 2022, Vol. 42
2022, Vol. 42文章信息
- 唐国, 胡雷, 宋小艳, 李香真, 王长庭
- TANG Guo, HU Lei, SONG Xiaoyan, LI Xiangzhen, WANG Changting
- 高寒草甸植物群落不同根序根系特征对降雨量变化的响应
- Response of plant roots in different diameter classes to changing precipitation in an alpine meadow
- 生态学报. 2022, 42(15): 6250-6264
- Acta Ecologica Sinica. 2022, 42(15): 6250-6264
- http://dx.doi.org/10.5846/stxb202104261096
-
文章历史
- 收稿日期: 2021-04-26
- 网络出版日期: 2022-03-03





2. 中国科学院成都生物研究所, 环境与应用微生物重点实验室, 环境微生物四川省重点实验室, 成都 610041
2. Key Laboratory of Environmental and Applied Microbiology, Sichuan Key Laboratory of Environmental Microbiology, Chengdu Institute of Biology, Chinese Academy of Sciences, Chengdu 610041, China
草地作为陆地生态系统中分布最广的类型之一, 约占地球陆地总面积的25%, 在调节区域气候和碳循环过程中起着重要作用[1]。草地植物的根系主要以细根(直径 < 2 mm)为主, 是土壤中变化最活跃的部分[2—3], 根系通过连接和固着地上部分, 穿插于土壤中, 为吸收养分和水分、固定地上部分的重要器官, 在储藏、输送营养物质, 调节植物生理功能等方面有重要作用[4], 同时也是陆地生态系统重要碳汇和养分库。根系的生长特征与陆地生态系统生产力息息相关, 是生态系统物质循环和能量流动的重要组成部分[5], 近年来已成为生态系统碳分配格局与过程研究的核心环节。因此加强根系空间分配格局、根系形态、结构特征[6—8]的研究有助于了解草地生态系统地下根系碳分配、碳转换对全球气候变化的响应机制。
人类活动的加剧导致全球气候变化, 据预测, 未来全球气温预计还将继续上升0.3—4.8℃, 降水的变异幅度将会增加, 其中区域性降雨格局变化正在继续和扩大[9]。作为影响植物根系形态特征最重要的环境因子[10], 由降雨量变化导致的土壤水分变化在很大程度上影响了植物根系的形态结构和在土壤中的垂直分布格局[11]。降雨量减少导致的干旱会胁迫植物根系向更深土层生长[12], 同时显著增加较细直径的根系数量, 减少较粗根系数量[13]以获取养分和水分。这主要是由于在植物根系系统中, 不同直径和根序的根系生态功能并不完全相同, 根据Fitter[14]根系发育理论, 生长在根系末端的根系为1级根, 1级起源于2级根、2级根起源于3级根, 以此类推形成根序, 其中低等级根系主要起吸收养分和水分等作用, 具有氮含量高和碳含量低、皮层组织发达的特点, 高等级根系起运输和支持作用, 皮层组织基本消失, 但碳含量丰富[15]。可见不同根序根系的生态功能决定了植物根系对降水变化响应的差异。然而, 关于降雨量变化对不同根序根系生长特征的研究结论并不一致, 有研究发现随着降雨量的增加, 直径小于0.4 mm的根系(低级根)现存量增加[16], 而大于0.5 mm的根系(高级根)现存量会减少;也有研究指出随着降雨减少, 主根(高级根)根系现存量增加[17]。尽管研究人员认识到水分胁迫对不同径级根系的生长特征具有不同影响, 但对于不同根序根系是如何响应降雨的变化研究较少。此外大多数研究集中在不同降雨量下森林生态系统中根系特征的变化, 而对不同降雨量下草地系统尤其是高寒草甸生态系统植物不同根序根系生长特征的研究还鲜见报道。
青藏高原高寒草甸是世界上海拔最高、面积最大、类型最独特的草地生态系统, 对北半球气候有着启动和调节的作用[18]。在全球变化下, 青藏高原降水事件预计将变得更加频繁和严重[19], 这将引起土壤水热条件季节性变化[20], 直接或者间接地影响植物地上植物群落和根系资源获取策略和生物量分配格局[21], 进而影响群落结构和物种多样性。因此研究高寒草甸植物根系对降水变化的适应性及其维持机制显得尤为重要。由于根系的不可见性、根系生产、死亡和周转的多变性以及根系研究手段的欠缺、常规研究方法(如挖掘法)采样破坏性大等, 限制了我们对根系生长特征的深入了解。微根管技术(minirhizotron)是目前应用最广的根系原位、重复测量的方法之一, 与根钻法和内生长法相比, 它最大的优点是在植物生长稳定后, 可在不被干扰生长环境的情况下, 实现对根系长期的定位观测。综上, 本文以川西北高寒草甸为研究对象, 利用微根管技术监测根系对降雨量改变的适应过程及特征, 结合样方法调查高寒草甸植物群落生物量, 旨在探讨降雨量改变下:1)高寒草甸群落及功能群生物量响应机制;2)不同根序根系现存量、生产量、死亡量的变化规律以及空间分布格局;3)根系寿命、周转对高寒草甸地下生态系统碳分配特征的影响;4)不同根序根系特征与土壤理化性质间的相互关系。以期进一步揭示高寒草甸不同径级根系特征及其对降雨变化的响应机制, 为青藏高原高寒草甸生态系统功能的管理和可持续利用提供理论依据。
1 材料和方法 1.1 研究区自然概况本实验设置于西南民族大学生态保护与畜牧高科技研发实验基地(32°49.823′ N, 103°35.237′ E), 位于青藏高原东部红原县。该区域属于高原寒温带半湿润季风气候, 平均海拔约3500 m, 年平均气温和年平均降水量分别为1.4℃、769.20 mm, 且80%降水集中在5—9月。土壤类型为亚高山草甸土, 土壤容重为1.04 g/cm3, 田间持水量为51.52%, 凋萎系数约为29.06%[22—24], 主要植被类型有垂穗披碱草(Elymus nutans)、四川嵩草(Kobresia setchwanensis)和剪股颖(Agrostis matsumurae)等, 杂类草植物包括钝苞雪莲(Saussurea nigrescens)、条叶银莲花(Anemone coelestina)和鹅绒委陵菜(Potentilla anserina)等[25]。
1.2 实验设计 1.2.1 实验样地设置在实验基地内选择地势相对平坦、地上植被分布较均匀的天然高寒草甸作为试验样地, 自2015年12月进行模拟降雨实验, 样地面积为50 m×50 m, 使用围栏进行围封以减少放牧干扰。本实验使用完全随机区组试验, 分五个模拟降雨处理(0.1P、0.5P、0.7P、1.0P和1.5P, 1.0P为自然降水, 该装置利用由高透光的有机玻璃作为挡雨板进行截流, PVC管材质的集雨管和集雨桶收集自然降雨, 采用均匀放置的面积为小区面积90%、50%和30%的高透光有机玻璃挡水板, 实现0.1P, 0.5P和0.7P3个减雨梯度, 其中将0.5P处理下挡水板截留的自然降雨收集在集雨桶中, 用人工浇水的方式均匀洒在1.5P小区中, 实现1.5P的增雨梯度, 挡雨板长度超过每个样方20 cm, 减雨棚最低处距地面1 m, 以保证近地表空气流通, 最高处为2 m以便观测采样), 每个处理6个重复共30个样方。单个样方大小为2 m×2 m, 小样方之间留有至少2 m间隔, 同时样方四周用铝皮埋到40 cm处防止土壤中水分的横向流动。
同期, 在上述每个样方内安装一根聚碳酸酯材料微根管, 微根管长度为100 cm, 内径5 cm, 外径6 cm。在样方内用土钻钻取一个与地面呈30°, 深度为60 cm, 直径大小为7 cm的孔, 然后将微根管密封的一端插入孔中, 并把未密封的露出地面的一端盖上配套的黑色胶盖, 用黑色塑料袋包裹露出地面的微根管(防止漏水和光照对实验产生干扰)。此外, 在微根管周围用钻出的土填平, 使土壤和微根管紧密接触, 安装好的微根管半年内不进行观测, 等待根系充分恢复。于2018年5月至9月进行根系图片的采集。此外2018年1.5P、1.0P、0.7P、0.5P、0.1P累计降雨量分别1291.2 mm、860.8 mm、602.56 mm、430.4 mm、86.08 mm(集雨桶测定值)。
1.3 数据采集和处理 1.3.1 群落地上生物量的测定于2018年8月中旬在所有样方中选取一个具有代表性的50 cm×50 cm的小样方, 并将小样方中所有植物齐地面刈割, 按功能群(禾本科、莎草科、豆科、杂类草)分类, 65 ℃下48 h烘干至恒重。
1.3.2 土壤紧实度和含水量的测定于2018年4月28日至同年9月24日每隔7天用土壤水分仪(Spectrum Field Scout TDR300, USA)和土壤紧实度仪(Spectrum Field Scout SC 900, USA)测定0—10 cm和10—20 cm土层土壤含水量(Soil water content, SWC)和紧实度(Soil compaction, SC)。
1.3.3 土壤样品的采集与测定于2018年8月中旬, 在2 m×2 m样方内, 用内径5 cm的土钻分层(0—10 cm和10—20 cm)按“S”形钻取3钻, 混合为1个土壤样, 每个处理重复3次, 并做好标记, 样品风干后过0.15 mm筛。用于测定土壤全碳(TC)、全氮(TN)、全磷(TP)含量。其中, 采用杜马斯燃烧法[26]测定全碳、全氮含量, 钼锑抗比色法[27]测定土壤全磷。
于2021年10月中旬, 用环刀法测量土壤容重(BD)、土壤田间持水量(FC)[28—30]。并将田间持水量(质量含水量)转换为体积含水量。
质量含水量FC转换为体积含水量FCv公式如下[31]:
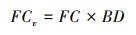
|
式中FC为质量含水量, BD为土壤容重。
1.3.4 根系图片的采集与分析于2018年5月8日至同年10月24日采用CI—600根系扫描仪(CID Bio-science, Camas, WA, USA)进行图像采集, 首先将微根管外包裹的黑色塑料袋打开, 打开顶端盖子, 用棉布将微根管内的水蒸气擦干, 等待15 min, 当微根管内外温差平衡后, 将标定好的摄像机放入微根管内并完成图片采集, 并用马克笔在微根管上标记好第一次摄像头放入的位置。每根微根管按垂直深度分为0—10 cm和10—20 cm土层分别采集图像数据。每次测定结束后, 将露出地面的根管按第一次的方法包裹好, 每2周采集一次图像, 共取12次图像, 总共追踪标记到6985条根系。
将采集的根系图片导入WinRHZIO Tron MF(CID Bio-Science Inc, Camas, WA, USA)根系分析软件中, 描绘图像中的根系, 所有根系长度的数据精确到0.1 mm。通过观察根系的颜色判断死活, 所有黑色和消失的根系作为死根, 褐色和白色的根系作为活根, 同时, 将所有根按直径分3个等级[32](分别为第1级0—0.3 mm、第2级0.3—0.8 mm、第3级0.8 mm以上)。根系的形态指标主要有根系现存量、生产量、寿命、死亡量和周转。其中根长密度、根系单位体积生物量、根系单位面积生物量、根系现存量、生产量、死亡量和周转按照史建伟等[33]的计算方法处理。
首先通过根系图像分析得到根系的长度, 并利用如下公式计算出单位体积的根长密度(RLDv, m/m3), 计算公式如下:
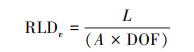
|
式中, L(m)为图像描绘得到的根系长度;A(m2)为观测窗的面积, 为0.1956 m×0.2159 m(根系扫描仪所采集图像大小);DOF(m)为微根管到周围土壤的距离, 一般在0.002—0.003 m[32]。本实验草地群落细根直径小, DOF取值为0.002 m[34]。
其次, 通过比根长(SRL, m/g)将RLDv转化为单位体积生物量RBDv(g/m3), 计算公式如下:
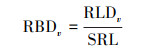
|
式中SRL表示用土钻法所钻取的单位质量的根系长度(m/g)。
最后, 将RBDv乘以取样土壤的剖面深度(D)得到单位面积根系生物量RBDA(g/m2), 计算公式如下:
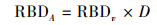
|
根系生产量定义为在t时不存在而在t+1时出现或伸长的所有新根的总和;同样, 根系死亡量量定义为t时至t+1时消失根和死根的总和;此外根系现存量定义为t时活根的总和。
根系周转率是运用上述方法所求出的根系生产量和现存量进行计算, 公式如下:
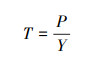
|
式中T表示根系周转率(1/a);P为细根年总产量(年生物量总产量);Y为活细根现存量平均值(平均生物量)。
根系寿命[35]定义为自第一次在影像出现时刻到第一次被定义为死亡或是消失的时间, 计算公式如下:
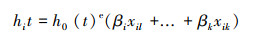
|
式中t表示在此时根系的死亡量, βi—βk表示死亡量增加的风险协变量, xil—xik表示危险因素。本研究中不同径级根系寿命的估算通过Excel 2007和IBM SPSS Statistics 22共同分析得出, 在Excel 2007中对不同径级根系的存活状态和存活天数进行计算, 再用IBM SPSS Statistics 22中Kaplan-Meler方法计算寿命, 做出根系存活曲线, 并求出中位生存时间, 即根系达到50%死亡率所需的天数。
1.4 统计分析利用IBM SPSS Statistics 22软件进行数据分析, 对不同降雨处理下植物地上生物量、土壤理化性质(土壤TN、TC、TP、SWC、SC、BD和FC)、根系现存量、生产量、死亡量、寿命和周转进行单因素方差分析(One—way analysis of variance, One—way ANOVA), 用最小显著差数法(Least significant different, LSD)进行处理间差异显著性检验(P < 0.05)。不同土层间(0—10 cm和10—20 cm)的差异则采用独立样本t检验。采用多因素方差法(Multi—factor analysis of variance)分析降雨处理、根系径级和土层深度及交互作用对根系现存量、生产量、死亡量、寿命和周转的影响。使用R4.0.3中corrplot包分析土壤理化性质与根系特征的皮尔森相关性。最后, 利用AMOS 22.0组建结构方程模型(SEM)分析根系特征和环境因子之间相互影响关系。
2 结果 2.1 土壤理化性质的变化在0—10 cm土层中, 与自然降雨(1.0P)相比, 田间持水量分别在增雨50%(1.5P)、减雨50%(0.5P)和减雨90%(0.1P)显著增加26%、18%和22%(P < 0.05)。土壤含水量随降雨量减少呈显著下降趋势, 且0—10 cm显著高于10—20 cm土层(P < 0.05)。与自然降雨(1.0P)相比, 在0—10 cm土层中, 土壤含水量在1.5P显著增加了6%, 在0.7P、0.5P和0.1P分别显著降低5%、11%和33%(P < 0.05);10—20 cm土层, 与自然降雨(1.0P)相比, 1.5P显著提高了土壤含水量(5%), 减雨梯度则分别降低了土壤含水量3%、7%和25%(表 1)。在0.1P中, 0—10 cm土层和10—20 cm土层土壤含水量分别为田间持水量的50.73%和44.11%, 处于轻度或中度水分胁迫[36]。
| 土层 Soil layer/cm |
降雨处理 Precipitation treatment |
田间持水量 FC/% |
含水量 SWC/% |
紧实度 SC/kPa |
容重 BD/(g/cm3) |
全碳 TC/(g/Kg) |
全氮 TN/(g/Kg) |
全磷 TP/(g/Kg) |
| 0—10 | 1.5P | 65.83±7.37 Ab | 44.31±0.51 Aa | 165.97±2.6 Ab | 1.07±0.03 Aab | 66.85±5.65 Aa | 2.19±0.10 Aa | 1.18±0.04 Aa |
| 1.0P | 52.11±3.05 Aa | 41.85±1.09 Ab | 172.32±7.93 Ab | 0.95±0.09 Aa | 68.87±9.54 Aa | 2.33±0.42 Aa | 1.23±0.06 Aa | |
| 0.7P | 51.49±2.16 Aa | 39.98±1.12 Ab | 177.51±4.22 Ab | 1.00±0.02 Aab | 78.13±2.72 Aa | 3.26±0.36 Aa | 1.22±0.07 Aa | |
| 0.5P | 61.28±4.75 Ab | 37.09±1.12 Ab | 178.83±4.85 Ab | 1.06±0.03 Aab | 72.74±5.65 Aa | 2.46±0.51 Aa | 1.14±0.04 Aa | |
| 0.1P | 63.57±11.8 Ab | 28.24±2.13 Ac | 224.28±3.55 Aa | 1.15±0.03 Ab | 79.73±5.36 Aa | 2.78±0.26 Aa | 1.15±0.03 Aa | |
| 10—20 | 1.5P | 44.37±4.61 Bab | 36.02±0.59 Ba | 150.33±8.13 Abc | 1.25±0.05 Aa | 52.34±3.90 Aa | 2.06±0.61 Aa | 1.03±0.04 Aa |
| 1.0P | 47.37±4.33 Bab | 34.27±0.69 Bab | 167.86±3.39 Abc | 1.18±0.04 Aa | 54.48±2.60 Aa | 2.00±0.19 Aa | 1.14±0.01 Aa | |
| 0.7P | 42.5±4.473 Ba | 33.24±0.62 Bab | 168.14±2.56 Ab | 1.12±0.1 Aa | 53.53±3.04 Ba | 1.76±0.12 Ba | 1.14±0.05 Aa | |
| 0.5P | 51.47±4 Bab | 31.82±1.09 Bb | 174.12±2.68 Ab | 1.16±0.03 Aa | 48.49±5.57 Ba | 1.83±0.22 Aa | 1.00±0.11 Aa | |
| 0.1P | 57.12±3.91 Ab | 25.81±2.1 Ac | 196.06±5.99 Ba | 1.21±0.02 Ba | 50.91±6.54 Ba | 2.19±0.56 Aa | 0.92±0.07 Ba | |
| 不同小写字母表示不同处理间相同土层土壤理化性质差异显著, 不同大写字母表示同一处理不同土层间土壤理化性质差异显著(P < 0.05);FC:田间持水量field capacity;SWC:土壤含水量Soil water content;SC:土壤紧实度Soil compaction;BD:容重Bulk density;TC:全碳Total carbon;TN:全氮Total nitrogen;TP:全磷Total phosphorus | ||||||||
土壤紧实度随降雨量减少呈上升趋势, 0—10 cm和10—20 cm土层土壤紧实度在0.1P分别提高了30%和17%(P < 0.05)。此外, 0—10 cm土层0.1P土壤容重显著升高。
与自然降雨相比, 土壤全碳在0—10 cm土层中随着降雨量减少呈上升趋势, 全氮、全磷随降雨量减少呈下降趋势;在10—20 cm土层, 土壤全碳、全氮和全磷随降雨量减少呈下降趋势。
2.2 植物群落特征的变化增雨和减雨没有显著改变植物群落总生物量。减雨处理显著增加禾本科植物生物量, 尤其是0.5P和0.1P分别显著增加禾本科植物生物量129%和151%(P < 0.05), 降低豆科和莎草科生物量, 但不显著;增雨对不同植物功能群生物量无显著影响(图 1)。

|
| 图 1 不同降雨处理下功能群生物量变化 Fig. 1 Changes of community above-ground biomass under different Precipitation treatment 小写字母表示不同处理之间差异显著(P < 0.05) |
降雨处理、土层及二者的交互作用对植物群落根系现存量影响不同(表 2)。0—10 cm土层, 0.7P和1.5P显著增加1级根现存量, 增雨和减雨均显著增加2、3级根现存量。10—20 cm土层, 1级根现存量在0.7P、0.1P显著增加(P < 0.05)。此外各土层总根系现存量在处理间无显著差异(图 2)。
| 差异来源 Source of variation |
根系现存量 Root standing crop |
根系生产量 Root production |
根系死亡量 Root Mortality |
根系周转 Root Turnover |
根系寿命 Root Life span |
|||||||||
| F | P | F | P | F | P | F | P | F | P | |||||
| 土层Soil layer | 20.646 | < 0.001 | 27.621 | < 0.001 | 24.22 | < 0.001 | 2.079 | 0.056 | 0.032 | 0.86 | ||||
| 处理Treatment | 1.241 | 0.326 | 3.513 | 0.05 | 4.774 | < 0.01 | 0.779 | 0.552 | 1.736 | 0.182 | ||||
| 根系径级Root diameter | 112.098 | < 0.001 | 149.07 | < 0.001 | 297.116 | < 0.001 | 3.899 | 0.05 | 14.185 | < 0.001 | ||||
| 深度×处理Depth×Treatment | 0.635 | 0.644 | 0.594 | 0.671 | 1.407 | 0.268 | 0.277 | 0.889 | 1.863 | 0.157 | ||||
| 深度×根系径级Depth×Root diameter | 10.939 | < 0.001 | 27.351 | < 0.001 | 0.309 | 0.872 | 0.309 | 0.872 | 0.659 | 0.422 | ||||
| 处理×根系径级Treatment×Root diameter | 0.333 | 0.992 | 2.714 | < 0.05 | 2.532 | < 0.001 | 0.812 | 0.15 | 0.298 | 0.878 | ||||
| 深度×处理×根系径级Depth×Treatment×Root diameter | 0.549 | 0.913 | 0.731 | 0.756 | 0.163 | 0.982 | 0.163 | 0.977 | 0.061 | 0.993 | ||||

|
| 图 2 不同径级根系现存量对不同降雨量的响应 Fig. 2 Standing crop of roots with different diameter classes in response to precipitation gradients 小写字母表示不同处理之间差异显著(P < 0.05), ns表示同一土层不同降雨处理之间差异不显著 |
根系生产量主要受到土层和处理的影响(表 2)。在0—10 cm土层, 增雨和减雨均降低了根系生产量, 且随降雨量增加显著降低, 1级根生产量分别在0.5P、0.1P显著降低了39%和56%(P < 0.05), 2级根系生产量在0.1P显著降低了66%(图 3)。在10—20 cm土层, 1级根、总根生产量在0.1P下显著降低(P < 0.05)。

|
| 图 3 不同径级根系生产量对不同降雨量的响应 Fig. 3 Production of roots with different diameter classes in response to precipitation gradients 小写字母表示不同处理之间差异显著(P < 0.05) |
土层和处理是影响根系死亡量的主要因素(表 2)。在0—10 cm土层, 1.5P、0.7P、0.5P和0.1P都降低了2、3级根和总根系死亡量, 且随着降雨程度显著降低(P < 0.05)。在10—20 cm土层, 1.5P、0.7P、0.5P和0.1P显著降低了2级根系死亡量(P < 0.05)(图 4)。

|
| 图 4 不同径级根系死亡量对不同降雨量的响应 Fig. 4 Mortality of roots with different diameter classes in response to precipitation gradients 小写字母表示不同处理之间差异显著(P < 0.05) |
在0—10 cm土层, 与自然降雨相比, 增雨和减雨均降低了1级根周转, 增加了2级根周转, 尤其是1.5P。在10—20 cm土层, 0.1P和1.5P明显抑制了1级根和2级根周转速率;0.5P抑制了1级根周转速率, 但明显增加了2级根周转;0.7P提高了1级和2级根周转速率(图 5)。

|
| 图 5 不同径级根系周转对不同降雨量的响应 Fig. 5 Turnover of roots with different diameter classes in response to precipitation gradients 小写字母表示不同处理之间差异显著(P < 0.05) |
不同径级根系寿命差异显著(F=14.185, P < 0.001)(表 2)。在0—10 cm中, 1级根系寿命在0.1P显著增加(P < 0.05);在10—20 cm土层, 2级根和总根寿命在0.1P显著增加(P < 0.05)(表 3)。
| 土层/cm Layer |
根级 Root diameter classes |
0.1 P | 0.5 P | 0.7 P | 1.0 P | 1.5 P |
| 0—10 | 1级根 | 102.64±2.21Ba | 99.91±14.60Aa | 111.76±10.72Aa | 102.99±12.81Ba | 105.15±6.075Aa |
| 2级根 | 134.16±18.08Aa | 112.00±22.26Ab | 125.29±6.52Ab | 121.75±10.45Ab | 126.25±24.93Ab | |
| 总根 | 103.03±20.49Ba | 100.41±14.38Aa | 112.45±9.78Aa | 103.674±5.31Aa | 106.15±5.34Aa | |
| 10—20 | 1级根 | 122.77±24.52Aa | 98.58±18.63Ab | 105.73±13.47Ab | 111.28±4.91Ab | 100.47±26.97Ab |
| 2级根 | 117.23±13.57Aa | 104.25±16.95Aa | 104.25±16.96Aa | 122.80±18.58Aa | 124.37±21.53Aa | |
| 总根 | 136.13±6.77Aa | 98.05±18.98Ab | 104.65±13.18Bb | 102.60±7.26Ab | 100.84±26.78Ab | |
| 不同大写字母表示不同土层根系寿命差异显著, 小写字母表示处理间根系寿命差异显著(P < 0.05) | ||||||
如图 6所示, 减雨一定程度上增加了10—20 cm土层根系存活率, 0.1P处理下1级和总根系存活率在生长季内显著升高。

|
| 图 6 不同径级根系累计存活率对不同降雨量的响应 Fig. 6 Life span of roots with different diameter classes in response to precipitation gradients |
对土壤理化性质和根系特征进行相关性分析, 结果表明根系现存量与SWC、TC、TN、TP显著正相关(P < 0.05);根系生产量与TN、TP显著增相关(P < 0.05);根系周转与TC显著负相关(P < 0.05);根系寿命与TC、TN显著正相关(P < 0.05)(图 7)。

|
| 图 7 土壤理化性质和根系特征的相关性 Fig. 7 Pearson correlation between soil physicochemical properties and root characteristics *表示P < 0.05;颜色接近白色, 表示相关性低;颜色越深, 表示相关性高; ST:现存量Standing crop;PR:生产量Production;TU:周转Turnover;MO:死亡量Mortality;LS:寿命Life span |
相关性分析结构表明土壤紧实度(SC)对根系特征无显著影响(图 7), 因此本文将土壤含水量作为主要环境因子纳入结构方程模型(SEM), 进一步分析根系特征与环境因子之间的相互影响关系(图 8)。降雨量通过直接改变土壤含水量进而影响根系生产量且显著正相关(P < 0.05);同时土层深度直接影响土壤含水量、土壤养分、根系生产量和现存量且显著负相关(P < 0.05);根系径级直接影响根系现存量、生产量和周转, 并极显著负相关(P < 0.001);此外, 土壤养分直接影响根系周转(P < 0.05)。

|
| 图 8 根系特征与环境因子结构方程模型 Fig. 8 Structural Equation Model of root system characteristics and environmental factors ***表示P < 0.001;**表示P < 0.01;*表示P < 0.05。红色实线表示显著正相关, 蓝色实线表示显著负相关, 虚线表示无显著相关 |
研究表明降水增加对藏北高寒草原化草甸植物地上生产力有显著正效应[37], 生产力对气候变化的响应依赖于主要功能群之间的补偿作用[38]。在本研究中植物地上生产力不随降雨量改变而显著变化, 但不同功能群生物量对水分变化的响应差异显著, 减雨处理(0.1P和0.5P)下禾本科生物量显著增加, 而莎草科生物量降低, 该结果与Liu等[39]在高寒草甸研究结果相似。这是因为禾本科植物属于旱生型[40], 莎草科植物一般属于湿生或中生型[41]。与莎草科相比, 禾本科植物根系更深, 高寒草甸生态系统中, 深层土壤水分的储存量大[42], 增加禾本科植物的丰度可以使群落整体获得更多的水分, 而水分获取能力的增加保障了群落度过干旱期。因此在干旱下高寒草甸生态系统禾本科增加的生物量补偿了莎草科减少的生物量, 功能群之间的补偿机制保障了生态系统的生产力和稳定性。此外在植物个体水平上, 随着土壤水分降低(表 1), 叶片气孔部分关闭, 蒸腾速率下降[43], 叶片水分流失减缓, 减少了水分亏缺对叶片的伤害[44], 而光合酶活性和水分利用效率在水分胁迫下增加[45], 使得叶片的光合速率保持正常水平, 生物量得以维持。
3.2 不同降雨量对根系现存量、死亡量和生产量的影响根系现存量是植物对土壤中资源有效性的反映[46]。本研究表明, 高寒草甸植物根系现存量主要集中在0—10 cm土层, 占总根系现存量的69.17%, 降雨处理并未显著改变总根系现存量, 但显著改变了土层间根系现存量的比例, 这与Zhang等[47]的研究结果相似, 在高寒草甸中, 土壤碳氮磷含量随土层深入而降低[48], 土壤理化性质的差异是导致根系现存量垂直下降的主要原因[49](图 7)。此外0—10 cm土层土壤有效水分涵养能力高于10—20 cm土层[50], 土壤水分的不同又会限制其他养分(如氮素)的有效性[51]。当10—20 cm土层有效水分和养分低于0—10 cm土层时, 植物为了获取更多有效资源, 会将更多光合产物分配到0—10 cm土层根系。值得我们注意的是随着水分减少, 0—10 cm土层2、3级根现存量显著低于对照(P < 0.05), 但1级根现存量呈增加趋势, 其中0.7P 1级根现存量显著增加(P < 0.05)(图 2)。这可能是因为1级根与2、3级根相比, 1细根细胞中具有发达的薄壁组织, 代谢更为旺盛, 对土壤环境变化会产生更明显的生理生化反应[52], 其主要的功能是吸收[53], 随着土壤含水量的降低, 植物将投入更多的光合产物到1级根以获取更多的水分, 而降低了对2、3级根的分配(表 2)。这种由降雨变化引起的植物对不同土层及径级根系生物量分配差异是植物面对恶劣条件下的生存策略[15], 地下生物量只在不同径级根系之间的转移和再分配是对全球变化下(如:降雨量改变)根系—土壤碳氮功能维持和权衡的适应性策略。
根系生产量和死亡量是影响碳循环的重要指标, 研究表明细根的生产量和死亡量占全球净初级生产的30%[54], 植物根系不仅能通过根系生产储存大量光合作用合成的有机碳, 而且也能通过周转将死亡根系中的有机碳转移到土壤中[55]。根据已有的报道, 在水分限制条件下, 土壤含水量是影响根系生产量的主要因素[56], 根系在适当干旱条件下会选择垂直或水平地延伸到土壤中[57], 根系生产量增加;随着干旱程度的加剧(0.1P), 根系生产量和死亡量显著降低(表 3), 这和李文娆等[13]的研究结果相似。当水分减少时, 根系通过增加在土壤中所占的体积和根长密度等策略来提高对土壤水分的获取能力[58], 更多光合产物将会分配到根系, 与此同时植物又会消耗更多的光合产物用于维持根系的正常生理活动, 这种供需关系使根系在水分胁迫下生长缓慢[46], 生产量降低。此外在养分和水分贫瘠的环境中, 植物会降低死亡来储存所需要的营养物质[59]。
本研究进一步发现根系生产主要集中发生在1级根系。一方面, 相同体积的细根较粗根有更大的表面积, 植物会选择促进根系变长、变细, 增大与土壤接触面积来扩大水分吸收范围, 加强对土壤水分的利用[13], 另一方面植物将有限的水分和养分集中用于幼嫩器官(1级根系)的保护及维持, 其目的是延长植物的存活时间, 增加存活的概率, 有利于复水后植物的恢复[60]。因此, 随着水分减少, 植物会将有限的碳分配给1级根系中, 通过1级根系的生长来扩大水分吸收的范围。
3.3 不同降雨量对根系寿命、周转的影响根系寿命及周转是影响生态系统生产力和养分循环的重要因素, 根据已有的报道, 根系直径增加1 mm, 寿命延长了99%[61]。这与本研究结果相似(表 3), 这是因为不同直径根系结构差异大, 直径较大的根细胞氮浓度低, 呼吸速率弱, 寿命也相对较长[15];其次粗根木质化程度高, 当面对土壤病原微生物或是植食性动物采食时, 粗根往往具有更高的抵抗性。同时史建伟等[35]对不同分枝根序寿命的研究表明, 1级根中值寿命表现随土层深度增加而增加趋势, 本实验也表明0.1P处理下1级根系在10—20 cm土层寿命高于0—10 cm, 且高于相同土层其余处理寿命(表 3)。本研究区位于海拔较高的高寒草甸生态系统, 昼夜温差大, 与10—20 cm土层相比, 0—10 cm土层土壤温度、水分波动更大, 而10—20 cm土层稳定的环境有助于细根寿命的维持。
细根周转不仅在植被碳库与土壤碳库交换过程中扮演着重要角色, 也是陆地生态系统土壤碳汇的重要途径, 通过细根周转进入土壤的碳占土壤总输入碳量的14—50%[62]。此外根系周转对全球变化, 如气候变暖、降水量改变等具有很强的敏感性[63]。因此研究根系周转及其影响因子是认识生态系统碳分配和养分循环的关键。具体来说, 根系周转是指根系从产生到死亡并分解消失的过程, 根系周转值越大, 植被利用营养元素的能力就越强[43], 有研究表明随着水分减少, 根系周转呈现出线性降低的趋势[64]。本研究结果发现, 根系周转随降雨减少而降低, 但处理间无显著差异, 这是因为根系周转主要受土壤养分、土壤温度和土壤水分的调控[39], 如氮的有效性增加会导致根系活性增加, 周转加快[65], 本实验中土壤含水量随着降雨减少而降低, 但土壤中养分含量却呈升高趋势(表 1), 在减雨处理中, 水分降低减缓了较高的氮从土壤表面向根系的质量流和扩散, 导致根系表面养分有效性降低[66], 土壤养分无法被植物充分利用, 根系呼吸速率变慢, 周转速率降低。因此随着降雨量的减少, 细根的周转率较低、寿命长, 减缓了根系的碳消耗, 这也可能是植物根系维持生长而获取更深层土壤水分的一种适应性表现。
3.4 根系特征与环境因子的相关性分析不同径级根系对降水减少和增加响应的异质性是多因素的, 确定高寒草甸细根在土壤剖面中对降水变化的响应对于预测地下碳循环具有重要意义。以往的研究表明, 土壤含水量和土层深度是影响根系垂直分布的主要因素, 本研究中也同样支持这个观点, 相关性分析表明土壤含水量与根系生产量、现存量正相关, 且随着土层深度增加, 根系生产量、现存量减少。这是因为高寒草甸土壤具有全量养分丰富而速效养分匮乏的特点[67], 植物生长季节短暂, 寒冷季漫长, 这种细根表聚现象可能是植物根系对高寒草甸恶劣环境的生存策略, 细根在0—10 cm土层有更多的生命活动是为了能够在恶劣环境内迅速地获取更多的养分。同时相关研究表明降水减少限制了直径较小的细根和细根生物量, 并且刺激了根系分解, 加速了周转[68]。而在本研究中, 随着水分的减少, 1级根系增加, 现存量受限制, 但根系死亡量减少, 根系周转减缓。此时不同直径根系碳分配优化, 细根生产所需消耗的碳及其获取资源能力的效益成为植物生长的关键, 在高寒草甸生态系统中, 植物更多的细根(1级根)和较长的寿命提供了根系可持续性的资源获取性和时间的保障, 同时1级根的碳利用率也得到提高。
4 结论1) 降雨变化对地上群落生物量无显著影响, 但轻度水分胁迫(0.1P)显著增加禾本科生物量。2)土壤水分是影响根系生产量和根系现存量的重要因子。在0—10 cm土层, 根系在较小降雨改变(1.5P、0.7P)下将分配更多生物量到1级根中, 用于资源的争夺和获取。3)1级根系对水分的变化更为敏感, 主要执行感知水分以及吸收功能, 2级、3级根系对水分变化无显著响应。4)轻度水分胁迫(0.1P)下, 10—20 cm土层1级和总根系寿命延长, 根系周转降低。
| [1] |
Scurlock J M O, Hall D O. The global carbon sink: a grassland perspective. Global Change Biology, 1998, 4(2): 229-233. DOI:10.1046/j.1365-2486.1998.00151.x |
| [2] |
李鹏, 李占斌, 鲁克新. 黄土区草本植被根系与土壤垂直侵蚀产沙关系研究. 植物生态学报, 2006, 30(2): 302-306. DOI:10.3321/j.issn:1005-264X.2006.02.013 |
| [3] |
Peters D P C. Plant species dominance at a grassland-shrubland ecotone: an individual-based gap dynamics model of herbaceous and woody species. Ecological Modelling, 2002, 152(1): 5-32. DOI:10.1016/S0304-3800(01)00460-4 |
| [4] |
王政权, 郭大立. 根系生态学. 植物生态学报, 2008, 32(6): 1213-1216. DOI:10.3773/j.issn.1005-264x.2008.06.001 |
| [5] |
Trumbore S E, Gaudinski J B. The secret lives of roots. Science, 2003, 302(5649): 1344-1345. DOI:10.1126/science.1091841 |
| [6] |
纪娇娇, 张秋芳, 杨智杰, 熊德成, 刘小飞, 胥超. 模拟氮沉降对中亚热带杉木幼树根系生物量的影响. 生态学报, 2020, 40(17): 6118-6125. |
| [7] |
Colchado-López J, Cervantes R C, Rosas U. A linear model to describe branching and allometry in root architecture. Plants, 2019, 8(7): 218. DOI:10.3390/plants8070218 |
| [8] |
Glimskär A. Estimates of root system topology of five plant species grown at steady-state nutrition. Plant and Soil, 2000, 227(1): 249-256. |
| [9] |
沈永平, 王国亚. IPCC第一工作组第五次评估报告对全球气候变化认知的最新科学要点. 冰川冻土, 2013, 35(5): 1068-1076. DOI:10.7522/j.issn.1000-0240.2013.0120 |
| [10] |
Pilon R, Picon-Cochard C, Bloor J M G, Revaillot S, Kuhn E, Falcimagne R, Balandier P, Soussana J F. Grassland root demography responses to multiple climate change drivers depend on root morphology. Plant and Soil, 2012, 364(1/2): 395-408. |
| [11] |
Zhang B W, Cadotte M W, Chen S P, Tan X R, You C H, Ren T T, Chen M L, Wang S S, Li W J, Chu C J, Jiang L, Bai Y F, Huang J H, Han X G. Plants alter their vertical root distribution rather than biomass allocation in response to changing precipitation. Ecology, 2019, 100(11): e02828. |
| [12] |
刘庚山, 郭安红, 任三学, 安顺清, 林日暖, 赵花荣. 人工控制有限供水对冬小麦根系生长及土壤水分利用的影响. 生态学报, 2003, 23(11): 2342-2352. DOI:10.3321/j.issn:1000-0933.2003.11.018 |
| [13] |
李文娆, 张岁岐, 丁圣彦, 山仑. 干旱胁迫下紫花苜蓿根系形态变化及与水分利用的关系. 生态学报, 2010, 30(19): 5140-5150. |
| [14] |
Fitter A H, Stickland T R. Architectural analysis of plant root systems 2. Influence of nutrient supply on architecture in contrasting plant species. New Phytologist, 1991, 118(3): 383-389. DOI:10.1111/j.1469-8137.1991.tb00019.x |
| [15] |
Guo D L, Xia M X, Wei X, Chang W J, Liu Y, Wang Z Q. Anatomical traits associated with absorption and mycorrhizal colonization are linked to root branch order in twenty-three Chinese temperate tree species. New Phytologist, 2008, 180(3): 673-683. DOI:10.1111/j.1469-8137.2008.02573.x |
| [16] |
Zhang J, Wang P, Xue K, Hao Y B, Wang Y F, Cui X Y. Trait complementarity between fine roots of Stipa purpurea and their associated arbuscular mycorrhizal fungi along a precipitation gradient in Tibetan alpine steppe. Journal of Mountain Science, 2019, 16(3): 542-547. DOI:10.1007/s11629-018-5311-9 |
| [17] |
段桂芳, 单立山, 李毅, 张正中, 张荣. 降水格局变化对红砂幼苗生长的影响. 生态学报, 2016, 36(20): 6457-6464. |
| [18] |
葛庆征, 魏斌, 张灵菲, 卫万荣, 黄彬, 江小雷, 张卫国. 草地恢复措施对高寒草甸植物群落的影响. 草业科学, 2012, 29(10): 1517-1520. |
| [19] |
许建伟, 高艳红, 彭保发, 王夏青. 1979-2016年青藏高原降水的变化特征及成因分析. 高原气象, 2020, 39(2): 234-244. |
| [20] |
Aanderud Z T, Schoolmaster Jr D R, Lennon J T. Plants mediate the sensitivity of soil respiration to rainfall variability. Ecosystems, 2011, 14(1): 156-167. DOI:10.1007/s10021-010-9401-y |
| [21] |
Zhu Y H, Weiner J, Yu M X, Li F M. Evolutionary agroecology: trends in root architecture during wheat breeding. Evolutionary Applications, 2019, 12(4): 733-743. DOI:10.1111/eva.12749 |
| [22] |
泽让东科, 文勇立, 艾鷖, 赵洪文, 陈有军. 放牧对青藏高原高寒草地土壤和生物量的影响. 草业科学, 2016, 33(10): 1975-1980. DOI:10.11829/j.issn.1001-0629.2016-0152 |
| [23] |
张光茹, 李红琴, 杨永胜, 王军邦, 祝景彬, 罗谨, 贺慧丹, 李英年. 基于主成分分析对退化高寒草甸不同恢复方式下草地质量的综合评价. 中国草地学报, 2020, 42(2): 76-82. |
| [24] |
Dai L C, Yuan Y M, Guo X W, Du Y G, Ke X, Zhang F W, Li Y K, Li Q, Lin L, Zhou H K, Cao G M. Soil water retention in alpine meadows under different degradation stages on the northeastern Qinghai-Tibet Plateau. Journal of Hydrology, 2020, 590: 125397. DOI:10.1016/j.jhydrol.2020.125397 |
| [25] |
罗雪萍, 阿的鲁骥, 字洪标, 杨有芳, 陈焱, 代迪, 王长庭. 高寒草甸土壤微生物功能多样性对积雪变化的响应. 冰川冻土, 2018, 40(5): 1016-1027. |
| [26] |
张薇, 付昀, 李季芳, 孟霞, 杨君. 基于凯氏定氮法与杜马斯燃烧法测定土壤全氮的比较研究. 中国农学通报, 2015, 31(35): 172-175. DOI:10.11924/j.issn.1000-6850.casb15060137 |
| [27] |
吴玉萍, 李应金, 赵立红, 刘亚丽, 夏振远. 连续流动分析法测定土壤中全磷、全钾的含量. 西南农业学报, 2013, 26(5): 1941-1945. |
| [28] |
杨伟德. 甘肃天祝亚高山草甸土的利用管理探讨. 草业科学, 2002, 19(8): 16-17. DOI:10.3969/j.issn.1001-0629.2002.08.005 |
| [29] |
祝景彬, 贺慧丹, 李红琴, 杨永胜, 未亚西, 罗谨, 李英年. 牧压梯度下高寒草甸土壤容重及持水能力的变化特征. 水土保持研究, 2018, 25(5): 66-71. |
| [30] |
金福一. 提高烘干法含水量数据准确度的研究. 中国水利, 2015(13): 56-57. DOI:10.3969/j.issn.1000-1123.2015.13.020 |
| [31] |
李炎, 王丹. 不同土壤水分测定方法的比较研究. 安徽农业科学, 2010, 38(17): 9110-9112. DOI:10.3969/j.issn.0517-6611.2010.17.106 |
| [32] |
Taylor H M, Huck M G, Klepper B, Lund Z F. Measurement of soil-grown roots in a Rhizotron. Agronomy Journal, 1970, 62(6): 807-809. DOI:10.2134/agronj1970.00021962006200060039x |
| [33] |
史建伟, 于水强, 于立忠, 韩有志, 王政权, 郭大立. 微根管在细根研究中的应用. 应用生态学报, 2006, 17(4): 715-719. DOI:10.3321/j.issn:1001-9332.2006.04.031 |
| [34] |
吴伊波, 车荣晓, 马双, 邓永翠, 朱敏健, 崔骁勇. 高寒草甸植被细根生产和周转的比较研究. 生态学报, 2014, 34(13): 3529-3537. |
| [35] |
史建伟, 秦晴, 陈建文. 柠条人工林细根不同分枝根序寿命估计. 生态学报, 2015, 35(12): 4045-4052. |
| [36] |
王琰, 陈建文, 狄晓艳. 不同油松种源光合和荧光参数对水分胁迫的响应特征. 生态学报, 2011, 31(23): 7031-7038. |
| [37] |
王志鹏, 张宪洲, 何永涛, 石培礼, 俎佳星, 牛犇, 李猛. 降水变化对藏北高寒草原化草甸降水利用效率及地上生产力的影响. 应用生态学报, 2018, 29(6): 1822-1828. |
| [38] |
Connell S D, Ghedini G. Resisting regime-shifts: the stabilising effect of compensatory processes. Trends in Ecology & Evolution, 2015, 30(9): 513-515. |
| [39] |
Liu H Y, Mi Z R, Lin L, Wang Y H, Zhang Z H, Zhang F W, Wang H, Liu L L, Zhu B, Cao G M, Zhao X Q, Sanders N J, Classen A T, Reich P B, He J S. Shifting plant species composition in response to climate change stabilizes grassland primary production. Proceedings of the National Academy of Sciences of the United States of America, 2018, 115(16): 4051-4056. DOI:10.1073/pnas.1700299114 |
| [40] |
李长慧, 李淑娟, 雷有升, 胡凯, 王佩羽, 孙启忠. 4种高原乡土禾草的抗旱生理比较. 草业科学, 2013, 30(9): 1386-1393. |
| [41] |
吴建国, 周巧富. 中国嵩草属植物地理分布模式和适应的气候特征. 植物生态学报, 2012, 36(3): 199-221. |
| [42] |
Huo L L, Chen Z K, Zou Y C, Lu X G, Guo J W, Tang X G. Effect of Zoige alpine wetland degradation on the density and fractions of soil organic carbon. Ecological Engineering, 2013, 51: 287-295. DOI:10.1016/j.ecoleng.2012.12.020 |
| [43] |
肖春旺, 张新时. 模拟降水量变化对毛乌素油蒿幼苗生理生态过程的影响研究. 林业科学, 2001, 37(1): 15-22. |
| [44] |
姬兰柱, 肖冬梅, 王淼. 模拟水分胁迫对水曲柳光合速率及水分利用效率的影响. 应用生态学报, 2005, 16(3): 408-412. DOI:10.3321/j.issn:1001-9332.2005.03.003 |
| [45] |
张仁和, 郑友军, 马国胜, 张兴华, 路海东, 史俊通, 薛吉全. 干旱胁迫对玉米苗期叶片光合作用和保护酶的影响. 生态学报, 2011, 31(5): 1303-1311. |
| [46] |
Eissenstat D M, Yanai R D. The ecology of root lifespan. Advances in Ecological Research, 1997, 27: 1-60. |
| [47] |
Zhang F Y, Quan Q, Song B, Sun J, Chen Y J, Zhou Q P, Niu S L. Net primary productivity and its partitioning in response to precipitation gradient in an alpine meadow. Scientific Reports, 2017, 7(1): 15193. DOI:10.1038/s41598-017-15580-6 |
| [48] |
顾振宽, 杜国祯, 朱炜歆, 索南吉, 张世虎. 青藏高原东部不同草地类型土壤养分的分布规律. 草业科学, 2012, 29(4): 507-512. |
| [49] |
Liu C, Xiang W H, Lei P F, Deng X W, Tian D L, Fang X, Peng C H. Standing fine root mass and production in four Chinese subtropical forests along a succession and species diversity gradient. Plant and Soil, 2014, 376(1/2): 445-459. |
| [50] |
王根绪, 胡宏昌, 王一博, 陈琳. 青藏高原多年冻土区典型高寒草地生物量对气候变化的响应. 冰川冻土, 2007, 29(5): 671-679. DOI:10.3969/j.issn.1000-0240.2007.05.001 |
| [51] |
陆姣云, 段兵红, 杨梅, 杨晗, 杨惠敏. 植物叶片氮磷养分重吸收规律及其调控机制研究进展. 草业学报, 2018, 27(4): 178-188. |
| [52] |
Cormier N, Twilley R R, Ewel K C, Krauss K W. Fine root productivity varies along nitrogen and phosphorus gradients in high-rainfall mangrove forests of Micronesia. Hydrobiologia, 2015, 750(1): 69-87. DOI:10.1007/s10750-015-2178-4 |
| [53] |
Pregitzer K S. Tree root architecture-form and function. New Phytologist, 2008, 180(3): 562-564. DOI:10.1111/j.1469-8137.2008.02648.x |
| [54] |
徐文静, 王政权, 范志强, 孙海龙, 贾淑霞, 吴楚. 遮荫对水曲柳幼苗细根衰老的影响. 植物生态学报, 2006, 30(1): 104-111. DOI:10.3321/j.issn:1005-264X.2006.01.015 |
| [55] |
裴智琴, 周勇, 郑元润, 肖春旺. 干旱区琵琶柴群落细根周转对土壤有机碳循环的贡献. 植物生态学报, 2011, 35(11): 1182-1191. |
| [56] |
Hodge A. The plastic plant: root responses to heterogeneous supplies of nutrients. New Phytologist, 2004, 162(1): 9-24. DOI:10.1111/j.1469-8137.2004.01015.x |
| [57] |
白文明, 左强, 黄元仿, 李保国. 乌兰布和沙区紫花苜蓿根系生长及吸水规律的研究. 植物生态学报, 2001, 25(1): 35-41. |
| [58] |
Han S H, Kim S, Li G L, Chang H N, Yun S J, An J, Son Y. Effects of warming and precipitation manipulation on fine root dynamics of Pinus densiflora Sieb. et Zucc. Seedlings. Forests, 2018, 9(1): 14. |
| [59] |
Pregitzer K S, Zak D R, Curtis P S, Kubiske M E, Teeri J A, Vogel C S. Atmospheric CO2, soil nitrogen and turnover of fine roots. New Phytologist, 1995, 129(4): 579-585. DOI:10.1111/j.1469-8137.1995.tb03025.x |
| [60] |
An Y Y, Liang Z S, Zhao R K, Zhang J, Wang X J. Organ-dependent responses of Periploca sepium to repeated dehydration and rehydration. South African Journal of Botany, 2011, 77(2): 446-454. DOI:10.1016/j.sajb.2010.11.003 |
| [61] |
邱俊, 谷加存, 姜红英, 王政权. 樟子松人工林细根寿命估计及影响因子研究. 植物生态学报, 2010, 34(9): 1066-1074. DOI:10.3773/j.issn.1005-264x.2010.09.007 |
| [62] |
Vogt K A, Grier C C, Vogt D J. Production, turnover, and nutrient dynamics of above-and belowground detritus of world forests. Advances in Ecological Research, 1986, 15: 303-377. |
| [63] |
Gill R A, Jackson R B. Global patterns of root turnover for terrestrial ecosystems. New Phytologist, 2000, 147(1): 13-31. DOI:10.1046/j.1469-8137.2000.00681.x |
| [64] |
Yuan Z Y, Chen H Y H. Fine root biomass, production, turnover rates, and nutrient contents in boreal forest ecosystems in relation to species, climate, fertility, and stand age: literature review and meta-analyses. Critical Reviews in Plant Sciences, 2010, 29(4): 204-221. DOI:10.1080/07352689.2010.483579 |
| [65] |
字洪标, 陈焱, 胡雷, 王长庭. 氮肥添加对川西北高寒草甸植物群落根系动态的影响. 植物生态学报, 2018, 42(1): 38-49. |
| [66] |
戴黎聪, 柯浔, 曹莹芳, 张法伟, 杜岩功, 李以康, 郭小伟, 李茜, 林丽, 马建军, 曹广民. 青藏高原矮嵩草草甸地下和地上生物量分配格局及其与气象因子的关系. 生态学报, 2019, 39(2): 486-493. |
| [67] |
林丽, 张德罡, 曹广民, 欧阳经政, 柯浔, 刘淑丽, 张法伟, 李以康, 郭小伟. 放牧强度对高寒嵩草草甸土壤养分特性的影响. 生态学报, 2016, 36(15): 4664-4671. |
| [68] |
Zhang X, Xing Y J, Yan G Y, Han S J, Wang Q G. Effects of precipitation change on fine root morphology and dynamics at a global scale: a meta-analysis. Canadian Journal of Soil Science, 2019, 99(1): 1-11. DOI:10.1139/cjss-2018-0114 |