 2022, Vol. 42
2022, Vol. 42文章信息
- 宋伦, 宋广军, 吴金浩, 杨国军, 刘印, 刘苏萱
- SONG Lun, SONG Guangjun, WU Jinhao, YANG Guojun, LIU Yin, LIU Suxuan
- 长山群岛海域真核微藻粒级结构及扇贝摄食选择性
- Particle size structure of eukaryotic microalgae and feeding selectivity of scallops in the waters of Changshan Islands
- 生态学报. 2022, 42(16): 6838-6852
- Acta Ecologica Sinica. 2022, 42(16): 6838-6852
- http://dx.doi.org/10.5846/stxb202105191311
-
文章历史
- 收稿日期: 2021-05-19
- 网络出版日期: 2022-04-20





2. 大连海洋大学, 大连 116023
2. Dalian Ocean University, Dalian 116023, China
大连长山群岛是海水贝类的主要增养殖区, 近40年来为地方经济发展做出了巨大贡献。但近10年, 增养殖双壳贝类死亡率普遍升高, 严重制约了贝类增养殖业的可持续健康发展。饵料微藻作为滤食性贝类的营养基础, 不同粒级营养含量差异较大, 被滤食性贝类利用率也不同[1—2]。
小粒级微藻会影响贝类营养储备和健康生长, 尤其对质量选择型的贝类影响更大[3]。因此研究小粒级微藻群落结构演变尤为重要, 但目前微藻的分类鉴定依然依靠传统的形态学镜检, 对小粒级微藻的检出率较低。高通量测序鉴定技术可加速小粒径微藻多样性的高效检测[4—7]。目前, 分析海水中微藻粒级的方法用的最多的是叶绿素a分级法, 将被测海水依次通过不同孔径的滤膜过滤, 检测其叶绿素a含量表征微藻粒级生物量, 但该方法会严重低估中、小粒级微藻的生物量, 而高估>20 μm大粒径微藻生物量的贡献, 研究发现, 该方法对小粒径微藻生物量的测算误差最高超过80%[8—10]。分子鉴定法相比传统形态学鉴定更加精准、高效、符合实际, 应用前景广阔。研究发现, 环境样本中真核生物rDNA序列多态性与细胞中rDNA的拷贝数成正比[11—12], 即真核微藻细胞体积(生物量)与rDNA的拷贝数成正比[13—15], 因此, 不同真核微藻分子鉴定获取的rDNA序列数之比即为生物量占比, 可用于微藻全粒级结构深度分析。宋等[16]采用该方法解析了长海县大长山岛邻近海域微藻粒级组成与环境因子的关联。
本研究在采集微藻样品的基础上, 对长山群岛海域微藻粒级结构在时空上的演变, 其与虾夷扇贝摄食选择性等进行分析, 探讨环境因子对各粒级微藻结构的影响关联, 采用高通量测序-分子鉴定分级技术对该海域微藻粒级结构进行估算, 研究海洋微藻粒级时空分布与环境关联及对扇贝摄食选择的影响, 为精准研究滤食性贝类供饵力提供参考。
1 材料与方法 1.1 样品采集及制备2019年在长山群岛海域垂岸布设9个站位(图 1), 分别于2019年5月(春)、8月(夏)、10月(秋)采集真核微藻分子鉴定样品。每个站位采集表层海水1 L, 现场用0.22 μm微孔滤膜收集全部微藻, 然后将滤膜转移至1.5 mL无菌离心管中, 置于-80℃冷冻保存、运输。同期进行海水的水深(Dep)、pH、盐度(Sal)、溶解氧(DO)、悬浮物(SS)、叶绿素a(Chla)、化学需氧量(COD)、石油类(Oil)、无机氮(DIN)[铵盐(NH4+)、硝酸盐(NO3-)、亚硝酸盐(NO2-)之和]、无机磷(DIP)、总氮(TN)、总磷(TP)、硅酸盐(SiO32-)、重金属汞(Hg)、铅(Pb)、铜(Cu)、锌(Zn)、镉(Cd)、铬(Cr)、砷(As)等指标的监测。调查、检测、质量控制方法均参照《海洋调查规范》(GB/T 12763—2007)[17]和《海洋监测规范》(GB17378- 2007)[18]执行。现场在5号站位分别于5、6、7月采集10只活力较好的筏养首层虾夷扇贝, 取出胃含物置于-80℃冷冻保存、运输, 用于微藻分子鉴定;同期采集表层海水1 L, 现场用0.22 μm微孔滤膜收集全部微藻, 然后将滤膜转移至1.5 mL无菌离心管中, 置于-80℃冷冻保存、运输, 用于微藻分子鉴定, 进行贝类摄食选择性分析。

|
| 图 1 采样站位示意 Fig. 1 Sampling sites |
采用CTAB法提取真核微藻宏基因组, 具体参考相关文献[16]。
1.3 18S rDNA V4可变区的PCR扩增利用自行开发的真核微藻18S rDNA的V4区基因扩增引物进行PCR扩增, 具体步骤参考相关文献[19]。
1.4 测序数据质量控制测序所得原始数据处理、有效数据质量过滤、物种聚类、微藻注释等质量控制方法参考相关文献[20]。
1.5 数据分析物种聚类、注释分类采用的方法和数据库参考相关文献[21]。根据相关研究, 不同粒径的真核微藻序列数比例更接近于生物量比例[13—16], 将各个样本微藻优势度超过0.1%的种类作为整体优势种参与粒级结构统计, 根据相关文献将筛查出的优势种按等效球径进行筛检分级[13—15, 22—25], 根据各粒径微藻序列占比统计其粒级结构, 同时利用序列数占比和总叶绿素a浓度可测算各粒级微藻的叶绿素a浓度, 相关公式:
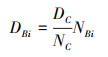
|
式中, DBi为某粒级微藻的叶绿素a浓度(μg/L);NBi为总叶绿素a浓度(μg/L);DC为某粒级微藻的序列数;NC为所有微藻的序列数。
物种生物量优势度(Y)表示微藻群落中某一物种质量所占的优势程度:
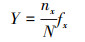
|
式中, nx为第x种微藻种类的OTUs (Operational Taxonomic Units)数, N为OTUs总数, fx为第x种微藻种类在各样品中出现的频率。
采用Ivlev选择指数E来分析贝类对微藻的摄食选择性[26]:
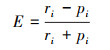
|
式中, ri表示微藻i在贝类胃含物中所占的百分比, pi表示微藻i在水体中的生物量百分比。E范围为[-1, 1], 当E为-1时, 表示贝类避食微藻i;当E为1时, 表示贝类主动选择微藻i;当E接近0时, 表示贝类随机摄食浮游植物i。
本文的小、中、大粒径微藻表述分别代指微微型藻类(0.22—3 μm)、微型藻类(3—20 μm)及小型藻类(>20 μm)。
对上述公式计算、数据分析、RDA排序图均通过WPS Office、SPSS 19.0、PRIMER 5.0软件完成。
2 结果与分析 2.1 测序数据质量和物种注释测序所得有效数据春季每个样品平均获得89616条原始序列, 夏季获得91419条原始序列, 秋季获得83722条原始序列, 经过拼接和质量过滤, 春夏秋季每个样品平均得到78709、81392、76988条有效序列, 高质量序列数均达到87%以上, 测序正确率达到98%以上, 测序数据准确可靠, 可进行OTUs聚类和物种分类分析。数据处理过程中各步骤得到的统计结果见表 1。
| 季节 Seasons |
原始序列数 Original Tags |
拼接序列数 Raw Tags |
过滤低质量和短长度后序列数 Clean Tags(#) |
有效序列数 Effective Tags |
有效碱基数 Base(nt) |
有效序列平均长度 Avglen/nt |
Q20/% | Q30/% | 有效序列数中GC碱基的含量GC/% | 有效序列数占比 Effective/% |
| 春Spring | 89616 | 84256 | 82867 | 78709 | 29517683 | 375 | 98 | 94 | 44.12 | 87.80 |
| 夏Summer | 91419 | 85537 | 84179 | 81392 | 30408251 | 374 | 98 | 93 | 43.21 | 89.00 |
| 秋Autumn | 83722 | 79636 | 78711 | 76988 | 28823487 | 374 | 98 | 93 | 42.79 | 92.00 |
| OTUs:分类操作单元Operational taxonomic units;Q20:表示序列中碱基质量值大于20(测序错误率小于1%)的碱基占比Indicates the proportion of bases with a base quality value greater than 20 (sequencing error rate less than 1%) in the sequence;Q30:表示序列中碱基质量值大于30(测序错误率小于0.1%)的碱基占比Indicates the proportion of bases with a base quality value greater than 30 (sequencing error rate less than 0.1%) in the sequence;GC:鸟嘌呤和胞嘧啶占比The ratio of Guanine and Cytosine OTUs:可操作分类单元Operational Taxonomic Units;Q20:测序质量值Quality, 表示序列中碱基质量值大于20(测序错误率小于1%)的碱基占比;Q30:测序质量值Quality, 表示序列中碱基质量值大于30(测序错误率小于0.1%)的碱基占比;GC:鸟嘌呤和胞嘧啶占比Guanine and cytosine | ||||||||||
长山群岛各粒级微藻优势种及优势度分析见表 2和图 2。其中, 春季融合微胞藻(Micromonas commoda)、曼吉尼刺囊甲藻(Blastodinium mangini)、细小微胞藻(Micromonas pusilla)在小粒级微藻群落中生物量优势度较高, 分别为0.166、0.119、0.114;夏季细小微胞藻、金牛微球藻(Ostreococcus tauri)、曼吉尼刺囊甲藻生物量优势度较高, 分别为0.342、0.257、0.057;秋季金牛微球藻、曼吉尼刺囊甲藻、细小微胞藻生物量优势度较高, 分别为0.263、0.231、0.163。综合来看, 细小微胞藻和金牛微球藻在小粒级微藻群落中占有绝对优势, 春夏秋季演替规律由细小微胞藻占优势转为金牛微球藻控制生态位。由于小粒级微藻粒径较小, 细胞中rDNA的拷贝数较少, 推测其数量优势度在整个微藻群落中占有绝对优势。
| 粒级结构 Size-fraction |
优势种 Dominant species |
春 Spring |
夏 Summer |
秋 Autumn |
| 小型微藻 | 柔弱几内亚藻Guinardia delicatula | 0.245 | — | — |
| Micro- | 北极多甲藻Islandinium minutum | 0.193 | — | — |
| 微型裸甲藻Gymnodinium microreticulatum | 0.151 | — | — | |
| 红色赤潮藻Akashiwo sanguinea | — | 0.092 | — | |
| 平野亚历山大藻Alexandrium hiranoi | — | 0.591 | — | |
| 多纹膝沟藻Gonyaulax polygramma | — | 0.057 | 0.905 | |
| 斯氏多沟藻Polykrikos schwartzii | — | — | 0.035 | |
| 微型微藻 | 剧毒卡尔藻Karlodinium veneficum | 0.457 | 0.362 | 0.678 |
| Nano- | 三叶原甲藻Prorocentrum triestinum | 0.151 | 0.121 | 0.084 |
| 虫黄甲藻Ansanella granifera | 0.048 | 0.128 | — | |
| 旋链海链藻Thalassiosira curviseriata | — | — | 0.048 | |
| 微微型微藻 | 融合微胞藻Micromonas commoda | 0.166 | — | — |
| Pico- | 细小微胞藻Micromonas pusilla | 0.114 | 0.342 | 0.163 |
| 曼吉尼刺囊甲藻Blastodinium mangini | 0.119 | 0.057 | 0.231 | |
| 金牛微球藻Ostreococcus tauri | — | 0.257 | 0.263 |

|
| 图 2 各粒级微藻第一优势种及优势度 Fig. 2 The first dominant species and dominant degree of microalgae in each grain size 物种下方数字为该物种在本粒级中优势度 |
春季剧毒卡尔藻(Karlodinium veneficum)和三叶原甲藻(Prorocentrum triestinum)在中粒级微藻群落中生物量占有绝对优势, 优势度分别为0.457、0.151;夏季剧毒卡尔藻(Karlodinium veneficum), 虫黄甲藻(Ansanella granifera), 三叶原甲藻生物量优势度较高, 分别为0.362、0.128、0.121;秋季剧毒卡尔藻和三叶原甲藻生物量优势度较高, 分别为0.678、0.084。综合来看,各季节剧毒卡尔藻在中粒级微藻群落中一直占有绝对优势。
春季柔弱几内亚藻(Guinardia delicatula), 北极多甲藻(Islandinium minutum), 微型裸甲藻(Gymnodinium microreticulatum), 矮小短棘藻(Detonula pumila), 中肋海链藻(Thalassiosira mala)在大粒级微藻群落中生物量优势度较高, 分别为0.245、0.193、0.151、0.142、0.131;夏季平野亚历山大藻(Alexandrium hiranoi)和红色赤潮藻(Akashiwo sanguinea)生物量优势度较高, 分别为0.591、0.092;秋季多纹膝沟藻(Gonyaulax polygramma)生物量占有绝对优势, 优势度为0.905。大粒级微藻优势种在各季节演替较大, 分别由不同种类占据主要生态位。
综合整个真核微藻种群优势分析, 春季由中粒径的剧毒卡尔藻占有绝对优势, 占比23.9%, 其他微藻占比均小于8%;夏季由大粒径的平野亚历山大藻占有绝对优势, 占比29.4%, 其他微藻占比均小于10%;秋季由大粒径的多纹膝沟藻占有绝对优势, 占比高达66.8%, 其他微藻占比均小于11%。由此可见, 虽然整个微藻群落由大粒级微藻控制生态位, 但都是有毒的甲藻, 对长山群岛海域增养殖的贝类贝毒累积风险较高。
2.3 微藻粒级结构特征图 3给出了春夏秋季不同粒级叶绿素a含量, 春季长山群岛海域微藻生物量由中小粒级控制, 夏季中小粒级生物量减低至与大粒级水平, 秋季大粒级生物量猛增, 主要是由有毒的多纹膝沟藻贡献所致。

|
| 图 3 春、夏、秋季真核微藻各粒级含量 Fig. 3 Size fractions of eukaryotic microalgae in spring, summer and autumn |
真核微藻粒级结构分析结果显示, 春季小粒级微藻组成平均(42±4)%, 中粒级微藻组成平均(47±4)%, 大粒级微藻组成平均(11±1)%(图 4);夏季小粒级微藻组成平均(39±13)%, 中粒级微藻组成平均(23±11)%, 大粒级微藻组成平均(38±21)%, 各站位分布差异比较大(图 5);秋季小粒级微藻组成平均(22±8)%, 中粒级微藻组成平均(18±6)%, 大粒级微藻组成平均(60±8)%, 各站位分布差异比较大(图 6)。

|
| 图 4 春季真核微藻粒级结构 Fig. 4 Size fractions of eukaryotic microalgae in spring |

|
| 图 5 夏季真核微藻粒级结构 Fig. 5 Size fractions of eukaryotic microalgae in summer |

|
| 图 6 秋季真核微藻粒级结构 Fig. 6 Size fractions of eukaryotic microalgae in autumn |
整体而言, 长山群岛海域春季真核微藻以中、小粒级为主, 整体生物量较高, 小、中、大粒级微藻生物量比例为42∶47∶11;夏季以小、大粒级为主, 各站位粒级结构波动较大, 整体生物量较低, 小粒级微藻主要分布在近岸海域, 大粒级微藻主要分布在离岸海域, 小、中、大粒级微藻比例为39∶23∶38;秋季以大粒级为主, 生物量上升较大, 各站位粒级结构相对比较均衡, 小、中、大粒级微藻比例为22∶18∶60。
2.4 扇贝对微藻粒级的摄食选择性采用Ivlev选择指数E对小长山岛海域5—7月虾夷扇贝胃含物与水体中微藻粒级进行了分析。发现筏养虾夷扇贝对小粒级和大粒级微藻的选择性较低, 对中粒级微藻的选择性较高(图 7—9), 尤其对水体中优势种剧毒卡尔藻一直表现出主动选择, 5、6、7月选择性指数均为正值, 分别为0.23、0.39、0.88(表 3)。图 10也显示了虾夷扇贝主要摄食甲藻, 主要种类也为剧毒卡尔藻。虾夷扇贝对水体中优势种小粒级的金牛微球藻、细小微胞藻、抑食金球藻(Aureococcus anophagefferens), 中粒级的阿米巴藻属(Amoebophrya sp.)、球形棕囊藻(Phaeocystis globosa)、雷克斯棕囊藻(Phaeocystis rex), 大粒级的旋转海链藻(Thalassiosira curviseriata)、丝状短棘藻(Detonula confervacea)、多米尼环沟藻(Gyrodinium dominans)、平野亚历山大藻显示出明显回避性, 选择性指数均为负值。

|
| 图 7 虾夷扇贝胃含物与水体中微藻粒级生物量占比 Fig. 7 The proportion of stomach contents and microalgae particle biomass in water of Mizuhopecten yessoensis |

|
| 图 8 虾夷扇贝胃含物与水体中小粒级微藻生物量占比 Fig. 8 Biomass ratio of stomach contents and small and medium sized microalgae in water of Mizuhopecten yessoensis |

|
| 图 9 虾夷扇贝对不同粒级微藻的选择指数 Fig. 9 Selectivity index of Mizuhopecten yessoensis prawns to microalgae of different grain sizes |
| 物种 Species |
粒级 Size-fraction |
类群 Group |
5月May | 6月June | 7月July | ||||||||
| G/% | W/% | E | G/% | W/% | E | G/% | W/% | E | |||||
| 金牛微球藻Ostreococcus tauri | 小 | 绿藻 | 0 | 0.6 | -1.00 | 0 | 0.3 | -1.00 | 0 | 4.2 | -1.00 | ||
| 细小微胞藻Micromonas pusilla | 小 | 绿藻 | 0 | 13.2 | -1.00 | 1.4 | 6.5 | -0.64 | 0 | 23.6 | -1.00 | ||
| 密球藻Pycnococcus provasolii | 小 | 绿藻 | 3.1 | 0.2 | 0.87 | 3.2 | 1.7 | 0.30 | 0 | 1.2 | -1.00 | ||
| 具翅冠突藻Pterosperma cristatum | 小 | 绿藻 | 2.3 | 0.1 | 0.93 | 1.1 | 0.7 | 0.21 | 0 | 0 | -1.00 | ||
| 海洋真核生物克隆Eukaryote marine clone ME1-21 | 小 | 甲藻 | 0 | 3.7 | -1.00 | 1.1 | 0 | 0.92 | 0 | 0 | — | ||
| 海洋真核生物克隆Eukaryote marine clone ME1-22 | 小 | 甲藻 | 0.1 | 0.5 | -0.67 | 12.6 | 2.4 | 0.68 | 1.0 | 1.0 | -0.03 | ||
| 海洋真核生物克隆Eukaryote marine clone MfE1-24 | 小 | 甲藻 | 0.1 | 0.4 | -0.58 | 0.4 | 8.4 | -0.92 | 0 | 4.6 | -1.00 | ||
| 海洋真核生物克隆Eukaryote marine clone ME1-20 | 小 | 甲藻 | 0 | 1.9 | -1.00 | 0 | 2.6 | -1.00 | 0 | 2.4 | -1.00 | ||
| 抑食金球藻Aureococcus anophagefferens | 小 | 金藻 | 0 | 0.3 | -1.00 | 0 | 2.1 | -1.00 | 0 | 0.1 | -1.00 | ||
| 剧毒卡罗藻Karlodinium veneficum | 中 | 甲藻 | 90.9 | 56.3 | 0.23 | 26.4 | 11.5 | 0.39 | 95.1 | 6.0 | 0.88 | ||
| 阿米巴藻属Amoebophrya sp1. | 中 | 甲藻 | 0 | 3.0 | -1.00 | 0 | 11.7 | -1.00 | 0 | 0.5 | -1.00 | ||
| 虫黄甲藻Ansanella granifera | 中 | 甲藻 | 0.8 | 0 | 0.89 | 25.3 | 0.1 | 0.99 | 0 | 0 | -1.00 | ||
| 三角异帽藻Heterocapsa triquetra | 中 | 甲藻 | 0 | 0.1 | -0.75 | 10.8 | 0.1 | 0.98 | 0 | 0 | -1.00 | ||
| 阿米巴藻Amoebophrya sp2. | 中 | 甲藻 | 0 | 0 | — | 0 | 8.8 | -1.00 | 0 | 0.1 | -1.00 | ||
| 球形棕囊藻Phaeocystis globosa | 中 | 定鞭藻 | 0 | 1.1 | -1.00 | 0 | 1.3 | -1.00 | 0 | 7.1 | -1.00 | ||
| 雷克斯棕囊藻Phaeocystis rex | 中 | 定鞭藻 | 0 | 9.6 | -1.00 | 0 | 0.2 | -1.00 | 0 | 0.1 | -1.00 | ||
| 旋转海链藻Thalassiosira curviseriata | 大 | 硅藻 | 0 | 0.4 | -0.96 | 0.4 | 3.8 | -0.82 | 0 | 0.7 | -1.00 | ||
| 斜生四链藻Tetradesmus obliquus | 大 | 硅藻 | 0 | 0 | 1.00 | 1.4 | 0 | 1.00 | 3.9 | 0 | 1.00 | ||
| 斜生四链藻Detonula confervacea | 大 | 硅藻 | 0 | 2.8 | -1.00 | 0 | 5.6 | -1.00 | 0 | 1.7% | -1.00 | ||
| 伪根管藻Rhizosolenia fallax | 大 | 硅藻 | 0 | 0 | 1.00 | 3.6 | 0 | 1.00 | 0 | 0 | — | ||
| 多米尼环沟藻Gyrodinium dominans | 大 | 甲藻 | 0 | 0.3 | -1.00 | 0 | 6.3 | -1.00 | 0 | 0.2 | -1.00 | ||
| 平野亚历山大藻Alexandrium hiranoi | 大 | 甲藻 | 0 | 0.2 | -1.00 | 0 | 0.1 | -1.00 | 0 | 36.7 | -1.00 | ||
| 科夫多沟藻Polykrikos kofoidii | 大 | 甲藻 | 0.2 | 0 | 1.00 | 4.3 | 0 | 1.00 | 0 | 0 | — | ||
| 红色赤潮藻Akashiwo sanguinea | 大 | 甲藻 | 0 | 0 | 1.00 | 2.2 | 0 | 1.00 | 0 | 0 | — | ||
| G:胃含物中微藻占比;W:水体中微藻占比;E:选择指数;“—”:该月扇贝胃含物和水体中均无该藻 | |||||||||||||

|
| 图 10 虾夷扇贝对不同类群微藻的选择指数 Fig. 10 Selectivity index of different groups of microalgae in Mizuhopecten yessoensis |
春、夏、秋季长山群岛真核微藻粒级结构与环境因子的RDA分析见图 11, RDA分析中微藻种类缩写编码见表 4, 环境因子的解释量及显著性检验见表 4。其中, 春季显著控制小粒级微藻群落结构的环境因素有重金属Cd(P < 0.01)、水深和COD(P < 0.05), 显著控制中粒级微藻群落结构的环境因素只有DO(P < 0.05), 显著控制大粒级微藻群落结构的环境因素只有COD(P < 0.05)。

|
| 图 11 春季小粒级微藻群落结构与环境因子RDA排序 Fig. 11 Community structure of microalgae and RDA ranking of environmental factors in spring RDA:冗余分析; COD:化学需氧量; DIP:无机磷; DIN:无机氮; DO:溶解氧; Dep:水深; Oil:石油类; Cd:镉; Kar mic微小卡罗藻Karlodinium micrum; Coc pol:阿米巴藻属有害甲藻Cochlodinium polykrikoides; Hal sp:海球藻属Halosphaera sp.; Bat pra:绿色葡萄藻Bathycoccus prasinos; Euk clone:真核微藻; Euk clo2:真核微藻; Mic Com:融合微胞藻Micromonas commoda; Mic pus:细小微胞藻Micromonas pusilla; Pte cri:具翅冠突藻Pterosperma cristatum; Cer fal:共甲藻Ceratoperidinium falcatum; Ost tau:金牛微球藻Ostreococcus tauri; Pyc pro:密球藻Pycnococcus provasolii; Bla man:曼吉尼刺囊甲藻Blastodinium mangini; Aur ano:抑食金球藻Aureococcus anophagefferens; Pra sp:青绿藻属Prasinophyceae sp.; Pro mic:海洋原甲藻Prorocentrum micans; Pro min:微小原甲藻Prorocentrum minimum; Eud cre:波缘杜波斯克藻Euduboscquella crenulata; Ima rot:棕囊藻属Imantonia rotunda; Mam gil:青绿藻Mamiella gilva; Bla con:扭曲刺囊甲藻Blastodinium contortum; Amo sp:阿米巴藻属Amoebophrya sp.; Mar sp:共球藻属Marsupiomonas sp.; Fav arc:杜波斯克属Favella arcuata |
| 季节 Seasons |
粒级 Size-fraction |
环境因子 Environmental factors |
环境因子解释量 Interpretation quantity |
pseudo-F | P |
| 春 | 小 | 水深 | 26.7 | 3.6 | 0.014 |
| 镉 | 18.7 | 3.1 | 0.010 | ||
| 化学需氧量 | 11.2 | 2.1 | 0.046 | ||
| 中 | 溶解氧 | 34.3 | 5.2 | 0.046 | |
| 大 | 化学需氧量 | 15.6 | 3.5 | 0.030 | |
| 夏 | 小 | 镉 | 28 | 3.9 | 0.014 |
| 中 | 无机氮 | 35.9 | 5.6 | 0.022 | |
| 石油烃 | 10 | 6.9 | 0.032 | ||
| 大 | 砷 | 18.9 | 4.6 | 0.042 | |
| 秋 | 小 | 盐度 | 73.4 | 27.5 | 0.010 |
| 化学需氧量 | 13 | 8.6 | 0.010 | ||
| 中 | 石油烃 | 38.8 | 6.3 | 0.024 | |
| 大 | 汞 | 50 | 10 | 0.006 | |
| 溶解氧 | 1.4 | 11.6 | 0.028 |
夏季显著控制小粒级微藻群落结构的环境因素有重金属Cd和DIN(P < 0.05), 显著控制中粒级微藻群落结构的环境因素有DIN、pH、Oil(P < 0.05), 显著控制大粒级微藻群落结构的环境因素有重金属As和盐度Sal(P < 0.05)。
秋季显著控制小粒级微藻群落结构的环境因素有COD和Sal(P < 0.01), 显著控制中粒级微藻群落结构的环境因素只有Oil(P < 0.05), 显著控制大粒级微藻群落结构的环境因素有重金属Hg(P < 0.01)和DO(P < 0.05)。总体来看, COD、DIN、DO、Oil及部分重金属Cd、As、Hg影响着整个真核微藻粒级结构时空演变。
3 讨论 3.1 真核微藻优势种及粒级结构演变本研究发现, 长山群岛海域第一优势种均为有毒甲藻, 春季为剧毒卡尔藻占比23.9%, 夏季为平野亚历山大藻占比29.4%, 秋季为多纹膝沟藻占比66.8%, 均高于黄海北部优势种平均值(3种优势种分别占比21.3%、23.6%、56.6%)[7], 说明该海域为有毒甲藻主要分布区, 应引起注意。从粒级结构看, 长山群岛海域小粒级真核微藻占比(春季42%、夏季39%、秋季22%)与黄海北部整体水平(39%、夏季40%、秋季26%)相差不大, 春夏季略微偏高, 不利于滤食性的贝类营养储备[7]。本次调查的小粒级真核微藻优势种主要为细小微胞藻和金牛微球藻, 而2018年大长山岛临近海域调查的优势种为抑食金球藻[16], 虽然优势种年度间发生演替, 但金牛微球藻和抑食金球藻作为褐潮致灾种, 应引起关注[10, 19, 27], 当然, 微藻粒级结构的测算方法需不断完善和优化, 使研究结果更趋于客观实际[3, 9, 16]。
3.2 不同微藻粒级对扇贝摄食的影响本研究发现筏养虾夷扇贝对小粒级和大粒级微藻的选择性都较低, 对中粒级微藻的选择性确较高, 尤其对水体中优势种甲藻剧毒卡尔藻一直表现出主动选择。有研究表明, 滤食性贝类对甲藻的摄食选择性要高于其他类群[26]。本研究发现, 同样作为水体中的优势种, 小粒级的金牛微球藻、细小微胞藻、抑食金球藻, 虾夷扇贝却选择避食, 表明扇贝摄食选择的复杂性。Cranford等[28]发现滤食性贝类偏好摄食微型藻类, Strohmeier等[29]发现紫贻贝(Mytilus edulis)对藻类的截留率从小粒径到大粒径逐渐增大, 最大的截留率在7—35 μm。Rosa等[30]研究发现贻贝对4 μm及以上的藻类有较高的截留率, Dunphy[31]也发现牡蛎6 μm微藻的截留效率低于15 μm粒径的微藻。张继红[32]发现紫贻贝对于2 μm的微藻截留率为19%、长牡蛎为17%、栉孔扇贝仅为8%, 对2 μm以下的颗粒物截留效率更低。上述研究结论与本研究发现的现象一致。该海域作为虾夷扇贝主产区, 不但微藻粒级结构影响其摄食营养与存活[33], 有毒藻类更影响其质量安全, 因此应推进微藻分子鉴定法应用, 并加强贝毒潜在风险监测[34]。

|
| 图 12 春季中粒级微藻群落结构与环境因子RDA排序 Fig. 12 Community structure of medium size microalgae and RDA ranking of environmental factors in spring Sal:盐度;SS:悬浮物;Pro tri:三叶原甲藻Prorocentrum triestinum;Cha sp:角毛藻属Chaetoceros sp.;Tha con:凹海链藻Thalassiosira concaviuscula;Tha nor:诺登海链藻Thalassiosira nordenskioeldii;Gui str:斯氏几内亚藻Guinardia striata;Ske sub:敏盐骨条藻Skeletonema subsalsum;Ast gla:冰河拟星杆藻Asterionellopsis glacialis;Ske men:曼氏骨条藻Skeletonema menzellii;Cha mue:牟氏角毛藻Chaetoceros muellerii;Tha nod:结线形海链藻Thalassiosira nodulolineata;Pyr sp:青绿皮藻属Pyramimonas sp.;Chr lea:里氏金色藻Chrysochromulina leadbeateri;Ans gra:虫黄甲藻Ansanella granifera;Kar ven:剧毒卡尔藻Karlodinium veneficum;Nep pyr:双鞭绿藻Nephroselmis pyriformis;Spu sp:色金藻属Spumella sp.;Tha pro:深海链藻Thalassiosira profunda;Chl kuw:库瓦达衣藻Chlamydomonas kuwadae;Ske tro:热带骨条藻Skeletonema tropicum;Hem cry:球半隐藻Hemiselmis cryptochromatica |
水体中环境因子对微藻生长、繁殖影响较大, 进而控制其种群及群落在时空上的演变。本研究发现, 不同季节控制各粒级微藻群落结构空间分布的环境因素不尽相同, 总体而言, COD(春秋季)对小粒级微藻群落结构影响较大, 说明某些小粒级微藻可对有机物进行高效利用[10], 一般真核微藻都偏好无机营养盐, 如无机氮、磷、硅都是微藻生长的必须元素[35—37]。同时发现春夏季重金属Cd都显著控制小粒级微藻群落结构在空间上的分布, 且对小粒级真核微藻优势种细小微胞藻和金牛微球藻均是正向影响, 应加强河流陆源入海重金属的消减防控。当然, 微藻粒级结构演变除受环境因子的上行效应影响之外, 贝类及浮游动物大量摄食的下行效应也会影响微藻的粒级结构[38—39]。该海域几十年来主要增养殖扇贝、牡蛎等滤食性贝类, 对天然饵料微藻需求量较大。随着陆源氮磷的消减, 海域无机营养盐逐渐减少, 可能会加剧微藻小型化趋势, 希望能得到有关方面的关注。
4 结论(1) 长山群岛海域真核微藻春季以中(47%)、小粒级(41%)为主, 主要优势种为剧毒卡尔藻(23.9%);夏季以小(39%)、大粒级(38%)为主, 主要优势种为平野亚历山大藻(29.4%);秋季以大粒级(60%)为主, 主要优势种为多纹膝沟藻(66.8%)。

|
| 图 13 春季大粒级微藻群落结构与环境因子RDA排序 Fig. 13 Community structure of macroalgae and RDA ranking of environmental factors in spring Gym mic:微型裸甲藻Gymnodinium microreticulatum;Lev fis:菲斯环沟藻Levanderina fissa;Per qui:五刺多甲藻Peridinium quinquecorne endosymbiont;Oxy oxy:高氧鳍藻Oxyphysis oxytoxoides;Tha mal:中肋海链藻Thalassiosira mala;Ale aff :相关亚历山大藻Alexandrium affine;Pse mul :多列拟菱形藻Pseudo-nitzschia multiseries;Gra ton:通洋多甲藻Grammatodinium tongyeonginum;Cer pel:大洋角管藻Cerataulina pelagica;Gui del:柔弱几内亚藻Guinardia delicatula;Tha rot:圆海链藻Thalassiosira rotula;Gon spi:具刺膝沟藻Gonyaulax spinifera;Det pum :矮小短棘藻Detonula pumila;Par imp:无孔近囊胞藻Paraphysomonas imperforata |

|
| 图 14 夏季小粒级微藻群落结构与环境因子RDA排序 Fig. 14 Community structure of microalgae in summer and RDA ranking of environmental factors |

|
| 图 15 夏季中粒级微藻群落结构与环境因子RDA排序 Fig. 15 Community structure of medium size microalgae and RDA ranking of environmental factors in summer |

|
| 图 16 夏季大粒级微藻群落结构与环境因子RDA排序 Fig. 16 Community structure of macroalgae and RDA ranking of environmental factors in summer Ale lee:李氏亚历山大藻Alexandrium leei;War sp:单眼藻属Warnowia sp.;Pol sch:斯氏多沟藻Polykrikos schwartzii;Aka san:红色赤潮藻Akashiwo sanguinea;Ale hir:平野亚历山大藻Alexandrium hiranoi;Pol kof :科夫多沟藻Polykrikos kofoidii;Isl min:北极多甲藻Islandinium minutum;Tha hen:亨氏海链藻Thalassiosira hendeyi;Cos sp:圆筛藻属Coscinodiscus sp;Gon pol:多纹膝沟藻Gonyaulax Polygramma;Pyr ste:斯氏扁甲藻Pyrophacus steinii |

|
| 图 17 秋季小粒级微藻群落结构与环境因子RDA排序 Fig. 17 Community structure of microalgae and RDA ranking of environmental factors in autumn |

|
| 图 18 秋季中粒级微藻群落结构与环境因子RDA排序 Fig. 18 Community structure of medium grain microalgae and RDA ranking of environmental factors in autumn |
(2) 小粒径微藻中金牛微球藻和抑食金球藻曾在渤海引发褐潮, 潜在威胁贝类养殖业。另外有毒甲藻在该海域中占有绝对优势, 中粒径的剧毒卡尔藻在春季优势明显, 优势度在0.2以上, 贝毒累积风险较高。
(3) 虾夷扇贝对小粒级和大粒级微藻的选择性较低, 对中粒级微藻的选择性较高, 尤其对水体中优势种剧毒卡尔藻一直表现出主动选择。

|
| 图 19 秋季大粒级微藻群落结构与环境因子RDA排序 Fig. 19 Community structure of macroalgae and RDA ranking of environmental factors in autumn Lau bor:北方劳德藻Lauderia borealis |
(4) COD、DIN、DO、Oil及部分重金属Cd、As、Hg影响着整个长山群岛海域真核微藻粒级结构时空演变。
| [1] |
宋伦, 宋广军, 王年斌, 赵海勃, 田金, 杨爽, 杜静. 辽东湾网采浮游植物粒级结构的胁迫响应. 中国环境科学, 2015, 35(9): 2764-2771. DOI:10.3969/j.issn.1000-6923.2015.09.031 |
| [2] |
董波, 薛钦昭, 李军. 滤食性贝类摄食生理的研究进展. 海洋科学, 2000, 24(7): 31-34. DOI:10.3969/j.issn.1000-3096.2000.07.012 |
| [3] |
吴文广, 张继红, 刘毅, 王巍, 蔺凡. 獐子岛及邻近海域秋季浮游植物的粒级结构及其影响因素. 生态学报, 2018, 38(4): 1418-1426. |
| [4] |
乔玲, 甄毓, 米铁柱. 抑食金球藻(Aureococcus anophagefferens)褐潮研究概述. 海洋环境科学, 2016, 35(3): 473-480. |
| [5] |
刘卫东, 宋伦, 吴景. 环境样本中微型和微微型浮游植物高通量测序的引物优化. 生态学报, 2017, 37(12): 4208-4216. |
| [6] |
Kong F Z, Yu R C, Zhang Q C, Yan T, Zhou M J. Pigment characterization for the 2011 bloom in Qinhuangdao implicated "brown tide" events in China. Chinese Journal of Oceanology and Limnology, 2012, 30(3): 361-370. DOI:10.1007/s00343-012-1239-z |
| [7] |
宋伦, 毕相东, 付杰, 宋广军, 吴金浩, 刘印, 刘苏萱. 黄海北部真核微藻粒级结构及环境关联. 中国环境科学, 2021, 41(3): 1336-1344. DOI:10.3969/j.issn.1000-6923.2021.03.037 |
| [8] |
孙军, 刘东艳, 钟华, 张利永. 浮游植物粒级研究方法的比较. 青岛海洋大学学报, 2003, 33(6): 917-924. DOI:10.3969/j.issn.1672-5174.2003.06.010 |
| [9] |
Song L, Wu J, Du J, Li N, Wang K, Wang P. Comparison of two methods to assess the size structure of phytoplankton community assemblages, in Liaodong Bay, China. Journal of Ocean University of China, 2019, 18(5): 1207-1215. DOI:10.1007/s11802-019-3960-0 |
| [10] |
宋伦, 吴景, 李楠, 杜静, 杨爽, 王鹏. 辽东湾潜在褐潮生物时空分布及环境关联. 中国环境科学, 2018, 38(8): 3060-3071. DOI:10.3969/j.issn.1000-6923.2018.08.035 |
| [11] |
郭立亮. 海洋微藻DNA条形码基因的评估及环境样本多样性分析和定量基因的开发[D]. 青岛: 中国海洋大学, 2015.
|
| [12] |
Gong J, Dong J, Liu X H, Massana R. Extremely high copy numbers and polymorphisms of the rDNA operon estimated from single cell analysis of Oligotrich and Peritrich ciliates. Protist, 2013, 164(3): 369-379. DOI:10.1016/j.protis.2012.11.006 |
| [13] |
Zhu F, Massana R, Not F, Marie D, Vaulot D. Mapping of picoeucaryotes in marine ecosystems with quantitative PCR of the 18S rRNA gene. FEMS Microbiology Ecology, 2005, 52(1): 79-92. DOI:10.1016/j.femsec.2004.10.006 |
| [14] |
Prokopowich C D, Gregory T R, Crease T J. The correlation between rDNA copy number and genome size in eukaryotes. Genome, 2003, 46(1): 48-50. DOI:10.1139/g02-103 |
| [15] |
Godhe A, Asplund M E, Harnstrom K, Saravanan V, Tyagi A, Karunasagar I. Quantification of diatom and Dinoflagellate biomasses in coastal marine seawater samples by real-time PCR. Applied and Environmental Microbiology, 2008, 74(23): 7174-7182. DOI:10.1128/AEM.01298-08 |
| [16] |
宋伦, 毕相东, 宋广军, 杜静, 吴金浩, 王志松, 胡超魁. 海洋真核微藻粒级结构及其环境影响因素. 中国环境科学, 2020, 40(6): 2627-2634. DOI:10.3969/j.issn.1000-6923.2020.06.033 |
| [17] |
中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会. GB/T 12763—2007海洋调查规范. 北京: 中国标准出版社, 2008.
|
| [18] |
中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会. GB 17378—2007海洋监测规范. 北京: 中国标准出版社, 2008.
|
| [19] |
宋伦, 吴景, 刘卫东, 宋永刚, 王年斌. 渤海长兴岛海域微型和微微型浮游植物多样性. 环境科学研究, 2016, 29(11): 1635-1642. |
| [20] |
Song L, Wu J, Du J, Li N, Song G J, Wang K, Sun M, Wang P. The characteristics and distribution of eukaryotic phytoplankton community in Liaodong Bay, China. Ocean Science Journal, 2019, 54(2): 183-203. DOI:10.1007/s12601-019-0007-9 |
| [21] |
Quast C, Pruesse E, Yilmaz P, Gerken J, Schweer T, Yarza P, Peplies J, Glöckner F O. The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Research, 2013, 41(D1): D590-D596. |
| [22] |
刘瑞玉. 中国海洋生物名录. 北京: 科学出版社, 2008: 301-870.
|
| [23] |
Xu X, Yu Z M, Cheng F J, He L Y, Cao X H, Song X X. Molecular diversity and ecological characteristics of the eukaryotic phytoplankton community in the coastal waters of the Bohai Sea, China. Harmful Algae, 2017, 61: 13-22. DOI:10.1016/j.hal.2016.11.005 |
| [24] |
于杰. 浮游生物多样性高效检测技术的建立及其在渤海褐潮研究中的应用[D]. 青岛: 中国海洋大学, 2014.
|
| [25] |
江雪娇. 北黄海微微型浮游植物的丰度及微微型真核浮游生物分子多样性研究[D]. 青岛: 中国海洋大学, 2009.
|
| [26] |
潘绘竹. 黄渤海海域几种典型双壳贝类对浮游植物的摄食规律研究[D]. 广州: 暨南大学, 2020.
|
| [27] |
宋伦, 吴景, 宋永刚, 刘卫东, 杨国军. 褐潮致灾种抑食金球藻在辽东湾的分布. 环境科学研究, 2017, 30(4): 537-544. |
| [28] |
Cranford P J, Li W, Strand Ø, Strohmeier T. Phytoplankton depletion by mussel aquaculture: high resolution mapping, ecosystem modeling and potential indicators of ecological carrying capacity. ICES, 2008.
|
| [29] |
Strohmeier T, Strand Ø, Alunno-Bruscia M, Duinker A, Cranford P J. Variability in particle retention efficiency by the mussel Mytilus edulis. J Exp Mar Biol Ecol, 2012, 412(31): 96-102. |
| [30] |
Rosa M, Ward J E, Ouvrard M, Holohan B A, Espinosa E P, Shumway S E, Allam B. Examining the physiological plasticity of particle capture by the blue mussel, Mytilus edulis (L.): confounding factors and potential artifacts with studies utilizing natural seston. J Exp Mar Biol Ecol, 2015, 473: 207-217. DOI:10.1016/j.jembe.2015.09.005 |
| [31] |
Dunphy B J, Hall J A, Jeffs A G, Wells R M G. Selective particle feeding by the Chilean oyster, Ostrea chilensis; implications for nursery culture and broodstock conditioning. Aquaculture, 2006, 261: 594-602. DOI:10.1016/j.aquaculture.2006.08.015 |
| [32] |
张继红. 滤食性贝类养殖活动对海域生态系统的影响及生态容量评估[D]. 青岛: 中国科学院研究生院(海洋研究所), 2008.
|
| [33] |
张继红, 吴文广, 刘毅, 蔺凡, 王巍, 牛亚丽. 虾夷扇贝动态能量收支生长模型. 中国水产科学, 2017, 24(3): 497-506. |
| [34] |
宋伦, 吴景, 李楠, 杜静, 王昆, 刘桂英, 王鹏. 辽东湾毒害微藻潜在风险分布及环境关联. 中国环境科学, 2018, 38(8): 3049-3059. DOI:10.3969/j.issn.1000-6923.2018.08.034 |
| [35] |
Luck M, Escobar F V, Glass K, Sabotke M I, Hagedorn R, Corellou F, Siebert F, Hildebrandt P, Hegemann P. Photoreactions of the histidine kinase rhodopsin Ot-HKR from the marine picoalga Ostreococcus tauri. Biochemistry, 2019, 58(14): 1878-1891. DOI:10.1021/acs.biochem.8b01200 |
| [36] |
Echeveste P, Agustí S, Dachs J. Cell size dependent toxicity thresholds of polycyclic aromatic hydrocarbons to natural and cultured phytoplankton populations. Environmental Pollution, 2010, 158(1): 299-307. DOI:10.1016/j.envpol.2009.07.006 |
| [37] |
Sunda W G, Huntsman S A. Effect of Zn, Mn, and Fe on Cd accumulation in phytoplankton: implications for oceanic Cd cycling. Limnology and Oceanography, 2000, 45(7): 1501-1516. DOI:10.4319/lo.2000.45.7.1501 |
| [38] |
蒋昊, 赵亮, 张晶. 黄海溶解无机氮及颗粒有机氮收支与转化模型. 中国环境科学, 2020, 40(9): 3981-3991. DOI:10.3969/j.issn.1000-6923.2020.09.031 |
| [39] |
张雪, 宋伦, 付杰, 王昆, 刘印, 刘苏萱. 黄海北部滤食性贝类供饵力区划研究. 水产科学, 2021, 40(3): 416-421. |