 2022, Vol. 42
2022, Vol. 42文章信息
- 王俊, 潘鸿, 谢磊, 崔国发
- WANG Jun, PAN Hong, XIE Lei, CUI Guofa
- 华南五针松濒危机制生态学研究
- An ecological study on the endangered mechanism of Pinus kwangtungensis
- 生态学报. 2022, 42(15): 6311-6319
- Acta Ecologica Sinica. 2022, 42(15): 6311-6319
- http://dx.doi.org/10.5846/stxb202103120670
-
文章历史
- 收稿日期: 2021-03-12
- 网络出版日期: 2022-04-07





种群分布格局不仅能反映某个种群适应生境后对空间占有的一种形式, 而且在不同生境梯度下植物种群在其定植、生长的过程中所经历的生态学过程不尽相同[1—5]。在一定的尺度范围内, 通过对种群分布格局的分析, 可以对种群的水平结构进行定量描述, 也可以进一步揭示植物群落特征的基础和格局的成因, 阐述种群及群落的动态变化等, 从而为植物保护、就地保护或迁地保护的实施提供一定的科学依据[6]。
群落组成与结构是由决定性因素(如生态位需求和种间作用因素)、随机因素(定居和干扰)两个方面决定的, 且群落组成种的相对多度和格局与其相对竞争能力有关[7—8]。植物受竞争影响程度取决于自身及相邻植物大小、数量、植物利用资源的特性和空间影响区域, 且种间竞争是植物间竞争的重要组成部分[9—11]。
华南五针松(Pinus kwangtungensis)又名广东松, 虽然其生态适应性较强, 现代地理分布区范围跨越大, 但呈现出分布零散、数量少、海拔高等特点[12—16]。前人对华南五针松的研究集中在局部分布区的种群、群落特征和种间关系[17—21], 以及华南五针松地理分布的主要限制因子, 并讨论了在气候变化的情况下, 华南五针松分布区的变化趋势等[16, 22]。至此, 但在群落生态学上, 国内外还未见有从华南五针松种群空间格局和生存压力等方面探讨其濒危机制的研究。
当前, 在全球气温升高、自然资源过度开发、环境污染日趋严重的情况下, 通过在种群生态学基础上定量计算不同海拔濒危植物的生存压力指数, 提出植物间“竞争驱赶”假说, 并从理论和实践两方面进行验证, 为濒危植物的就地保护或近地保护研究提供理论基础。华南五针松的自然演替处于衰退之中[23], 研究华南五针松的濒危原因不但为其他五针松组植物的濒危机制提供重要素材外, 也为其它物种的濒危机制研宄提供了案例。
1 研究区概况广东南岭国家级自然保护区地处广东省北部韶关市的乳源县、清远市的阳山县和连州市行政境界内, 保护区面积58368.4hm2。地理坐标为112°30′—113°04′E, 24°37′—24°57′N。总体上属于花岗岩山地的中山山地, 区内山高谷深, 地势峻峭, 最高峰石坑岭海拔1902m, 为广东省第一高峰, 其次为紫云山海拔为1888m, 小黄山最高海拔为1600m。气候属典型的亚热带温湿气候, 因地势较高, 具山地气候特色, 年平均气温17.7℃, 最高温34.4℃, 最低温-3.6℃;降水量较充沛, 多集中3—8月, 年平均达1705mm, 最高年份达2495mm, 年平均相对湿度84%;土壤类型随海拔高度不同, PH值在4—5.5, 海拔900—1800之间为山地黄壤[24]。
广东南岭国家级自然保护区是广东珍稀濒危针叶树种最集中的分布区, 在乳阳林区中山山地的小黄山和紫云山, 保存着1300hm2的华南五针松原始林, 植株多, 面积大, 以华南五针松、五列木(Pentaphylax euryoidex)、长苞铁杉(Tsuga longibracteata)、福建柏(Fokienia hodginsii)为优势种的原始针阔混交林。
南岭地质历史悠久, 是植物物种南北交流和东西交汇的重要过渡地区, 是我国中亚热带与南亚热带重要的气候分界线, 在阻挡南北气流的运行方面发挥着重要作用, 是华南五针松的核心分布区, 同时也是我国亚热带常绿阔叶林的主要分布区[25]。
2 研究方法 2.1 文献收集法根据《中国植被图(1 ∶ 100万)》和《中国植物志》等相关植物方面的书籍, 查找出在南岭地区不同海拔华南五针松所处的典型植被, 并根据不同植被型中的建群种或共优种, 记录其生物学高度、胸径和冠幅等, 计算出华南五针松的平均压迫力。
2.2 样地调查法于2018年4月底至5月初, 在植物标本采集和踏查的基础上, 根据植物的分布和具体生境, 采用典型抽样调查法, 在华南五针松生存最低海拔下限和最高海拔上限之间, 沿垂直高度以60—200m为海拔梯度设置样地, 利用“种—面积曲线”法求算群落的最小样地面积。在同一分布区内的两个山头, 共设置样地14个, 其中1个人工林(实验区的树木园)于1977年栽种, 每个样地面积400m2。样地调查详细情况如表 1。
| 样地编号 Sample number |
地点 Site |
经纬度 Longitude and latitude |
海拔/m Altitude |
坡度% Gradient |
坡向 Slope |
郁闭度 Canopy density |
林分起源 Stand origin |
| 001 | 乳峰楼 | 24°53′55.81″ 113°1′11.02″ | 1575 | 60—65 | 正南 | 0.8 | 天然林 |
| 002 | 乳阳小黄山 | 24°53′46.6″ 113°1′7.82″ | 1373 | 28—34 | 西南 | 0.7 | 人工林 |
| 003 | 乳阳小黄山 | 24°53′52″ 113°1′12.67″ | 1509 | 37—43 | 正南 | 0.82 | 天然林 |
| 004 | 乳阳小黄山 | 24°53′57.74″ 113°1′22.39″ | 1447 | 50—60 | 正南 | 0.83 | 天然林 |
| 005 | 乳阳小黄山 | 24°53′44.77″ 113°1′39.6″ | 1186 | 50—55 | 东南 | 0.78 | 天然林 |
| 006 | 乳阳小黄山 | 24°53′48.5″ 113°1′34″ | 1276 | 60—65 | 东南 | 0.75 | 天然林 |
| 007 | 乳阳小黄山 | 24°53′50.9″ 113°1′28.78″ | 1370 | 45—50 | 东北 | 0.8 | 天然林 |
| 008 | 九重山 | 24°53′47.89″ 113°2′24.19″ | 1032 | 47—52 | 正南 | 0.5 | 天然林 |
| 009 | 瀑布亭 | 24°53′49″ 113°2′29.47″ | 994 | 51—56 | 东南 | 0.6 | 天然林 |
| 010 | 紫云山 | 24°53′17.93″ 113°0′20.65″ | 1685 | 40—45 | 东南 | 0.56 | 天然林 |
| 011 | 紫云山 | 24°53′12.83″ 113°0′24.53″ | 1630 | 40—50 | 东南 | 0.55 | 天然林 |
| 012 | 紫云山 | 24°53′6.63″ 113°0′29.23″ | 1512 | 42—47 | 东南 | 0.73 | 天然林 |
| 013 | 紫云山 | 24°53′19″ 113°0′19″ | 1710 | 5—7 | 正南 | 0.2 | 天然林 |
| 014 | 树木园 | 24°55′24.43″ 113°4′35.71″ | 569 | 4—8 | 正东 | 0.85 | 人工林 |
样地内华南五针松幼苗极小, 且数量少, 所以对样方内胸径(DBH)>4cm的乔木采用“每木调查法”, 测定其胸径、基径、树高、枝下高、冠幅等指标, 简单记录灌木、草本植物的种类, 同时以每个乔木样方的两边为坐标轴测定每株树木的相对坐标(X, Y), 并记录每个大样方的生境因子, 例如经纬度、海拔、坡度、坡向、坡位、郁闭度等。
2.3 计算方法考虑到在植物群落内, 植株个体之间的竞争往往取决于植株对生存环境和空间资源的争夺, 尤其表现在对光资源和水资源的竞争。在水资源不成为限制因子时, 植株树高越高、冠幅越大对光资源的竞争力越强, 从而对其他植株的影响程度也就越大。胸径相等的不同树种间植株个体的树高和冠幅大小往往存在很大差异, 再者, 树冠大小相同而处在不同方位的植株个体对珍稀濒危树种个体造成的生存压力也不同。
鉴于此, 采用一种珍稀濒危树种个体生存压力计算方法[11, 26—27], 能够客观、定量地反映珍稀濒危树种个体在植物群落中的生存压力和平均压迫力大小, 并在此方法上进行了改进。
计算方法包括以下步骤:
(1) 竞争木的确定:当某树种的植株个体树高即H大于等于其与珍稀濒危树种个体距离即L时, 确定此植株为竞争木;
(2) 计算珍稀濒危树种个体与竞争木的树冠大小V, 树冠近似看作为椭圆柱, 用椭圆柱体积量化树冠大小;
(3) 计算珍稀濒危树种个体承受的来自某一株竞争木的单株实际生存压力指数PIij, 理论压迫力P;
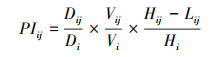
|
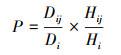
|
式中, PIij为单株实际生存压力指数, 即珍稀濒危树种第i株个体承受第j株竞争木的生存压力, PIij≥0;当竞争木与珍稀濒危树种个体的距离Lij等于其树高Hij时, 该竞争木无竞争效应, PIij为最小值0;当竞争木与珍稀濒危树种个体距离越近, 树冠越大, 所处方位越接近珍稀濒危树种个体的正南方向时, PIij越大, 当PIij≥1时, 认为珍稀濒危树种第i株个体承受第j株竞争木的生存压力较大;P为理论压迫力;
Vij为第j株竞争木的树冠大小, 单位为m3;
Vi为珍稀濒危树种第i株个体的树冠大小, 单位为m3;
Di和Dij分别为第i株珍稀濒危树种和第j株竞争木的胸径;
(4) 计算珍稀濒危树种个体承受的来自某个竞争树种即t的生存压力指数PIit;
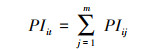
|
式中, PIit为单个树种生存压力指数, 即珍稀濒危树种第i株个体承受的来自竞争树种t的生存压力;m为竞争树种t的竞争木的株数;
(5) 计算珍稀濒危树种个体承受的来自所有竞争树种的生存压力指数PIi:
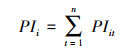
|
式中, PIi为单个树种总生存压力指数, 即珍稀濒危树种第i株个体承受的来自所有竞争树种的生存压力;n为竞争树种总数。
(6) 计算珍稀濒危树种承受的来自所有竞争树种的平均生存压力指数PI:
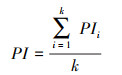
|
式中, PI为濒危树种平均生存压力指数, 即珍稀濒危树种承受的来自所有竞争树种t的平均生存压力;k为珍稀濒危树种总数。
3 结果与分析 3.1 沿海拔梯度上种群空间格局种群的空间格局的计算方法很多, 采用目前较为常用的具有相同本质或较大相似性的8种方法(方差/均值比率、t值检验、负二项参数、扩散型指数、Cassie指标、丛生指标、平均拥挤度和聚块性指标等)综合分析华南五针松种群的分布格局[6, 28—31], 计算结果见表 2。
| 海拔 Altitude |
分布格局参数 Distribution pattern parameter |
聚集强度参数 Aggregation intensity parameter |
分布型 Distribution type |
||||||||||
| 均值 Mean value |
方差 Variance |
C | t | CA | GI | I | PI | Iδ | K | m* | |||
| 低海拔 Low altitude (< 1300m) |
6.5 | 7 | 1.07 | 0.06* | 0.01 | -0.2 | 0.07 | 1.01 | 5.39 | 84.5 | 6.57 | C或U | |
| 中海拔 Mid altitude (1300—1500m) |
21 | 133 | 6.33 | 4.48* | 0.25 | 55.5 | 5.33 | 1.25 | 65.83 | 3.93 | 26.33 | C | |
| 高海拔 High altitude (1500—1600m) |
24.3 | 586.3 | 24.09 | 19.42* | 0.94 | 280.5 | 23.09 | 1.94 | 122.5 | 1.05 | 47.42 | C | |
| 较高海拔 Higher altitude (>1600m) |
2.66 | 4.33 | 1.62 | 0.52 | 0.2 | 0.33 | 0.62 | 1.23 | 0.4 | 4.26 | 3.29 | R或C | |
| *:P < 0.05;C:扩散系数Coefficient of diffusion;t:t检验test;CA:Cassie指标Cassie index;GI:Green指数Green index;I:丛生指数Clumping index; PI:聚块性指标Clustering index;Iδ:Moristia指数Moristia index;K:负二项参数Negative binomial parameter;m*:平均拥挤度Mean crowding;C:聚集Cluster;U:均匀Uniform;R: 随机Random | |||||||||||||
由表 2可知, 在不同海拔梯度上C、Iδ和PI均大于1, I和CA均大于0, 而GI小于0。以上6个指标表明种群在中、高海拔处为聚集分布, 低海拔为聚集或均匀, 且C通过了t检验, 进而验证沿海拔梯上种群的分布格局与预期假设呈显著差异。
随海拔的升高, Iδ和m*的值逐渐增大, 同时K值逐渐减少, 由此表明, 随海拔梯度的增加(1600m除外), 种群聚集强度增加, 种群的拥挤程度也相应增加, 种间和种内竞争激烈, 竞争压力增大;当海拔在1600m以上时, K值又变大, 表明聚集程度变缓, 生存压力减小。
3.2 “竞争驱赶”假说的提出与验证植物为了其更好生存需最大限度的加大对养分、能源和空间等的争夺, 当两个或者多个植物所要求的资源, 空间等发生重叠时就发生种内和种间竞争, 树高冠密的个体就会占据优势, 欺负其他植物, 驱使他们逃离到生存压力小的地方或死亡, 这就是“竞争驱赶”假说。
本次调查中, 在华南五针松生存的最低海拔下限和最高海拔上限之间, 沿小黄山和紫云山共设置13块样地, 共测175棵华南五针松。从图 1中可以得出:沿海拔梯度的上升, 华南五针松种群平均生存压力指数整体上呈递减趋势(R2=0.31, P < 0.05), 海拔较高的区域生存压力指数较小, 利于华南五针松种群的生存。

|
| 图 1 不同海拔梯度上华南五针松种群平均生存压力指数 Fig. 1 The everage survival pressure index of the population of Pinus kwangtungensis along different elevation gradients |
在中高海拔1509m处物种丰富, 种群密度大, 伴生种的平均树高和冠幅均高于华南五针松, 种群竞争激烈, 造成华南五针松种群平均生存压力指数较大, 在拟合回归中出现突变点。
南岭属亚热带季风湿润气候, 其水平地带性植被为中亚热带南部亚地带常绿阔叶林。由表 3得出, 不同海拔区段呈现出不同的植被型, 华南五针松适生于中上海拔的针阔混交林内, 这跟实际调查的华南五针松分布海拔是一致的;《中国植物志》中记录华南五针松高达30 m, 胸径1.5m, 分布海拔为700—1600 m, 而在实际调查中最低海拔为994 m, 最高海拔接近1710 m, 这种分布海拔区域的变化极有可能是由于阔叶树种的存在, 且树高冠密, 平均压迫力较大, 从而驱使针叶树种有一定程度上的提升。
| 海拔 Altitude/m |
植被型 Vegetation type |
建群种或共优种 Constructive species or co-optimal species |
平均树高/m Average tree height |
平均胸径/m Mean DBH |
平均压迫力 Average pressure |
| 100—300 | 亚热带河滩灌丛 | 柳叶润楠Machilus salicina | 5 | ||
| 200—800 | 亚热带常绿阔叶林 | 樟Cinnamomum camphora、木荷、南岭栲Castanopsis fordii、蕈树Altingia chinensis | 26.25 | 1.4 | 0.87 |
| 800—1200 | 亚热带常绿与落叶阔叶林 | 水青冈Fagus longipetiolata、银钟花Halesia macgregorii、光皮桦、甜槠 | 22.65 | 0.66 | 0.33 |
| 1000—1500 | 亚热带常绿针阔叶混交林 | 华南五针松、福建柏、长苞铁杉、硬斗柯Lithocarpus hancei、木荷 | 23.8 | 1.07 | 0.5 |
| >1500 | 亚热带常绿针阔混交山顶矮林 | 南华杜鹃Rhododendron simiarum、宽叶粗榧Cephalotaxus latifolia | 3 | ||
| DBH: 胸径diameter at breast height | |||||
不论从理论(表 3)和实际调查(图 1)中发现, 华南五针松是有一个暂时稳定的群落环境, 但随着群落内植物种内和种间竞争驱赶, 全球气温升高, 且华南五针松种群喜温凉湿润, 而高海拔生存压力较小, 受到的阻力就小, 种群更加倾向于高海拔分布, 但当种群分布上移到达山顶时, 会出现温度急剧降低, 风大雾多, 土层浅薄等现象, 若干年后华南五针松则无处存活被迫退出群落, 导致种群的灭亡。
3.3 同海拔生存压力表 4、图 2和图 3表明:在土壤质地、坡向和气候基本相似的情况下, 在海拔1509m处, 华南五针松种群平均生存压力指数为120, 而在海拔1512m处的平均生存压力指数是2.6, 前者是后者的46倍。在1509m处, 华南五针松成片存在, 密度大, 物种多样性丰富, 平均树高9.33m, 平均胸径和冠幅分别为12.73cm和36m3, 而竞争木像马蹄荷(Exbucklandia populnea)、木荷(Schima superba)、南方铁杉(Tsuga chinensis)、五列木等树种的平均树高为12m, 平均冠幅是100m3, 种内和种间相互竞争, 且以种内竞争为主, 伴生种的主要竞争顺序为木荷>甜槠(Castanopsis eyrei)>五列木>马蹄荷(Exbucklandia populnea)>马尾松(Pinus massoniana)>长苞铁杉;在1512m处, 华南五针松种群的群落结构较简单, 只有少量的南方铁杉和长苞铁杉等树种, 平均树高不高于4.5m, 以种内竞争为主, 其他竞争树种对其竞争生存压力较小。
| 海拔 Altitude/m |
平均树高 Mean height/m |
最高 Highest/m |
平均胸径 Mean DBH/ cm |
种间竞争指数 Index of interspecific competition |
种内竞争指数 Index of intraspecific competition |
平均生存压力指数 Everage survival pressure index |
| 1512 | 4.24 | 5.4 | 11.54 | 4.81 | 18.75 | 2.6 |
| 1509 | 9.33 | 17 | 12.73 | 2312.61 | 5331.02 | 120 |
| 1370 | 20.09 | 25.3 | 32.57 | 139.11 | 873.97 | 34 |
| 1373 | 13.43 | 17.3 | 15.54 | 2987.49 | 1090.73 | 200 |

|
| 图 2 海拔1509m华南五针松群落特征及种间生存压力 Fig. 2 The characteristic of the community of Pinus kwangtungensis and the survival pressure of interspecific at 1509m |

|
| 图 3 海拔1512m华南五针松群落特征及种间生存压力 Fig. 3 The characteristic of the community of Pinus kwangtungensis and the survival pressure of interspecific at 1512m |
由表 4、图 4和图 5得出:在土壤质地、坡向和气候基本相似的情况下, 在海拔1370m, 华南五针松种群平均生存压力指数为34, 而在海拔1373m处的生存压力指数为200, 两者相差约160。这主要是由于在在海拔1370m处, 样地为半阴坡, 华南五针松天然生存, 平均树高20.09m, 以种内竞争为主;虽为优势种, 但主要受青冈(Cyclobalanopsis glauca)、木荷、甜槠等物种的竞争, 竞争顺序为青冈>木荷>甜槠>五列木;而在海拔1373m处, 样地为半阳坡, 属华南五针松人工林, 种群竞争压力大, 以种间竞争为主, 伴生种的竞争顺序为吴茱萸(Evodia rutaecarpa)>拟赤杨(Alniphyllum fortunei)>红楠(Machilus thunbergii)>光皮桦(Betula luminifera)>仁昌木莲(Manglietia chingii)>乳源木莲(Manglietia yuyuanensis), 群落结构不合理造成竞争压力巨大。

|
| 图 4 海拔1370m华南五针松群落特征及种间生存压力 Fig. 4 The characteristic of the community of Pinus kwangtungensis and the survival pressure of interspecific at 1370m |

|
| 图 5 海拔1370m华南五针松群落特征及种间生存压力 Fig. 5 The characteristic of the community of Pinus kwangtungensis and the survival pressure of interspecific at 1373m |
在低海拔569m处, 保护区于1977年育苗种植14棵华南五针松, 经过近40年的时间, 现今只有4棵活立木, 且种群平均生存压力指数为50, 竞争压力大, 若不加以严格保护, 及时对竞争树种进行整枝或择伐处理, 预计4棵华南五针松将会在10—20年时间里成为濒死木。
4 讨论种群空间格局随尺度的变化而不同, 不同的空间格局特征由不同的因素所致, 而本文只是在一定区域采用样方分析法研究种群空间格局, 且随尺度的不断变大, 生物因素和非生物因素的作用加强, 种群空间格局会相应的改变, 下一步将从生态位的分化理论及对不同生境的偏好程度来分析华南五针松种群的适应性、物种共存和不同尺度下的分布格局。
植物濒危或灭亡是由内在和外在等各种因素共同作用的结果, 且物种间的空间分离减弱种间竞争, 阻止或减缓竞争优势物种对竞争劣势物种的竞争排除作用, 这有利于维持物种共存和物种多样性维持[32]。华南五针松虽在主林层占据显著地位, 但在演替层和更新层中的优势远不及其他优势种[19], 随着群落内种间、种内竞争的激烈, 华南五针松种群在中高海拔处生存压力最大, 而华南五针松种群在沿海拔梯度上升过程中平均生存压力指数整体上呈递减趋势, 海拔较高的区域利于华南五针松种群的生存, 物种会倾向于向高海拔迁移, 降低其他优势物种的竞争强度, 这与在温度升高下, 华南五针松分布的最低海拔将不断上升, 南亚热带和北热带地区的华南五针松已经上移到山顶的结论[16]和群落种间竞争驱赶致使群落正向演替是华木莲濒危的一个重要原因[33]有相似之处。而本研究只是在一定区域内通过沿海拔梯度上计算种群生存压力, 提出“竞争驱赶”假说, 并进行了验证, 但随着时间的推移, 华南五针松种群的数量会不会减少以及分布区的上限和下限会不会缩短都将进一步需要验证。
华南五针松作为一种比较原始和古老的物种, 幼苗难以成活, 种群更新困难, 对温度、湿度比较敏感。由于气候变暖将导致物种适宜生境的丧失和破碎化, 已造成欧洲南部山脉地区大量物种的数量在过去10年里正在逐步减少, 在地中海地区14个物种向山顶移动, 分布区逐渐缩小, 生存压力大和不适应, 从而引起高山植物灭绝或被其他物种取代[34—36], 及在温度对华南五针松地理分布起主导作用, 其对高温条件的要求极为严格, 喜温凉湿润的环境下[16], 随着气候带北移, 利用土壤、湿度、物候和降水等因素, 结合区域气候等模型, 探讨华南五针松分布区是否会北移及向高海拔迁移, 以及关键致危因子和濒危理论机制还需要进一步的研究。
造成物种濒危的因素主要是内因或者外因, 抑或是两者的结合。“竞争驱赶”假说是在种群和群落基础上推断物种濒危的一个方面, 属第一次提出, 是对外因的一个重要补充。其生存压力指数的计算综合考虑了树高、树冠、距离、胸径等生态因子, 优于诸如邻体干扰和Hegyi单木竞争等模型的计算方法, 能够更客观、真实、定量反映珍稀濒危树种个体及种群在群落中的生存状况, 为物种濒危的推断提供了一个全新模式。
5 结论华南五针松种群在中、高海拔处为聚集分布, 低海拔为聚集或均匀分布, 且随海拔的升高, Iδ和m*的值逐渐增大, 同时K值逐渐减少, 种群聚集强度增加, 当达到一定高度时, 聚集强度降低, 种内和种间竞争减缓, 生存压力变小, 验证“竞争驱赶”假说的存在。
在南岭国家级自然保护区内, 华南五针松种群天然分布的垂直最低海拔为1000m左右, 最高海拔上限1710m左右, 在理论分布上有所抬升, 且随海拔升高, 华南五针松平均生存压力变小, 海拔较高的区域利于华南五针松种群的生存, 但当物种继续向高海拔迁移, 这些物种将由于分布区逐渐缩小、生存压力大和不适应等因素造成最终灭绝, 因此本研究认为物种之间存在“竞争驱赶”, 这可能是导致华南五针松濒危的原因之一, 为华南五针松植物濒危机制的理论研究提供一个示例。
海拔高度和温度影响植物的群落结构, 在立地条件相似的情况下, 由于华南五针松复杂的群落结构及激烈的种内、种间竞争, 造成同海拔的华南五针松种群呈现不同的生存压力及华南五针松人工林群落中种群生存压力指数大于天然林群落。
| [1] |
张金屯. 植被数量生态学方法. 北京: 中国科学技术出版社, 1995.
|
| [2] |
Condit R, Ashton P S, Baker P, Bunyavejchewin S, Gunatilleke S, Gunatilleke N, Hubbell S P, Foster R B, Itoh A, LaFrankie J V, Lee H S, Losos E, Manokaran N, Sukumar R, Yamakura T. Spatial patterns in the distribution of tropical tree species. Science, 2000, 288(5470): 1414-1418. DOI:10.1126/science.288.5470.1414 |
| [3] |
Wiles L J. Spatial pattern analysis in plant ecology. Weed Technology, 2001, 15(1): 195-196. DOI:10.1614/0890-037X(2001)015[0195:]2.0.CO;2 |
| [4] |
Wiegand T, Gunatilleke S, Gunatilleke N. Species associations in a heterogeneous Sri Lankan dipterocarp forest. The American Naturalist, 2007, 170(4): E77-E95. DOI:10.1086/521240 |
| [5] |
董雪, 杜昕, 孙志虎, 谷会岩, 陈祥伟. 生境梯度影响下的天然红松种群空间格局与种内关联. 生态学报, 2020, 40(15): 5239-5246. |
| [6] |
张瑾, 陈文业, 张继强, 赵明, 吴三雄, 王志广, 袁海峰, 窦英杰, 康建军, 罗文莉. 甘肃敦煌西湖荒漠-湿地生态系统优势植物种群分布格局及种间关联性. 中国沙漠, 2013, 33(2): 349-357. |
| [7] |
Aguiar M R, Lauenroth W K, Peters D P. Intensity of intra-and interspecific competition in coexisting shortgrass species. Journal of Ecology, 2001, 89(1): 40-47. DOI:10.1046/j.1365-2745.2001.00523.x |
| [8] |
高艳. 黄土丘陵区撂荒群落共存种植物竞争对土壤水分变化的响应[D]. 咸阳: 中国科学院研究生院(教育部水土保持与生态环境研究中心), 2016.
|
| [9] |
Weiner J. Neighbourhood interference amongst Pinus rigida individuals. Journal of Ecology, 1984, 72(1): 183-195. DOI:10.2307/2260012 |
| [10] |
王会梅. 黄土区几种牧草植物的生态适应性与种间竞争研究[D]. 咸阳: 西北农林科技大学, 2006.
|
| [11] |
王泳腾, 黄治昊, 王俊, 张童, 郎立华, 孙国明, 崔国发. 濒危植物黄檗的生存压力研究. 北京林业大学学报, 2021, 43(1): 49-57. |
| [12] |
王献溥, 李信贤. 广西环江县石灰岩山地广东松林群落学特点的研究. 植物研究, 1989, 9(3): 77-86. |
| [13] |
李林初, 钱吉. 两种中国特有松属植物核型的研究兼论松属的系统位置. 云南植物研究, 1993, 15(1): 47-56. |
| [14] |
王发国, 张荣京, 邢福武, 吴世捷, 叶育石, 陈焕强. 海南鹦哥岭自然保护区的珍稀濒危植物与保育. 武汉植物学研究, 2007, 25(3): 303-309. DOI:10.3969/j.issn.2095-0837.2007.03.017 |
| [15] |
黄练忠, 张尚坤, 陈进, 熊基粦. 东莞银瓶山森林公园广东松+红花荷群落的基本特征. 广东林业科技, 2011, 27(5): 13-18. DOI:10.3969/j.issn.1006-4427.2011.05.003 |
| [16] |
陶翠, 李晓笑, 王清春, 崔国发. 中国濒危植物华南五针松的地理分布与气候的关系. 植物科学学报, 2012, 30(6): 577-583. |
| [17] |
杜道林, 刘玉成, 苏杰. 茂兰喀斯特山地广东松种群结构和动态初步研究. 植物生态学报, 1996, 20(2): 159-166. |
| [18] |
杨沅志, 张璐, 陈北光, 苏志尧. 珍稀濒危植物广东松林的群落特征. 华南农业大学学报, 2006, 27(2): 70-73. DOI:10.3969/j.issn.1001-411X.2006.02.018 |
| [19] |
张璐, 苏志尧, 陈北光. 中国特有植物——华南五针松群落的种间关系. 生态学报, 2006, 26(4): 1063-1072. DOI:10.3321/j.issn:1000-0933.2006.04.012 |
| [20] |
王厚麟, 缪绅裕, 邓敏, 邹滨, 沈孝清, 扶建庭. 广东乐昌杨东山-十二渡水保护区广东松群落的特征. 生态科学, 2007, 26(2): 115-119. DOI:10.3969/j.issn.1008-8873.2007.02.005 |
| [21] |
王俊, 王泳腾, 段洋波, 谢磊, 崔国发. 广东南岭国家级自然保护区华南五针松种内和种间关系研究. 北京林业大学学报, 2020, 42(5): 25-32. |
| [22] |
李越, 李胜利, 杨昌腾, 旷远文, 曹宗英. 南岭华南五针松树轮宽度对气候因子的响应. 亚热带资源与环境学报, 2016, 11(1): 26-31, 51-51. DOI:10.3969/j.issn.1673-7105.2016.01.005 |
| [23] |
徐燕千. 建立南岭国家级自然保护区的重大意义. 生态科学, 1993(1): 14-20. |
| [24] |
王发国, 陈振明, 陈红锋, 邢福武, 董安强. 南岭国家级自然保护区植物区系与植被. 武汉: 华中科技大学出版社, 2013.
|
| [25] |
祁承经, 萧育檀, 曹铁如, 林亲众. 南岭植被的研究. 中南林学院学报, 1992, 12(1): 1-10. |
| [26] |
魏永久. 濒危植物海南五针松初步调查研究[D]. 北京: 北京林业大学, 2015.
|
| [27] |
张建亮, 刘方正, 崔国发. 一种珍稀濒危树种个体生存压力计算方法: 中国, CN104094814A. 2014-10-15.
|
| [28] |
张金屯. 植物种群空间分布的点格局分析. 植物生态学报, 1998, 22(4): 344-349. DOI:10.3321/j.issn:1005-264X.1998.04.008 |
| [29] |
赵常明, 陈庆恒, 乔永康, 潘开文. 青藏高原东缘岷江冷杉天然群落的种群结构和空间分布格局(英文). 植物生态学报, 2004, 28(3): 341-350. DOI:10.3321/j.issn:1005-264X.2004.03.009 |
| [30] |
周洪华, 陈亚宁, 李卫红. 塔里木河下游绿洲-荒漠过渡带植物多样性特征及优势种群分布格局. 中国沙漠, 2009, 29(4): 688-696. |
| [31] |
高文强. 不同地理梯度上栓皮栎种群动态及其环境解释[D]. 北京: 中国林业科学研究院, 2017.
|
| [32] |
姚杰, 张春雨, 赵秀海. 吉林蛟河阔叶红松林树种空间分布格局及其种间关联性. 林业科学, 2018, 54(8): 23-31. |
| [33] |
杨清培, 肖智勇, 胡晓东, 欧阳明, 陈昕, 林国江, 徐佳文, 杨光耀. 珍稀植物华木莲的现代濒危机制: 探析与展望. 广西植物, 2017, 37(5): 653-660. |
| [34] |
陈建国, 杨扬, 孙航. 高山植物对全球气候变暖的响应研究进展. 应用与环境生物学报, 2011, 17(3): 435-446. |
| [35] |
Klanderud K, Totland Ø. Diversity-Stability relationships of an alpine plant community under simulated environmental change. Arctic, Antarctic, and Alpine Research, 2008, 40(4): 679-684. DOI:10.1657/1523-0430(07-075)[KLANDERUD]2.0.CO;2 |
| [36] |
Pauli H, Gottfried M, Dullinger S, Abdaladze O, Akhalkatsi M, Alonso J L B, Coldea G, Dick J, Erschbamer B, Calzado R F, Ghosn D, Holten J I, Kanka R, Kazakis G, Kollár J, Larsson P, Moiseev P, Moiseev D, Molau U, Mesa J M, Nagy L, Pelino G, Puşcaş M, Rossi G, Stanisci A, Syverhuset A O, Theurillat J P, Tomaselli M, Unterluggauer P, Villar L, Vittoz P, Grabherr G. Recent plant diversity changes on Europe's mountain summits. Science, 2012, 336(6079): 353-355. DOI:10.1126/science.1219033 |