 2022, Vol. 42
2022, Vol. 42文章信息
- 张力天, 杨时海, 刘炜, 王晓丽, 秦金萍, 董瑞珍, 边巴普尺, 米玛顿珠, 索朗次仁, 张树淼, 张敏, 仁增曲扎, 马玉寿
- ZHANG Litian, YANG Shihai, LIU Wei, WANG Xiaoli, QIN Jinping, DONG Ruizhen, BIANBA Puchi, MIMA Dunzhu, SUOLANG Ciren, ZHANG Shumiao, ZHANG Min, RENZENG Quzha, MA Yushou
- 雅鲁藏布江中上游白草主要分布区植物群落特征
- Characteristics of plant communities in distribution areas of Pennisetum centrasiaticum in the upper and middle reaches of the Yarlung Zangbo River
- 生态学报. 2022, 42(15): 6111-6124
- Acta Ecologica Sinica. 2022, 42(15): 6111-6124
- http://dx.doi.org/10.5846/stxb202107091853
-
文章历史
- 收稿日期: 2021-07-09
- 网络出版日期: 2022-04-07





2. 日喀则市百亚成农牧产品加工有限公司, 日喀则 857800;
3. 西藏云旺实业有限公司, 日喀则 857000;
4. 兰州大学生命科学学院, 兰州 730000
2. Shigatse Baiyacheng Agricultural Products Processing Co., Ltd., Shigatse 857800, China;
3. Tibet Yunwang Industrial Co., Ltd., Shigatse 857000, China;
4. School of Life Sciences, Lanzhou University, Lanzhou 730000, China
种群结构反映了种群内不同年龄个体数量的组成情况、种群数量动态及其发展趋势, 并在很大程度上体现种群与环境之间的相互关系, 以及种群在群落中的作用和地位[1]。植物群落是植物种群与环境因子共同构成的一个有机整体, 不同物种种群在群落中所处空间分布格局取决于物种的生物学特性、生境条件及其交互作用[2—3]。探究植物群落多样性及其维持机制, 是了解植物群落生态功能和稳定的基础, 也一直是群落生态学研究的中心议题[3]。海拔梯度下, 水、热因子可在相对较小范围内急剧变化[4—5], 成为物种丰富度梯度格局研究的重要方面[6], 特别是在陆地生态系统中江河流域的河岸区是生物种群和各种环境因子相互影响的潜在敏感地区[1], 掌握河岸生物多样性分布格局与各种环境因子的关系, 对于河岸生态系统保护特别是流域生态安全具有十分重要的科学意义[7—10]。
白草(Pennisetum centrasiaticum)是禾本科(Gramineae)黍族(Trib. Paniceae)狼尾草属(Pennisetum)多年生草种, 具横走根茎, 秆直立, 单生或丛生, 产我国黑龙江、吉林、辽宁、内蒙古、河北、山西、陕西、甘肃、青海、四川(西北部)、云南(北部)、西藏等省区, 多生于海拔800—4600 m山坡和较干燥之处[11]。白草是一种根茎型禾本科牧草, 地下部可蔓延形成网状地下根茎系统[12]而具有强大的扩展能力[13]。目前, 对于白草的研究有核型分析[14—15]、构件生物量分配[16—17]、耐旱研究[13, 18—19]、营养价值[20—21]、栽培研究[22—23]以及生态修复[24—25]等。
雅鲁藏布江是青藏高原上的最重要的河流, 同时也是我国重要的国际河流之一[26], 其发源于西藏自治区日喀则市仲巴县海拔5590 m的杰马央宗冰川[27], 由西向东横贯西藏南部, 绕过喜马拉雅山脉最东端的南迦巴瓦峰转向南流, 经巴昔卡出中国境, 而后进入印度、孟加拉国, 最后注入印度洋[28]。在雅鲁藏布江流域, 随着经度的增加, 植被类型逐渐由荒漠草原向高山草原演变, 进而发展为灌木草原[29]。上游地区以蒿属、针茅属、狼尾草属、嵩草属等为主, 是典型的荒漠草原群落[30]。中游地区除上述种类外, 还包括蔷薇科和豆科灌木, 以高寒草地群落和灌木群落为主[31—32]。在雅鲁藏布江中上游这一生态敏感区域开展野外样方调查, 可明确白草在该区域分布范围、白草群落特征及其与环境因子的关系, 为该流域植物多样性评估和保护利用提供科学依据, 对确保青藏高原主体作为我国重要生态屏障有着十分重要的科学意义。
1 材料与方法 1.1 研究区概况研究区位于西藏自治区雅鲁藏布江上游日喀则市仲巴县帕羊镇至中游林芝市米林县派镇, 研究样点经度(83°44′16″—94°51′44″E), 纬度(28°11′24″—29°6′1″N), 平均海拔3890.65 m。
1.2 研究方法 1.2.1 样地设置根据白草在西藏的分布范围和生境特征, 在雅鲁藏布江中上游地区确定26个典型分布区域(图 1)。

|
| 图 1 雅鲁藏布江中上游白草主要分布区样地 Fig. 1 Sampling sites of P. centrasiaticum communities in the upper and middle reaches of the Yarlung Zangbo River |
通过路线踏察在研究区域选取白草分布较集中的样地, 利用手持GPS仪(eTrex Venture, Garmin, USA)记录采样点的经度、纬度、海拔等信息, 并根据经纬度信息在International Water Management Institute(IWMI)在线气候服务平台(http://wcatlas.iwmi.org/Default.asp)获取降雨量和平均温度数据(表 1)。
| 群落分布区域 Community distribution site |
经度 Longitude |
纬度 Latitude |
海拔 Altitude/m |
降水量 Precipitation/(mm/a) |
平均温度 Temperature/℃ |
| 昂仁县 | 87°27′36″ | 29°6′1″ | 4024 | 223.63 | 3.35 |
| 白朗县 | 89°21′42″ | 29°4′49″ | 3870 | 153.08 | 1.97 |
| 定结县 | 87°44′14″ | 28°11′59″ | 4254 | 209.09 | —1.73 |
| 定日县 | 86°55′36″ | 28°42′8″ | 4404 | 174.69 | 0.41 |
| 岗巴县 | 88°40′18″ | 28°18′22″ | 4567 | 170.90 | —1.03 |
| 贡嘎县 | 90°49′37″ | 29°16′30″ | 3549 | 315.27 | 5.60 |
| 吉隆县 | 85°18′42″ | 28°52′36″ | 4096 | 210.88 | —3.01 |
| 加查县 | 92°46′24″ | 29°4′37″ | 3104 | 284.14 | 1.13 |
| 江孜县 | 89°32′36″ | 28°57′27″ | 4010 | 221.71 | 3.68 |
| 康马县 | 89°38′24″ | 28°46′56″ | 4091 | 251.27 | —0.59 |
| 拉孜县 | 87°40′43″ | 29°11′45″ | 3857 | 277.65 | 4.68 |
| 朗县 | 93°17′59″ | 28°59′41″ | 3039 | 396.85 | 4.28 |
| 米林县 | 94°51′44″ | 29°30′59″ | 2883 | 589.67 | 3.50 |
| 乃东区 | 91°39′50″ | 29°15′18″ | 3520 | 266.50 | 4.63 |
| 南木林县 | 89°15′12″ | 29°26′28″ | 3989 | 248.12 | 4.62 |
| 尼木县 | 90°10′38″ | 29°26′16″ | 3763 | 211.17 | 3.00 |
| 聂拉木县 | 85°58′58″ | 28°11′24″ | 3783 | 289.06 | —0.64 |
| 仁布县 | 89°48′51″ | 29°16′46″ | 3815 | 168.50 | 1.82 |
| 萨嘎县 | 85°15′57″ | 29°19′13″ | 4423 | 215.75 | —1.52 |
| 萨迦县 | 88°27′32″ | 29°10′5″ | 3895 | 262.53 | 4.25 |
| 桑日县 | 91°53′45″ | 29°16′57″ | 3535 | 250.59 | 3.43 |
| 桑珠孜区 | 88°40′13″ | 29°19′32″ | 3818 | 293.99 | 5.14 |
| 谢通门县 | 88°19′14″ | 29°29′13″ | 4284 | 154.94 | 0.82 |
| 亚东县 | 89°23′59″ | 28°12′59″ | 4460 | 149.36 | —0.98 |
| 扎囊县 | 91°21′18″ | 29°19′22″ | 3581 | 304.38 | 5.78 |
| 仲巴县 | 83°44′16″ | 29°52′27″ | 4543 | 240.02 | —1.01 |
2020年8—9月, 在植物生长旺盛期, 于每个样地选择群落结构和组成分布均匀的区域, 随机选取3个50 cm×50 cm的调查研究样方, 调查群落内各物种的高度、盖度(对于样方内存在的矮小灌木, 如砂生槐等, 在本实验中, 按照草本植物的调查方法处理)。分类齐地面剪取样方内各物种地上部分, 带回室内于65℃下烘干至恒重, 记录干重, 作为各物种的地上生物量。地上总生物量采用各物种地上生物量干重之和计算。
1.2.3 植物功能群的划分植物功能群参照王长庭[33]及李里[34]的标准进行划分。根据拉琼[35]等对雅鲁藏布江上游源头区至下游对河岸植物物种丰富度分布格局的研究结果将本研究中的植物功能群划分为禾本科(Gramineae)、豆科(Leguminosae)、菊科(Asteraceae)和杂类草(Weeds)。
1.2.4 地下生物量测定地下生物量的测定采用土柱法, 用特制的圆形钢筒取样。钢筒直径5 cm, 高30 cm, 端口削尖。地下生物量的取样与地上生物量同步进行, 每个地上的生物量样方内设一个地下生物量样方。每个样方取3层, 分0—10 cm, 10—20 cm, 20—30 cm。样品按层装入塑封袋中, 标记好带回室内。用80目筛冲洗、分离, 将根系按层装入纸袋中, 在65℃烘箱内烘干至恒重, 称其干重。根据土柱内各层地下生物量测定值, 按比例换算成单位面积地下生物量。
1.2.5 土壤养分测定用上述圆形钢筒进行土壤样品取样, 每个样方采集3钻0—10 cm表层土, 每个样点的土样混合装入密封袋带回实验室, 自然风干后去除杂草、根系、石块等, 过筛备用。土壤养分调查参考鲍士旦《土壤农化分析》[36], 土壤pH测定采用pH计测定, 土壤含水量采用称重法测定, 土壤有机质(SOM)采用重铬酸钾氧化法测定, 土壤全氮(TNC)采用重铬酸钾—硫酸消化法测定, 全磷(TPC)采用高氯酸—硫酸法测定, 速效磷(APC)采用碳酸氢钠浸提—钼锑抗比色法测定, 速效氮(ANC)采用扩散法测定。
1.2.6 数据分析丰富度:
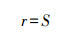
|
植物物种相对重要值计算:
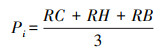
|
Shannon-Weiner指数计算:
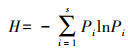
|
Simpson指数计算:
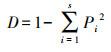
|
物种均匀度指数计算:
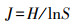
|
式中:S为所调查样方内物种总数, RC为相对盖度, RH为相对高度, RB为相对生物量。
用IBM SPSS Statistics 20进行单因素方差分析(One-way ANOVA)和Duncan多重比较(显著水平0.05), 并用于计算指标间的Pearson线性相关系数;采用Canoco 4.5软件进行白草群落特征与环境因子之间的冗余分析(Redundancy analysis, RDA)。用Microsoft Excel 2016对原始数据进行汇总并进行图表的绘制, 数据以平均值±标准误差(Mean ± SE)来表示。
2 结果与分析 2.1 雅鲁藏布江中上游地区白草主要分布区植物群落结构组成和植物多样性特征从雅鲁藏布江上游仲巴县帕羊镇至米林县派镇白草主要分布区26个样地中, 各白草群落共记录到植物有35科96属共119个物种。其中含种数较多的有禾本科(Gramineae)15属16种, 代表物种:白草、固沙草、垂穗披碱草(Elymus nutans)等;菊科(Asteraceae)14属20种, 代表物种:藏沙蒿、白莲蒿、狗娃花(Heteropappus hispidus)等;豆科(Leguminosae)9属12种, 代表物种:砂生槐、笔直黄耆(Astragalus strictus)、黄华(Thermopsis lupinoides)等。
根据样地群落中物种重要值大小划分为白草群落、藏沙蒿群落、砂生槐群落、白莲蒿群落和固沙草群落5种群落类型, 群落类型所在样地和次优势种统计见表 2。
| 群落编号 Community code |
群落类型 Community type |
样地 Sample sites |
次优势种 Subdominant species |
| Ⅰ | 白草群落 P. centrasiaticum community |
昂仁、白朗、定结、定日、吉隆、江孜、聂拉木、南木林、仁布、萨迦、桑珠孜、谢通门、仲巴、贡嘎、加查、拉孜、乃东、尼木 | 笔直黄耆、藏沙蒿、固沙草、牻牛儿苗(Erodium stephanianum)、垂穗披碱草、三角草(Trikeraia hookeri)、砂生槐、针茅(Stipa capillata)、猪毛蒿(Artemisia scoparia) |
| Ⅱ | 藏沙蒿群落 A. wellbyi community |
岗巴、康马、亚东、朗县 | 白草、马蔺(Iris lactea)、冰草(Agropyron cristatum) |
| Ⅲ | 砂生槐群落 S. moorcroftiana community |
桑日、扎囊 | 固沙草、白草 |
| Ⅳ | 白莲蒿群落 A. sacrorum community |
米林 | 藏沙蒿、白草 |
| Ⅴ | 固沙草群落 O. thoroldii community |
萨嘎 | 白草、笔直黄耆 |
雅鲁藏布江中上游白草主要分布区植物群落多样性特征统计结果见表 3。各白草群落物种丰富度变幅较大, 桑日县最低(S=12), 江孜县最高(S=37), 植物物种数量相差25个。Shannon-Weiner指数(H)变幅为1.28(仁布县)—2.30(米林县);Simpson指数(D)变幅为0.52(拉孜县)—0.86(米林县);物种均匀度指数(J)变幅为0.54(昂仁县)—0.80(朗县)。
| 群落分布区域 Community distribution site |
物种丰富度 Richness S |
Shannon-Weiner指数 Shannon-Weiner index H |
Simpson指数 Simpson index D |
物种均匀度指数 Species evenness inex J |
| 昂仁县 | 27 | 1.65±0.05 | 0.59±0.00 | 0.54±0.02 |
| 白朗县 | 26 | 1.54±0.07 | 0.60±0.01 | 0.59±0.01 |
| 定结县 | 30 | 2.11±0.10 | 0.79±0.03 | 0.72±0.04 |
| 定日县 | 27 | 1.82±0.15 | 0.74±0.03 | 0.70±0.03 |
| 岗巴县 | 24 | 2.07±0.20 | 0.80±0.04 | 0.73±0.03 |
| 贡嘎县 | 25 | 2.21±0.09 | 0.83±0.01 | 0.78±0.01 |
| 吉隆县 | 29 | 1.85±0.20 | 0.69±0.06 | 0.64±0.05 |
| 加查县 | 18 | 1.95±0.08 | 0.81±0.01 | 0.76±0.01 |
| 江孜县 | 37 | 2.24±0.18 | 0.82±0.05 | 0.71±0.05 |
| 康马县 | 30 | 2.29±0.07 | 0.85±0.01 | 0.76±0.02 |
| 拉孜县 | 19 | 1.60 | 0.52 | 0.54 |
| 朗县 | 16 | 2.03±0.01 | 0.84±0.00 | 0.80±0.01 |
| 米林县 | 28 | 2.30±0.08 | 0.86±0.01 | 0.78±0.00 |
| 乃东区 | 14 | 1.44±0.05 | 0.70±0.01 | 0.69±0.02 |
| 南木林县 | 16 | 1.39±0.11 | 0.63±0.04 | 0.62±0.03 |
| 尼木县 | 21 | 1.78±0.07 | 0.76±0.01 | 0.70±0.01 |
| 聂拉木县 | 23 | 1.72±0.09 | 0.73±0.05 | 0.70±0.04 |
| 仁布县 | 18 | 1.28±0.09 | 0.54±0.02 | 0.55±0.01 |
| 萨嘎县 | 17 | 1.93±0.04 | 0.79±0.01 | 0.72±0.00 |
| 萨迦县 | 15 | 1.48±0.18 | 0.64±0.06 | 0.66±0.04 |
| 桑日县 | 12 | 1.37±0.06 | 0.60±0.01 | 0.59±0.00 |
| 桑珠孜区 | 18 | 1.31±0.28 | 0.55±0.07 | 0.58±0.05 |
| 谢通门县 | 29 | 1.94±0.32 | 0.67±0.10 | 0.61±0.09 |
| 亚东县 | 21 | 2.05±0.10 | 0.83±0.01 | 0.77±0.02 |
| 扎囊县 | 13 | 1.40±0.03 | 0.65±0.01 | 0.63±0.03 |
| 仲巴县 | 20 | 1.76±0.04 | 0.73±0.00 | 0.69±0.01 |
| 因资料遗失, 拉孜县有一个样方调查表 | ||||
对各群落内白草高度、盖度、生物量和重要值进行统计分析, 结果见表 4。白草平均高度为64.22 cm, 变幅为26.97 cm(仲巴县)—96.38 cm(米林县);白草平均盖度为56.57%, 变幅为10.33%(桑日县)—90.67%(萨迦县);白草平均生物量为250.87 g/m2, 变幅为48.77 g/m2(桑日县)—507.84 g/m2(尼木县);白草平均重要值为39.38%, 变幅为11.88%(桑日县)—67.52%(拉孜县)。
| 群落分布区域 Community distribution site |
白草高度 Height/cm |
白草盖度 Coverage/% |
白草生物量 Biomass/(g/m2) |
白草重要值 Importance value/% |
| 昂仁县 | 90.13±15.13 | 78.00±10.00 | 377.52±48.40 | 62.47±1.09 |
| 白朗县 | 67.71±3.32 | 85.25±1.31 | 402.38±6.21 | 61.75±0.76 |
| 定结县 | 51.25±1.43 | 66.00±7.02 | 195.36±20.79 | 38.96±4.89 |
| 定日县 | 43.82±8.03 | 65.00±0.58 | 179.40±1.59 | 45.83±3.35 |
| 岗巴县 | 36.80±4.62 | 33.67±1.86 | 74.07±4.08 | 21.91±3.06 |
| 贡嘎县 | 80.12±6.23 | 46.67±1.67 | 345.33±12.33 | 32.81±3.14 |
| 吉隆县 | 48.27±7.67 | 80.67±2.96 | 245.23±9.01 | 52.78±5.88 |
| 加查县 | 82.56±4.20 | 51.67±6.01 | 314.13±36.54 | 29.18±2.79 |
| 江孜县 | 61.50±5.75 | 50.00±10.00 | 222.00±44.40 | 33.38±7.97 |
| 康马县 | 66.00±7.38 | 27.00±3.79 | 109.08±15.30 | 17.09±1.87 |
| 拉孜县 | 38.20 | 88.00 | 228.80 | 67.52 |
| 朗县 | 82.33±4.10 | 36.00±3.06 | 241.92±20.53 | 20.92±1.06 |
| 米林县 | 96.38±1.38 | 25.00±5.00 | 198.00±39.60 | 14.06±3.61 |
| 乃东区 | 92.86±3.02 | 57.67±1.45 | 389.83±9.82 | 39.94±1.04 |
| 南木林县 | 76.80±11.07 | 79.33±5.21 | 399.84±26.24 | 55.61±4.12 |
| 尼木县 | 87.33±0.73 | 69.00±2.08 | 507.84±15.32 | 39.32±1.87 |
| 聂拉木县 | 35.22±0.78 | 55.00±4.04 | 116.60±8.57 | 45.12±6.72 |
| 仁布县 | 40.20±5.49 | 74.33±6.17 | 205.16±17.04 | 66.51±1.03 |
| 萨嘎县 | 42.17±4.38 | 26.00±4.58 | 67.60±11.91 | 22.50±1.95 |
| 萨迦县 | 90.27±5.66 | 90.67±2.96 | 464.21±15.17 | 54.89±6.36 |
| 桑日县 | 49.72±3.57 | 10.33±0.88 | 48.77±4.16 | 11.88±0.77 |
| 桑珠孜区 | 78.07±9.95 | 84.67±0.33 | 447.04±1.76 | 64.89±5.62 |
| 谢通门县 | 91.33±11.67 | 80.50±5.50 | 421.82±28.82 | 55.18±9.65 |
| 亚东县 | 35.13±1.11 | 22.33±1.45 | 50.92±3.31 | 14.22±1.29 |
| 扎囊县 | 87.60±0.53 | 25.00±2.89 | 178.00±20.55 | 22.10±1.14 |
| 仲巴县 | 26.97±0.22 | 77.00±4.51 | 98.56±5.77 | 46.89±0.29 |
将群落地下0—10 cm, 10—20 cm, 20—30 cm土层地下生物量分别统计, 结果如表 5。各群落0—10 cm土层中, 地下生物量均为最大, 随土层的加深, 地下生物量降低。各群落中, 0—10 cm、10—20 cm、20—30 cm土层和总的地下生物量平均为1969.30 g/m2, 921.77 g/m2, 483.16 g/m2和3374.23 g/m2;各土层地下生物量占比分别为60.38%, 26.36%和13.26%。
| 群落分布区域 Community distribution site |
地下生物量(0—10 cm) Belowground biomass in 0—10cm soil layer/ (g/m2) |
占比(0—10 cm) The proportion of biomass in 0—10cm soil layer/% |
地下生物量(10—20 cm) Belowground biomass in 10—20cm soil layer/ (g/m2) |
占比(10—20 cm) The proportion of biomass in 10—20cm soil layer/% |
地下生物量(20—30 cm) Belowground biomass in 20—30cm soil layer/ (g/m2) |
占比(20—30 cm) The proportion of biomass in 20—30cm soil layer/% |
地下生物量 Belowground biomass/ (g/m2) |
| 昂仁县 | 1100.64±315.81a | 63.72 | 433.12±122.33 | 25.07 | 193.63±62.89b | 11.21 | 1727.39±499.71 |
| 白朗县 | 1319.75±155.59a | 66.93 | 514.65±60.36b | 26.10 | 137.58±71.58c | 6.98 | 1971.97±240.25 |
| 定结县 | 1600.00±342.38a | 63.05 | 621.66±90.58b | 24.50 | 315.92±66.31b | 12.45 | 2537.58±485.05 |
| 定日县 | 1793.63±16.38a | 65.19 | 694.69±68.19b | 25.25 | 263.27±20.66c | 9.57 | 2751.59±64.52 |
| 岗巴县 | 2383.01±39.94a | 70.01 | 647.13±44.52b | 19.01 | 373.67±72.32c | 10.98 | 3403.82±23.35 |
| 贡嘎县 | 1900.64±206.37a | 56.60 | 957.96±61.43b | 28.53 | 499.36±82.84b | 14.87 | 3357.96±336.40 |
| 吉隆县 | 1467.52±189.36a | 72.73 | 387.26±54.96b | 19.19 | 163.06±87.07b | 8.08 | 2017.83±310.17 |
| 加查县 | 1696.82±112.14a | 61.21 | 587.69±124.96b | 21.20 | 487.47±88.52b | 17.59 | 2771.97±269.63 |
| 江孜县 | 1284.08±75.06a | 70.19 | 402.55±63.71b | 22.01 | 142.68±34.68c | 7.80 | 1829.30±33.93 |
| 康马县 | 2145.22±538.16a | 56.43 | 1187.26±390.37 | 31.23 | 468.79±95.46b | 12.33 | 3801.27±1016.27 |
| 拉孜县 | 692.99±86.92a | 60.18 | 346.50±54.01b | 30.09 | 112.10±38.24c | 9.73 | 1151.59±156.97 |
| 朗县 | 2542.68±135.10a | 52.69 | 1350.32±428.78b | 27.98 | 932.48±280.21b | 19.32 | 4825.48±626.81 |
| 米林县 | 2741.40±257.43a | 50.68 | 1812.31±355.01a | 33.50 | 856.05±171.31b | 15.82 | 5409.77±764.85 |
| 乃东区 | 2150.32±269.26a | 51.67 | 1494.69±77.52b | 35.92 | 516.35±119.04c | 12.41 | 4161.36±400.95 |
| 南木林县 | 1467.52±462.66a | 61.15 | 636.94±129.34 | 26.54 | 295.54±84.81b | 12.31 | 2400.00±625.16 |
| 尼木县 | 2680.25±470.86a | 59.91 | 1136.31±283.36b | 25.40 | 657.32±171.47b | 14.69 | 4473.89±923.56 |
| 聂拉木 | 907.01±92.14a | 64.73 | 275.16±35.67b | 19.64 | 219.11±31.82b | 15.64 | 1401.27±134.82 |
| 仁布县 | 636.94±60.79a | 62.19 | 229.30±20.38b | 22.39 | 157.96±64.92b | 15.42 | 1024.20±91.34 |
| 萨嘎县 | 1987.26±669.93a | 58.12 | 810.19±291.96a | 23.70 | 621.66±139.76a | 18.18 | 3419.11±1057.76 |
| 萨迦县 | 1467.52±498.11a | 66.72 | 580.89±380.00 | 26.41 | 151.17±44.26b | 6.87 | 2199.58±864.66 |
| 桑日县 | 3543.10±649.47a | 55.78 | 1746.07±340.63b | 27.49 | 1063.27±213.41b | 16.74 | 6352.44±650.62 |
| 桑珠孜区 | 1151.59±166.00a | 60.43 | 555.41±44.42b | 29.14 | 198.73±39.58c | 10.43 | 1905.73±176.59 |
| 谢通门县 | 3373.25±451.33a | 55.47 | 1870.06±107.37b | 30.75 | 837.37±55.92c | 13.77 | 6080.68±417.10 |
| 亚东县 | 4081.53±257.67a | 54.47 | 2019.53±602.28b | 26.95 | 1392.78±302.35b | 18.59 | 7493.84±346.05 |
| 扎囊县 | 3016.56±301.41a | 55.47 | 1610.19±446.85b | 29.61 | 811.89±212.19b | 14.93 | 5438.64±414.87 |
| 仲巴县 | 2070.49±724.47a | 54.18 | 1058.17±410.37a | 27.69 | 692.99±193.56a | 18.13 | 3821.66±1222.81 |
| 不同小写字母表示白草主要分布区不同土层地下生物量差异显著(P < 0.05) | |||||||
各样地中, 亚东县地下生物量最高, 达到7493.84 g/m2, 仁布县地下生物量最低, 为1024.20 g/m2。在0—10 cm土层, 亚东县生物量最高, 为4081.53 g/m2, 吉隆县生物量占比最高, 为72.73%;在10—20 cm土层中, 亚东县生物量最高, 为2019.53 g/m2, 乃东区生物量占比最高, 为35.92%;在20—30 cm土层中, 亚东县生物量最高, 为1392.78 g/m2, 朗县生物量占比最高, 为19.32%。
2.3 白草种群特征及群落特征与主要环境因子间的相关性白草种群特征与主要环境因子间相关关系如表 6所示。白草高度与海拔(r=-0.60), 白草高度与土壤pH(r=-0.53), 白草生物量与土壤pH(r=-0.55)呈极显著负相关关系(P < 0.01)。白草高度与全磷含量(r=0.42), 白草盖度与速效氮含量(r=0.43), 白草盖度与有机质含量(r=0.41), 白草生物量与全磷含量(r=0.40), 白草重要值与速效氮含量(r=0.40), 白草重要值与有机质含量(r=0.46)呈显著正相关关系。
| 环境因子 Climatic factors |
白草高度 Height |
白草盖度 Coverage |
白草生物量 Biomass |
白草重要值 Importance value |
| 海拔Altitude | -0.60** | 0.21 | -0.27 | 0.21 |
| 含水量Soil moisture | 0.25 | 0.17 | 0.16 | 0.14 |
| 土壤PH Soil pH | -0.53** | -0.29 | -0.55** | -0.21 |
| 土壤全氮含量TNC | 0.13 | 0.36 | 0.21 | 0.39 |
| 土壤全磷含量TPC | 0.42* | 0.25 | 0.40* | 0.20 |
| 土壤速效氮含量ANC | 0.19 | 0.43* | 0.29 | 0.40* |
| 土壤速效磷含量APC | -0.08 | 0.26 | 0.09 | 0.22 |
| 土壤有机质含量SOC | -0.17 | 0.41* | 0.06 | 0.46* |
| *表示在0.05水平(双侧)上显著相关, **表示在0.01水平(双侧)上显著相关 | ||||
冗余分析(RDA)是一种回归分析与主成分分析相结合的排序方法, 是多响应变量回归分析的拓展[37], 能够整体反映环境因子与白草群落常见物种组成和个体分布之间的关系。将白草和群落中常见的12种植物(白草、砂生槐、笔直黄耆、猪毛蒿、菊叶香藜Chenopodium foetidum、狗娃花、风毛菊Saussurea japonica、固沙草、蒲公英Taraxacum mongolicum、平车前Plantago depressa、毛瓣棘豆Oxytropis sericopetala、垂穗披碱草、藏沙蒿)的重要值为基础数据建立响应变量矩阵, 以9个环境因子(7个土壤养分含量、年平均温度、年平均降雨量)作为解释变量进行RDA分析, 结果见图 2。

|
| 图 2 白草主要分布区常见物种重要值与环境因子的RDA排序 Fig. 2 RDA ordination of environmental factors and importance value of common species in P. centrasiaticum distribution areas |
轴1和轴2共解释全部方差的40.60%, 在环境—群落的解释中起主导作用。其中RDA1轴与土壤pH、年平均降雨量、全氮含量、速效氮含量、速效磷含量、有机质含量、土壤含水量有较强的关系性, 单独解释了方差的23.30%, 说明土壤养分因子及降雨量对白草群落结构产生影响。RDA2轴与年平均温度、全磷含量有较强相关性, 单独解释变异的17.30%。总体上, 白草主要分布区土壤pH与其它土壤理化特征之间呈负相关, 白草重要值和土壤含水量、全氮、全磷、速效氮、速效磷、有机质含量有较强的相关性。
2.4 白草主要分布区聚类分析利用白草主要分布区的海拔数据对雅鲁藏布江中上游26个样点进行聚类分析, 将雅鲁藏布江中上游地区白草分布区划分为梯度Ⅰ、梯度Ⅱ和梯度Ⅲ, 见图 3。

|
| 图 3 雅鲁藏布江中上游白草主要分布区聚类图 Fig. 3 Clusters of P. centrasiaticum distribution areas in the middle and upper reaches of the Yarlung Zangbo River |
梯度Ⅰ(2883—3104 m, 平均海拔3008.67 m), 包含加查县、朗县和米林县3个样点;梯度Ⅱ(3520 m—4096 m, 平均海拔3824.75 m), 包含萨迦县、桑珠孜区、南木林县等16个样点;梯度Ⅲ(4254 m—4567 m, 平均海拔4419.29 m), 包含谢通门县、定日县、定结县等7个样点。
2.4.1 不同海拔植株功能群特征不同植物功能群在不同海拔梯度的高度、盖度、地上生物量和重要值如图 4所示。26个样地不同功能群平均高度顺序为:禾本科(32.78 cm)>豆科(20.11 cm)>菊科(17.00 cm)>杂类草(9.16 cm)。各功能群的平均盖度顺序为:禾本科(65.50%)>菊科(14.25%)>豆科(11.01%)>杂类草(9.01%)。各功能群平均地上生物量顺序为:豆科(327.78 g/m2)>禾本科(305.15 g/m2)>菊科(255.55 g/m2)>杂类草(67.23 g/m2)。各功能群平均重要值顺序为:禾本科(52.51%)>菊科(16.90%)>豆科(16.04%)>杂类草(14.55%)。

|
| 图 4 不同海拔梯度植株功能群特征 Fig. 4 The Characteristics of plant functional groups in different altitude gradients 不同大写字母表示相同海拔不同植物功能群特征差异显著, 不同小写字母表示相同植物功能群特征在不同海拔差异显著(P < 0.05) |
豆科、禾本科、菊科和杂类草功能群的高度均随着海拔上升而显著降低(P < 0.05)。禾本科功能群的盖度在梯度Ⅱ(70.22%)显著高于梯度Ⅰ(40.93%)(P < 0.05), 与梯度Ⅲ(65.27%)差异不显著;菊科功能群的盖度(42.29%)在梯度Ⅰ最大, 显著高于梯度Ⅱ(8.44%)和梯度Ⅲ(15.52%)(P < 0.05);豆科和杂类草功能群的盖度在3个海拔下差异不显著。菊科功能群的地上生物量在梯度Ⅰ(1255.58 g/m2)显著高于梯度Ⅱ(118.42 g/m2)和梯度Ⅲ(140.38 g/m2)(P < 0.05), 杂类草功能群的地上生物量在梯度Ⅰ(178.01 g/m2)显著大于梯度Ⅱ(42.45 g/m2)(P < 0.05), 与梯度Ⅲ(76.40 g/m2)差异不显著;豆科和禾本科功能群地上生物量在3个海拔下差异不显著。禾本科功能群重要值在梯度Ⅱ(55.85%)和梯度Ⅲ(55.58%)差异不显著, 显著高于梯度Ⅰ(27.50%)(P < 0.05);菊科功能群的重要值在梯度Ⅰ(39.53%)显著高于梯度Ⅱ(12.47%)和梯度Ⅲ(17.32%)(P < 0.05);豆科和杂类草功能群的重要值在3个海拔下差异不显著。
2.4.2 不同海拔植物群落地下生物量分配不同海拔白草群落的地下生物量占总生物量比例的计算结果如图 5所示。随着海拔的增大, 地下生物量占比显著增加(P < 0.05)。在梯度Ⅲ, 地下生物量占比为89.08%, 显著高于梯度Ⅱ(77.37%)和梯度Ⅰ(65.44%)(P < 0.05), 梯度Ⅱ地下生物量占比显著高于梯度Ⅰ(P < 0.05)。

|
| 图 5 不同海拔地下生物量占比 Fig. 5 Percentage of underground biomass in different altitude gradients 不同小写字母表示地下生物量占比在不同海拔差异显著(P < 0.05) |
植物群落多样性在各海拔梯度的变化见表 7, 群落物种丰富度在3个海拔梯度差异不显著, 其变化趋势为:梯度Ⅲ(24.00)>梯度Ⅱ(21.44)>梯度Ⅰ(20.67)。群落多样性指数(H、D、J)在不同海拔的变化趋势为:梯度Ⅰ>梯度Ⅲ>梯度Ⅱ, 梯度Ⅰ显著大于梯度Ⅱ(P < 0.05), 梯度Ⅲ和另2个梯度的差异不显著。
| 海拔 Altitude |
物种丰富度 S |
Shannon-Weiner指数 H |
Simpson指数 D |
物种均匀度指数 J |
| 梯度Ⅰ Altitude gradient Ⅰ | 20.67±3.71a | 2.09±0.11a | 0.84±0.01a | 0.78±0.01a |
| 梯度Ⅱ Altitude gradient Ⅱ | 21.44±1.77a | 1.66±0.08b | 0.67±0.03b | 0.64±0.02b |
| 梯度Ⅲ Altitude gradient Ⅲ | 24.00±1.85a | 1.95±0.05 | 0.76±0.02 | 0.71±0.02 |
| 不同小写字母表示不同海拔植物群落多样性指数差异显著(P < 0.05) | ||||
功能群被认为是与系统的某种功能直接相关的物种群, 功能群内物种对系统的作用有很大相似性, 因而用功能群数作为物种多样性的指标研究多样性与生态系统功能的关系值得肯定[33, 38]。用功能群以解释不同演替梯度或生境中物种多样性格局的变化及与生态系统功能的关系, 使复杂的问题较为简化且更具说服力[34, 39]。因研究角度的不同, 不同科学家对植物功能群的划分也不同[34, 40—41]。拉琼[35]研究表明:雅鲁藏布江上游源头区至下游出现频度最高的前三大科分别是菊科、禾本科和豆科, 这与本研究的结果相同。以白草为代表的禾本科功能群叶面积和叶位相对较大, 在雅鲁藏布江中上游26个样地中有最大的高度、盖度和重要值, 在生长方面占据光照和养分的优势[40, 42—43], 抑制了豆科、菊科和杂类草的生长。功能群之间具有补偿作用, 存在生态位重叠的现象, 当一种功能群的数量或生产力下降时, 会为另一些功能群提供更大的空间, 使之占据更多的资源[34]。梯度Ⅰ的禾本科功能群盖度低(40.93%), 菊科功能群迅速进行补偿, 提高其高度(36.34 cm)、盖度(42.29%)、生物量(1255.58 g/m2)和重要值(39.53%)。其它研究也表明禾本科牧草盖度的降低将导致毒害草和杂类草增多;禾本科牧草盖度的提升, 使得杂类草减少[40—44]。植物的这种生态补偿作用是生命实体对环境变化和干扰的一种响应对策, 通过功能群之间的相互作用来实现植物功能群对群落稳定性的影响[34]。
3.2 雅鲁藏布江中上游白草主要分布区群落结构特征与环境解释植物之间均存在着广泛而复杂的相互作用[45], 植物种群结构可反映种群的数量动态、发展趋势与环境之间的相互关系[46], 也可揭示植物种群与生境相适应的结果, 对预测群落未来发展趋势具有重要意义[47—48]。植物种群多样性特征及其空间分布格局能够反映环境对种群内个体生长和生存的影响, 也能反映和指示植物种群的生态适应对策[49]。在草地发展过程中, 受到多种环境因素的影响, 其中降水和温度作为重要的气候因子, 成为草地生长发育因素研究中的重点[50—51]。受海拔和经度的影响, 雅鲁藏布江流域自西向东形成湿热增加的梯度, 植物多样性沿该梯度逐渐增加[29]。拉琼[35]等沿海拔梯度从雅鲁藏布江上游源头区至下游记录的维管束植物238种, 物种分布格局呈现出物种丰富度在下游最高、中游偏低、上游略上升的趋势。另外, 在拉萨河上游的研究[52]发现物种丰富度在山腰低, 在山脚和山顶高, 其成因被认为是半山腰高密度灌木使物种丰富度降低。在本实验中, 白草主要分布区共记录植物物种数119个, 随海拔上升, 群落丰富度增加。在白草主要分布区, 禾本科植物在不同的植物功能群中高度和盖度最大, 占据光照和养分的优势, 导致杂草物种的排除[53], 使本实验记录的植物物种总数低于拉琼的调查。梯度Ⅰ温度高、降雨多, 白莲蒿和藏沙蒿等成为白草主要分布区的优势种或者次优势种, 相对高大的菊科植物降低了样方内物种丰富度[54], 使其物种丰富度低于梯度Ⅱ。同理, 梯度Ⅱ的物种丰富度低于梯度Ⅲ, 这是物种之间对光照竞争的结果[55]。白草主要分布区土壤pH与其它土壤理化特征呈负相关, 与罗巧玉[45]对发草适生地土壤以及王文颖[56]对高寒藏嵩草草甸土壤的研究结果相同。土壤pH与海拔呈正相关关系, 禾本科和杂类草植物数量随土壤pH值逐渐增加[57], 因而白草群落丰富度随海拔上升增加。白草主要分布区禾本科植物的重要值为梯度Ⅱ>梯度Ⅲ>梯度Ⅰ, 而群落多样性指数呈现相反趋势。这与周选博[43]和张峰[58]研究得出禾本科植物重要值增加, 将导致Shannon-Weiner指数、Simpson指数和物种均匀度指数变小这一结果相同。
白草主要分布区各群落中, 地下生物量随土层深度增加而递减, 在0—10 cm土层地下生物量占比达到60.38%。表层通气条件较好, 地下生物量大, 随着土壤深度的增加, 土壤温度、含水量和通气条件变劣, 地下生物量相应减少[59]。雅鲁藏布江中上游白草群落的地下生物量大都分布在表土层中以获取更多的热量、水分和矿质营养, 表明高原植物对严酷环境有适应性。青藏高原高寒地区, 强辐射、生长季节短等因素阻碍植物生长[60], 寒冷天气降低植物细胞分裂的速度, 植物难以发育成大个体[61], 导致高海拔地区植物个体矮小, 地上生物量降低;同时, 以白草为代表的多年生植物常将更多生物量投资到根部[16, 62], 使雅鲁藏布江中上游流域白草主要分布区的地下生物量占比随海拔上升显著增加。
4 结论以上研究对雅鲁藏布江中上游地区白草主要分布区植物群落特征进行调查, 发现白草主要分布区有119个植物物种, 属35科96属, 表土层地下生物量占比最高。RDA分析表明白草重要值和土壤含水量、全氮、全磷、速效氮、速效磷、有机质含量有较强的相关性。聚类分析揭示4个功能群的高度均随海拔上升而下降, 群落物种丰富度和群落地下生物量随海拔上升而占比增加。以上结果反映出白草是雅鲁藏布江中上游广泛分布的乡土草种, 其群落特征多样, 与环境因子有较强的相关性。
| [1] |
李海东, 沈渭寿, 方颖, 燕守广, 张慧, 赵卫. 雅鲁藏布江中游河岸带几种主要沙生植物种群点格局分析. 植物生态学报, 2011, 35(8): 834-843. |
| [2] |
Klanderud K, Vandvik V, Goldberg D. The importance of Biotic vs. abiotic drivers of local plant community composition along regional bioclimatic gradients. PLoS One, 2015, 10(6): e0130205. DOI:10.1371/journal.pone.0130205 |
| [3] |
Xu M H, Zhang S X, Wen J, Yang X Y. Multiscale spatial patterns of species diversity and biomass together with their correlations along geographical gradients in subalpine meadows. PLoS One, 2019, 14(2): e0211560. DOI:10.1371/journal.pone.0211560 |
| [4] |
Walter H. Vegetation of the Earth and Ecological Systems of the Geo-Biosphere. 2nd ed. New York: Springer, 1979: 1-41.
|
| [5] |
唐志尧, 方精云. 植物物种多样性的垂直分布格局. 生物多样性, 2004, 12(1): 20-28. |
| [6] |
Gaston K J. Global patterns in biodiversity. Nature, 2000, 405(6783): 220-227. DOI:10.1038/35012228 |
| [7] |
Acker S A, Gregory S V, Lienkaemper G, McKee W A, Swanson F J, Miller S D. Composition, complexity, and tree mortality in riparian forests in the central Western Cascades of Oregon. Forest Ecology and Management, 2003, 173(1/3): 293-308. |
| [8] |
Kuglerová L, Jansson R, Ågren A, Laudon H, Malm-Renöfält B. Groundwater discharge creates hotspots of riparian plant species richness in a boreal forest stream network. Ecology, 2014, 95(3): 715-725. DOI:10.1890/13-0363.1 |
| [9] |
Sabo J L, Sponseller R, Dixon M, Gade K, Harms T, Heffernan J, Jani A, Katz G, Soykan C, Watts J, Welter J. Riparian zones increase regional species richness by harboring different, not more, species. Ecology, 2005, 86(1): 56-62. DOI:10.1890/04-0668 |
| [10] |
O'Donnell J, Fryirs K, Leishman M R. Digging deep for diversity: riparian seed bank abundance and species richness in relation to burial depth. Freshwater Biology, 2014, 59(1): 100-113. DOI:10.1111/fwb.12249 |
| [11] |
中国科学院中国植物志编辑委员会. 中国植物志. 北京: 科学出版社, 1990, 368. |
| [12] |
郭金丽, 李青丰. 克隆整合对白草克隆生长的影响. 中国草地学报, 2008, 30(6): 43-48. |
| [13] |
郭金丽. 三种根茎型禾草繁殖特性及其生态适应性研究[D]. 呼和浩特: 内蒙古农业大学, 2007.
|
| [14] |
Mehra P N, Remanandan P. Cytological investigations on W. himalayan panicoideae. Cytologia, 1973, 38(2): 259-270. DOI:10.1508/cytologia.38.259 |
| [15] |
李春红. 白草染色体核型与C-显带研究. 草业科学, 1988, 5(4): 30-33. |
| [16] |
高飞翔. 兰州本土地被植物调查与白草生态生物学特性研究[D]. 兰州: 兰州大学, 2008.
|
| [17] |
朱志诚, 贾东林. 白草群落生物量初步研究. 草业科学, 1996, 13(5): 1-3. |
| [18] |
Zhang G Y, Wang J W, Zhang H R, Fu G, Shen Z X. Comparative study of the impact of drought stress on P. centrasiaticum at the seedling stage in Tibet. Journal of Resources and Ecology, 2020, 11(3): 322-328. DOI:10.5814/j.issn.1674-764x.2020.03.010 |
| [19] |
任安芝, 高玉葆, 梁宇, 陈世苹, 刘爽, 刘宁. 白草和赖草无性系生长对干旱胁迫的反应. 中国沙漠, 1999, 19(S1): 30-34. |
| [20] |
曹仲华, 呼天明, 曹社会, 张中岳. 西藏山南地区6种野生牧草营养价值评定. 草地学报, 2010, 18(3): 414-420. |
| [21] |
薛树媛, 金海, 郭雪峰, 永西修, 巴雅斯胡良. 内蒙古荒漠草原优势牧草营养价值评价. 中国草地学报, 2007, 29(6): 22-27. |
| [22] |
李佃勇. 白草驯化及栽培技术的研究. 四川草原, 1983(2): 55-57. |
| [23] |
张怀山, 赵桂琴, 代立兰, 杨世柱, 王平. 黄土高原半干旱区狼尾草属牧草品比试验. 草原与草坪, 2014, 34(3): 42-45, 50-50. |
| [24] |
Szczęs'niak E. Pennisetum flaccidum (Cenchrus flaccidus) (Poaceae) - new anthropophyte in flora of Poland. Fragmenta Floristica et Geobotanica Polonica, 2011, 18(2): 295-300. |
| [25] |
李海燕, 杨允菲. 科尔沁草原区火电厂储灰池植被恢复过程中白草种群分蘖株的年龄结构. 生态学报, 2014, 34(20): 5907-5914. |
| [26] |
何大明, 汤奇成. 中国国际河流. 北京: 科学出版社, 2000: 127-137.
|
| [27] |
西藏在线. 雅鲁藏布江. (2018-03-20)[2021-07-09]. http://www.tibetol.cn/html/2013/hl_0917/1291.html.
|
| [28] |
何萍, 郭柯, 高吉喜, 史培军, 张永泽, 庄红翔. 雅鲁藏布江源头区的植被及其地理分布特征. 山地学报, 2005, 23(3): 267-273. |
| [29] |
Li C, Xu W L, Li Q K, Wang J S. Community structure and diversity distribution pattern of sandy plants in the middle and upper reaches of the Yarlung Zangbo River. Journal of Resources and Ecology, 2021, 12(1): 11-21. |
| [30] |
李晖, 文雪梅, 于顺利. 羌塘高原及雅鲁藏布江上游地区植物种质资源调查与评价. 植物分类与资源学报, 2013, 35(3): 327-334. |
| [31] |
Wang T, Wang J S, Ding Y K, Liu W J, Bao X T, LI C. Quantitative classification and ordination of plant communities in the upper and middle reaches of the Yarlung Zangbo River Basin. Journal of Resources and Ecology, 2019, 10(4): 389-396. DOI:10.5814/j.issn.1674-764x.2019.04.006 |
| [32] |
张剑, 覃家理, 邓莉兰, 李双智, 黄桂林. 西藏雅鲁藏布江中游陆生植被调查与评价. 林业资源管理, 2008(4): 118-123. |
| [33] |
王长庭, 龙瑞军, 丁路明. 高寒草甸不同草地类型功能群多样性及组成对植物群落生产力的影响. 生物多样性, 2004, 12(4): 403-409. |
| [34] |
李里, 刘伟. 退化草地植物功能群和物种丰富度与群落生产力关系的研究. 草地学报, 2011, 19(6): 917-921, 999-999. |
| [35] |
拉琼, 扎西次仁, 朱卫东, 许敏, 钟扬. 雅鲁藏布江河岸植物物种丰富度分布格局及其环境解释. 生物多样性, 2014, 22(3): 337-347. |
| [36] |
鲍士旦. 土壤农化分析(第三版). 北京: 中国农业出版社, 2000: 14-97.
|
| [37] |
Lozano E, García-Orenes F, Bárcenas-Moreno G, Jiménez-Pinilla P, Mataix-Solera J, Arcenegui V, Morugán-Coronado A, Mataix-Beneyto J. Relationships between soil water repellency and microbial community composition under different plant species in a Mediterranean semiarid forest. Journal of Hydrology and Hydromechanics, 2014, 62(2): 101-107. DOI:10.2478/johh-2014-0017 |
| [38] |
Bengtsson J. Which species? What kind of diversity? Which ecosystem function? Some problems in studies of relations between biodiversity and ecosystem function. Applied Soil Ecology, 1998, 10(3): 191-199. DOI:10.1016/S0929-1393(98)00120-6 |
| [39] |
Bonet A. Secondary succession of semi-arid Mediterranean old-fields in south-eastern Spain: insights for conservation and restoration of degraded lands. Journal of Arid Environments, 2004, 56(2): 213-233. DOI:10.1016/S0140-1963(03)00048-X |
| [40] |
王超, 王晓丽, 施建军, 吴建丽, 邢云飞, 才仁求吉, 义西措毛. 返青期休牧对玛多高寒草原植物群落特征的影响. 草地学报, 2021, 29(4): 763-771. |
| [41] |
谢乐乐, 王晓丽, 马玉寿, 王彦龙, 罗少辉, 秦金萍, 周选博. 春季休牧对三江源区藏嵩草草甸植被群落特征与物种多样性的影响. 青海畜牧兽医杂志, 2021, 51(1): 11-16, 22-22. |
| [42] |
李佳琪, 赵敏, 魏斌, 胡廷花, 于应文. 蘑菇圈形成对高寒草甸群落植被结构及稳定性的作用. 草业学报, 2018, 27(4): 1-9. |
| [43] |
周选博, 王晓丽, 马玉寿, 王彦龙, 罗少辉, 秦金萍, 谢乐乐. 返青期休牧初期对三江源区高寒草甸植被特征的影响. 青海大学学报, 2020, 38(6): 1-8. |
| [44] |
李青丰, 赵钢, 郑蒙安, 阿民布和. 春季休牧对草原和家畜生产力的影响. 草地学报, 2005, 13(S1): 53-56, 66-66. |
| [45] |
罗巧玉, 王彦龙, 杜雷, 刘念, 李丽, 马玉寿. 黄河源区发草适生地植物群落特征及其土壤因子解释. 草业学报, 2021, 30(4): 80-89. |
| [46] |
Xu M H, Ma L, Jia Y Y, Liu M. Integrating the effects of latitude and altitude on the spatial differentiation of plant community diversity in a mountainous ecosystem in China. PLoS One, 2017, 12(3): e0174231. DOI:10.1371/journal.pone.0174231 |
| [47] |
Barker C A, Turley N E, Orrock J L, Ledvina J A, Brudvig L A. Agricultural land-use history does not reduce woodland understory herb establishment. Oecologia, 2019, 189(4): 1049-1060. DOI:10.1007/s00442-019-04348-6 |
| [48] |
Litza K, Diekmann M. The effect of hedgerow density on habitat quality distorts species-area relationships and the analysis of extinction debts in hedgerows. Landscape Ecology, 2020, 35(5): 1187-1198. DOI:10.1007/s10980-020-01009-5 |
| [49] |
Albrecht M A, Becknell R E, Long Q. Habitat change in insular grasslands: Woody encroachment alters the population dynamics of a rare ecotonal plant. Biological Conservation, 2016, 196: 93-102. DOI:10.1016/j.biocon.2016.01.032 |
| [50] |
Gill R A, Kelly R H, Parton W J, Day K A, Jackson R B, Morgan J A, Scurlock J M O, Tieszen L L, Castle J V, Ojima D S, Zhang X S. Using simple environmental variables to estimate below-ground productivity in grasslands. Global Ecology and Biogeography, 2002, 11(1): 79-86. DOI:10.1046/j.1466-822X.2001.00267.x |
| [51] |
Yang Y H, Fang J Y, Ji C J, Han W X. Above- and belowground biomass allocation in Tibetan grasslands. Journal of Vegetation Science, 2009, 20(1): 177-184. DOI:10.1111/j.1654-1103.2009.05566.x |
| [52] |
Qiong L, Grytnes J A, Birks H J B. Alpine vegetation and species-richness patterns along two altitudinal gradients in the Gyama Valley, south-central Tibet, China. Plant Ecology & Diversity, 2010, 3(3): 235-247. |
| [53] |
Li W, Wen S J, Hu W X, Du G Z. Root-shoot competition interactions cause diversity loss after fertilization: a field experiment in an alpine meadow on the Tibetan Plateau. Journal of Plant Ecology, 2011, 4(3): 138-146. DOI:10.1093/jpe/rtq031 |
| [54] |
方精云, 神崎护, 王襄平, 依田恭二, 孙世洲, 下田胜久. 西藏珠峰-卓奥友峰普士拉地区高山稀疏植被的群落特征及小地形的影响. 生物多样性, 2004, 12(1): 190-199. |
| [55] |
Grace J B, Pugesek B H. A structural equation model of plant species richness and its application to a coastal wetland. The American Naturalist, 1997, 149(3): 436-460. DOI:10.1086/285999 |
| [56] |
王文颖, 周华坤, 杨莉, 李锦萍, 汪新川. 高寒藏嵩草(Kobresia tibetica)草甸植物对土壤氮素利用的多元化特征. 自然资源学报, 2014, 29(2): 249-255. |
| [57] |
卢慧, 丛静, 刘晓, 王秀磊, 唐军, 李迪强, 张于光. 三江源区高寒草甸植物多样性的海拔分布格局. 草业学报, 2015, 24(7): 197-204. |
| [58] |
张峰, 杨阳, 乔荠瑢, 贾丽欣, 赵天启, 赵萌莉. 利用方式对大针茅草原植物多样性、功能性状及地上生物量的影响. 中国草地学报, 2019, 41(1): 1-8. |
| [59] |
鄢燕, 张建国, 张锦华, 范建容, 李辉霞. 西藏那曲地区高寒草地地下生物量. 生态学报, 2005, 25(11): 2818-2823. |
| [60] |
Zhao Z G, Wang Y K. Selection by pollinators on floral traits in generalized Trollius ranunculoides (Ranunculaceae) along Altitudinal Gradients. PLoS One, 2015, 10(2): e0118299. DOI:10.1371/journal.pone.0118299 |
| [61] |
Friend A D, Woodward F I. Evolutionary and ecophysiological responses of mountain plants to the growing season environment. Advances in Ecological Research, 1990, 20: 59-124. |
| [62] |
郭丽珠, 黄顶, 张丛, 李佳欢, 赵欢, 王堃. 退化典型草原狼毒生物量分配及异速生长分析. 中国草地学报, 2019, 41(6): 53-59. |