 2022, Vol. 42
2022, Vol. 42文章信息
- 孙龙, 岳阳, 胡同欣
- SUN Long, YUE Yang, HU Tongxin
- 干扰对生态系统多功能性的影响研究进展
- Research progress on the effects of disturbance on ecosystem multifunctionality
- 生态学报. 2022, 42(15): 6066-6075
- Acta Ecologica Sinica. 2022, 42(15): 6066-6075
- http://dx.doi.org/10.5846/stxb202104231072
-
文章历史
- 收稿日期: 2021-04-23
- 网络出版日期: 2022-04-07

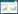




生态系统多功能性(ecosystem multifunctionality, EMF)是指生态系统同时行使多个生态系统功能的能力或性质[1], 是评价生态系统多个功能和性质的一个综合指标, 也是全面认识生态系统功能与结构的有效途径[2], 生态系统功能即为生态系统的过程或性质, 其中生态系统过程包括养分循环、初级生产及分解作用等, 另一方面, 生态系统功能是生态系统本身所具备的一种基本属性, 如其物理结构、生态系统稳定性、恢复力等[3—4]。目前关于EMF的研究越来越注重生态系统功能和生态系统服务, 2018年Manning等[5]在Nature上提出, EMF可以归纳为两个独立的研究领域, 即生态系统功能多功能性及生态系统服务多功能性[5], 2021年井新等[6]通过分析生态系统功能和服务的特点, 首次提出了生态系统多服务性的概念。生态系统并不是以孤立形式存在的, 它通过与周围的水、空气、生物体等物质与外界环境变化做出反应, 因此在对生态系统进行评价时, 需考虑多个生态系统功能或服务。Hölting等[7]通过对“多功能性”进行文献检索发现, 所研究的507篇文献主要集中于农业管理、景观规划、生态和土壤科学、城乡发展、林业五组学科研究领域中, 其中在对EMF进行量化的文章中属于“生态学与土壤学科”领域最多(49%)。
干扰作为能够显著改变系统正常格局的一种事件[8], 通常分为自然干扰和人为干扰两种, 诸如火灾、病虫害、飓风等为自然干扰, 放牧、营养元素添加、城市化等为人为干扰。由于干扰活动在连续空间内极易传播, 如火灾、植物病虫害等干扰活动更容易在抵抗力弱且连续性强的植被中蔓延, 因此, 对于某些斑块化明显或由多种斑块组成的景观, 干扰程度会有所减弱。干扰通过改变群落结构、组成和功能, 改变物理环境等方式破坏生态系统平衡[9]。例如, 东北次生林生态系统中线虫群落的多样性随干扰度的增加而减少[6, 10], 生态系统功能变化的主要原因侧重于由干扰所导致的生物多样性丧失[11]。同时, 干扰被当作一种管理工具来实现土地管理者的目的, 近年来由于人为干扰的频率、范围和强度不断增大, 生态系统的生物多样性和功能所面临的威胁也在不断加剧[6]。
到目前为止, 关于EMF的测度方法、度量指标还未达到共识, 评估EMF的研究很少考虑研究过程中所处环境受到扰动事件的后果, 以及因此导致的群落结构和环境变化。本文通过查阅大量相关文献, 总结分析EMF的测度方法、影响因素以及干扰对EMF的影响, 通过探讨目前各研究进展, 提出具有针对性的展望, 以期为今后该领域的研究提供借鉴, 推动该领域的发展。
1 生态系统多功能性概念研究进展多样性对生态系统过程(或功能)的潜在影响最早可以追溯到达尔文时期[12]。2004年Sanderson等[13]在Crop Science上首次提出生态系统是由几个生态系统服务或过程来管理或评估的, 即生态系统“多功能”。2007年Hector和Bagchi等[12]在Nature上提出EMF是生态系统的多种服务或过程, 同时将EMF进行量化。2012年Maestre等[14]首次对全球干旱地区植物物种丰富度与EMF之间的关系进行评估。
通过对国际数据库ISI Web of Science进行主题(Topic)为ecosystem multifunctionality, 检索时间为2001—2020年共计20年的文献检索, 数据库设置为“核心合集”, 检索到的文献为735篇, 剔除无关文献后为654篇(文献类型仅保留article)。在此基础上, 根据对ISI Web of Science数据库进行以多个不同生态系统类型为主题词的搜索(2001—2020年, 图 1), 共计493篇, 其中森林生态系统(forest ecosystem, 36.8%)>草原生态系统(grassland ecosystem, 27.2%)>农田生态系统(field ecosystem, 22.8%), 三者体现了目前EMF研究在不同生态系统中的关注焦点。在近10年的研究中, 对于城市生态系统(urban ecosystem, 17.2%)进行的研究明显增加, 可能是由于生态系统服务的提出, 对于景观生态系统的关注度上升, 增强了研究人员对城市EMF的重视。目前, 针对EMF的部分研究成果如表 1。

|
| 图 1 以不同生态系统类型为主题关键词的ISI Web of Science文献检索结果(2001—2020) Fig. 1 The search result for various ecosystem types as key words of references collected in the ISI Web of Science(2001—2020) 文献数量小于5%时隐藏标签 |
| 研究区域 Study area |
结论 Conclusion |
参考文献 References |
| 森林生态系统 Forest ecosystem |
云南松天然次生林中物种丰富度对多功能性的影响最大, 表现为极显著正相关 | [15] |
| 总体多样性效应与多功能性呈正相关, 功能丰富度对单功能影响比对多功能性的影响更大 | [16] | |
| 农田生态系统 Field ecosystem |
作物行间距模式对多功能性有直接的正效应 | [17] |
| 草原生态系统 Grassland ecosystem |
牲畜多样化可以促进草原生物多样性和生态系统多功能性 | [18] |
| 地上和地下生物多样性能够更有效地预测放牧对生态系统多功能性的影响 | [19] | |
| 城市生态系统 Urban ecosystem |
城市化水平阻碍了生态系统服务功能 | [20] |
| 湿地生态系统 Wetlands ecosystem |
物种栖息地同质化可能导致湿地提供的功能减少 | [21] |
| 在全球増温能力降低的背景下由加性多功能指数法(AMF)表征的生态系统多功能性指数随物种丰富度升高而增强 | [1] | |
| 旱地生态系统 Arid ecosystems |
生态系统多功能性与物种丰富度有积极的正效应 | [14] |
迄今为止, 许多研究者们试图选取不同类型的生态系统功能来代表整个EMF。然而, 由于生态系统功能和服务种类繁多, 且存在相互作用[5], 因此Grace等[22]选择通过将数据与理论结合, 运用结构方程建模来对EMF进行定性分析。EMF量化指标的选取还会因所研究生态系统类型的不同而有差异, 选取指标时还应充分考虑不同生态系统类型中影响生态系统功能的不同量化指标。考虑的功能越多, 结果就越接近整个EMF的真实值。目前针对EMF的量化指标主要集中于土壤多功能性(C、N、P循环)、土壤含水率、pH值、植物元素含量(叶片、根系)、生物多样性以及土壤酶活性等方面。
由于生态系统服务与人类生活密切相关, 故在进行生态系统服务研究时, 首先应明确研究所选的具体生态系统服务指标。Jochem等[23]在研究德国整体森林区域时提出, 应重点关注木材生产(供应)、森林蓄积量和林木产品的碳固存(调节)以及娱乐(文化)这三种生态系统服务。Jönsson等[24]在计算生态系统服务时将树木生物量生产、土壤表层碳储量以及与落叶树种的数量分别作为生态系统供应服务调节作用和文化支持服务的测度指标。目前针对森林生态系统服务的研究主要放在木材生产力这一森林服务功能上, 对于其他生态系统服务功能研究甚少[24]。
2.2 生态系统多功能性的测度方法由于EMF具有一定的复杂性, 且量化方法众多, 使其至今还没有确定的量化方法, 并且不同研究之间难以进行比较, 限制其进一步发展[25]。针对目前量化EMF的方法原理及优缺点进行分析(表 2), 其中使用最多的是平均值法和阈值法。
| 方法 Method |
原理 Theory |
优点 Advantage |
缺点 Disadvantage |
| 单功能法 Single-approach[26] |
将所研究的功能指标与生物多样性进行线性回归分析, 根据结果判断生态系统功能和生物多样性之间的关系[26] | 计算简便, 有助于说明EMF变化的具体驱动因子及生物多样性对EMF的影响[25] | 易忽略多个生态因子间的相互作用, 无法对EMF进行定性分析[27], 会受到功能性指标的影响 |
| 功能—物种替代法(也称重叠法) Turnover approach[12] |
通过最小AIC(Akaike Information Criterion)标准确定对生态系统功能影响最大的物种组成, 通过每个样点上的物种重叠率计算影响生态系统功能所有可能的物种组成 | 可以准确识别出对每种生态系统功能有影响的具体物种组成, 并测试这些物种组合在不同生态系统功能之间是否有差异 | 不能对其进行量化, 无法得知物种丰富度对EMF具体的影响程度[27], 忽略了功能对EMF的负效应, 数据量大[25], 也可能会出现两个极端情况[12, 25] |
| 平均值法 Averaging approach[28] |
将每个功能的测定值进行转化, 然后将每个功能的值进行标准化, 计算每个功能的平均值[25—26, 29] | 可以测定多个生态系统功能 | 适用于样方面积过大、植被覆盖率较低时[14], 默认所有功能对于EMF的贡献度都相同 |
| 单阈值法 Single threshold approach[30] |
计算每个生态系统中达到某一阈值的功能数来求得一个指数, 该指数表示在该阈值条件下这个生态系统整体功能的水平[25, 31] | 选择空间大, 应用范围广[25] | 没有确定的标准阈值, 计算的生态系统功能值会随阈值的选择而变化[27], 容易受到研究者的主观态度影响[25] |
| 多阈值法 Multiple-threshold approach[27] |
计算多功能性指数的方法以及阈值的确定方法均与单阈值法相同, 但多阈值法包含了0—100%之间的所有阈值, 对每个阈值都计算了相应的多功能数[27] | 能够定量的评估EMF的性质和范围, 且计算简便, 应用范围广 | 将所有功能的作用都视为等效, 没有反映特定生态系统功能或服务的重要性, 结果会因阈值方法的不同而改变[25, 27] |
| 多元模型法 Multivariate model approach[32] |
以多样性交互模型[33]为基础, 将一个生态系统功能扩展到一个多变量框架中, 并且同时模拟生物多样性与EMF之间的关系[32] | 同时评估多功能性的多种驱动因素, 可以测试驱动因素在生态系统功能之间的相对重要性, 合并生态系统功能之间的相关性, 检验整体功能在何时达到最大值[32] | 目前还没有全面的多功能度量, 计算过程复杂, 数据量大且难以全部获得, 很难实现广泛应用 |
Byrnes等[27]于2014年在对现有研究生物多样性与EMF的关系中提出, 阈值法提供了最全面和最有效的方法, 建议将其应用于未来的研究。
2.3 生态系统多功能性的影响因素生物多样性[34]是指地球上动物、植物、微生物及其与环境形成的生态复合体, 以及与此相关的各种生态过程, 是生态系统功能最主要的驱动因素[14], 生物多样性随多功能性指数的增加而增加[6, 16], 同时也提高了生态系统功能的稳定性[35]。由全球变化引起的生物多样性丧失虽对EMF产生了消极影响[14, 36], 但并未改变对生态系统功能的影响[37]。Gamfeldt等[30]通过模拟物种丰富度和整体生态系统功能之间的关系得出, 物种丰富度可能会通过水分和养分动态、营养物质相互作用或干扰机制的变化, 影响生态系统的结构和功能, 进而影响生态系统的整体功能。生物多样性对不同生态系统功能的影响随着资源分布空间异质性的增加而增加[38], 如Byrnes等[27]在对横跨欧洲大陆的8个草原生态系统进行的平行实验中发现, 物种丰富度对地上生物量和植物总氮影响明显。此外, 井新等[6]人从生物多样性维度、时空依赖性和驱动因子多样化三个角度分别论述了与EMF的特点, 王凯等[39]在研究生物多样性与EMF的关系时提出, 干扰会直接或间接地影响生物多样性与EMF的关系。但是目前关于生物多样性与生态系统功能之间关系的研究主要集中在植物群落上, 对于地表和地下生物多样性对生态系统多功能影响的研究还很单薄。
植物多样性在维持生态系统功能和调解气候变化对生态系统功能的影响中发挥着至关重要的作用[40]。Wang等[19]通过研究中度放牧处理后EMF与植物多样性的关系指出, EMF与植物多样性呈正相关, 当植被覆盖率低于25%时, 多功能性崩溃, 对应的生态系统功能减少。此外, 树木多样性可影响不同的生态系统服务功能[36]。植物功能多样性是解释旱地生态系统EMF的关键因素之一[41], 功能多样性也可通过改善微生物环境条件和增加凋落物来对EMF产生正相关效应[42]。
土壤微生物[43]是土壤中一切肉眼看不见或看不清楚的微小生物的总称, 严格意义上应包括细菌、古菌、真菌、病毒、原生动物和显微藻类。同时, 在全球变暖的背景下, 生态系统通过减少水分来抑制微生物活性、养分循环、植物生长以及植被覆盖度[44]。Ma等[45]首次研究了不同剂量氮、磷添加对参与土壤氮循环的9种主要微生物群落的影响, Bell等[46]通过研究细菌群落对生物多样性的影响得出, 细菌多样性与群落呼吸作用呈负相关, 且不同种类的细菌之间存在的协同作用和细菌群落的组成在决定生态系统功能水平方面具有重要意义。He等[47]通过研究得出, 随着土壤细菌多样性的丧失, 生态系统功能数量也将减少。此外, 土壤动物也对EMF有影响, 例如蚯蚓是通过影响土壤群落中的细菌群落间接增强EMF[48]。
除生物因素外, 土地管理方式和其他非生物因素也会对EMF产生影响[6]。受人类活动影响, 土地利用变化会不同程度的扩大土地利用面积或减少农业用地, 致使这些农业生态系统的生态系统功能退化乃至消失。Lindborg等[49]指出, 生态系统服务随当地物质和社会经济条件的变化而不同, 很可能受到与其利益相关者对当地环境以及管理土地方式决策的影响。Allan等[6, 50]在150个农业及草原生态系统中证明土地利用方式是通过生物多样性丧失和生态系统功能构成的变化影响生态系统服务。Xue等[51]研究得出不同土地利用方式之间通过影响土壤细菌多样性影响EMF的结论。随着世界人口及资源消耗的增加, 土地资源逐渐匮乏, 土地利用规模改变, 农业用地面积增加, 但由此产生的景观异质性也可以促进景观尺度生态系统的多功能性[52]。Zirbel[29]通过比较多个草原多样性得出, 由景观异质性造成多功能性变化的影响几乎是功能多样性的四倍。
3 干扰对生态系统多功能性的影响 3.1 自然干扰全球气候变化是地球生态系统中一个巨大而持续的干扰活动[53], 气候变暖通过直接影响生态系统的净初级生产力(NPP)、有机物产量、溶解营养物质等直接影响生态系统功能, 从而增大EMF[54]。气候变暖又能够降低草原生态系统的物种丰富度、生物量和土壤有机物含量[55]。预计未来几十年在气候变暖的背景下, 火灾和病虫害对森林生态系统的干扰可能会增加[9]。
极端气候事件包括干旱、洪水、热浪、霜冻和飓风等[56—57], 现已成为描述全球气候变化的主要特征[58], 极端气候事件发生频率的增加, 对陆地生态系统服务功能和人类社会生产生活造成了严重影响[59]。极端干旱通过高温抑制植物叶片的蒸散发, 降低植物的光合作用和呼吸作用, 进而减少植物的生物量[60], 干旱[61]又可以通过微生物群落的结构和功能影响生态系统, 极端降水使得水分向土壤更深层次渗透, 有效降低土壤蒸散发损失, 延长土壤有效水的供给时间[62—63], 极端的热浪事件正在造成整个海洋生态系统的生态系统功能退化[64]。
火干扰作为森林生态系统碳循环的重要影响因子, 改变了整个森林生态系统的格局与过程, 对全球的碳循环产生了重要影响[65—66], 火烧后土壤中除Cu和Co外的主要元素和微量元素含量均高于未火烧后的土壤[67]。通过陆地植物群落的结构和功能影响全球生物群落分布[68]。火灾可以改变生态系统的养分循环与能量传递[69—70], 通过植物群落的变化, 改变土壤物理和化学环境, 并最终改变土壤微生物群落[71]。在草原生态系统中发生的火灾可以导致植物物种丰富度和多样性在短期内增加[72]。火干扰对生态系统的影响还取决于火干扰的强度和频率, 对于某些低频率低强度的区域, 随着时间的推移, 会形成在空间上连续的、富含可燃物的环境, 一旦发生火灾, 危险性极大。
此外, 外来害虫和病原体的入侵也能够对生态系统造成威胁[6, 73], 遭遇病虫害的森林营养缺乏, 植被稀疏, 透光性强, 极易受到飓风等外界干扰, 从而增加可燃物载量。酸雨对森林土壤、植物及生态系统各层面中的钙离子均造成了长期广泛的影响[74]。随着自然干扰对生态系统造成的威胁日益严峻, 且相关研究甚少。因此, 对生态系统进行具有针对性的干扰研究具有重大意义。
3.2 人为干扰放牧能不同程度地削弱多项生态系统服务及功能间的权衡关系[6, 75], 放牧通过破坏草地植被光合组织, 降低草地生态系统碳交换的能力, 减弱草地生态系统碳汇的功能[76]。不同程度的放牧试验表明, 适度放牧有利于草地生态系统的多功能性, 可以改善养分循环和土壤碳固存[19], 过度放牧主要通过减少植物物种多样性降低其生产力, 破坏生物多样性并降低生态系统中的氮循环[77], 禁牧更有利于其生态系统有机碳、氮储量积累[78]。
氮(N)和磷(P)是陆地生态系统中的两个主要的营养元素, 不同水平N、P元素的添加会对生态系统功能产生重要影响[79]。通过在青藏高原高寒草甸进行的氮、磷添加实验得出, 氮、磷添加可缓解植物生长的营养限制, 促进植物地上部分的生长[80], 氮添加能够显著降低土壤pH、全磷以及碱性磷酸酶的活性[81]。张秀兰等[82]在亚热带地区的一项研究表明, 森林土壤碳的稳定性主要受到磷含量的调控, 短期磷添加易导致表层土壤活性有机碳分解, 增加土壤碳稳定性。
Zhang等[6, 83]通过研究农耕对土壤多功能性的影响得出, 免耕可以通过控制微生物群落减轻干旱对土壤功能的影响, 改变生态系统稳定性, 提高EMF。Castioni等[84]通过研究草场清理对土壤多功能性的影响得出, 草场的全部去除和高度去除导致土壤有机碳减少, 物理性能退化。然而, 当人口持续增加, 土地资源更加稀缺, 土地利用率高, 生态系统退化。
徐煖银等[85]在赣南地区的研究表明生态系统服务价值和人为干扰度值呈现出极强的负相关关系。相对于自然干扰而言, 人为干扰更容易控制, 适度的人为干扰有助于生态系统的稳定。
4 结论与展望生态系统的价值在于它能够同时提供多种生态功能。目前对EMF的研究因出发点不同, 发展较快[86]。国外Gamfeldt等[87]于2008年对生物多样性对水生生态系统的影响做了较为详细的综述。国内徐炜等[6, 25, 88]于2016年对生物多样性与EMF的研究进展做了详细的介绍, 测度方法主要针对的是生物多样性和EMF的关系, 即生态系统功能多功能性, 张宏锦等[89]人通过对相关文献进行计量分析, 系统归纳了此领域的发展现状。但就目前EMF的应用情况而言, 仍存在诸多问题。目前对于EMF的测度方法较为复杂, 仅依据经验确定计算方法;针对EMF具体的测度因子还未确定, 会对实验结果产生影响;另外, 跨时间尺度较长的实验易受到其他外界影响。因此, 为加强我国EMF的研究, 今后研究应从以下几个方面开展:
(1) 加强生态系统多功能性的应用性研究。与农业和草原生态系统相比, 对森林生态系统等复杂的动态系统的研究较少[6, 16], 关于森林生态系统中生物多样性与各生态系统功能之间关系的研究仍然匮乏[90]。研究发现, 在大面积的空间范围内(欧洲和全世界的草原), 环境变化对多功能性的影响有时比生物多样性大[29]。目前关于EMF的研究还存在概念不清、方法不明确、测度指标不定、针对性不强的特点。今后应加强对不同生态系统类型、不同空间尺度EMF具有针对性的研究。
(2) 加强评估各生态系统功能之间的相互作用对生态系统多功能性的影响研究。在进行对生态系统评估时, 若仅从某单一功能指标推断其对生态系统的作用, 不仅忽略了生态系统的其他功能, 还忽略了各生态系统功能间的相互作用, 实际上并未对生态系统的响应做出综合评价。假设各功能影响相同, 确定具体相关指标[5, 91]。由于物种间多种功能之间的协同作用, 生态系统的整体功能比单一功能更容易受到物种损失的影响[6, 30]。对EMF的评估不仅只考虑影响多功能性的多个功能和系统的研究上, 还应评估它们之间的相互作用对EMF的影响。
(3) 加强干扰对生态系统多功能性影响的机制性研究。外界干扰具有一定的复杂性, 在生态系统受到影响时, 生态系统会通过一个或几个功能或服务对其做出响应。随着EMF概念的引入以及深入研究, 已有部分文献针对干扰对EMF的影响机制进行探索, 如气候变暖、放牧、元素添加等[19, 54, 79], 但由于生态系统的复杂性难以概括全部生态系统影响机制, 还需对干扰过后生态系统产生的影响进行全面综合性的评价, 明确具体干扰对EMF的影响机制, 量化其影响程度, 从而在干扰过后能够做出及时有效的应变措施。因此, 应结合干扰对生态系统的影响机制研究, 来为今后提出行之有效的稳定生态系统的建议。
| [1] |
刘阳. 人工湿地中植物多样性与生态系统多功能性. 杭州: 浙江大学, 2015.
|
| [2] |
Jing X, Sanders N J, Shi Y, Chu H Y, Classen A T, Zhao K, Chen L T, Shi Y, Jiang Y X, He J S. The links between ecosystem multifunctionality and above- and belowground biodiversity are mediated by climate. Nature Communications, 2015, 6: 8159. DOI:10.1038/ncomms9159 |
| [3] |
吕亭亭. 草本植物群落功能多样性与生态系统功能关系研究. 长春: 东北师范大学, 2014.
|
| [4] |
冯剑丰, 李宇, 朱琳. 生态系统功能与生态系统服务的概念辨析. 生态环境学报, 2009, 18(4): 1599-1603. |
| [5] |
Manning P, van der Plas F, Soliveres S, Allan E, Maestre F T, Mace G, Whittingham M J, Fischer M. Redefining ecosystem multifunctionality. Nature Ecology & Evolution, 2018, 2(3): 427-436. |
| [6] |
井新, 贺金生. 生物多样性与生态系统多功能性和多服务性的关系: 回顾与展望. 植物生态学报, 2021, 45: 1-18. |
| [7] |
Hölting L, Beckmann M, Volk M, Cord A F. Multifunctionality assessments-More than assessing multiple ecosystem functions and services? A quantitative literature review. Ecological Indicators, 2019, 103: 226-235. DOI:10.1016/j.ecolind.2019.04.009 |
| [8] |
王广慧, 乌兰, 于军. 干扰与生态系统的关系. 内蒙古草业, 2007, 19(1): 15-18. |
| [9] |
Seidl R, Thom D, Kautz M, Martin-Benito D, Peltoniemi M, Vacchiano G, Wild J, Ascoli D, Petr M, Honkaniemi J, Lexer M J, Trotsiuk V, Mairota P, Svoboda M, Fabrika M, Nagel T A, Reyer C P O. Forest disturbances under climate change. Nature Climate Change, 2017, 7: 395-402. DOI:10.1038/nclimate3303 |
| [10] |
焦向丽, 朱教君, 张金鑫, 闫巧玲, 杨凯. 干扰对东北次生林生态系统土壤线虫群落的影响. 生态学杂志, 2008, 27(12): 2129-2135. |
| [11] |
Villnäs A, Norkko J, Hietanen S, Josefson A B, Lukkari K, Norkko A. The role of recurrent disturbances for ecosystem multifunctionality. Ecology, 2013, 94(10): 2275-2287. DOI:10.1890/12-1716.1 |
| [12] |
Hector A, Bagchi R. Biodiversity and ecosystem multifunctionality. Nature, 2007, 448(7150): 188-190. DOI:10.1038/nature05947 |
| [13] |
Sanderson M A, Skinner R H, Barker D J, Edwards G R, Tracy B F, Wedin D A. Plant species diversity and management of temperate forage and grazing land ecosystems. Crop Science, 2004, 44(4): 1132-1144. DOI:10.2135/cropsci2004.1132 |
| [14] |
Maestre F T, Quero J L, Gotelli N J, Escudero A, Ochoa V, Delgado-Baquerizo M, García-Gómez M, Bowker M A, Soliveres S, Escolar C, García-Palacios P, Berdugo M, Valencia E, Gozalo B, Gallardo A, Aguilera L, Arredondo T, Blones J, Boeken B, Bran D, Conceição A A, Cabrera O, Chaieb M, Derak M, Eldridge D J, Espinosa C I, Florentino A, Gaitán J, Gatica M G, Ghiloufi W, Gómez-González S, Gutiérrez J R, Hernández R M, Huang X W, Huber-Sannwald E, Jankju M, Miriti M, Monerris J, Mau R L, Morici E, Naseri K, Ospina A, Polo V, Prina A, Pucheta E, Ramírez-Collantes D A, Romão R, Tighe M, Torres-Díaz C, Val J, Veiga J P, Wang D L, Zaady E. Plant species richness and ecosystem multifunctionality in global drylands. Science, 2012, 335(6065): 214-218. DOI:10.1126/science.1215442 |
| [15] |
黄小波, 李帅锋, 苏建荣, 刘万德, 郎学东. 云南松天然次生林物种丰富度与生态系统多功能性的关系. 生物多样性, 2017, 25(11): 1182-1191. |
| [16] |
Mensah S, Salako K V, Assogbadjo A, Glèlè Kakaï R, Sinsin B, Seifert T. Functional trait diversity is a stronger predictor of multifunctionality than dominance: evidence from an Afromontane forest in South Africa. Ecological Indicators, 2020, 115: 106415. DOI:10.1016/j.ecolind.2020.106415 |
| [17] |
Zhang G Z, Kou X C, Zhang X K, Bai W, Liang W J. Effect of row spacings on soil nematode communities and ecosystem multifunctionality at an aggregate scale. Scientific Reports, 2020, 10(1): 4779. DOI:10.1038/s41598-020-61498-x |
| [18] |
Wang L, Delgado-Baquerizo M, Wang D L, Isbell F, Liu J, Feng C, Liu J S, Zhong Z W, Zhu H, Yuan X, Chang Q, Liu C. Diversifying livestock promotes multidiversity and multifunctionality in managed grasslands. Proceedings of the National Academy of Sciences of the United States of America, 2019, 116(13): 6187-6192. DOI:10.1073/pnas.1807354116 |
| [19] |
Wang X Y, Li F Y, Wang Y N, Liu X M, Cheng J W, Zhang J Z, Baoyin T, Bardgett R D. High ecosystem multifunctionality under moderate grazing is associated with high plant but low bacterial diversity in a semi-arid steppe grassland. Plant and Soil, 2020, 448(1/2): 265-276. |
| [20] |
Kim I, Kwon H, Kim S, Jun B. Identification of landscape multifunctionality along urban-rural gradient of coastal cities in South Korea. Urban Ecosystems, 2020, 23(5): 1153-1163. DOI:10.1007/s11252-020-00993-5 |
| [21] |
Pendleton M C, Sedgwick S, Kettenring K M, Atwood T B. Ecosystem functioning of great salt lake wetlands. Wetlands, 2020, 40(6): 2163-2177. DOI:10.1007/s13157-020-01333-1 |
| [22] |
Grace J B, Anderson T M, Olff H, Scheiner S M. On the specification of structural equation models for ecological systems. Ecological Monographs, 2010, 80(1): 67-87. DOI:10.1890/09-0464.1 |
| [23] |
Bösch M, Jochem D, Weimar H, Dieter M. Physical input-output accounting of the wood and paper flow in Germany. Resources, Conservation and Recycling, 2015, 94: 99-109. DOI:10.1016/j.resconrec.2014.11.014 |
| [24] |
Jönsson M, Snäll T. Ecosystem service multifunctionality of low-productivity forests and implications for conservation and management. Journal of Applied Ecology, 2020, 57(4): 695-706. DOI:10.1111/1365-2664.13569 |
| [25] |
徐炜, 井新, 马志远, 贺金生. 生态系统多功能性的测度方法. 生物多样性, 2016, 24(01): 72-84. |
| [26] |
Duffy J E, Richardson J P, Canuel E A. Grazer diversity effects on ecosystem functioning in seagrass beds. Ecology Letters, 2003, 6(7): 637-645. DOI:10.1046/j.1461-0248.2003.00474.x |
| [27] |
Byrnes J E K, Gamfeldt L, Isbell F, Lefcheck J S, Griffin J N, Hector A, Cardinale B J, Hooper D U, Dee L E, Duffy J E. Investigating the relationship between biodiversity and ecosystem multifunctionality: challenges and solutions. Methods in Ecology and Evolution, 2014, 5(2): 111-124. DOI:10.1111/2041-210X.12143 |
| [28] |
Hooper D U, Vitousek P M. Effects of plant composition and diversity on nutrient cycling. Ecological Monographs, 1998, 68(1): 121-149. DOI:10.1890/0012-9615(1998)068[0121:EOPCAD]2.0.CO;2 |
| [29] |
Zirbel C R, Grman E, Bassett T, Brudvig L A. Landscape context explains ecosystem multifunctionality in restored grasslands better than plant diversity. Ecology, 2019, 100(4): e02634. |
| [30] |
Gamfeldt L, Hillebrand H, Jonsson P R. Multiple functions increase the importance of biodiversity for overall ecosystem functioning. Ecology, 2008, 89(5): 1223-1231. DOI:10.1890/06-2091.1 |
| [31] |
Zavaleta E S, Pasari J R, Hulvey K B, Tilman G D. Sustaining multiple ecosystem functions in grassland communities requires higher biodiversity. Proceedings of the National Academy of Sciences of the United States of America, 2010, 107(4): 1443-1446. DOI:10.1073/pnas.0906829107 |
| [32] |
Dooley Á, Isbell F, Kirwan L, Connolly J, Finn J A, Brophy C. Testing the effects of diversity on ecosystem multifunctionality using a multivariate model. Ecology Letters, 2015, 18(11): 1242-1251. DOI:10.1111/ele.12504 |
| [33] |
Kirwan L, Connolly J, Finn J A, Brophy C, Lüscher A, Nyfeler D, Sebastià M T. Diversity-interaction modeling: estimating contributions of species identities and interactions to ecosystem function. Ecology, 2009, 90(8): 2032-2038. DOI:10.1890/08-1684.1 |
| [34] |
Mori A S, Lertzman K P, Gustafsson L. Biodiversity and ecosystem services in forest ecosystems: a research agenda for applied forest ecology. Journal of Applied Ecology, 2017, 51(1): 12-27. |
| [35] |
Cardinale B J, Duffy J E, Gonzalez A, Hooper D U, Perrings C, Venail P, Narwani A, Mace G M, Tilman D, Wardle D A, Kinzig A P, Daily G C, Loreau M, Grace J B, Larigauderie A, Srivastava D S, Naeem S. Biodiversity loss and its impact on humanity. Nature, 2012, 489(7401): 59-67. |
| [36] |
Xie H T, Wang G G, Yu M K. Ecosystem multifunctionality is highly related to the shelterbelt structure and plant species diversity in mixed shelterbelts of eastern China. Global Ecology and Conservation, 2018, 16: e00470. DOI:10.1016/j.gecco.2018.e00470 |
| [37] |
Jing X, Prager C M, Classen A T, Maestre F T, He J S, Sanders N J. Variation in the methods leads to variation in the interpretation of biodiversity-ecosystem multifunctionality relationships. Journal of Plant Ecology, 2020, 13(4): 431-441. DOI:10.1093/jpe/rtaa031 |
| [38] |
Hillebrand H, Matthiessen B. Biodiversity in a complex world: consolidation and progress in functional biodiversity research. Ecology Letters, 2009, 12(12): 1405-1419. DOI:10.1111/j.1461-0248.2009.01388.x |
| [39] |
王凯, 王聪, 冯晓明, 伍星, 傅伯杰. 生物多样性与生态系统多功能性关系研究进展. 生态学报, 2022(01): 1-13. |
| [40] |
Yan Y Z, Zhang Q, Buyantuev A, Liu Q F, Niu J M. Plant functional β diversity is an important mediator of effects of aridity on soil multifunctionality. Science of the Total Environment, 2020, 726: 138529. DOI:10.1016/j.scitotenv.2020.138529 |
| [41] |
Gross N, Le Bagousse-Pinguet Y, Liancourt P, Berdugo M, Gotelli N J, Maestre F T. Functional trait diversity maximizes ecosystem multifunctionality. Nature Ecology & Evolution, 2017, 1(5): 0132. |
| [42] |
Mouillot D, Villéger S, Scherer-Lorenzen M, Mason N W H. Functional structure of biological communities predicts ecosystem multifunctionality. PLoS One, 2011, 6(3): e17476. DOI:10.1371/journal.pone.0017476 |
| [43] |
沈仁芳, 赵学强. 土壤微生物在植物获得养分中的作用. 生态学报, 2015, 35(20): 6584-6591. |
| [44] |
Ye J S, Delgado-Baquerizo M, Soliveres S, Maestre F T. Multifunctionality debt in global drylands linked to past biome and climate. Global Change Biology, 2019, 25(6): 2152-2161. DOI:10.1111/gcb.14631 |
| [45] |
Ma W B, Jiang S J, Assemien F, Qin M S, Ma B B, Xie Z, Liu Y J, Feng H Y, Du G Z, Ma X J, Le Roux X. Response of microbial functional groups involved in soil N cycle to N, P and NP fertilization in Tibetan alpine meadows. Soil Biology and Biochemistry, 2016, 101: 195-206. DOI:10.1016/j.soilbio.2016.07.023 |
| [46] |
Bell T, Newman J A, Silverman B W, Turner S L, Lilley A K. The contribution of species richness and composition to bacterial services. Nature, 2005, 436(7054): 1157-1160. DOI:10.1038/nature03891 |
| [47] |
He J Z, Ge Y, Xu Z H, Chen C R. Linking soil bacterial diversity to ecosystem multifunctionality using backward-elimination boosted trees analysis. Journal of Soils and Sediments, 2009, 9(6): 547. DOI:10.1007/s11368-009-0120-y |
| [48] |
Liu T, Chen X Y, Gong X, Lubbers I M, Jiang Y Y, Feng W, Li X P, Whalen J K, Bonkowski M, Griffiths B S, Hu F, Liu M Q. Earthworms coordinate soil biota to improve multiple ecosystem functions. Current Biology, 2019, 29(20): 3420-3429. DOI:10.1016/j.cub.2019.08.045 |
| [49] |
Lindborg R, Stenseke M, Cousins S A O, Bengtsson J, Berg Å, Gustafsson T, Sjödin N E, Eriksson O. Investigating biodiversity trajectories using scenarios-Lessons from two contrasting agricultural landscapes. Journal of Environmental Management, 2009, 91(2): 499-508. DOI:10.1016/j.jenvman.2009.09.018 |
| [50] |
Allan E, Manning P, Alt F, Binkenstein J, Blaser S, Blüthgen N, Böhm S, Grassein F, Hölzel N, Klaus V H, Kleinebecker T, Morris E K, Oelmann Y, Prati D, Renner S C, Rillig M C, Schaefer M, Schloter M, Schmitt B, Schöning I, Schrumpf M, Solly E, Sorkau E, Steckel J, Steffen-Dewenter I, Stempfhuber B, Tschapka M, Weiner C N, Weisser W W, Werner M, Westphal C, Wilcke W, Fischer M. Land use intensification alters ecosystem multifunctionality via loss of biodiversity and changes to functional composition. Ecology Letters, 2015, 18(8): 834-843. DOI:10.1111/ele.12469 |
| [51] |
Xue Y F, Tian J, Quine T A, Powlson D, Xing K X, Yang L Y, Kuzyakov Y, Dungait J A J. The persistence of bacterial diversity and ecosystem multifunctionality along a disturbance intensity gradient in karst soil. Science of the Total Environment, 2020, 748: 142381. DOI:10.1016/j.scitotenv.2020.142381 |
| [52] |
van der Plas F, Allan E, Fischer M, Alt F, Arndt H, Binkenstein J, Blaser S, Blüthgen N, Böhm S, Hölzel N, Klaus V H, Kleinebecker T, Morris K, Oelmann Y, Prati D, Renner S C, Rillig M C, Schaefer H M, Schloter M, Schmitt B, Schöning I, Schrumpf M, Solly E F, Sorkau E, Steckel J, Steffan-Dewenter I, Stempfhuber B, Tschapka M, Weiner C N, Weisser W W, Werner M, Westphal C, Wilcke W, Manning P. Towards the development of general rules describing landscape heterogeneity-multifunctionality relationships. Journal of Applied Ecology, 2019, 56(1): 168-179. DOI:10.1111/1365-2664.13260 |
| [53] |
Schmitz O J, Post E, Burns C E, Johnston K M. Ecosystem responses to global climate change: moving beyond color mapping. BioScience, 2003, 53(12): 1199-1205. DOI:10.1641/0006-3568(2003)053[1199:ERTGCC]2.0.CO;2 |
| [54] |
Antiqueira P A P, Petchey O L, Romero G Q. Warming and top predator loss drive ecosystem multifunctionality. Ecology Letters, 2018, 21(1): 72-82. DOI:10.1111/ele.12873 |
| [55] |
Walker T W N, Janssens I A, Weedon J T, Sigurdsson B D, Richter A, Peñuelas J, Leblans N I W, Bahn M, Bartrons M, De Jonge C, Fuchslueger L, Gargallo-Garriga A, Gunnarsdóttir G E, Marañón-Jiménez S, Oddsdóttir E S, Ostonen I, Poeplau C, Prommer J, Radujković D, Sardans J, Sigurðsson P, Soong J L, Vicca S, Wallander H, Ilieva-Makulec K, Verbruggen E. A systemic overreaction to years versus decades of warming in a subarctic grassland ecosystem. Nature Ecology & Evolution, 2020, 4(1): 101-108. |
| [56] |
Pan S F, Yang J, Tian H Q, Shi H, Chang J F, Ciais P, Francois L, Frieler K, Fu B J, Hickler T, Ito A, Nishina K, Ostberg S, Reyer C P O, Schaphoff S, Steinkamp J, Zhao F. Climate extreme versus carbon extreme: responses of terrestrial carbon fluxes to temperature and precipitation. Journal of Geophysical Research: Biogeosciences, 2020, 125(4): e2019JG005252. |
| [57] |
Frank D, Reichstein M, Bahn M, Thonicke K, Frank D, Mahecha M D, Smith P, van der Velde M, Vicca S, Babst F, Beer C, Buchmann N, Canadell J G, Ciais P, Cramer W, Ibrom A, Miglietta F, Poulter B, Rammig A, Seneviratne S I, Walz A, Wattenbach M, Zavala M A, Zscheischler J. Effects of climate extremes on the terrestrial carbon cycle: concepts, processes and potential future impacts. Global Change Biology, 2015, 21(8): 2861-2880. DOI:10.1111/gcb.12916 |
| [58] |
Dodd R J, Chadwick D R, Harris I M, Hines A, Hollis D, Economou T, Gwynn-Jones D, Scullion J, Robinson D A, Jones D L. Spatial co-localisation of extreme weather events: a clear and present danger. Ecology Letters, 2021, 24(1): 60-72. DOI:10.1111/ele.13620 |
| [59] |
朴世龙, 张新平, 陈安平, 刘强, 连旭, 王旭辉, 彭书时, 吴秀臣. 极端气候事件对陆地生态系统碳循环的影响. 中国科学: 地球科学, 2019, 49(9): 1321-1334. |
| [60] |
Reichstein M, Bahn M, Ciais P, Frank D, Mahecha M D, Seneviratne S I, Zscheischler J, Beer C, Buchmann N, Frank D C, Papale D, Rammig A, Smith P, Thonicke K, van der Velde M, Vicca S, Walz A, Wattenbach M. Climate extremes and the carbon cycle. Nature, 2013, 500(7462): 287-295. DOI:10.1038/nature12350 |
| [61] |
Bardgett R D, Caruso T. Soil microbial community responses to climate extremes: resistance, resilience and transitions to alternative states. Philosophical Transactions of the Royal Society B: Biological Sciences, 2020, 375(1794): 20190112. DOI:10.1098/rstb.2019.0112 |
| [62] |
张彬, 朱建军, 刘华民, 潘庆民. 极端降水和极端干旱事件对草原生态系统的影响. 植物生态学报, 2014, 38(9): 1008-1018. |
| [63] |
Knapp A K, Beier C, Briske D D, Classen A T, Luo Y Q, Reichstein M, Smith M D, Smith S D, Bell J E, Fay P A, Heisler J L, Leavitt S W, Sherry R, Smith B, Weng E S. Consequences of more extreme precipitation regimes for terrestrial ecosystems. Bioscience, 2008, 58(9): 811-821. DOI:10.1641/B580908 |
| [64] |
Ainsworth T D, Hurd C L, Gates R D, Boyd P W. How do we overcome abrupt degradation of marine ecosystems and meet the challenge of heat waves and climate extremes?. Global Change Biology, 2020, 26(2): 343-354. DOI:10.1111/gcb.14901 |
| [65] |
Hu T X, Hu H Q, Li F, Dou X, Sun L. Changes in the non-growing season soil heterotrophic respiration rate are driven by environmental factors after fire in a cold temperate forest ecosystem. Annals of Forest Science, 2021, 78(2): 38. DOI:10.1007/s13595-021-01060-5 |
| [66] |
Hu T X, Zhao B Q, Li F, Dou X, Hu H Q, Sun L. Effects of fire on soil respiration and its components in a Dahurian larch (Larix gmelinii) forest in northeast China: Implications for forest ecosystem carbon cycling. Geoderma, 2021, 402: 115273. DOI:10.1016/j.geoderma.2021.115273 |
| [67] |
Campos I, Abrantes N, Keizer J J, Vale C, Pereira P. Major and trace elements in soils and ashes of eucalypt and pine forest plantations in Portugal following a wildfire. Science of the Total Environment, 2016, 572: 1363-1376. DOI:10.1016/j.scitotenv.2016.01.190 |
| [68] |
Batllori E, Parisien M A, Krawchuk M A, Moritz M A. Climate change-induced shifts in fire for Mediterranean ecosystems. Global Ecology and Biogeography, 2013, 22(10): 1118-1129. DOI:10.1111/geb.12065 |
| [69] |
胡海清, 罗碧珍, 罗斯生, 魏书精, 王振师, 李小川, 刘菲. 林火干扰对森林生态系统碳库的影响研究进展. 林业科学, 2020, 56(4): 160-169. |
| [70] |
Hu T X, Hu H Q, Li F, Zhao B Q, Zhu G Y, Sun L. Long-term effects of post-fire restoration types on nitrogen mineralisation in a Dahurian larch (Larix gmelinii) forest in boreal China. Science of the Total Environment, 2019, 679: 237-247. DOI:10.1016/j.scitotenv.2019.05.008 |
| [71] |
Pérez-Valera E, Verdú M, Navarro-Cano J A, Goberna M. Soil microbiome drives the recovery of ecosystem functions after fire. Soil Biology and Biochemistry, 2020, 149: 107948. DOI:10.1016/j.soilbio.2020.107948 |
| [72] |
Beal-Neves M, Ely C V, Esteves M W, Blochtein B, Lahm R A, Quadros E L L, Ferreira P M A. The influence of urbanization and fire disturbance on plant-floral visitor mutualistic networks. Diversity, 2020, 12(4): 141. DOI:10.3390/d12040141 |
| [73] |
Jactel H, Desprez-Loustau M L, Battisti A, Brockerhoff E, Santini A, Stenlid J, Björkman C, Branco M, Dehnen-Schmutz K, Douma J C, Drakulic J, Drizou F, Eschen R, Franco J C, Gossner M M, Green S, Kenis M, Klapwijk M J, Liebhold A M, Orazio C, Prospero S, Robinet C, Schroeder M, Slippers B, Stoev P, Sun J H, van den Dool R, Wingfield M J, Zalucki M P. Pathologists and entomologists must join forces against forest pest and pathogen invasions. NeoBiota, 2020, 58: 107-127. DOI:10.3897/neobiota.58.54389 |
| [74] |
吴飞华, 刘廷武, 裴真明, 郑海雷. 酸雨引起森林生态系统钙流失研究进展. 生态学报, 2010, 30(4): 1081-1088. |
| [75] |
汤永康, 武艳涛, 武魁, 郭之伟, 梁存柱, 王敏杰, 常佩静. 放牧对草地生态系统服务和功能权衡关系的影响. 植物生态学报, 2019, 43(5): 408-417. |
| [76] |
邢鹏飞, 李刚, 陈晓鹏, 李德颖, 王常慧, 董宽虎, 赵祥. 放牧对晋北农牧交错带赖草草地生态系统碳交换的影响. 草业学报, 2019, 28(10): 1-11. |
| [77] |
Zhang R Y, Wang J S, Niu S L. Toward a sustainable grazing management based on biodiversity and ecosystem multifunctionality in drylands. Current Opinion in Environmental Sustainability, 2021, 48: 36-43. DOI:10.1016/j.cosust.2020.09.005 |
| [78] |
李文, 曹文侠, 师尚礼, 李小龙, 陈建刚, 徐长林. 放牧管理模式对高寒草甸生态系统有机碳、氮储量特征的影响. 草业学报, 2016, 25(11): 25-33. |
| [79] |
Huang J Y, Yu H L, Liu J L, Luo C K, Sun Z J, Ma K B, Kang Y M, Du Y X. Phosphorus addition changes belowground biomass and C: N: P stoichiometry of two desert steppe plants under simulated N deposition. Scientific Reports, 2018, 8: 3400. DOI:10.1038/s41598-018-21565-w |
| [80] |
杨晓霞, 任飞, 周华坤, 贺金生. 青藏高原高寒草甸植物群落生物量对氮、磷添加的响应. 植物生态学报, 2014, 38(2): 159-166. |
| [81] |
田沐雨, 于春甲, 汪景宽, 丁凡, 陈振华, 姜楠, 蒋晖, 陈利军. 氮添加对草地生态系统土壤pH、磷含量和磷酸酶活性的影响. 应用生态学报, 2020, 31(9): 2985-2992. |
| [82] |
张秀兰, 王方超, 方向民, 何平, 张宇飞, 陈伏生, 王辉民. 亚热带杉木林土壤有机碳及其活性组分对氮磷添加的响应. 应用生态学报, 2017, 28(2): 449-455. |
| [83] |
Zhang B, Liang A Z, Wei Z B, Ding X L. No-tillage leads to a higher resistance but a lower resilience of soil multifunctionality than ridge tillage in response to dry-wet disturbances. Soil and Tillage Research, 2019, 195: 104376. DOI:10.1016/j.still.2019.104376 |
| [84] |
Castioni G A F, Cherubin M R, de Oliveira Bordonal R, Barbosa L C, Menandro L M S, Carvalho J L N. Straw removal affects soil physical quality and sugarcane yield in brazil. BioEnergy Research, 2019, 12(4): 789-800. DOI:10.1007/s12155-019-10000-1 |
| [85] |
徐煖银, 孙思琦, 薛达元, 郭泺. 基于地形梯度的赣南地区生态系统服务价值对人为干扰的空间响应. 生态学报, 2019, 39(1): 97-107. |
| [86] |
Hector A, Joshi J, Scherer-Lorenzen M, Schmid B, Spehn E M, Wacker L, Weilenmann M, Bazeley-White E, Beierkuhnlein C, Caldeira M C, Dimitrakopoulos P G, Finn J A, Huss-danell K, Jumpponen A, Leadley P W, Loreau M, Mulder C P H, Neẞhöver C, Palmborg C, Read D J, Siamantziouras A S D, Terry A C, Troumbis A Y. Biodiversity and ecosystem functioning: reconciling the results of experimental and observational studies. Functional Ecology, 2007, 21(5): 998-1002. DOI:10.1111/j.1365-2435.2007.01308.x |
| [87] |
Gamfeldt L, Hillebrand H. Biodiversity effects on aquatic ecosystem functioning-maturation of a new paradigm. International Review of Hydrobiology, 2008, 93(4/5): 550-564. |
| [88] |
徐炜, 马志远, 井新, 贺金生. 生物多样性与生态系统多功能性: 进展与展望. 生物多样性, 2016, 24(1): 55-71. |
| [89] |
张宏锦, 王娓. 生态系统多功能性对全球变化的响应: 进展、问题与展望. 植物生态学报, 2021, 45: 1-15. |
| [90] |
Scherer-Lorenzen M, Potvin C, Koricheva J, Schmid B, Hector A, Bornik Z, Reynolds G, Schulze E D. The design of experimental tree plantations for functional biodiversity research//Scherer-Lorenzen M, Körner C, Schulze E D, eds. Forest Diversity and Function: Temperate and Boreal Systems. Berlin, Heidelberg: Springer, 2005: 347-376.
|
| [91] |
Meyer S T, Ptacnik R, Hillebrand H, Bessler H, Buchmann N, Ebeling A, Eisenhauer N, Engels C, Fischer M, Halle S, Klein A M, Oelmann Y, Roscher C, Rottstock T, Scherber C, Scheu S, Schmid B, Schulze E D, Temperton V M, Tscharntke T, Voigt W, Weigelt A, Wilcke W, Weisser W W. Biodiversity-multifunctionality relationships depend on identity and number of measured functions. Nature Ecology & Evolution, 2018, 2(1): 44-49. |