎, 王乃昂, 牛震敏, 孙杰, 张律吕. 巴丹吉林沙漠高大沙山柽柳地土壤水分时空动态. 水土保持研究, 2018, 25(4): 92-97. |
| [28] |
公维昌, 庄丽, 赵文勤, 占东霞, 张莉. 多枝柽柳与梭梭光合器官形态解剖结构的生态适应性. 中国沙漠, 2011, 31(1): 129-136. |
| [29] |
徐贵青, 李彦. 共生条件下三种荒漠灌木的根系分布特征及其对降水的响应. 生态学报, 2009, 29(1): 130-137. DOI:10.3321/j.issn:1000-0933.2009.01.016 |
| [30] |
丁爱强. 民勤绿洲荒漠过渡带退化柽柳灌丛沙堆植被群落与土壤特性研究[D]. 北京: 中国林业科学研究院, 2018.
|
| [31] |
冯绳武. 民勤绿洲的水系演变. 地理学报, 1963, 18(3): 241-249. DOI:10.3321/j.issn:0375-5444.1963.03.006 |
| [32] |
满多清, 刘世增, 魏振海, 刘虎俊, 李银科, 刘淑娟. 石羊河中下游流域植被的演替历史. 中国沙漠, 2013, 33(2): 613-618. |
| [33] |
俄有浩, 严平, 仲生年, 韩福贵. 民勤沙井子地区地下水动态研究. 中国沙漠, 1997, 17(1): 70-76. DOI:10.3321/j.issn:1000-694X.1997.01.013 |
| [34] |
马金珠, 魏红. 民勤地下水资源开发引起的生态与环境问题. 干旱区研究, 2003, 20(4): 261-265. |
| [35] |
杨自辉. 民勤沙井子地区40年来荒漠植被变迁初探. 中国沙漠, 1999, 19(4): 395-398. DOI:10.3321/j.issn:1000-694X.1999.04.016 |
| [36] |
马全林, 王继和, 刘虎俊, 杨自辉, 詹科杰. 民勤绿洲边缘柽柳荒漠林的时空变化及其驱动因素. 中国沙漠, 2006, 26(5): 802-808. DOI:10.3321/j.issn:1000-694X.2006.05.024 |
| [37] |
徐高兴, 王立, 徐先英, 金红喜, 柴成武. 民勤绿洲边缘地下水埋深对柽柳灌丛生长及物种多样性的影响. 草原与草坪, 2017, 37(2): 49-56. DOI:10.3969/j.issn.1009-5500.2017.02.008 |
| [38] |
曾凡江, 张希明, 李小明. 柽柳的水分生理特性研究进展. 应用生态学报, 2002, 13(5): 611-614. DOI:10.3321/j.issn:1001-9332.2002.05.023 |
| [39] |
Gay L W, Fritschen L J. An energy budget analysis of water use by Saltcedar. Water Resources Research, 1979, 15(6): 1589-1592. DOI:10.1029/WR015i006p01589 |
| [40] |
Busch D E, Ingraham N L, Smith S D. Water uptake in woody riparian phreatophytes of the southwestern United States: a stable isotope study. Ecological Applications, 1992, 2(4): 450-459. DOI:10.2307/1941880 |
| [41] |
Sun Z Y, Long X, Ma R. Water uptake by saltcedar (Tamarix ramosissima) in a desert riparian forest: responses to intra-annual water table fluctuation. Hydrological Processes, 2016, 30(9): 1388-1402. DOI:10.1002/hyp.10688 |
| [42] |
朱林, 许兴, 毛桂莲. 宁夏平原北部地下水埋深浅地区不同灌木的水分来源. 植物生态学报, 2012, 36(7): 618-628. |
| [43] |
麦尔哈巴·尼加提, 比拉力·依明, 师庆东, 戴岳, 史浩伯, 肖合来提·巴义. 沙漠腹地达里雅布依绿洲柽柳幼苗的水分利用来源. 干旱区资源与环境, 2021, 35(8): 159-166. |
| [44] |
孟德坤, 闵孝君, 臧永新, 马剑英. 塔克拉玛干沙漠南缘人工防护林和自然群落多枝柽柳叶片养分特征与水分利用策略. 生态学杂志, 2019, 38(11): 3245-3253. |
| [45] |
苏鹏燕, 张明军, 王圣杰, 邱雪, 王家鑫, 杜勤勤, 郭蓉, 车存伟. 基于氢氧稳定同位素的黄河兰州段河岸植物水分来源. 应用生态学报, 2020, 31(6): 1835-1843. |
| [46] |
Yan X, Zhou M X, Dong X C, Zou S B, Xiao H L, Ma X F. Molecular mechanisms of foliar water uptake in a desert tree. AoB PLANTS, 2015: 10.1093/aobpla/plv129.
|
| [47] |
Li S, Xiao H L, Zhao L, Zhou M X, Wang F. Foliar water uptake of Tamarix ramosissima from an atmosphere of high humidity. The Scientific World Journal, 2014: 10.1155/2014/529308
|
| [48] |
赵文智, 周宏, 刘鹄. 干旱区包气带土壤水分运移及其对地下水补给研究进展. 地球科学进展, 2017, 32(9): 908-918. |