 2022, Vol. 42
2022, Vol. 42文章信息
- 白燕娇, 李义学, 刘任涛, 常海涛, 唐希明
- BAI Yanjiao, LI Yixue, LIU Rentao, CHANG Haitao, TANG Ximing
- 干旱绿洲区枸杞林地面节肢动物功能群结构季节分布特征
- Seasonal distribution of functional groups of ground-active arthropods in Lycium. barbarum plantations in arid oasis region
- 生态学报. 2022, 42(15): 6239-6249
- Acta Ecologica Sinica. 2022, 42(15): 6239-6249
- http://dx.doi.org/10.5846/stxb202104271110
-
文章历史
- 收稿日期: 2021-04-27
- 网络出版日期: 2022-04-07
- 修订日期: 2022-04-07





2. 西北土地退化与生态恢复国家重点实验室培育基地, 银川 750021;
3. 西北退化生态系统恢复与重建教育部重点实验室, 银川 750021;
4. 中卫市自然资源局治沙林场, 中卫 755000
2. Breeding Base for Sate Key Laboratory of Land Degradation and Ecological Restoration in Northwestern China, Yinchuan 750021, China;
3. Key Laboratory for Restoration and Reconstruction of Degraded Ecosystems in Northwestern China of Ministry of Education, Ningxia University, Yinchuan 750021, China;
4. Sand-Fixing Forestry Station of Natural Resource Bureau in Zhongwei, Zhongwei 755000, China
在干旱区, 绿洲是政治、经济、文化等活动的重要场所, 绿洲的利用直接影响着一个地区的发展[1—2]。绿洲是干旱区背景下人类局部优化的生存环境, 是自然与人类矛盾最为激烈和复杂的一种生态环境[3—4]。在宁夏卫宁平原上, 中宁枸杞依赖绿洲而发展成具有特色的支柱产业, 成为宁夏重要的经济树种[5]。随着季节更替, 降雨和温度发生改变, 驱动枸杞林地资源分布变化, 直接关系到地面节肢动物群落的时空分布[6]。由于不同地面节肢动物随季节变化而在体形、活动能力及食性等方面各不相同, 从而表现出不同的功能群分布来适应环境变化[7]。在绿洲农业生态系统中, 利用地面节肢动物功能群可以反应生境梯度变化和生态系统的功能, 并且通过不同营养级之间强烈的上行效应和下行效应, 可以有效调控食物链的组成和结构[8—9]。因此, 深入研究枸杞林地地面节肢动物功能群的组成和季节分布特征, 对于宁夏绿洲生态系统的稳定以及可持续发展具有重要的理论与实践意义。
近些年来, 关于土壤动物功能类群的研究主要集中在环境因子、季节变化、土地管理方式不同产生的影响等方面。徐帅博等[10]对宝天曼自然保护区中小型土壤动物进行研究, 发现捕食性和腐食性土壤动物与土壤含水量呈正相关, 杂食性土壤动物与土壤有机质和全氮呈正相关, 植食性土壤动物与全氮呈正相关。叶岳等[11]对七星岩大型土壤动物进行研究, 发现大型土壤动物功能群的类群数和生物量与土壤温度呈负相关, 与pH值、有机质相关性不显著。陈蔚等[8]对宁夏荒漠草原进行研究, 发现地面节肢动物功能群多样性受季节变化影响深刻, 且功能群多样性与植物多样性间呈现出复杂的线性关系。并且, 不同管理方式会导致农田土壤空间格局改变将影响土壤动物功能类群的多样性和功能[12], Quinn等[13]发现施用N肥后植物根组织N的浓度增加可能会导致植食性土壤动物数量增加。但是, 目前对干旱区绿洲枸杞林的地面节肢动物功能类群结构和季节动态分布特征尚不清楚。因此, 研究枸杞林地地面节肢动物功能群对季节更替的响应规律, 有助于揭示枸杞林地土壤动物功能群的季节节律性和动态特征。
鉴于此, 本文以宁夏干旱区绿洲枸杞林作为研究样地, 分别于2018年春季、夏季和秋季, 调查地面节肢动物, 并结合土壤理化性质, 分析枸杞林地地面节肢动物功能类群的组成、分布特征以及与环境因子的相关关系。探讨不同地面节肢动物功能群的季节动态变化以及与枸杞林地土壤的相互影响, 旨在为干旱区绿洲农业的发展和绿洲生态系统的安全提供有价值的参考。
1 材料与方法 1.1 研究区概况研究区位于宁夏回族自治区中部西侧(36°49′—37°47′ N, 105°15′—106°05′ E, 海拔1348 m), 地处黄河两岸, 属北温带大陆性干旱气候。年平均降水量为202.1 mm, 降水多集中于6—8月, 占全年降水量的61%;年蒸发量1947.1 mm, 为年平均降水量的9.6倍。年平均气温为9.5℃, 日平均气温≥0℃, 积温3200—3300℃。该区域日照充足, 温差较大, 热量丰富, 无霜期较长。并且, 该区域处在清水河与黄河交汇的冲积平原上, 土壤矿物质含量丰富, 腐殖质多, 熟化度高;土壤为灌淤土淡灰钙, 沙质壤土, 土质均匀;境内的清水河与黄河交汇, 便于灌溉[14—15]。中宁县气候干燥, 四季分明, 昼夜温差大, 光照强烈, 为枸杞生长提供了优越自然环境。
本试验选取的枸杞林地是同一个农业合作社, 管理方式相同, 本底条件一致。4—6月, 枸杞处于现蕾期;5—7月, 枸杞处于开花期和幼果期;6—8月, 枸杞处于果熟期;8—9月, 枸杞处于秋蕾开花期;9—10月, 枸杞处于秋果果熟期[16]。在果熟期和秋果果熟期, 对枸杞林地施用有机肥。枸杞林为单行种植, 密度是220株/hm2, 规格:行距1.7 m, 株距1.1 m。本试验调查于2018年开展, 研究区域内年降雨量为234.2 mm, 平均气温为11.3℃。其中, 5月份降雨量为19.1 mm, 平均气温为20.1℃;7月份降雨量为113.2 mm, 平均气温为25.5℃;9月份降雨量为22.2 mm, 平均气温为16.6℃(图 1)。

|
| 图 1 中宁县2018年各月平均降雨量和气温 Fig. 1 Mean monthly rainfall and air temperature of Zhongning in 2018 |
在宁夏中宁县枸杞种植基地, 选择枸杞林为研究样地。为了覆盖枸杞林的整个生长过程, 全面反映枸杞林节肢动物的总体季节分布特征, 本研究样地包括不同林龄的枸杞林:1, 3, 5, 8, 10年。每个研究样地设置5个调查样区, 每个样区间距15—20 m, 面积分别为50 m×80 m, 共25个样本单位。每个样区内设置4个样点, 每个调查样点间距30 m以上。每个样点选择1株长势良好的枸杞灌丛进行标记, 在枸杞灌丛根部(避免田间管理对土壤动物的扰动而影响捕获的有效性)布设1个陷阱杯。共得到100个陷阱杯(5个样地×5个样区×4个样点×1陷阱杯/样点), 分别于2018年春季(5月)、夏季(7月)和秋季(9月)调查。
1.3 地面节肢动物取样与鉴定在每个调查样点附近, 采用陷阱诱捕法调查地面节肢动物功能群的组成与数量分布。具体方法为:在每个样点将收集器杯口齐地面(上、下直径分别为14 cm和7 cm, 高度10 cm)埋入土中, 在杯内加入无色无味的玻璃水和酒精混合液, 以增加诱捕的有效性。每次布设陷阱的持续时间均为14 d, 每3 d回收一次, 防止标本腐烂。在室内将收集的地面节肢动物标本保存于75%酒精中待鉴定。
将采集到的地面节肢动物根据《宁夏贺兰山昆虫》[17]、《昆虫分类》[18]和《中国土壤动物检索图鉴》[19]等参考书进行鉴定。然后, 将鉴定和统计后的地面节肢动物烘干并称重(精度为0.0001 g), 得到节肢动物生物量。本研究中, 将所有地面节肢动物标本鉴定到科水平, 并且, 由于地面节肢动物的成虫与幼虫在环境中有不同生态功能, 所以将成虫与幼虫个体数分别进行统计[20]。依据取食类型不同将收集到的地面节肢动物划分为植食性(phytophagous, Ph)、捕食性(predatory, Pr)、杂食性(omnivorous, Om)和腐食性(saprophagous, Sa)4个营养功能群[8, 21]。
1.4 土壤样品采集与分析在春季、夏季和秋季的每个调查样点, 利用铝盒采集剖面土壤测定土壤含水量(0—15 cm)。然后, 用5点取样法在枸杞根部调查样点附近采集0—15 cm的表层混合土样1个, 装入自封袋, 带回实验室用于土壤理化性质分析, 共取土样75个(5个调查样点×5个重复样区×3个季节)。在实验室, 将采集到的土壤样品在自然状态下进行风干后, 过2 mm土壤筛, 去除其中的杂质(如根、叶、石块等), 然后用于测定土壤pH、土壤电导率、土壤全碳、土壤全氮、土壤全磷、土壤全钾、土壤硒等理化性质。
土壤含水量(%)采用烘干称重法测定。土壤pH和土壤电导率(μS/m)均采用1:5土水比浸提后, 分别使用PHS—3C酸度计与便携式电导率仪(雷磁DDSJ—308F)进行测定。土壤全碳(g/kg)和土壤全氮(g/kg)采用元素分析仪(意大利DK6, UDK140)进行测定, 土壤全磷(g/kg)采用Na2CO3熔融法测定, 土壤全钾(g/kg)采用NaOH熔融法测定, 土壤硒含量(g/kg)采用原子荧光光谱法测定[22]。
1.5 数据处理(1) 群落多样性
分别将春、夏、秋季每个样地采样点数据进行整理, 统计地面节肢动物每个功能群的个体数(只/陷阱)、类群数和生物量, 在此基础上进行每个功能群多样性指标计算[23]。
(2) 功能多样性
对于功能多样性, 目前主要从功能丰富度、功能均匀度和功能离散度3方面来描述。本文选择功能丰富度(FRic)、Raos二次熵指数(RaoQ)、功能离散度(FDis)和功能优势值(CWM)来评价功能多样性[24]。对于功能多样性的计算, 本研究选择食性来代表动物功能特性。功能参数计算方法见表 1。
| 名称Name | R代码R code | 公式Formula | 说明Note |
| 功能多样性 Rao′s quadratic entropy |
dbFD (X, comm) $RaoQ |
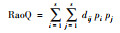 |
式中, dij为物种i和j的功能距离特征, pi为物种i的多度, s为研究的性状总数。 |
| 功能离散度 Functional dispersion |
dbFD (X, comm) $FDis |
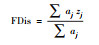 |
式中, aj为物种j的多度, zj为物种j到加权质心的距离。 |
| 功能优势值 Community-level weighted means |
Functcomp (X, comm) $CWM |
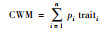 |
式中, pi是物种i的多度, traiti为物种i的性状值, n为物种个数 |
本试验所有数据均采用IBM SPSS Statistics 25.0进行统计分析, 通过单因素方差分析(One-way ANOVA)和最小显著差异法(LSD)比较不同处理间的差异性, 利用方差齐性检验和非参数检验统计是否在P=0.05水平上差异显著。多样性计算使用的软件为R, 其中FD软件包完成功能多样性指数计算。采用Pearson法对地面节肢动物群落指数与环境因子进行相关分析, 并进行相关系数检验(P < 0.05)。应用Origin 2018进行作图分析。
(3) 冗余分析
通过对3个季节地面节肢动物功能群分布进行除趋势对应分析(Detrended correspondence analysis, DCA), 计算出排序轴的梯度长度(Lengths of gradient, LGA), LGA=2.82<4, 故采用线性模型的冗余对应分析(Redundancy analysis, RDA)来确定地面节肢动物功能群分布与土壤因子间的关系。本文对地面节肢动物功能群与环境因子进行RDA排序分析, 得到功能群分布与环境因子的二维排序图。同时, 采用偏RDA分析(Partial RDA)和蒙特卡洛置换检验(Monte Carlo permutation test), 定量评价每个因子对地面节肢动物功能群分布变化的贡献率(即独立解释量)。在偏RDA分析的基础上, 选择对地面节肢动物功能群分布有较高影响的解释变量, 绘制地面节肢动物功能群分布与解释变量关系的二维排序图(Biplot)。应用排序软件CANOCO 5进行分析运算。
2 结果与分析 2.1 土壤环境特征由表 2可知, 在3个季节间环境因子均存在显著差异。土壤全碳在夏季最高, 秋季次之, 春季最低(P < 0.05)。土壤全氮和土壤pH均表现为夏季显著低于春季和秋季(P < 0.05), 而后两个季节间无显著差异。土壤全磷表现为秋季显著低于春季和夏季(P < 0.05), 而后两个季节间无显著差异。土壤全钾表现为夏季显著高于春季(P < 0.05), 而两者均与秋季无显著差异。土壤硒表现为春季显著低于夏季和秋季(P < 0.05), 而后两个季节间无显著差异。土壤含水量表现为秋季显著高于春季和夏季(P < 0.05), 而后两个季节间无显著差异。土壤温度表现为夏季显著高于春季和秋季(P < 0.05), 而后两个季节间无显著差异。土壤电导率表现为春季显著高于夏季和秋季(P < 0.05), 而后两个季节间无显著差异。
| 季节 Season |
全碳 Soil total carbon/(g/kg) |
全氮 Soil total nitrogen/(g/kg) |
全磷 Soil total phosphorus/(g/kg) |
全钾 Soil total potassium/(g/kg) |
硒 Soil selenium/(g/kg) |
含水量 Soil moisture/% |
温度 Soil temperature/℃ |
土壤pH Soil pH |
电导率 Electrical conductivity/(μS/m) |
| 春Spring | 22.54±0.54c | 0.7±0.09a | 1.87±0.21a | 1.95±0.03b | 0.11±0.02b | 0.16±0.01b | 19.76±0.29b | 7.6±0.02a | 558.43±91.02a |
| 夏Summer | 30.73±0.74a | 0.36±0.06b | 1.63±0.19a | 2.13±0.05a | 0.24±0.01a | 0.16±0.01b | 29.18±0.42a | 7.26±0.08b | 284.27±24.94b |
| 秋Autumn | 26.28±0.92b | 0.66±0.09a | 0.97±0.09b | 2.06±0.05ab | 0.26±0.01a | 0.27±0.01a | 19.72±0.28b | 7.5±0.07a | 194.25±12.91b |
| 同列不同字母表示显著差异(P < 0.05) | |||||||||
由表 3可知, 在3个季节内, 共捕获634只地面节肢动物, 隶属于5纲9目26个类群, 划分为4种功能群。其中, 植食性动物7个类群, 捕食性动物9个类群, 腐食性动物3个类群, 杂食性动物7个类群。
| 类群 Group |
功能群 Functional group |
春 Spring |
夏 Summer |
秋 Autumn |
类群 Group |
功能群 Functional group |
春 Spring |
夏 Summer |
秋 Autumn |
|
| 狼蛛科Lycosidae | Pr | 4(1.65) | 8(5.23) | 7(2.94) | 蝽科Pentatomidae | Ph | 1(0.65) | |||
| 平腹蛛科Gnaphosidae | Pr | 10(4.12) | 1(0.65) | 8(3.36) | 步甲科Carabidae | Pr | 157(64.61) | 123(80.39) | 207(86.97) | |
| 光盔蛛科Liocranidae | Pr | 1(0.65) | 象甲科Curculionidae | Ph | 15(6.17) | |||||
| 蟹蛛科Thomisidae | Pr | 1(0.41) | 隐食甲科Cryptophagidae | Om | 5(2.06) | |||||
| 跳蛛科Salticidae | Pr | 5(2.06) | 拟步甲科Tenebrionidae | Ph | 9(3.7) | 1(0.65) | ||||
| 近管蛛科Anyphaonidae | Pr | 1(0.65) | 粪金龟科Geotrupidae | Sa | 8(3.29) | 1(0.65) | 1(0.42) | |||
| 盲蛛科Phalangidae | Om | 1(0.42) | 隐翅虫科Staphylinidae | Pr | 14(5.76) | 1(0.65) | 1(0.42) | |||
| 蜱螨目Arachnoidea | Om | 1(0.41) | 3(1.26) | 金龟甲科Scarabacidae | Sa | 1(0.41) | 1(0.65) | |||
| 潮虫科Oniscidae | Om | 2(0.82) | 2(1.31) | 6(2.52) | 阎甲科Histeridae | Om | 1(0.41) | |||
| 马陆科Julidae | Ph | 1(0.65) | 锹甲科Lucanidae | Om | 2(0.82) | |||||
| 蜈蚣目幼虫Scolopendra larva | Pr | 1(0.65) | 薪甲科Lucanidae | Sa | 1(0.65) | |||||
| 红蝽科Pyrrhocoridae | Ph | 1(0.65) | 鞘翅目幼虫Coleoptera larvae | Ph | 4(2.61) | 2(0.84) | ||||
| 盲蝽科Miridae | Ph | 2(0.82) | 4(2.61) | 蚁科Formicidae | Om | 6(2.47) | 2(0.84) | |||
| Ph:植食性phytophagous;Pr:捕食性predatory;Sa:腐食性sacrificial;Om:杂食性omnivorous; 括弧内数据表示个体数占总个体数的百分比(%) | ||||||||||
春季, 植食性类群有盲蝽科(Miridae)、象甲科(Curculionidae)和拟步甲科(Tenebrionidae)3类, 个体数占10.70%;捕食性类群有狼蛛科(Lycosidae)、平腹蛛科(Gnaphosidae)、蟹蛛科(Thomisidae)、跳蛛科(Salticidae)、步甲科(Carabidae)和隐翅虫科(Staphylinidae)6类, 个体数占78.60%;腐食性类群有粪金龟科(Geotrupidae)和金龟甲科(Scarabacidae)2类, 个体数占3.70%;杂食性类群有蜱螨目(Arachnoidea)、潮虫科(Oniscidae)、隐食甲科(Cryptophagidae)、阎甲科(Histeridae)、锹甲科(Lucanidae)和蚁科(Formicidae)6类, 个体数占7.00%。
从春季到夏季, 植食性类群除了盲蝽科(Miridae)和拟步甲科(Tenebrionidae)2类外, 另外增加了马陆科(Julidae)、红蝽科(Pyrrhocoridae)、蝽科(Pentatomidae)和鞘翅目幼虫(Coleoptera larvae)4类, 个体数占7.84%;捕食性类群除了狼蛛科(Lycosidae)、平腹蛛科(Gnaphosidae)、步甲科(Carabidae)和隐翅虫科(Staphylinidae)4类外, 另外增加了光盔蛛科(Liocranidae)、近管蛛科(Anyphaonidae)和蜈蚣目幼虫(Scolopendra larva)3类, 个体数占88.89%;腐食性类群增加了1类, 为薪甲科(Lathridiidae), 个体数占1.96%;杂食性类群减少至1类, 为潮虫科(Oniscidae), 个体数占1.31%。
夏季之后, 植食性类群和腐食性类群均有且仅有1类, 分别为鞘翅目幼虫(Coleoptera larvae)和粪金龟科(Geotrupidae), 个体数分别占0.84%和0.42%;捕食性类群减少至4类, 分别为狼蛛科(Lycosidae)、平腹蛛科(Gnaphosidae)、步甲科(Carabidae)和隐翅虫科(Staphylinidae), 个体数占93.70%;杂食性类群增加了3类, 分别为盲蛛科(Phalangidae)、蜱螨目(Arachnoidea)和蚁科(Formicidae), 个体数占5.04%。
2.3 地面节肢动物功能群结构特征图 2可知, 植食性地面节肢动物个体数和生物量表现为春季显著高于秋季(P < 0.05), 而两者均与夏季无显著差异;类群数表现为夏季显著高于秋季(P < 0.05), 而两者均与春季无显著差异。捕食性地面节肢动物生物量表现为夏季和秋季显著高于春季(P < 0.05), 而前两个季节间无显著差异;个体数和类群数在不同季节间均无显著差异(P>0.05)。腐食性地面节肢动物个体数、生物量和类群数在不同季节间均无显著差异(P>0.05)。杂食性地面节肢动物个体数和类群数表现为春季显著高于夏季(P < 0.05), 而两者均与秋季无显著差异;生物量在不同季节间均无显著差异(P>0.05)。

|
| 图 2 地面节肢动物功能群多样性指数(均值±标准误, n=75) Fig. 2 Functional groups index of ground-active arthropods(Mean ea SE, n=75) |
由图 3可知, 功能丰富度指数和功能优势值均表现为春季显著高于夏季(P < 0.05), 而两者均与秋季无显著差异。Rao′s二次熵指数和功能离散度指数在不同季节间均无显著差异(P>0.05)。

|
| 图 3 地面节肢动物功能多样性分析(均值±标准误, n=75) Fig. 3 Analysis of functional diversity of ground-active arthropods(Mean ea SE, n=75) |
由图 4可知, 第1典型轴(F=13.453, P=0.036)和所有典型轴(F=2.218, P=0.014)在统计学上均达到了显著水平, 说明排序轴能够很好地反映枸杞林地地面节肢动物功能群分布与土壤因子的关系。并且, 前两个排序轴累计解释了15.07%的地面节肢动物功能群组成变异。

|
| 图 4 地面节肢动物功能群分布与土壤因子关系的RDA二维排序图 Fig. 4 RDA two-dimensional diagram of the relationship between functional groups of ground-active arthropods and environmental factors SM:土壤含水量Soil moisture;EC:电导率Electrical conductivity;AT:土壤温度Soil temperature;TP:土壤全磷Soil total phosphorus;TK:土壤全钾Soil total potassium;Se:土壤硒Soil selenium;pH:Soil pH;TC:土壤全碳Soil total carbon;TN:土壤全氮Soil total nitrogen |
由图 4和表 4可以得知, 植食性地面节肢动物与排序轴1呈负相关关系。其中, 植食性地面节肢动物分布与土壤含水量呈显著负相关关系(P < 0.01)。捕食性和腐食性地面节肢动物与排序轴1呈正相关关系。其中, 捕食性地面节肢动物分布与土壤含水量呈正相关关系(P < 0.05), 腐食性地面节肢动物分布与土壤全磷含量呈显著正相关关系(P < 0.01)。杂食性地面节肢动物与排序轴2呈负相关关系。其中, 杂食性地面节肢动物分布与土壤温度和全碳呈负相关关系(P < 0.05), 与土壤全氮呈正相关关系(P < 0.05)。
| 土壤含水量 Soil moisture/% |
温度 Soil temperature/℃ |
全碳 Soil total carbon/(g/kg) |
全氮 Soil total nitrogen/(g/kg) |
全磷 Soil total phosphorus/(g/kg) |
全钾 Soil total potassium/(g/kg) |
硒 Soil selenium/(g/kg) |
土壤pH Soil pH |
电导率 Electrical conductivity/(μS/m) |
|
| Ph | -0.308** | 0.003 | -0.158 | -0.123 | -0.123 | -0.086 | -0.062 | 0.135 | -0.063 |
| Pr | 0.250* | -0.113 | 0.063 | 0.190 | 0.147 | -0.067 | 0.075 | 0.199 | -0.088 |
| Sa | 0.120 | -0.012 | 0.071 | 0.093 | 0.426** | -0.01 | -0.208 | 0.092 | 0.145 |
| Om | -0.012 | -0.333** | -0.235* | 0.242* | -0.215 | -0.047 | -0.031 | 0.133 | 0.105 |
| *P < 0.05;**P < 0.01 | |||||||||
偏RDA结果分析表明(表 5), 土壤含水量对地面节肢动物分布产生显著影响, 对地面节肢动物个体数的贡献率是6.2%(P=0.032), 其余环境因子对地面节肢动物个体数影响不显著。因此, 土壤含水量是影响枸杞灌丛地面节肢动物功能群的主要环境因子。
| 变量 Variable |
贡献率 Contribution rate/% |
F | P | 变量 Variable |
贡献率 Contribution rate/% |
F | P | |
| SM | 6.2 | 4.8 | 0.032 | TN | 0.6 | 0.5 | 0.504 | |
| pH | 3.1 | 2.5 | 0.098 | EC | 0.5 | 0.4 | 0.556 | |
| TP | 1.7 | 1.3 | 0.228 | TC | 0.3 | 0.2 | 0.682 | |
| TK | 1.4 | 1.1 | 0.234 | AT | < 0.1 | < 0.1 | 0.850 | |
| Se | 1.4 | 1.1 | 0.306 | |||||
| SM:土壤含水量Soil moisture;EC:电导率Electrical conductivity;AT:土壤温度Soil temperature;TP:土壤全磷Soil total phosphorus;TK:土壤全钾Soil total potassium;Se:土壤硒Soil selenium;pH:Soil pH;TC:土壤全碳Soil total carbon;TN:土壤全氮Soil total nitrogen | ||||||||
在干旱区绿洲农业生态系统中, 气候变化和人类活动往往对该生态系统内的土壤理化性质及养分元素等环境条件产生深刻的影响[25]。夏季为枸杞的生长季, 为了保证果树健康生长, 保障果实良好的品质, 因此需要施用大量的有机肥, 这会显著增加土壤中全碳、全钾以及硒元素的含量[26]。相关研究表明, 土壤有机质对硒元素具有吸附与固定的作用, 土壤有机质含量越高, 硒元素的吸附能力越强, 总含量也越高[27]。本研究中, 土壤全氮在夏季最低, 而土壤全磷在秋季最低。这与枸杞的生长发育有关, 土壤全氮和全磷随果树生长周期发生季节性变化, 并且与施肥管理有一定的关系[28—29]。枸杞林土壤含水量受到降雨、温度和灌溉的影响[30]。春季降雨少, 温度较低, 夏季降雨多, 温度较高, 蒸发量大, 导致春季和夏季土壤含水量较低。夏季土壤温度高于其它两季。这与研究区域的气候有关, 夏季温度较高导致土壤温度也较高。夏季土壤pH显著低于春季和秋季。这与常海涛等[23]的研究结果相同。说明在枸杞生长季节, 植被发育过程中根系分泌的有机酸及其与微生物的相互作用降低了表层土壤的pH值[31]。春季土壤电导率显著高于夏季和秋季。这可能是由于春季降雨量较少, 蒸发垂直向上, 导致盐分在土壤表层积累, 而到了夏季与秋季, 随着降雨量和灌溉次数的增加, 土壤盐分随水分下渗, 使电导率降低[32]。
3.2 季节变化对地面节肢动物功能群组成与结构的影响在干旱区绿洲农业生态系统中, 土壤理化性质在随季节变化的同时, 地面节肢动物功能群结构分布特征也随之发生显著变化。植食性地面节肢动物的个体数和生物量均表现为春季>夏季>秋季。夏季到秋季为枸杞的采摘期, 为了保证果实产量和质量, 果农在此期间喷施了农药, 导致植食性动物数量减少[33]。从春季到夏季, 植食性动物类群增加了马陆科、红蝽科、蝽科和鞘翅目幼虫4类, 但个体数极少。这是由于夏季温度和降水充足, 食物资源丰富, 良好的生境条件为植食性类群的生存提供了保障[34]。
捕食性地面节肢动物共占总捕获数的86.75%, 是枸杞林地生境的主要功能类群。这与刘继亮和李锋瑞[35]的研究结果相似, 即高强度的农业管理措施对地面节肢动物产生较大的影响[36]。在枸杞林地内捕食性动物主要以步甲科和游猎型蜘蛛为主, 迁移能力强, 活动范围广, 受人为干扰影响较小[37—38]。捕食性动物的个体数在不同季节间无显著差异, 但生物量表现为夏季和秋季高于春季。步甲科在捕食性动物中所占比例较大, 主要活动在夏季和秋季, 所以捕食性动物的生物量会显著增加[37]。并且, 植食性动物随季节变化生物量减少, 而捕食性动物生物量增加, 这形成了一种自下而上的上行控制效应[39]。捕食性动物类群主要由蜘蛛目昆虫和鞘翅目昆虫组成, 很少出现新的类群, 表明枸杞林地的生态系统较为稳定[40]。
腐食性地面节肢动物所占比例最少, 这与刘长海[41]等人的研究结果相似。腐食性动物的主要食物来源是枯枝落叶和动物残体, 但由于枸杞林地常进行除草和翻耕管理, 地面植被较少[16], 导致腐食性动物的数量和类群较少, 仅有粪金龟科、金龟甲科和薪甲科3类。这与林琳等[42]的研究结果一致, 即在环境压力和人为干扰程度较大的枸杞林地内腐食性类群的生存和发育受到抑制。腐食性动物个体数、生物量和类群数在不同季节间均无显著差异。可能是受不同季节凋落物分解矿化过程影响, 调查取样时间不在凋落物分解较多的时期, 影响腐食性动物的生存及变化[43]。
杂食性地面节肢动物的个体数和类群数表现为春季显著高于夏季。一方面, 杂食性类群与土壤温度呈显著负相关(表 3), 表明杂食性动物对土壤温度具有很强的敏感性, 较高的土壤温度会抑制杂食性类群的繁殖[44]。另一方面, 在食物资源有限的环境中, 杂食性动物与植食性和捕食性动物竞争有限的资源所致[45]。杂食性动物的生物量在不同季节间均无显著差异。这可能是因为杂食性动物具有较宽的生态位, 即使是在资源较为匮乏的春季和秋季, 杂食性类群也能够维持相对稳定的生物量[46]。
3.3 季节变化对地面节肢动物功能多样性的影响功能多样性指数用于表征生态系统的服务功能, 反映生态系统的生产力和抗干扰能力[47]。本研究中, 功能丰富度表现为春季>秋季>夏季。说明春季地面节肢动物在群落中所占据的功能空间较大, 而夏季较小。其原因在于春季资源开始丰富, 竞争释放导致地面节肢动物群落功能特性多样化, 有利于物种以不同方式对有限的资源进行最大利于[48]。而夏季和秋季, 枸杞林中不仅存在高强度管理, 而且采摘导致地面被频繁践踏, 导致功能丰富度降低。从Rao′s二次熵指数和功能离散度指数来看, 枸杞林地中不同季节间均无显著差异, 可能是由于在人为管理的枸杞林中, 环境压力较大, 地面节肢动物群落组成的物种性状将被限制在一定的范围内, 从而导致Rao′s二次熵指数和功能离散度指数在不同季节间无显著差异[49]。功能优势值均表现为春季显著高于夏季, 说明优势物种在春季产生更大影响[43]。综合分析表明, 在人为管理的枸杞林生态系统中, 环境压力较大, 但功能较稳定。
3.4 地面节肢动物功能群分布与土壤因子的关系在绿洲生态系统中, 环境因子对地面节肢动物功能群的分布会产生不同的影响。地面节肢动物功能群的分布与土壤含水量、土壤温度、土壤全碳、土壤全氮和土壤全磷含量的相关性较大。说明地面节肢动物功能群的分布与环境因子间关系密切, 即地面节肢动物的功能群分布会受到土壤因子的相互作用的影响[9]。RDA分析结果表明, 土壤含水量是影响地面节肢动物功能群数量分布的主要因素。这与张安宁[50]的研究结果有相似之处。地面节肢动物功能类群对土壤含水量响应程度极高, 土壤动物的各种生命过程均离不开水, 同时决定了动物的部分生存环境[10, 51], 因此土壤含水量也就成为土壤动物的重要限制因子, 可以影响地面节肢动物功能群的数量分布。
本研究中, 相关环境因子对地面节肢动物功能群分布的总贡献率为15.3%, 这说明在农田生态系统中, 土壤理化性质的影响较小, 更多的是人为活动的影响, 如施肥、耕地、喷施农药等。有研究表明, 大量配施化肥对某些土壤动物有抑制作用, 降低了土壤动物的种类, 显著减少了土壤中小型节肢动物的均匀性[52]。同时, 低浓度杀虫剂使中小型土壤动物个体数和类群数减少[33]。
4 结论综合分析表明:(1)随着季节变化, 枸杞林地土壤养分和土壤理化性质均具有显著差异。(2)枸杞林地中, 捕食性地面节肢动物是功能群组成的主要营养类群。(3)随着季节变化, 地面节肢动物功能多样性较稳定。(4)地面节肢动物功能群与土壤含水量、土壤温度、土壤全碳、土壤全氮和土壤全磷含量的关系密切。并且, 土壤含水量是影响绿洲地面节肢动物功能群分布的主要限制因子。
| [1] |
雷志栋, 杨汉波, 倪广恒, 杨诗秀, 丛振涛, 李平, 张玉平, 朱刚, 王教堂, 陈俊鹏. 干旱区绿洲耗水分析. 水利水电技术, 2006, 37(1): 15-20. DOI:10.3969/j.issn.1000-0860.2006.01.004 |
| [2] |
师庆东, 郭玉川, 周小龙, 戴岳, 依力亚斯江·努尔麦麦提, 李诚志. 塔克拉玛干沙漠克里雅河尾闾达理雅博依绿洲地表水、地下水对植被格局的影响机制. 新疆大学学报: 自然科学版, 2019, 36(3): 253-259, 286-286. |
| [3] |
贾宝全. 绿洲景观若干理论问题的探讨. 干旱区地理, 1996, 19(3): 58-65. |
| [4] |
杜梦迪, 李兰, 罗光明, 董克鹏, 师庆东. 气候与土地利用变化对于田绿洲农业耗水特征的影响. 水土保持通报, 2020, 40(3): 103-109. |
| [5] |
张德纯. 宁夏中宁枸杞. 中国蔬菜, 2021(1): 65-65. |
| [6] |
叶岳, 姜玉霞. 黑石顶自然保护区土壤动物功能类群对季节动态的响应. 广东农业科学, 2017, 44(5): 78-85. |
| [7] |
李萌. 贡嘎山东坡土壤动物群落结构及多样性时空动态[D]. 成都: 西南民族大学, 2014.
|
| [8] |
陈蔚, 黄兴科, 刘任涛, 张安宁, 常海涛. 宁夏荒漠草原植物多样性对地面节肢动物功能群多样性的影响. 草地学报, 2019, 27(6): 1587-1595. |
| [9] |
Gaudêncio M J, Cabral H N. Trophic structure of macrobenthos in the Tagus estuary and adjacent coastal shelf. Hydrobiologia, 2007, 587(1): 241-251. DOI:10.1007/s10750-007-0686-6 |
| [10] |
徐帅博, 李艳红, 冯小燕, 李理, 朱连奇. 宝天曼中小型土壤动物功能类群及其影响因素研究. 南阳师范学院学报, 2021, 20(1): 21-30. DOI:10.3969/j.issn.1671-6132.2021.01.004 |
| [11] |
叶岳, 姜玉霞, 陈华. 大型土壤动物功能类群对小生境环境因子的响应. 江苏农业科学, 2019, 47(3): 253-257. |
| [12] |
刘鹏飞, 红梅, 平翠枝, 美丽, 王文东, 赵乌英嘎. 玉米间作种植对黑土区农田中小型土壤动物群落的影响. 中国土壤与肥料, 2018(6): 91-97. |
| [13] |
Quinn M A, Hower A A. Effects of root nodules and taproots on survival and abundance of Sitona hispidulus (Coleoptera: Curculionidae) on Medicago sativa. Ecological Entomology, 1986, 11(4): 391-400. DOI:10.1111/j.1365-2311.1986.tb00318.x |
| [14] |
王秀娟, 梁勇, 王银, 闵庆文, 张永勋. 宁夏中宁枸杞种植系统申报国家级重要农业文化遗产的可行性研究. 现代化农业, 2015(12): 30-33. DOI:10.3969/j.issn.1001-0254.2015.12.015 |
| [15] |
朱建国, 何月红, 谢施祎. 宁夏中宁县枸杞产业发展的优势分析. 北京农业, 2015(6): 239-239, 240-240. DOI:10.3969/j.issn.1000-6966.2015.06.187 |
| [16] |
李丁仁, 李爽, 曹弘哲. 宁夏枸杞. 银川: 宁夏人民出版社, 2012: 70-72.
|
| [17] |
王新谱, 杨贵军. 宁夏贺兰山昆虫. 银川: 宁夏人民出版社, 2010: 156-342.
|
| [18] |
郑乐怡, 归鸿. 昆虫分类上. 南京: 南京师范大学出版社, 1999: 525-882.
|
| [19] |
尹文英. 中国土壤动物检索图鉴. 北京: 科学出版社, 2000: 299-606.
|
| [20] |
Swift M J, Heal O W, Anderson J M. Decomposition in terrestrial ecosystems. Studies in Ecology, 1979, 5(14): 2772-2774. |
| [21] |
刘继亮, 赵文智, 李锋瑞. 黑河中游荒漠地面节肢动物分布特征. 干旱区研究, 2015, 32(6): 1192-1200. |
| [22] |
鲍士旦. 土壤农化分析(第三版). 北京: 中国农业出版社, 2000: 39-114.
|
| [23] |
常海涛, 刘佳楠, 赵娟, 张静, 罗雅曦, 刘任涛, 左小安. 乌拉特荒漠草原红砂(Reaumuria soongorica)灌丛林地地面节肢动物群落季节分布特征. 生态学报, 2020, 40(9): 3121-3130. |
| [24] |
蒋万祥, 渠晓东, 吴乃成, 贾兴焕, 蔡庆华. 神农架地区香溪河流域水生昆虫物种多样性与功能多样性研究. 长江流域资源与环境, 2021, 30(6): 1428-1436. |
| [25] |
苏芝屯, 黄维, 乔斌, 何彤慧, 冯艳琼, 陈向全, 崔乔. 不同开发程度对沙坡头南山台子绿洲土壤性质的影响. 土壤通报, 2019, 50(5): 1045-1052. |
| [26] |
王庆鲁, 祝鹏飞, 丁凡. 地膜覆盖和有机肥施用对农田土壤和作物C、N、P化学计量学的影响. 生态学杂志, 2020, 39(4): 1191-1197. |
| [27] |
伍艳, 王玮屏, 蔡怀森, 杨忠芳. 黄河下游滩区原阳段硒元素分布特征及影响因素. 人民黄河: 1-7. [2021-09-02]. http://www.cnki.com.cn/Article/CJFDTotal-RMHH20190307000.htm.
|
| [28] |
孙娇, 任福聪, 陈刚, 郭鑫年, 梁锦秀, 周涛. 宁杞7号周年干物质与氮磷钾累积动态特征. 西北农林科技大学学报: 自然科学版, 2021, 49(5): 130-137. |
| [29] |
肖蓉, 白军红, 高海峰, 邓伟, 崔保山. 封闭性和开放性沼泽湿地土壤全磷的季节变化特征. 草业学报, 2010, 19(3): 88-93. |
| [30] |
李锋, 刘晓彤, 罗健航, 赵营, 王海廷, 张学军. 环境因子远程监测技术对枸杞园土壤水分动态变化规律的影响. 现代农业科技, 2020(2): 61-63. |
| [31] |
常海涛, 刘任涛, 刘佳楠, 赵娟, 罗雅曦, 张静, 马继. 草方格造林固沙过程中土壤性质变化及分形特征——以腾格里沙漠东南缘为例. 水土保持学报, 2018, 32(6): 58-65, 165-165. |
| [32] |
韩跃, 马风云, 解国磊, 秦光华, 马胜国. 黄河三角洲盐碱地混交林土壤电导率的空间异质性. 中国水土保持科学, 2014, 12(5): 84-89. DOI:10.3969/j.issn.1672-3007.2014.05.014 |
| [33] |
张淑花, 周利军, 郑丽颖, 彭万臣. 低浓度杀虫剂对农田中小型土壤动物群落结构的影响. 绥化学院学报, 2015, 35(2): 142-145. DOI:10.3969/j.issn.2095-0438.2015.02.039 |
| [34] |
刘任涛, 郗伟华, 朱凡. 宁夏荒漠草原地面节肢动物群落组成及季节动态特征. 草业学报, 2016, 25(6): 126-135. |
| [35] |
刘继亮, 李锋瑞. 干旱区绿洲扩张方式对土壤生物优势类群及功能的影响. 生物多样性, 2018, 26(10): 1116-1126. DOI:10.17520/biods.2018130 |
| [36] |
刘洁. 东北黑土区农田地表大型节肢动物多样性格局研究[D]. 长春: 吉林大学, 2019.
|
| [37] |
高九思, 赵增渠, 来学仁. 河南省苹果园鞘翅目天敌昆虫资源发生种类记述——步甲科Carabdae. 河南农业科学, 2007(9): 61-63. DOI:10.3969/j.issn.1004-3268.2007.09.020 |
| [38] |
胡文浩, 段美春, 那书豪, 张锋, 宇振荣. 坝上地区农田及两种恢复生境中蜘蛛多样性与群落特征. 应用生态学报, 2020, 31(2): 643-650. |
| [39] |
Hunter M D, Price P W. Playing chutes and ladders: heterogeneity and the relative roles of bottom-up and top-down forces in natural communities. Ecology, 1992, 73(3): 724-732. |
| [40] |
樊云龙, 马福军, 阮英慧, 宮连虎, 陶飞. 贵阳乌当盆地农田土壤动物群落特征研究. 贵州师范学院学报, 2021, 37(6): 1-6. DOI:10.3969/j.issn.1674-7798.2021.06.001 |
| [41] |
刘长海, 刘世鹏, 王晓涧, 陈宗礼, 廉振民. 陕北枣林土壤动物功能类群及其害虫生态调控. 土壤通报, 2011, 42(2): 316-319. |
| [42] |
林琳, 邬天媛, 李景科, 张雪萍. 大庆草甸草原区大型土壤动物功能类群. 地理研究, 2013, 32(1): 41-54. |
| [43] |
张宇晨, 红梅, 赵巴音那木拉, 叶贺, 闫瑾, 李静, 梁志伟. 荒漠草原土壤线虫对氮沉降及降雨变化的响应. 中国环境科学, 2021, 41(6): 2788-2797. DOI:10.3969/j.issn.1000-6923.2021.06.033 |
| [44] |
郭玉梅, 殷秀琴, 马辰. 长白山地不同地貌类型农田生态系统土壤动物群落特征及季节动态. 应用与环境生物学报, 2016, 22(6): 972-977. |
| [45] |
Lindberg N. Soil Fauna and Global Change: Responses to Experimental Drought, Irrigation, Fertilisation and Soil Warming[D]. Uppsala: Swedish University of Agricultural Sciences, 2003: 37-37.
|
| [46] |
Rosenheim J A, Corbett A. Omnivory and the indeterminacy of predator function: can a knowledge of foraging behavior help?. Ecology, 2003, 84(10): 2538-2548. DOI:10.1890/02-0469 |
| [47] |
杨晓明, 韩雪梅, 梁子安, 龚子乐, 朱会会, 李玉英. 人类活动干扰下大型底栖动物功能多样性评价. 河南师范大学学报: 自然科学版, 2020, 48(4): 96-102. |
| [48] |
韩涛涛, 唐玄, 任海, 王俊, 刘楠, 郭勤峰. 群落/生态系统功能多样性研究方法及展望. 生态学报, 2021, 41(8): 3286-3295. |
| [49] |
李亚芳, 杜飞雁, 王亮根, 谷阳光, 宁加佳. 基于生物性状分析方法的不同恢复阶段无瓣海桑人工林湿地大型底栖动物生态功能研究. 南方水产科学, 2018, 14(3): 10-19. |
| [50] |
张安宁, 常海涛, 陈蔚, 刘任涛. 干旱风沙区灌丛林地地面节肢动物群落对放牧管理的响应. 应用生态学报, 2019, 30(11): 3931-3941. |
| [51] |
董炜华, 王子婷, 李锋瑞, 刘继亮. 黑河中游绿洲化过程中不同开垦年限农田大型土壤动物群落特征. 地理科学, 2014, 34(10): 1254-1261. |
| [52] |
孔云, 张婷, 李刚, 杨殿林, 赵建宁, 张贵龙, 王丽丽, 修伟明. 不同施肥方式下玉米田土壤中小型节肢动物的群落特征及稳定性. 玉米科学, 2020, 28(2): 156-162. |