 2022, Vol. 42
2022, Vol. 42文章信息
- 张子琦, 焦菊英, 陈同德, 林红, 陈玉兰, 徐倩, 程玉卓, 赵文婷
- ZHANG Ziqi, JIAO Juying, CHEN Tongde, LIN Hong, CHEN Yulan, XU Qian, CHENG Yuzhuo, ZHAO Wenting
- 拉萨河流域洪积扇不同植被类型土壤化学计量特征
- Soil stoichiometry characteristics of different vegetation types in alluvial fans of the Lhasa River Basin
- 生态学报. 2022, 42(16): 6801-6815
- Acta Ecologica Sinica. 2022, 42(16): 6801-6815
- http://dx.doi.org/10.5846/stxb202105171292
-
文章历史
- 收稿日期: 2021-05-17
- 网络出版日期: 2022-04-20





2. 中国科学院 水利部 水土保持研究所, 杨凌 712100
2. Institute of Soil and Water Conservation, Chinese Academy of Sciences and Ministry of Water Resources, Yangling 712100, China
碳(C)、氮(N)、磷(P)、钾(K)是植物生长所需的最重要的生命元素。C是构成生物体的骨架元素[1], 土壤是陆地生态系统最大的C库, 土壤中储存的C大约是植被的3倍, 大气的2倍, 因此土壤C储量较小幅度的变化就会引起大气CO2的巨大波动[2];N是调节陆地生态系统生产量、结构和功能的关键元素, 也是限制群落生产力的常见元素[1], 土壤中植物可直接利用的N素形态主要为NO3-和NH4+, 仅占土壤全N的1%左右;P是植物生长代谢过程中的必需元素[3], 土壤中全P的含量很低(约为0.02%—2%), 我国土壤普查估算全国近七成的土壤缺P;K是与植物体新陈代谢有关的重要元素, 通常是草地和寒带森林生态系统的限制性养分元素[4]。土壤和植物是陆地生态系统的重要组成部分, 二者相互作用, 相互影响。土壤为植被提供了生长条件, 土壤有机质和N、P、K等养分直接影响植被的生长发育, 植被在其生长发育过程中又通过凋落物的归还以及根系的分泌等方式反过来影响土壤的理化性质。
近年来生态化学计量比越来越受到生态学家的关注, 且被认为是生态过程中养分循环的重要指标, 可以有效地反映出生态系统中的养分限制状况[5—6]。生态化学计量学是在生态学背景下, 结合生态学与化学计量学的基本原理, 将各种构成有机体的基本元素联系起来, 是研究生态系统能量平衡和多重化学元素平衡的科学[7], 可以分析多重化学元素的质量平衡对生态交互作用的影响。土壤C∶N∶P直接反映了土壤肥力状况, 其与植物变量的联系也常被用来测定植物所处生态系统的限制性养分元素[6, 8]。土壤C∶N与土壤微生物的活动和繁殖有关, 当C∶N>25时, 表现为N素不足, 微生物与植物争夺土壤中的N素, 从而导致土壤缺N;土壤C∶N值过低则表明土壤有机质矿化速率较高, 土壤肥力水平低。土壤C∶P不仅与土壤有机质的分解相关, 还可以作为判别土壤P有效性高低的一个指标, 若土壤C∶P低, 则相应的土壤P的有效性高[9]。N∶P化学计量在陆地生态系统中的研究首先在植物组织中取得了重大发现, Koerselman和Meulman[10]发现植物叶片N∶P<14时表现为N限制, N∶P>16时表现为P限制, N∶P在14—16之间表现为N和P共同限制。而后发现土壤与植物组织中的N∶P显著相关[11], 又因土壤N∶P的测量相对方便, 所以对于土壤N∶P的研究蓬勃发展。例如, Bedford等[12]将植物叶片的N∶P养分限制阈值应用于北美温带沼泽土壤养分限制的研究; Yan和Lu[13]对青藏高原高寒草地的研究显示, 土壤N∶P与植物地上生物量显著正相关, 与物种多样性显著负相关;Shen等[14]通过对杉木人工林的长期施N实验表明土壤N∶P可作为判别土壤N和P限制的指标, 可以反映土壤微生物群落组成和土壤微生物活性。植物N∶K是生态系统组成和物种多样性变化的原因之一, Roem和Berendse[15]通过植物N∶K和N浓度与植被地上生物量的回归分析表明, 当N∶K值为1—1.5时植物有最大的生产量, 而目前对土壤N∶K化学计量的研究较少。
拉萨河流域属于藏南及喜马拉雅中段生态安全屏障区中的雅鲁藏布江中游宽谷土地沙化和水土流失控制及经济重点发展亚区, 具有重要的生态功能和独特的生物区系[16]。洪积扇是拉萨河流域珍贵的土地资源, 随着人口数量日益增加, 人地矛盾突出, 许多村落依洪积扇而建, 洪积扇已经成为人类进行农业生产与放牧的重要场地[17], 而对土壤养分状况的研究是对洪积扇进行科学合理开发利用的基础。此外, 目前国内在青藏高原地区对土壤生态化学计量特征的研究多集中于高寒草地[18]、高寒草甸[19]以及高山林草交错带[20], 而对洪积扇土壤与植被的相互关系及生态化学计量特征的研究较少。为此, 本研究选取拉萨河流域的10个洪积扇, 研究其上土壤有机碳(SOC)、全氮(TN)、全磷(TP)、全钾(TK)、碱解氮(AN)、速效磷(AP)、速效钾(AK)的含量及其化学计量比随植被类型的变化特征, 探究土壤生态化学计量比对植物群落物种组成的影响, 分析拉萨河流域洪积扇植被在生长过程中受到的养分限制状况, 为洪积扇植被的保护与可持续发展提供理论依据。
1 材料与方法 1.1 研究区概况拉萨河发源于念青唐古拉山脉中段北侧的罗布如拉, 沿途流经墨竹工卡县, 达孜区, 最后经过拉萨市, 在拉萨市曲水县汇入雅鲁藏布江, 全长568 km, 流域面积31760 km2, 平均海拔5200 m, 属于典型的高原温带半干旱季风气候, 是世界上海拔最高的河流之一, 位于29°20′—31°15′N、90°05′—93°20′E之间, 气候寒冷干燥, 年平均气温-7.1—9.2℃, 极端气温-14℃和31℃。年平均降雨量400—500 mm, 大多数降雨集中在夏季(5—9月), 冬天干旱几乎无雪。土壤类型以草毡土、草甸土及黑毡土为主, 还包括寒冻土、棕冷钙土、沼泽土等多种土壤类型, 各类型土壤中均含有较多的石砾;植被类型主要有山地稀疏森林、山地灌丛草原、寒冷半湿润高山草甸、灌丛及流石滩稀疏植被等[21]。
1.2 野外调查野外调查前, 在室内通过Google Earth对拉萨河流域内的洪积扇进行提取与选择, 综合考虑空间分布和土地利用, 结合实地情况, 最终选取了拉萨河流域的10个典型洪积扇作为调查对象。其主要分布于堆龙德庆、当雄、林周、墨竹工卡4县区辖区内(图 1), 各洪积扇基本状况见表 1。

|
| 图 1 研究样区分布图 Fig. 1 Distribution map of study sample area |
| 洪积扇 Alluvial fans |
经度 Longitude |
纬度 Latitude |
海拔 Elevation/m |
面积 Area/km2 |
干扰因素 Interference factors |
| 1 | 90°51′ | 29°42′ | 3857 | 2.58 | 耕种+放牧 |
| 2 | 91°10′ | 29°59′ | 3929 | 1.39 | 耕种+放牧 |
| 3 | 91°39′ | 30°04′ | 3934 | 4.41 | 耕种+放牧 |
| 4 | 91°50′ | 30°11′ | 4479 | 0.42 | 放牧 |
| 5 | 92°08′ | 30°03′ | 4300 | 0.20 | 耕种+放牧 |
| 6 | 92°20′ | 30°07′ | 4413 | 0.09 | 放牧 |
| 7 | 90°45′ | 30°01′ | 4070 | 1.62 | 耕种+放牧 |
| 8 | 90°36′ | 30°11′ | 4525 | 1.26 | 耕种+放牧 |
| 9 | 90°46′ | 30°20′ | 4435 | 49.75 | 放牧 |
| 10 | 90°07′ | 30°32′ | 4435 | 0.90 | 放牧 |
于2020年7—8月在上述区域进行了植被调查和土壤样本采集。调查过程中根据各个洪积扇的植被分布状况, 共布设20个样地, 每个样地按洪积扇的上中下布设3个样点作为重复, 其中:自然草本样地9个, 布设2 m×2 m的样方, 调查其内草本的种名、高度、多度以及盖度;灌丛样地5个, 布设5 m×5 m的样方, 调查其内灌丛的种名、高度、多度、盖度、冠幅, 灌丛下的草本调查同草本样地;农田样地6个, 布设2 m×2 m的样方, 调查其内农作物的种名、高度、盖度以及农田田坎上草本的种名;土壤取样时在各个样方内遵循S形取样法, 选5点采取表层0—20 cm土壤, 然后将采取的土样混合, 依次过20 mm筛、10 mm筛除去石块和植物根系后装入布袋中带回实验室, 在实验室风干之后将样品过1 mm筛用于土壤AN, AP和AK的测定;过0.149 mm筛用于土壤TN、TP、TK和SOC的测定。
1.3 土壤性质测定SOC采用K2Cr2O4容量法测定;土壤TN是土样经硫酸-催化剂消解后用全自动定氮仪测定(型号:福斯Kjetltec8400);土壤TP是土样加入硝酸、高氯酸和氢佛酸蒸干, 然后加水摇匀消解后用AA3全自动连续流动分析仪测定(Seal AA3, 产地:德国);土壤TK是土样经硝酸、高氯酸和氢佛酸消解后用火焰光度计测定(上海分析仪器有限公司FP640);土壤AN用碱解扩散法测定(国标);土壤AP是土样经0.5 mol/L浸提后用UV-1780紫外分光度计(岛津)测定;土壤AK用火焰光度计法测定(上海分析仪器有限公司FP6410)。
1.4 数据分析重要值是以综合数值表示植物种在群落中的相对重要性。本研究中各物种的重要值(IV, %)通过如下公式计算[22]:

|
(1) |
式中, 相对盖度为样方中某物种的盖度与样方中全部物种盖度之和的比值;相对多度为样方中某物种的个体数与样方中全部物种的个体总数的比值;相对高度为样方中某物种的平均高度与样方中全部物种平均高度之和的比值。
植物多样性指数是研究植物群落构造和功能的重要参数之一, 本研究选取物种数S、Shannon-Winner多样性指数、Margalef丰富度指数和Pielou均匀度指数来研究洪积扇植物群落的多样性特征, 其中Shannon-Wiener多样性指数综合表征群落物种组成的丰富度及均匀度, 其值越大, 群落内物种越丰富;物种数S和Margalef指数表征群落内物种数的多少, 其值越大, 表明群落内物种越多;Pielou指数表征群落内各物种分布的相对多度, 其值越大, 表明群落内物种分布越均匀。其计算公式分别如下[22]:
Margalef丰富度指数:
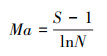
|
(2) |
Shannon-Wiener多样性指数:
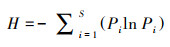
|
(3) |
Pielou均匀度指数:
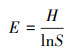
|
(4) |
式中, S为样方中物种的种数, N为样方中全部物种的个体总数, Pi为物种i的个体数占全部物种个体总数的比例。
SOC、TN、TP、TK的含量及其化学计量比的空间变异性用变异系数(COV)表示, 其计算公式如下[23]:

|
(5) |
式中, COV < 10%属于弱变异, 10% < COV < 100%属于中等变异, COV>100%属于强变异。
本研究中的化学计量比值均为土壤全量养分的质量比。
采用SPSS 26.0软件进行Pearson相关性分析, 探究土壤养分之间以及土壤养分和植物群落特征值之间的相关关系;采用Duncan显著性检验分析不同植被类型下土壤养分含量及其化学计量比之间的差异(P < 0.05);通过双因素方差分析(Two-way ANOVA)研究洪积扇和植被类型及其交互作用对土壤养分及其化学计量比的影响;采用CANOCO 5.0软件进行典范对应分析(Canonical Correspondence Analysis, CCA), 以物种多度数据作为响应变量, 土壤养分及化学计量比值作为解释变量, 探究土壤生态化学计量对植物群落物种组成变化的解释程度。本文中的图件利用Origin 2018和R 4.0.5绘制。
2 结果与分析 2.1 植被类型及物种组成特征本研究调查分析结果显示(表 2和表 3), 10个洪积扇的60个样方共发现87种植物, 分属于29科79属, 其中以禾本科和菊科的物种居多, 分别占总属数的19.0%和13.9%, 总种数的17.2%和16.1%。本调查中, 发现自然草本群落共8种, 包含有水葱(Scirpus validus)群落、中华草沙蚕(Tripogon chinensis)群落、绢毛委陵菜(Potentilla sericea)群落、高原毛茛(Ranunculus tanguticus)群落、草血竭(Polygonum paleaceum)群落、牛筋草(Eleusine indica)群落、鼠麴草(Gnaphalium affine)群落和黄耆(Astragalus membranaceus)群落, 草本种类相对丰富, 总计27科74属81种。其中, 水葱几乎在各个样地的重要值均较高, 水葱喜湿, 多生长于湖边或浅水塘中, 说明拉萨河流域洪积扇的水分条件较好。灌丛样地共5个, 灌丛种类少, 仅发现6种灌木, 分属于3科5属, 包含小檗(Berberis thunbergii)+扁刺峨眉蔷薇(Rosa omeiensis f. pteracantha)群落、小檗+金露梅(Potentilla fruticosa)群落、小檗+高山柏(Sabina squamata)+扁刺峨眉蔷薇群落、扁刺峨眉蔷薇群落和金露梅群落等, 自然灌丛生长状况较差, 覆盖度小于50%, 多分布于河流两侧、山脚, 其下草本种类多且覆盖度高, 但大多仅有单一的优势种存在。农田样地共6个, 农作物主要为青稞(Hordeum vulgare var. nudum)和油菜(Brassica napus), 分别属于禾本科大麦属和十字花科芸薹属, 农作物种植密度大, 除9号农田样地外, 其余农田样地盖度在80%左右。本调查中各草本群落Shannon-Wiener多样性指数、Margalef丰富度指数和Pielou均匀度指数的均值分别为1.35、1.67和0.54;灌丛群落特征值均小于草本群落, 分别为0.50、0.57和0.51(表 3), 表明洪积扇灌丛群落多样性和稳定性较草本群落较差。
| 物种名Species name | 科Family | 属Genus | 物种名Species name | 科Family | 属Genus | |
| 水葱Scirpus validus | 莎草科 | 藨草属 | 绢毛委陵菜Potentilla sericea | 蔷薇科 | 委陵菜属 | |
| 糠稷Panicum bisulcatum | 禾本科 | 黍属 | 草血竭Polygonum paleaceum | 蓼科 | 蓼属 | |
| 高原毛茛Ranunculus tanguticus | 毛茛科 | 毛茛属 | 白草Pennisetum centrasiaticum | 禾本科 | 狼尾草属 | |
| 火绒草Leontopodium leontopodioides | 菊科 | 火绒草属 | 佛甲草Sedum lineare | 景天科 | 景天属 | |
| 黄耆Astragalus membranaceus | 豆科 | 黄耆属 | 大针茅Stipa grandis | 禾本科 | 针茅属 | |
| 裂叶委陵菜Potentilla chinensis | 蔷薇科 | 委陵菜属 | 马兰Kalimeris indica | 菊科 | 马兰属 | |
| 橐吾Ligularia sibirica | 菊科 | 橐吾属 | 牛筋草Eleusine indica | 禾本科 | 穇属 | |
| 喜马拉雅米口袋Tibetia himalaica | 豆科 | 高山豆属 | 油菜Brassica napus | 十字花科 | 芸薹属 | |
| 肉果草Lancea tibetica | 玄参科 | 肉果草属 | 青稞Hordeum vulgare var. nudum | 禾本科 | 大麦属 | |
| 龙胆Gentiana scabra | 龙胆科 | 龙胆属 | 水栒子Cotoneaster multiflorus. | 蔷薇科 | 栒子属 | |
| 鼠麴草Gnaphalium affine | 菊科 | 鼠曲草属 | 高山柏Sabina squamata | 柏科 | 圆柏属 | |
| 中华草沙蚕Tripogon chinensis | 禾本科 | 草沙蚕属 | 金露梅Potentilla fruticosa | 蔷薇科 | 委陵菜属 | |
| 小蓝雪花Ceratostigma minus | 白花丹科 | 蓝雪花属 | 扁刺峨眉蔷薇Rosa omeiensis f. pteracantha | 蔷薇科 | 蔷薇属 | |
| 画眉草Eragrostis pilosa | 禾本科 | 画眉草属 | 小檗Berberis thunbergii | 小檗科 | 小檗属 | |
| 朝天委陵菜Potentilla supina | 蔷薇科 | 委陵菜属 |
| 样地 Plots |
主要物种 Main species |
物种数 Species numbers |
草本盖度 Grass coverage/% |
灌丛盖度 Shrub coverage/% |
H | Ma | E |
| 草地1 Grassland 1 | Sci+Pan+Pot+Leo | 26 | 73 | — | 1.90±0.12a | 2.49±0.41a | 0.70±0.04a |
| 草地2 Grassland 2 | Tri+Gna+Lan+Cer | 13 | 40 | — | 1.54±0.49ab | 1.64±0.33bc | 0.64±0.19a |
| 草地3 Grassland 3 | Poten+Sti+Erag+Sci | 19 | 75 | — | 1.86±0.19a | 1.91±0.14b | 0.70±0.07a |
| 草地4 Grassland 4 | Sci+Trig+Pote+Tha | 19 | 88 | — | 0.98±0.31cd | 1.65±0.39bc | 0.38±0.10b |
| 草地6 Grassland 6 | Pol+Ran+Pote+Sci | 19 | 98 | — | 1.75±0.31ab | 1.85±0.52b | 0.67±0.06a |
| 草地7 Grassland 7 | Tha+Ligu+Ast+Gen | 11 | 58 | — | 0.50±0.16e | 0.98±0.06d | 0.23±0.07b |
| 草地8 Grassland 8 | Gna+Sci+Ast+Sed | 15 | 53 | — | 1.36±0.05bc | 1.51±0.19bcd | 0.57±0.04a |
| 草地9 Grassland 9 | Gna+Sci+Ast+Kal | 14 | 45 | — | 0.83±0.13de | 1.20±0.36cd | 0.38±0.05b |
| 草地10 Grassland10 | Ast+Sed+Him+Pan | 20 | 70 | — | 1.40±0.15bc | 1.77±0.15bc | 0.57±0.10a |
| 灌丛样地3 Shrubland 3 | Ber+Rosa+Cot-Tha | 47 | 67 | 55 | 0.72±0.21a | 0.88±0.35a | 0.75±0.15a |
| 灌丛样地4 Shrubland 4 | Ber+Pott+Rosa+Sab | 29 | 65 | 55 | 0.78±0.38a | 0.86±0.50a | 0.66±0.16a |
| 灌丛样地5 Shrubland 5 | Rosa+Ber+Cha-Pen | 36 | 83 | 49 | 0.88±0.17a | 0.97±0.36a | 0.93±0.68a |
| 灌丛样地7 Shrubland 7 | Rosa+Ber-Tha+Gen | 21 | 11 | 33 | 0.14±0.24b | 0.17±0.29b | 0.20±0.34b |
| 灌丛样地9 Shrubland 9 | Pott-Gen+Sci+Tha | 18 | 47 | 42 | 0b | 0b | 0b |
| 农田1 Cropland 1 | Hor+Bra | — | 87 | — | — | — | — |
| 农田2 Cropland 2 | Hor | — | 65 | — | — | — | — |
| 农田3 Cropland 3 | Hor | — | 82 | — | — | — | — |
| 农田5 Cropland 5 | Bra | — | 67 | — | — | — | — |
| 农田7 Cropland 7 | Bra | — | 87 | — | — | — | — |
| 农田8 Cropland 8 | Hor | — | 53 | — | — | — | — |
| Sci:水葱Scirpus validus;Pot:裂叶委陵菜Potentilla chinensis;Ligu:橐吾Ligularia sibirica; Leo:火绒草Leontopodium leontopodioides;Tri:中华草沙蚕Tripogon chinensis;Gna:鼠麴草 Gnaphalium affine;Lan:肉果草 Lancea tibetica;Cer:小蓝雪花Ceratostigma minus;Gen:龙胆 Gentiana scabra;Tha:牛筋草Eleusine indica;Poten:绢毛委陵菜Potentilla sericea;Sti:大针茅Stipa grandis; Erag:画眉草 Eragrostis pilosa;Pote:朝天委陵菜 Potentilla supina;Pol:草血竭Polygonum paleaceum;Ran:高原毛茛Ranunculus tanguticus;Ast:黄耆Astragalus membranaceus;Kal:马兰Kalimeris indica;Sed:佛甲草Sedum lineare;Pen:白草Pennisetum centrasiaticum;Him:喜马拉雅米口袋Tibetia himalaica;Pan:糠稷Panicum bisulcatum;Ber:小檗Berberis thunbergii;Rosa:扁刺峨眉蔷薇Rosa omeiensis;Cot:水栒子Cotoneaster multiflorus;Pott:金露梅Potentilla fruticosa;Sab:高山柏Sabina squamata;Hor:青稞Hordeum vulgare;Bra:油菜Brassica napus;H:Shannon-Wiener多样性指数Shannon-Wiener diversity index;Ma:Margalef丰富度指数Margalef richness index;E:Pielou均匀度指数Pielou evenness index;不同的字母代表该植被类型内各植物多样性指数在不同样方之间差异显著(P < 0.05) | |||||||
由图 2和表 4可得, 研究样地SOC、TN、TP和TK的平均含量分别为34.38 g/kg、2.77 g/kg、0.39 g/kg和19.68 g/kg。其中, SOC、TN和TP含量在不同植被类型下均表现为草地>灌丛>农田, 土壤TK表现为农田>草地>灌丛。土壤TP和TK在不同植被类型下的含量差异不显著(P>0.05), SOC和TN含量在草地和农田下差异显著(P < 0.05)。土壤AN、AP和AK的平均含量分别为130.78 mg/kg、3.36 mg/kg和189.79 mg/kg。其中, 土壤AN和AK的含量在草地和农田下差异显著(P < 0.05), 土壤AP在不同植被类型下差异不显著(P>0.05)。SOC、TN、AN和TP含量的COV分别为59%、58%、57%和60%, 属于中等程度的变异, 土壤AP含量的COV分别为171%, 属于强变异, 表明其空间变异性较大;土壤TK含量的COV为17%, 相对较小, 而AK含量的COV为114%, 属于强变异, 表明土壤K元素在研究样地总体分布均匀但其有效性差异大。

|
| 图 2 不同植被类型下的土壤养分含量 Fig. 2 Soil nutrient content under different vegetation types, different letters indicated significant differences in soil nutrients 不同的字母表示土壤养分含量在不同植被类型下存在显著差异(P < 0.05) |
| 土壤养分及计量比 Soil nutrient and measurement ratio |
变化范围 Variation range |
均值 Mean values |
标准差 Standard deviation |
变异系数 Coefficient of variation/% |
| SOC/(g/kg) | 10.28—109.75 | 34.38 | 20.14 | 59 |
| TN/(g/kg) | 0.78—8.97 | 2.77 | 1.60 | 58 |
| TP/(g/kg) | 0.04—1.27 | 0.39 | 0.23 | 60 |
| TK/(g/kg) | 7.71—25.57 | 19.68 | 3.38 | 17 |
| AN/(mg/kg) | 31.27—358.56 | 130.78 | 74.65 | 57 |
| AP/(mg/kg) | 0.12—15.31 | 3.36 | 3.48 | 171 |
| AK/(mg/kg) | 41.00—629.00 | 189.79 | 140.52 | 114 |
| C∶N | 4.55—19.17 | 12.75 | 2.21 | 17 |
| N∶P | 1.01—31.64 | 8.10 | 4.54 | 56 |
| C∶P | 16.02—465.76 | 102.50 | 64.20 | 63 |
| N∶K | 0.03—0.62 | 0.16 | 0.12 | 78 |
由表 4和图 3可得, 研究样地土壤C∶N、C∶P、N∶P、和N∶K值分别为12.75、8.10、102.50和0.16, 土壤C∶N的均值表现为农田>灌丛>草地, 土壤C∶P、N∶P和N∶K值均表现为草地>灌丛>农田, 其中只有土壤N∶P在农田下的值与在草地和灌丛下的值差异显著(P < 0.05)。土壤N∶P、C∶P和N∶K的COV分别为56%、63%和78%, 均属于中等程度的变异;土壤C∶N的COV为17%, 相对较小, 表明C∶N在研究样地变化不大。由图 4可得, 土壤C∶N(12.71)受到良好的约束, 在不同植被类型下变化不大, 而土壤C∶P、N∶P和N∶K相对来说不稳定, 变化较大。

|
| 图 3 不同植被类型下的土壤生态化学计量比 Fig. 3 Soil ecological stoichiometry under different vegetation types, different letters indicated that there were significant differences in soil ecological stoichiometry 不同的字母表示土壤生态化学计量比在不同植被类型下存在显著差异(P < 0.05) |

|
| 图 4 土壤生态化学计量特征 Fig. 4 Soil ecological stoichiometry characteristics |
土壤养分及其计量比之间的Pearson相关分析显示(图 5), SOC与TN、TP、AN、AK、N∶K之间均存在极显著的正相关关系(P < 0.01), 与TK、K: P之间均存在极显著的负相关关系(P < 0.01), 与AP、N∶P显著正相关(P < 0.05), 与土壤C∶P显著负相关(P < 0.05);土壤TN与TP、AN、AP、AK、N∶K之间均存在极显著的正相关关系(P < 0.01), 与TK之间存在极显著的负相关关系(P < 0.01), 与N∶P显著正相关(P < 0.05);土壤TP与AN、N∶K之间均存在极显著的正相关关系(P < 0.01), 与TK、N∶P、K: P、C∶P之间存在极显著的负相关关系(P < 0.01), 与AP、AK显著正相关(P < 0.05);土壤TK与AK显著负相关(P < 0.05);土壤AN与AK、N∶K之间均存在极显著的正相关关系(P < 0.01), 与AP、N∶P、C∶P显著正相关(P < 0.05);土壤AP与N∶K之间存在极显著的正相关关系(P < 0.01);土壤AK与N∶K之间存在极显著的正相关关系(P < 0.01);土壤N∶P与土壤K: P、C∶P之间均存在极显著的正相关关系(P < 0.01), 与N∶K显著正相关(P < 0.05)。

|
| 图 5 土壤养分与其化学计量比之间的相关关系 Fig. 5 Correlations between soil nutrients and their stoichiometric ratios **P < 0.01;*P < 0.05 |
植物群落特征指数与土壤生态化学计量比之间的Pearson相关分析显示, 仅有物种盖度与SOC和TN极显著正相关(P < 0.01, 表 5);而Shannon-Wiener多样性指数、Margalef丰富度指数、Pielou均匀度指数和物种数均与各土壤养分及其计量比不显著相关(P>0.05, 表 5)。CCA进一步显示, 土壤养分及其化学计量比对洪积扇植物群落物种组成影响显著(P=0.002, 图 6), 但仅解释了物种组成变化总变异的26.9%, 其中, 仅有TP、K∶P和AP通过蒙特卡洛检验(检验次数为499次), 对植物群落物种组成变化影响显著(P < 0.05), 分别解释了物种组成总变异的6.2%、5.5%和4.1%, 表明土壤TP、K∶P和AP是影响洪积扇植物群落物种组成变化的主要土壤生态化学计量因子。
| 群落特征值 Community characteristic index |
SOC | TN | TP | TK | C∶N | N∶P | C∶P | N∶K |
| 物种数Species number | -0.011 | -0.002 | 0.153 | -0.061 | -0.033 | 0.030 | 0.021 | 0.077 |
| 盖度Coverage | 0.412** | 0.424** | 0.288 | -0.112 | 0.058 | 0.081 | 0.095 | 0.289 |
| H | -0.207 | -0.232 | -0.088 | 0.163 | 0.168 | -0.094 | -0.055 | -0.129 |
| Ma | -0.112 | -0.107 | 0.079 | -0.005 | -0.034 | 0.004 | -0.006 | 0.001 |
| E | -0.217 | -0.224 | -0.051 | -0.069 | 0.010 | -0.076 | -0.062 | -0.061 |
| H:Shannon-Wiener多样性指数Shannon-Wiener diversity index;Ma:Margalef丰富度指数Margalef richness index;E:Pielou均匀度指数Pielou evenness index;**表示在0.01级别相关性显著 | ||||||||

|
| 图 6 群落主要物种与土壤养分及其计量比之间的典范对应分析 Fig. 6 Canonical Correspondence Analysis between community major species and soil nutrient and their measurement ratios 红色箭头代表解释变量, 红色星星代表不同的物种;Sci:水葱Scirpus validus;Pot:裂叶委陵菜Potentilla chinensis;Ligu:橐吾Ligularia sibirica; Leo:火绒草Leontopodium leontopodioides;Tri:中华草沙蚕Tripogon chinensis;Gna:鼠麴草 Gnaphalium affine;Lan:肉果草 Lancea tibetica;Cer:小蓝雪花Ceratostigma minus;Gen:龙胆 Gentiana scabra;Tha:牛筋草Eleusine indica;Poten:绢毛委陵菜Potentilla sericea;Sti:大针茅Stipa grandis; Era:画眉草 Eragrostis pilosa;Pote:朝天委陵菜 Potentilla supina;Pol:草血竭Polygonum paleaceum;Ran:高原毛茛Ranunculus tanguticus;Ast:黄耆Astragalus membranaceus;Kal:马兰Kalimeris indica;Sed:佛甲草Sedum lineare;Pen:白草Pennisetum centrasiaticum;Him:喜马拉雅米口袋Tibetia himalaica;Pan:糠稷Panicum bisulcatum;Ber:小檗Berberis thunbergii;Rosa:扁刺峨眉蔷薇Rosa omeiensis;Cot:水栒子Cotoneaster multiflorus;Pott:金露梅Potentilla fruticosa;Sab:高山柏Sabina squamata |
本研究中, 洪积扇SOC和TN平均含量均高于我国土壤SOC(29.51 g/kg)和TN(2.30 g/kg)平均含量, 可能有以下3方面的原因。其一是洪积扇普遍存在放牧现象, 牲畜排泄粪便到土壤中直接增加SOC和TN的含量, 这也是造成SOC和TN空间变异性大的主要原因;其二是洪积扇植被覆盖度高, 尤其是草地, 平均盖度达65%, 枯落物层较厚, 因而土壤中有机质的含量高;其三是由于近年来农业化肥的广泛使用和化石燃料燃烧的增加导致全球大气N沉降增加[24], 全球N沉降的增加会提升土壤中C、N元素的含量, 此外, 这种变化在不同的陆地生态系统(农田、草地和林地生态系统)和气候背景下均存在[25], 而青藏高原地区独特的地理位置以及较高的海拔, 使得其对大气N沉降的响应更为敏感[26]。本研究中SOC和TN在不同植被类型下的含量均表现为草地>灌丛>农田, 且在草地和农田下差异显著(P < 0.05), 一方面是因为草地和农田下枯落物量差异大, 另一方面是由于洪积扇的地理位置、形成时间等引起的洪积扇之间的差异导致的(表 6)。土壤TP主要来源于土壤母质及岩石的风化, 因此其在土壤中的含量相对稳定。洪积扇土壤TP平均含量为0.39 g/kg, 略低于全国土壤TP含量0.52—0.78 g/kg, 显著低于世界土壤TP平均含量2.8 g/kg, 处于极度缺P状态。这主要与土壤母质的类型有关, 西藏土壤成土母质组成复杂, 但总体上含P量不高[27]。本研究洪积扇土壤TK的平均含量为19.68 g/kg, 根据全国第二次土壤养分普查标准[28], 洪积扇土壤K元素的含量水平处于第三等级, 属于相对富K的土壤。西藏土壤富K的原因主要有以下两方面[29]:其一是因为土壤中K元素主要来源于土壤母质, 而西藏地区土壤母质中K元素较丰富;其二是因为西藏地区土壤K元素风化淋溶程度普遍很低, 因此土壤TK含量较高。土壤养分之间的相关分析表明, 土壤TK与SOC、TN、TP之间均呈极显著的负相关(P < 0.01), 这可能是因为土壤矿物质中K的含量高于有机质中K的含量。李建平等[30]发现富有机质土壤中有机质与矿物态K之间存在“稀释效应”, 即土壤有机质的增加会导致土壤全K的相对减少, 但这种稀释与K元素的风化淋溶不同, 并没有实质性的损失。
| 因素Factors | 显著性Significance | ||
| 植被类型 Vegetation types |
洪积扇 Alluvial fans |
植被类型×洪积扇 Vegetation types×Alluvial fans |
|
| SOC | 0.046* | 0.000** | 0.138 |
| TN | 0.026* | 0.000** | 0.310 |
| TP | 0.996 | 0.000** | 0.686 |
| TK | 0.325 | 0.003** | 0.551 |
| AN | 0.038* | 0.000** | 0.090 |
| AP | 0.095 | 0.071 | 0.504 |
| AK | 0.031* | 0.000** | 0.596 |
| C∶N | 0.093 | 0.009** | 0.020 |
| N∶P | 0.025* | 0.361 | 0.890 |
| C∶P | 0.05 | 0.314 | 0.649 |
| N∶K | 0.126 | 0.004** | 0.655 |
| *表示在0.05级别影响显著;**表示在0.01级别影响显著 | |||
土壤AN、AP、AK等速效养分是表征土壤直接的肥力供应能力的良好指标, 研究样地土壤AN、AP和AK的平均含量分别为130.78 mg/kg、3.36 mg/kg、189.79 mg/kg。根据全国第二次土壤普查养分分级标准[28, 31](表 7), 研究样地土壤AN和AK含量均处于稍丰水平, AP的含量处于缺水平, 这与土壤TP含量低有关, 此外有研究显示青藏高原不同草地类型与土壤P循环相关的碱性磷酸酶活性低[32], 进而使得土壤AP含量极低。
| 丰缺度 Abundance degree |
养分分级 Nutrient classification |
AN/ (mg/kg) | AP/ (mg/kg) | AK/ (mg/kg) |
| 丰abundant | 1 | >150 | >40 | >200 |
| 稍丰slightly abundant | 2 | 120—150 | 20—40 | 150—200 |
| 中等medium | 3 | 90—120 | 10—20 | 100—150 |
| 稍缺slightly deficient | 4 | 60—90 | 5—10 | 50—100 |
| 缺deficient | 5 | 30—60 | 3—5 | 30—50 |
| 极缺extremely deficient | 6 | < 30 | < 3 | < 30 |
本研究中, 土壤C∶N值在草地、灌丛和农田的均值分别为12.47、12.79和13.40, 略高于我国土壤C∶N的平均值11.38[33], 均小于理论上较适宜的C∶N值25∶1, 表明洪积扇SOC的矿化速率较高。土壤C∶N值在各植被类型下表现为农田>灌丛>草地, 这与Liu和Wang[34]在黄土高原高寒地区的研究结果一致。土壤C∶N值主要受所输入的植被枯落物的质与量的影响, 向土壤中输入的枯落物越多, 微生物分解有机质所需的基质就越充足, 从而有机质的分解速率就越快[35]。农田向土壤中输入的枯落物最少, 其下微生物分解有机质所需的基质最少, 因此有机质的分解速率就最低, 从而其土壤C∶N值最高。灌丛凋落物具有较多的生化难降解物质(尤其是脂肪族生物聚合物), 与草地凋落物相比不适合作为微生物基质, 因此其分解速率比草地慢, C∶N值高于草地。SOC和TN含量在草地和农田下差异显著(P < 0.05)(图 2), 研究区土壤C∶N(12.71)在不同植被类型下均表现出良好的约束(图 4), 这与Cleveland和Liptzin[36]、曾全超等[37]的研究结果一致。而SOC与TN含量之间极显著正相关(P < 0.01), 说明不同植被类型对SOC和TN的响应具有一致性。土壤C∶P值在各植被类型下表现为草地>灌丛>农田, 其均值分别为122.70、107.90和69.36, 均小于200。一般来说, C∶P < 200意味着土壤有机质的净矿化[9]。这与C∶N值结果相一致, 均说明了本研究洪积扇土壤有机质的矿化速率较高, 从而导致了土壤肥力的下降。
目前关于土壤化学计量比的研究多集中于C∶N∶P[38—39], 本研究中土壤N∶K值与SOC、TN、TP、AN、AP、AK含量均极显著相关(P < 0.01), 与土壤N∶P显著正相关(P < 0.05), 表明N∶K值也可以作为评价洪积扇土壤养分状况的指标。土壤N∶P的值与气候、土壤和植被等因素有关, 本研究中土壤N∶P在草地的含量最高, 可能与草本样地中有较多的黄耆、喜马拉雅米口袋等豆科植物有关。土壤TN、TP与N∶P的Pearson相关分析表明, 土壤TN与N∶P呈显著的正相关(P < 0.05), 土壤TP与N∶P呈极显著的负相关(P < 0.01), 说明土壤TP对N∶P的影响大于TN。Du和Gao[40]在青藏高原东部退化高寒草甸中的研究显示, 围栏封育9年后植被在生长发育过程中受到土壤P元素的限制, 本研究洪积扇土壤N∶P(8.10)与其研究区土壤N∶P(6.60)接近, 且土壤TP(0.39 g/kg)远低于其研究区的TP含量(1.18 g/kg), 说明洪积扇植被在生长发育过程中更加受到土壤P元素的限制。事实上, 在不断变化的气候条件和风化阶段, 由于对N沉降的敏感响应, 西藏地区也越来越频繁观察到P限制[41—45]。也有研究指出, 植被在生长发育过程中最终受到土壤P元素的限制, 因为当土壤中的磷酸盐耗尽后, 土壤P元素便不会再得到补充[46]。
3.3 土壤生态化学计量因子对洪积扇植物群落物种组成变化的影响植物群落特征指数与土壤养分及其计量比之间的Pearson相关分析显示, Shannon-Wiener多样性指数和Margalef丰富度指数与各土壤养分及其计量比之间均不存在显著的相关关系(P>0.05), 这和一些学者的研究不一致[47—48], 可能与气候和放牧两方面的因素有关。首先, 拉萨河流域雨季集中在5—9月, 冬天干旱, 因此洪积扇植物种多为短命植物, 植物多样性指数低, 植物群落结构不稳定, 从而导致植物群落特征指数与土壤养分及其化学计量比之间的相关性弱;其次, 洪积扇普遍存在放牧现象, 放牧在一定程度上会使植物群落的多样性降低[49], 从而加剧了两者之间的不相关程度。本研究中Pielou均匀度指数与土壤养分及其计量比之间也不显著相关, 这与赵景学[50]、高国刚[51]、文海燕[52]和王顺忠等[53]的研究结果相一致。Pielou均匀度指数反映了群落内各物种的相对多度, 其值的大小与物种的适生性有关, 而不同物种因生活习性的差异对各养分的需求度不同, 因此均匀度的值可能与某种特定的养分因子相关。洪积扇植物群落盖度与SOC和TN极显著正相关(P < 0.01), 说明在拉萨河流域洪积扇这样的植物群落组成不稳定的地区, 群落盖度可能比上述3种常用的植物多样性指数更适合表征植被特征与土壤养分之间的关系。CCA进一步显示, 土壤养分及其计量比对洪积扇植物群落物种组成变化的影响极其显著(P=0.002), 但其解释率仅有29.6%。Liu等[54]对黄土高原次生演替过程中植被群落与土壤、植物和微生物生态化学计量比的研究显示, 土壤、植物和微生物C∶N∶P共解释了群落物种组成变化总变异的80.8%, 说明除了土壤生态化学计量因子之外, 土壤微生物生物量和植物本身的C∶N∶P也是解释植物群落物种组成变化的重要因子。所以本研究中CCA的解释率低的原因可能是因为还有许多重要的解释变量如水分、温度、海拔和土壤微生物活性等未考虑, 由洪积扇和植被类型对土壤养分的影响的双因素方差分析(表 5)也表明, 洪积扇自身的差异也是造成土壤养分差异的重要因素, 在后续的研究中应当予以重视。CCA结果还显示, 土壤TP、K∶P和AP是洪积扇植物群落物种组成的主要土壤生态化学计量因子(P < 0.05), 在一定程度上印证了本研究洪积扇植被在生长发育过程中受到土壤P元素限制的观点。
4 结论本研究通过对拉萨河流域洪积扇的植被调查, 并采集其下土壤, 研究拉萨河流域不同植被类型下的土壤化学计量特征, 具体结果如下:
(1) 拉萨河流域洪积扇植物种相对丰富, 主要为草本物种, 其中以禾本科和菊科物种居多;灌丛较少, 灌丛群落的多样性、丰富度、均匀度均较草本群落差;基本无天然乔木分布。;
(2) 洪积扇水分条件较好, C、N、K养分含量相对充足, TP和AP含量较稀缺, 植被在生长发育过程中受到土壤P元素的限制。虽然洪积扇SOC和TN含量相对丰富, 但土壤C∶N值低, 土壤有机质矿化速率高, 从而导致洪积扇的土壤肥力低。此外, 土壤C∶N值在不同植被类型下受到良好的约束, 差异不大。土壤N∶K与SOC、TN、TP、AN、AP、AK均极显著相关(P < 0.01), 说明N∶K同C∶N∶P一样, 也可作为评价土壤养分状况的生态化学计量指标。
(3) CCA结果显示, 土壤生态化学计量因子对植物群落组成影响显著(P=0.002), 其中土壤TP、K∶P和AP是洪积扇植物群落物种组成的主要土壤生态化学计量因子。但其总体解释率仅有26.9%, 在未来的研究中还应充分考虑土壤水分、温度以及微生物活性等因子的影响。
(4) 本研究通过调查拉萨河流域洪积扇不同植被类型下的土壤养分含量, 分析其生态化学计量特征, 阐明拉萨河流域洪积扇整体的土壤养分稀缺状况。然而, 植被只是影响洪积扇土壤养分状况的一个方面, 洪积扇的成土过程, 形成时间及其所处气候条件等均对其影响显著, 本研究在这方面的讨论涉及较少, 在后续更加深入、具体的研究中应当加强对洪积扇自身条件差异的研究。
| [1] |
Ågren G I. Stoichiometry and nutrition of plant growth in natural communities. Annual Review of Ecology, Evolution, and Systematics, 2008, 39: 153-170. DOI:10.1146/annurev.ecolsys.39.110707.173515 |
| [2] |
Wang Y G, Li Y, Ye X H, Chu Y, Wang X P. Profile storage of organic/inorganic carbon in soil: from forest to desert. Science of the Total Environment, 2010, 408(8): 1925-1931. DOI:10.1016/j.scitotenv.2010.01.015 |
| [3] |
金强, 安婉丽, 刘旭阳, 陈晓旋, 林少颖, 王维奇. 模拟酸雨对福州沿江稻田水稻叶片碳氮磷含量及其生态化学计量学特征的影响. 生态学报, 2020, 40(9): 3085-3095. |
| [4] |
Venterink H O, van der Vliet R, Wassen M J. Nutrient limitation along a productivity gradient in wet meadows. Plant and Soil, 2001, 234(2): 171-179. DOI:10.1023/A:1017922715903 |
| [5] |
Elser J J, Dobberfuhl D R, MacKay N A, Schampel J H. Organism size, life history, and N∶P stoichiometry: toward a unified view of cellular and ecosystem processes. BioScience, 1996, 46(9): 674-684. DOI:10.2307/1312897 |
| [6] |
Hessen D O. Stoichiometry in food webs-Lotka revisited. Oikos, 1997, 79(1): 195-200. DOI:10.2307/3546108 |
| [7] |
曾德慧, 陈广生. 生态化学计量学: 复杂生命系统奥秘的探索. 植物生态学报, 2005, 29(6): 1007-1019. DOI:10.3321/j.issn:1005-264X.2005.06.018 |
| [8] |
Fan H B, Wu J P, Liu W F, Yuan Y H, Hu L, Cai Q K. Linkages of plant and soil C∶N∶P stoichiometry and their relationships to forest growth in subtropical plantations. Plant and Soil, 2015, 392(1/2): 127-138. |
| [9] |
Paul E A. Soil Microbiology, Ecology and Biochemistry. 4th ed. Waltham: Elsevier, 2014.
|
| [10] |
Koerselman W, Meuleman A F M. The vegetation N∶P ratio: a new tool to detect the nature of nutrient limitation. Journal of Applied Ecology, 1996, 33(6): 1441-1450. DOI:10.2307/2404783 |
| [11] |
Wassen M J, Venterink H G M O, de Swart E O A M. Nutrient concentrations in mire vegetation as a measure of nutrient limitation in mire ecosystems. Journal of Vegetation Science, 1995, 6(1): 5-16. DOI:10.2307/3236250 |
| [12] |
Bedford B L, Walbridge M R, Aldous A. Patterns in nutrient availability and plant diversity of temperate North American wetlands. Ecology, 1999, 80(7): 2151-2169. DOI:10.1890/0012-9658(1999)080[2151:PINAAP]2.0.CO;2 |
| [13] |
Yan Y, Lu X Y. Are N, P, and N∶P stoichiometry limiting grazing exclusion effects on vegetation biomass and biodiversity in alpine grassland?. Global Ecology and Conservation, 2020, 24: e01315. DOI:10.1016/j.gecco.2020.e01315 |
| [14] |
Shen F F, Wu J P, Fan H B, Liu W F, Guo X M, Duan H L, Hu L, Lei X M, Wei X H. Soil N/P and C/P ratio regulate the responses of soil microbial community composition and enzyme activities in a long-term nitrogen loaded Chinese fir forest. Plant Soil, 2019, 436(1): 91-107. |
| [15] |
Roem W J, Berendse F. Soil acidity and nutrient supply ratio as possible factors determining changes in plant species diversity in grassland and heathland communities. Biological Conservation, 2000, 92(2): 151-161. DOI:10.1016/S0006-3207(99)00049-X |
| [16] |
包小婷, 丁陆彬, 姚帅臣, 王景升, 石培礼, 王彤, 李超, 刘文婧. 拉萨河流域植物群落的数量分类与排序. 生态学报, 2019, 39(3): 779-786. |
| [17] |
陈同德, 焦菊英, 林红, 王颢霖, 赵春敬, 章志鑫. 扇形地的类型辨析及区分方法探讨. 水土保持通报, 2020, 40(4): 190-198. |
| [18] |
詹天宇, 侯阁, 刘苗, 孙建, 付顺. 青藏高原不同退化梯度高寒草地植被与土壤属性分异特征. 草业科学, 2019, 36(4): 1010-1021. |
| [19] |
罗亚勇, 孟庆涛, 张静辉, 赵学勇, 秦彧. 青藏高原东缘高寒草甸退化过程中植物群落物种多样性、生产力与土壤特性的关系. 冰川冻土, 2014, 36(5): 1298-1305. DOI:10.7522/j.issn.1000-0240.2014.0155 |
| [20] |
刘美, 马志良. 青藏高原东部乔灌交错带地被物和土壤碳氮磷化学计量特征. 应用与环境生物学报, [2021-05-10]. https://doi.org/10.19675/j.cnki.1006-687x.2020.05040.
|
| [21] |
林红, 焦菊英, 陈同德, 赵春敬. 西藏拉萨河流域中下游洪积扇植被的物种组成与多样性特征. 水土保持研究, 2021, 28(5): 67-75. |
| [22] |
张金屯. 数量生态学(第三版). 北京: 科学出版社, 2018.
|
| [23] |
卢同平, 张文翔, 牛洁, 林永静, 武梦娟. 典型自然带土壤氮磷化学计量空间分异特征及其驱动因素研究. 土壤学报, 2017, 54(3): 682-692. |
| [24] |
Galloway J N, Townsend A R, Erisman J W, Bekunda M, Cai Z C, Freney J R, Martinelli L A, Seitzinger S P, Sutton M A. Transformation of the nitrogen cycle: recent trends, questions, and potential solutions. Science, 2008, 320(5878): 889-892. DOI:10.1126/science.1136674 |
| [25] |
Sun Y, Wang C T, Chen H Y H, Ruan H H. Responses of C∶N stoichiometry in plants, soil, and microorganisms to nitrogen addition. Plant and Soil, 2020, 456(1/2): 277-287. |
| [26] |
张江, 袁旻舒, 张婧, 李函微, 王洁仪, 张贤, 鞠佩君, 蒋海波, 陈槐, 朱求安. 近30年来青藏高原高寒草地NDVI动态变化对自然及人为因子的响应. 生态学报, 2020, 40(18): 6269-6281. |
| [27] |
安新固, 关树森. 浅谈西藏农区土壤缺磷问题. 西藏农业科技, 1981(1): 28-36. |
| [28] |
全国土壤普查办公室. 中国土壤. 北京: 中国农业出版社, 1998.
|
| [29] |
高丽丽. 西藏土壤有机质和氮磷钾状况及其影响因素分析[D]. 雅安: 四川农业大学, 2004.
|
| [30] |
李建平, 姚祖芳, 刘世全. 西藏自治区土壤资源. 北京: 科学出版社, 1994.
|
| [31] |
罗红, 吴建普, 边巴多吉, 嘎玛群宗, 朱雪林. 西藏人工造林作业区土壤养分特征研究. 土壤学报, 2017, 54(2): 421-433. |
| [32] |
汪子微, 万松泽, 蒋洪毛, 胡扬, 马书琴, 陈有超, 鲁旭阳. 青藏高原不同高寒草地类型土壤酶活性及其影响因子. 植物生态学报, 2021, 45(5): 528-538. |
| [33] |
李启权, 岳天祥, 范泽孟, 杜正平, 陈传法, 卢毅敏. 中国表层土壤全氮的空间模拟分析. 地理研究, 2010, 29(11): 1981-1992. |
| [34] |
Liu R S, Wang D M. Soil C, N, P and K stoichiometry affected by vegetation restoration patterns in the alpine region of the Loess Plateau, Northwest China. PLoS One, 2020, 15(11): e0241859. DOI:10.1371/journal.pone.0241859 |
| [35] |
Pang D B, Wang G Z, Li G J, Sun Y L, Liu Y G, Zhou J X. Ecological stoichiometric characteristics of two typical plantations in the karst ecosystem of southwestern China. Forests, 2018, 9(2): 56. DOI:10.3390/f9020056 |
| [36] |
Cleveland C C, Liptzin D. C∶N∶P stoichiometry in soil: is there a "Redfield ratio" for the microbial biomass?. Biogeochemistry, 2007, 85(3): 235-252. DOI:10.1007/s10533-007-9132-0 |
| [37] |
曾全超, 李鑫, 董扬红, 安韶山. 黄土高原延河流域不同植被类型下土壤生态化学计量学特征. 自然资源学报, 2016, 31(11): 1881-1891. DOI:10.11849/zrzyxb.20160038 |
| [38] |
Hui D F, Yang X T, Deng Q, Liu Q, Wang X, Yang H, Ren H. Soil C∶N∶P stoichiometry in tropical forests on Hainan Island of China: Spatial and vertical variations. Catena, 2021, 201: 105228. DOI:10.1016/j.catena.2021.105228 |
| [39] |
Wang Z C, He G X, Hou Z H, Luo Z, Chen S X, Lu J, Zhao J. Soil C∶N∶P stoichiometry of typical coniferous (Cunninghamia lanceolata) and/or evergreen broadleaved (Phoebe bournei) plantations in South China. Forest Ecology and Management, 2021, 486: 118974. DOI:10.1016/j.foreco.2021.118974 |
| [40] |
Du C J, Gao Y H. Grazing exclusion alters ecological stoichiometry of plant and soil in degraded alpine grassland. Agriculture, Ecosystems & Environment, 2021, 308: 107256. |
| [41] |
Zhao G S, Shi P L, Wu J S, Xiong D P, Zong N, Zhang X Z. Foliar nutrient resorption patterns of four functional plants along a precipitation gradient on the Tibetan Changtang Plateau. Ecology and Evolution, 2017, 7(18): 7201-7212. DOI:10.1002/ece3.3283 |
| [42] |
Zhang J J, Yan X B, Su F L, Li Z, Wang Y, Wei Y N, Ji Y G, Yang Y, Zhou X H, Guo H, Hu S J. Long-term N and P additions alter the scaling of plant nitrogen to phosphorus in a Tibetan alpine meadow. Science of the Total Environment, 2018, 625: 440-448. DOI:10.1016/j.scitotenv.2017.12.292 |
| [43] |
Hong J T, Wang X D, Wu J B. Stoichiometry of root and leaf nitrogen and phosphorus in a dry alpine steppe on the northern Tibetan Plateau. PLoS One, 2014, 9(10): e109052. DOI:10.1371/journal.pone.0109052 |
| [44] |
Zhang H R, Sun W, Li S W, Han F S, Yu C Q, Zhang X Z, Wang J S, He Y T, Zhong Z M. Impact of extra nitrogen on ecological stoichiometry of alpine grasslands on Tibetan Plateau: meta-analysis. Polish Journal of Ecology, 2019, 66(4): 315-324. DOI:10.3161/15052249PJE2018.66.4.001 |
| [45] |
Tian L M, Zhao L, Wu X D, Fang H B, Zhao Y H, Hu G J, Yue G Y, Sheng Y, Wu J C, Chen J, Wang Z W, Li W P, Zou D F, Ping C L, Shang W, Zhao Y G, Zhang G L. Soil moisture and texture primarily control the soil nutrient stoichiometry across the Tibetan grassland. Science of the Total Environment, 2018, 622-623: 192-202. DOI:10.1016/j.scitotenv.2017.11.331 |
| [46] |
Bui E N, Henderson B L. C∶N∶P stoichiometry in Australian soils with respect to vegetation and environmental factors. Plant and Soil, 2013, 373(1/2): 553-568. |
| [47] |
汪攀, 王霖娇, 盛茂银. 西南喀斯特石漠化生态系统植物多样性、土壤生态化学计量特征及其相关性分析. 南方农业学报, 2018, 49(10): 1959-1969. DOI:10.3969/j.issn.2095-1191.2018.10.09 |
| [48] |
白永飞, 李凌浩, 王其兵, 张丽霞, 张焱, 陈佐忠. 锡林河流域草原群落植物多样性和初级生产力沿水热梯度变化的样带研究. 植物生态学报, 2000, 24(6): 667-673. DOI:10.3321/j.issn:1005-264X.2000.06.005 |
| [49] |
王琳, 张金屯, 上官铁梁, 樊龙锁. 历山山地草甸的物种多样性及其与土壤理化性质的关系. 应用与环境生物学报, 2004, 10(1): 18-22. DOI:10.3321/j.issn:1006-687X.2004.01.004 |
| [50] |
赵景学, 曲广鹏, 多吉顿珠, 尚占环. 藏北高寒植被群落物种多样性与土壤环境因子的关系. 干旱区资源与环境, 2011, 25(6): 105-108. |
| [51] |
高国刚, 胡玉昆, 李凯辉, 肖洪亮, 公延明, 尹伟. 高寒草地群落物种多样性与土壤环境因子的关系. 水土保持通报, 2009, 29(3): 118-122. |
| [52] |
文海燕, 傅华, 赵哈林. 退化沙质草地植物群落物种多样性与土壤肥力的关系. 草业科学, 2008, 25(10): 6-9. DOI:10.3969/j.issn.1001-0629.2008.10.002 |
| [53] |
王顺忠, 陈桂琛, 柏玉平, 周国英, 孙菁. 青海湖鸟岛地区植物群落物种多样性与土壤环境因子的关系. 应用生态学报, 2005, 16(1): 186-188. DOI:10.3321/j.issn:1001-9332.2005.01.038 |
| [54] |
Liu W C, Fu S Y, Yan S J, Ren C J, Wu S J, Deng J, Li B Y, Han X H, Yang G H. Responses of plant community to the linkages in plant-soil C∶N∶P stoichiometry during secondary succession of abandoned farmlands, China. Journal of Arid Land, 2020, 12(2): 215-226. DOI:10.1007/s40333-020-0009-6 |