 2022, Vol. 42
2022, Vol. 42文章信息
- 贺慧丹, 李红琴, 祝景彬, 张法伟, 杨永胜, 王春雨, 罗方林, 李英年
- HE Huidan, LI Hongqin, ZHU Jingbin, ZHANG Fawei, YANG Yongsheng, WANG Chunyu, LUO Fanglin, LI Yingnian
- 祁连山南麓高寒湿地生态系统呼吸季节、年际动态及影响因素
- Seasonal and inter-annual dynamics and influencing factors of ecosystem respiration in an alpine wetland at the southern foot of Qilian Mountain
- 生态学报. 2022, 42(14): 5956-5965
- Acta Ecologica Sinica. 2022, 42(14): 5956-5965
- http://dx.doi.org/10.5846/stxb202108172274
-
文章历史
- 收稿日期: 2021-08-17
- 网络出版日期: 2022-03-23





2. 中国科学院西北高原生物研究所, 西宁 810001;
3. 洛阳师范学院生命科学学院, 洛阳 471934;
4. 中国科学院高原生物适应与进化重点实验室, 西宁 810001
2. Northwest Institute of Plateau Biology, Chinese Academy of Sciences, Xining 810001, China;
3. College of Life Sciences, Luoyang Normal University, Luoyang 471934, China;
4. Key Laboratory of Adaptation and Evolution of Plateau Biota, Chinese Academy of Sciences, Xining 810001, China
大气中温室气体浓度的升高造成“温室效应”, 持续的全球变暖对陆地生态系统碳循环产生广泛的影响[1—2]。湿地生态系统只占陆地表面积的5%—8%, 但却占陆地生态系统土壤有机碳的25%—30%, 湿地具有释放储存碳的潜力[3—4]。青藏高原的湿地大约占全国总湿地面积的三分之一, 其包含了中国湿地土壤有机质的30%—40%[5], 该地区正经历着强烈的地表温度升高[2]。由于高海拔、低温环境, 高寒湿地碳动态对全球气候变化非常敏感[6]。因此, 了解高寒湿地生态系统的碳平衡对于准确预测未来气候变化中的全球碳循环具有重要意义[7]。
生态系统呼吸作为生态系统最大的碳交换途径之一, 其微小的波动都会引起大气中CO2浓度的显著变化[8]。生态系统呼吸对气候变化有着较高的敏感性, 尤其是对全球变暖[2, 8], 研究结果显示气候变暖可能会导致生态系统呼吸的增强[9—10]、减弱或不变[11]。弄清气候环境因子对生态系统呼吸的影响机制, 对准确评估生态系统碳循环具有重要意义[9, 11]。全球变化背景下, 诸多地区的气温均呈升高趋势, 尤其是青藏高原表面的气温上升幅度要高于全球平均气温的上升幅度[11—12]。由于青藏高原气候严酷, 其陆地表层生态系统脆弱, 而生态系统呼吸对气候变化特别是温度的升高的响应极其敏感[13]。目前, 研究人员在青藏高原开展了一系列关于气候变化对高寒草地生态系统碳排放相关研究, 但并未得到一致结论[11, 14—15]。
研究表明, 近50年来青藏高原的年平均气温和降水量每十年增加0.3℃和9.1 mm[16—17]。然而, 关于青藏高原水热条件对生态系统呼吸影响的研究大多集中在高寒草甸和高寒灌丛上, 关于高寒湿地生态系统碳排放的研究相对薄弱[18—19]。此外, 以往对湿地碳排放动态及其影响机制的分析仅限于短期观测[20]。因此, 基于多年原位测量的研究可以进一步认识高寒湿地生态系统碳排放动态及其影响因素[11, 15]。并且, 冻土融化和冰川退缩可能会在青藏高原上形成更大或新的湿地[20]。高寒湿地范围的暖化扩张可能会导致土壤碳储量和区域碳平衡的变化[15, 21]。涡度协方差法(EC)是测量大气与各种生态系统之间碳收支的最有用方法之一, 本研究采用EC法测定了青藏高原祁连山南麓高寒湿地生态系统2004—2016年的碳排放数据。本研究的总体目标是:(1)量化高寒湿地生态系统呼吸(RE)在季节和年际尺度上的变异特征。(2)明晰高寒湿地RE在季节和年际尺度上的影响因素, 以提高预测未来气候变化背景下高寒湿地生态系统碳平衡响应特征的准确性。
1 材料与方法 1.1 试验地概况试验在青海海北高寒草甸生态系统国家野外科学观测研究站(海北站) 附近的高寒湿地中进行。海北站地处青藏高原东北隅祁连山北支冷龙岭东段南麓坡地(37°29′—37°45′N, 101°12′—101°23′E, 海拔3200—3600 m)。该地区属高原大陆性气候, 有明显的干冷(11月至次年4月)和暖湿(5月至10月)季节[22]。年平均气温约为-1.0℃, 最温暖的月份在7月, 温度约为10℃;最冷的月份在1月, 温度约为-15℃。年平均降水量约480 mm, 其中80%的降水量集中在暖湿季节。本研究的高寒湿地为泥炭沼泽地, 其泥炭层厚度在0.2—2.0 m, 土壤呈微碱性, 有机质含量丰富[23]。湿地植被组成较为贫乏, 以寒冷湿生、多年生地下芽、具有发达通气组织的草本植物莎草科、毛茛科为主, 以帕米尔苔草(Carex pamirensis)为建群种, 次优势种有华扁穗草(Blysmus sinocompressus)、黑褐苔草(Carex atrofusca)、乳白香青(Anaphalis lectea)等[24]。
1.2 研究材料与分析方法 1.2.1 通量和相关监测涡度相关观测系统设置于地势平坦、地形开阔, 且具有足够大“风浪区”的试验场内。通量塔高为2.5 m, 通量观测的主要仪器为开路式涡度相关观测系统和常规气象观测系统, 其中涡度相关系统包括三维超声风速仪(CSAT3, Campbell Scientific, Logan, USA)、红外气体分析仪(LI-7500, LI-COR Lincoln, USA) 和数据采集器(CR5000, Campbell Scientific, Logan, USA), 主要用于观测植被与大气界面的CO2通量, 数据采集频率为10 Hz, 每30 min输出平均值。常规气象观测系统主要用于连续气象要素采集, 数据采集(Model CR23X, Campbell Scientific, Logan, USA)时段与通量数据相同, 观测项目主要包括温度、湿度、辐射、风向、风速等。光量子通量密度(PPFD)传感器(L1190SB, LI-COR, Lincoln, USA)安装在高1.2 m的支架上。空气温度/湿度传感器(Model HMP45C, Vaisala, Helsinki, Finland)安装在防辐射罩内(Model 41002, RM Young Company, Michigan, USA)。土壤温度(Ts)主要是利用热电偶温度传感器(107-L, Campbell Scientific, Logan, USA)分别测定5、10、20 cm和40 cm处的Ts。因湿地地表积水, 未测量土壤含水量、土壤热通量等。本研究中, 将5—9月定义为生长季, 10月为枯黄期定义为生长季与非生长季的过渡时期, (1—4月及11—12月)定义为非生长季[23—24]。
1.2.2 数据处理和插补由于供电间断、探头结露或仪器异常等情况, 涡度相关系统采集到的原始数据缺失或异常现象十分普遍[25]。通量数据的处理按照ChinaFLUX推荐的通量数据处理标准流程进行[26], 在经过WPL校正后[27], 以|NEE| < 1.0 mg CO2 m-2 s-1为阈值标准, 再以10d数据为窗口, 3δ (标准差)为标准进行数据的异常剔除。针对夜间(PPFD < 10μmol m-2 s-1)湍流发展不充分, 舍去摩擦速度u* < 0.15 m/s所对应的通量数据。缺失的通量数据, 一般通过建立环境因子与有效通量数据的非线性经验方程进行插补。本研究夜间通量数据(Reco, n)采用其与5 cm深处土壤温度(Ts)的Van′t Hoff函数(方程(1))进行插补, 生长季白天的碳通量数据(NEE)则利用与PPFD的Michaelis-Menten函数(方程(2))进行插补[28—29]。
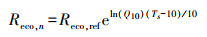
|
(1) |
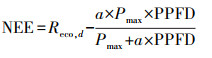
|
(2) |
式中, Reco, REf为生态系统在10℃时的参考呼吸速率, Q10是生态系统呼吸熵(呼吸随温度升高10℃所增加的倍数), 反映了生态系统呼吸对温度的敏感性。, Reco, d是生态系统白天的呼吸速率, a和Pmax分别为生态系统表观光量子效率和潜在最大CO2同化速率(即PPFD趋向极大时生态系统净光合速率)。以上参数均为方程拟合值。涡度相关系统无法直接测定生态系统呼吸(RE), 需利用公式外推得到。涡度相关系统观测的CO2通量为NEE, 将夜间呼吸函数外延至白天(Reco, d), 即可得生态系统的总呼吸量(RE), 日RE为白天呼吸(Reco, d)和夜间呼吸(Reco, n)之和。
1.2.3 数据分析分类回归树(classification and regression trees, CART)分析能够有效识别变量的相对重要性, CART是一种基于图形的非参数模型, 与传统分析方法相比更易于理解。本研究中采用CART用于识别气候环境因素(空气温度(Ta)、5 cm深处土壤温度(Ts)、生长季积温(GDD)、光量子通量密度(PPFD)、降水(PPT)、水汽饱和亏缺(VPD))对生态系统呼吸(RE)变化的关键控制作用。线性回归和相关分析用来探究RE的变异特征以及对气候环境因子的响应机制。本研究中所测定的数据利用SYSTAT 13.0软件对数据进行统计分析。
2 结果与分析 2.1 气候因子变化特征图 1可见, 高寒湿地在2004年1月至2016年12月的气候环境因子具有较高变异性, 整体上平均月(Ta、Ts、GDD、PPFD、PPT、RH、VPD)都呈单峰变化趋势, 但其峰值出现的月份并不一致。月Ta的峰值出现在7月((10.3 ±0.9)℃), 月Ts的峰值出现在8月((12.7 ±2.0)℃), 并且只在4月、5月和6月Ta高于Ts;年Ta、Ts的均值分别为(-1.1±0.4)℃和(2.8±0.5)℃, 整体上高寒湿地的Ts高于Ta。月GDD和月PPT在7月达到峰值, 分别为(317.3±26.4)℃d和(99.3±19.8) mm, 全年GDD和PPT的均值分别为(1076.9±78.5)℃d和(476.6±55.7) mm。月VPD和月RH分别在7月和9月达到峰值, 分别为(0.3±0.02) kPa和(77.3±1.7)%。

|
| 图 1 高寒湿地在2004—2016年平均月Ta, Ts, GDD, PPFD, PPT, RH, VPD Fig. 1 The average value of monthly air temperature (Ta), soil temperature (Ts), and aggregated growing season degree day (GDD); photosynthetic photon flux density (PPFD) and precipitation (PPT); relative humidity (RH) and vapor pressure deficit (VPD) of alpine wetland from 2004 to 2016 |
图 2可见, 高寒湿地在2004—2016年的月平均RE表现为单峰变化趋势, 在8月达到峰值((107.0±16.9)g C m-2月-1), 最小值出现在1月(17.1±4.1) g C m-2月-1)。图 3可见, 年RE具有一定变异性, 整体上表现为逐年升高的趋势(r2=0.36, P=0.031), 年RE均值为(608.9±65.6)g C m-2 a-1;湿地生长季RE的均值为(405.9±54.7)g C m-2季度-1, 湿地非生长季RE的均值为(152.2±30.8)g C m-2季度-1。

|
| 图 2 高寒湿地在2004—2016年的月平均RE Fig. 2 The average value of monthly RE of alpine wetland for the period 2004—2016 |

|
| 图 3 高寒湿地在2004—2016年的年RE、生长季RE、非生长季RE Fig. 3 The annual RE, growing seasonal RE, non-growing seasonal RE of alpine wetland for the period 2004—2016 |
高寒湿地生长季RE、非生长季RE的均值占全年RE的比例约为66.7%和25.0%, 线性回归分析表明生长季RE(r2=0.66, P=0.001)、非生长季RE(r2=0.47, P=0.01)与全年RE呈极显著正相关(图 4), 通过比较r2值, 说明生长季RE对全年RE有更强的控制作用。但, 本结果表明虽然高寒湿地非生长季RE占全年RE的比例相对较低, 但依然与全年RE呈极显著正相关, 暗示在未来高寒湿地生态系统碳排放的研究过程中不能忽略非生长季CO2通量排放的作用。

|
| 图 4 高寒湿地在2004—2016年的生长季RE、非生长季RE与年RE的关系 Fig. 4 The relationship between growing seasonal RE, non-growing seasonal RE and annual RE of alpine wetland for the period 2004—2016 |
图 5可见, 分类回归树(CART)表明全年-(月RE)、生长季-(月RE)、非生长季-(月RE)最主要的控制因素是Ts;线性回归分析表明, 通过比较r2值可知热量条件(Ts、Ta和GDD)是高寒湿地在不同时期月RE变化的主导因素, 其中高寒湿地的月Ts与全年-(月RE)、生长季-(月RE)、非生长季-(月RE)相关性最强(表 1)。此外, 通过进行逐步线性回归方程分析, 最终模型为:全年-(月RE) =3.63Ts+0.74GDD+33.25(r2=0.90, n=156, P < 0.001), 高寒湿地的Ts和GDD共同解释月RE变化的89.5%;生长季-(月RE) =4.98Ts+0.05GDD+25.65(r2=0.73, n=65, P < 0.05), 说明在生长季高寒湿地的Ts和GDD共同解释月RE变化的73.4%;非生长季-(月RE) =2.52Ts+0.03PPFD+23.37(r2=0.61, n=78, P < 0.01), 说明在非生长季高寒湿地的Ts和PPFD共同解释月RE变化的61.2%。综上所述, 月RE在全年、生长季、非生长季最主要的控制因子是Ts。

|
| 图 5 分类回归树分析高寒湿地的月RE与环境因子的关系 Fig. 5 Regression trees for monthly RE from environmental variables of alpine wetland PRE:消减误差比例Proportional Reduction in Error;MEAN:平均值Mean value;SD:标准差Standard deviation |
| 月份 Month |
全年-(月RE) Annual-(monthly respiration) | 生长季-(月RE) Growing season-(monthly respiration) | 非生长季-(月RE) Non-growing season-(monthly respiration) |
||||||||
| Linear Equation | r2 | P | Linear Equation | r2 | P | Linear Equation | r2 | P | |||
| 气温Ta(x1) | y=0.16x12+3.81x1+43.24 | 0.82 | 0.00 | y=7.27x1+26.53 | 0.50 | 0.00 | y=1.18x1+35.45 | 0.50 | 0.00 | ||
| 相对湿度RH(x2) | y=0.11x22-11.6x1+302.2 | 0.71 | 0.00 | y=3.34 x2-163.12 | 0.38 | 0.00 | y=0.40x2+2.45 | 0.09 | 0.009 | ||
| 光量子通量密度PPFD(x3) | y=0.21x3-12.96 | 0.27 | 0.00 | y=-0.17x3+142.93 | 0.10 | 0.00 | y=0.07x3+7.86 | 0.33 | 0.00 | ||
| 土壤温度Ts(x4) | y=4.90x4+36.16 | 0.88 | 0.00 | y=5.9x4+27.69 | 0.72 | 0.00 | y=2.80x4+32.02 | 0.52 | 0.00 | ||
| 降水PPT(x5) | y=0.66x5+24.66 | 0.72 | 0.00 | y=0.48x5+41.25 | 0.24 | 0.00 | y=0.44x5+22.68 | 0.20 | 0.00 | ||
| 水汽饱和亏缺VPD(x6) | y=284.51x6-4.24 | 0.56 | 0.00 | y=89.52x6+56.68 | 0.03 | 0.19 | y=98.14x6+12.55 | 0.38 | 0.00 | ||
| 生长季积温GDD(x7) | y=0.23x7+30.02 | 0.77 | 0.00 | y=0.18x7+42.39 | 0.49 | 0.00 | |||||
| Ta:气温air temperature;Ts:土壤温度soil temperature;GDD:生长季积温aggregated growing season degree day;PPFD:光量子通量密度photosynthetic photon flux density;PPT:降水precipitation;RH:相对湿度relative humidity;VPD:水汽饱和亏缺vapor pressure deficit | |||||||||||
在年际尺度上, 线性回归分析表明2004—2016高寒湿地生态系统的气候环境因子(Ta、Ts、GDD、PPFD、PPT、RH、VPD)与年RE没有显著相关性(P>0.05)。但是, 生长季GDD与生长季RE呈显著正相关(r2=0.42, P=0.015), 而生长季PPT与生长季RE呈显著负相关(r2=0.45, P=0.012)(图 6)。此外, 非生长季Ta(r2=0.77, P < 0.001)、PPT(r2=0.38, P=0.025)与非生长季RE呈显著正相关(图 6), 且非生长季Ta比PPT对高寒湿地在非生长季的碳排放控制作用更强(0.77>0.38)。

|
| 图 6 线性回归分析高寒湿地生长季RE与GDD、PPT的关系和非生长季RE与Ta、PPT的关系 Fig. 6 Linear regressions between growing season RE and GDD, PPT and linear regressions between non-growing season RE and Ta, PPT |
2004—2016年湿地的年RE均值为608.9 g C m-2 a-1, 高于附近高寒灌丛(437.4 g C m-2 a-1)和高寒草甸(513.6 g C m-2 a-1)的全年生态系统呼吸[13, 18], 三种植被类型相距较近, 具有相似的气候环境条件, 三种高寒草地碳排放的差异主要受到群落结构以及土壤理化性质差异的控制, 此外高寒湿地相对于高寒灌丛、草甸其土壤中含有更多未分解的有机质, 暗示高寒湿地生态系统的土壤呼吸可能对生态系统碳排放的贡献率更大[14, 18, 21]。过去的研究大多采用静态箱法观测的生态系统CO2排放通量, 主要包含植物、根系和土壤微生物呼吸以及极少部分土壤动物呼吸和化学氧化[14, 30], 主要观测了生态系统在生长季的碳排放, 而由于非生长季的碳排放较少, 往往忽略了对非生长季生态系统呼吸的研究[21]。本研究中, 虽然非生长季RE仅占全年RE的比例约25.0%, 但非生长季RE(r2=0.47, P=0.01)与全年RE呈极显著正相关(图 4), 由此说明高寒湿地在非生长季的碳排放对于整个生态系统的碳排放至关重要, 这是因为青藏高原高海拔、低温的特殊环境条件使得非生长季较为漫长, 并且由于土壤中含有大量未分解的有机质, 暗示在未来高寒生态系统碳排放的研究过程中应给予非生长季的碳排放足够的重视[14, 17, 30]。
由于不同生态系统的植被群落结构、土壤理化性质、水热条件等生物非生物因素的差异, 导致RE对热量条件的改变具有不同的响应方式[31—32]。有些研究表明, 温度的升高加强了生态系统的碳排放[33], 但有些研究表明, 温度的升高有利于提高自养呼吸, 但抑制了异氧呼吸, 致使RE对于增温的响应并不敏感, 暗示RE通过调节其内源成分的响应具有抵御气候变暖的潜力[34]。分类回归树分析表明, 高寒湿地的月RE的变化主要受到Ts的控制(图 5), 在生长季、非生长季Ts仍是月RE变化的主导因素(图 5)。许多研究表明, 高寒草地生态系统土壤温度显著影响CO2的释放和氮的矿化, 高寒生态系统土壤微生物生物量受到低温的限制[21, 25]。因此, 土壤温度成为生态系统呼吸的主导因素。这可能是因为较高的土壤温度刺激微生物活动和酶活性, 促进了土壤呼吸, 而高寒草地生态系统由于高海拔、低温使土壤中含有大量未分解的有机质, 所以造成高寒生态系统较高的土壤呼吸, 并且对土壤温度极其敏感, 因此使得土壤温度成为高寒草地生态系统的CO2排放的首要控制因素[1, 13, 18]。线性回归分析和逐步回归分析也表明高寒湿地生态系统在不同月份的碳排放更加依赖于土壤温度的变化, 暗示高寒湿地不同月份的土壤呼吸可能比植被自养呼吸对不同月份热量条件差异的响应更加敏感。
在年际尺度上, 线性回归分析表明2004—2016高寒湿地生态系统的气候环境因子(Ta、Ts、GDD、PPFD、PPT、RH、VPD)与年RE没有显著相关性(P>0.05), 这可能是由于时间尺度较短导致的[7, 10], 大量研究表明, 不同的时间尺度上气候环境因子可能对碳通量具有不同的影响[28]。但是, 生长季GDD与生长季RE呈显著正相关(r2=0.42, P=0.015)(图 6), 由于热量条件的累积效应是打破植被休眠的首要限制因素, 并对植被的物候发育和持续代谢生长至关重要[21], 在生长季植被生长发育旺盛, 说明整个生长季GDD的增加促进了高寒湿地整个生长季的碳排放。生长季PPT与生长季RE呈显著负相关(r2=0.45, P=0.012)(图 6), 大量研究表明湿地平均CO2排放通量与表面的积水存在显著负相关[14], 由于湿地常年积水, 使植被凋落物、土壤有机质的分解受到了抑制, 积水越多越不利于大气中的O2扩散到土壤中, 从而抑制了微生物的有氧呼吸, 进而不利于土壤有机质的分解, 使得减弱了CO2排放通量[7, 35];另一方面, 积水的增多抑制了微生物对土壤有机质的分解, 进而不利于土壤对植被生长代谢的营养供应, 从而不利于植被地上生物量的生长, 也间接减弱了植被自养呼吸, 最终使得CO2排放通量减少[35]。
非生长季Ta(r2=0.77, P < 0.001)、PPT(r2=0.38, P=0.025)与非生长季RE呈显著正相关(图 6), 在非生长季温度越高, 越利于微生物的分解, 促进了碳排放[14];在非生长季, 积雪对土壤温度具有保温作用, 减弱了土壤温度的变化, 使之处于一个较为稳定的阶段, 有利于土壤有机质的分解, 促进了高寒湿地的碳排放, 另外地表积雪的存在增加了大气中的水汽浓度, 地表有积雪时显著高于地表无积雪时的水汽通量, 由此提高了CO2交换速度, 进而促进了高寒湿地的CO2排放通量[15, 21, 23, 36]。综上, 由于在年际尺度上生长季、非生长季的热量条件对于CO2排放通量的促进作用, 暗示未来全球气候变化背景下温度的升高将会促进高寒湿地的CO2排放, 而生长季、非生长季PPT对于CO2排放通量的不同响应方式, 加之未来青藏高原PPT的变化的不确定性, 暗示在分析高寒生态系统碳排放对未来水热条件响应机制时需要更加谨慎。
4 结论(1) 高寒湿地生长季RE(P < 0.01)、非生长季RE(P < 0.01)与全年RE呈极显著正相关, 虽然生长季RE对全年RE有更强的控制作用, 但过去对非生长季RE的研究相对薄弱, 暗示在未来应加强对高寒湿地生态系统非生长季CO2排放通量的研究。
(2) 高寒湿地生态系统呼吸主要受到热量条件的关键控制, 而生长季、非生长季PPT对碳排放的差异化响应, 暗示在分析高寒湿地生态系统CO2排放对未来水热条件响应机制时需要更加谨慎。
| [1] |
吴琴, 胡启武, 曹广民, 李东. 高寒矮嵩草草甸冬季CO2释放特征. 生态学报, 2011, 31(18): 5107-5112. |
| [2] |
IPCC. Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge, UK: Cambridge University Press, 2021.
|
| [3] |
Gao J Q, Ouyang H, Lei G C, Xu X L, Zhang M X. Effects of temperature, soil moisture, soil type and their interactions on soil carbon mineralization in Zoigê alpine wetland, Qinghai-Tibet Plateau. Chinese Geographical Science, 2011, 21(1): 27-35. DOI:10.1007/s11769-011-0439-3 |
| [4] |
Bridgham S D, Megonigal J P, Keller J K, Bliss N B, Trettin C. The carbon balance of North American wetlands. Wetlands, 2006, 26(4): 889-916. DOI:10.1672/0277-5212(2006)26[889:TCBONA]2.0.CO;2 |
| [5] |
林春英, 李希来, 孙海松, 孙华方, 马程彪, 韩辉邦, 张宇鹏, 李成一. 黄河源高寒湿地有机碳组分对不同退化程度的响应. 草地学报, 2021, 29(7): 1540-1548. |
| [6] |
Barichivich J, Briffa K R, Myneni R B, Osborn T J, Melvin T M, Ciais P, Piao S L, Tucker C. Large-scale variations in the vegetation growing season and annual cycle of atmospheric CO2 at high northern latitudes from 1950 to 2011. Global Change Biology, 2013, 19(10): 3167-3183. DOI:10.1111/gcb.12283 |
| [7] |
Shen M G, Piao S L, Jeong S J, Zhou L M, Zeng Z Z, Ciais P, Chen D L, Huang M T, Jin C S, Li L Z X, Myneni R B, Yang K, Zhang G X, Zhang Y G, Yao T D. Evaporative cooling over the Tibetan Plateau induced by vegetation growth. Proceedings of the National Academy of Sciences of the United States of America, 2015, 112(30): 9299-9304. DOI:10.1073/pnas.1504418112 |
| [8] |
Luo Y Q. Terrestrial carbon-cycle feedback to climate warming. Annual review of Ecology, Evolution, and Systematics, 2007, 38: 683-712. DOI:10.1146/annurev.ecolsys.38.091206.095808 |
| [9] |
李愈哲, 樊江文, 胡中民, 邵全琴. 温性草原利用方式对生态系统碳交换及其组分的影响. 生态学报, 2018, 38(22): 8194-8204. |
| [10] |
Wang N, Quesada B, Xia L L, Butterbach-Bahl K, Goodale C L, Kiese R. Effects of climate warming on carbon fluxes in grasslands-a global meta-analysis. Global Change Biology, 2019, 25(5): 1839-1851. DOI:10.1111/gcb.14603 |
| [11] |
Chen J, Luo Y Q, Xia J Y, Shi Z, Jiang L F, Niu S L, Zhou X H, Cao J J. Differential responses of ecosystem respiration components to experimental warming in a meadow grassland on the Tibetan Plateau. Agricultural and Forest Meteorology, 2016, 220: 21-29. DOI:10.1016/j.agrformet.2016.01.010 |
| [12] |
Chen N, Zhang Y J, Zhu J T, Cong N, Zhao G, Zu J X, Wang Z P, Huang K, Wang L, Liu Y J, Zheng Z T, Tang Z, Zhu Y X, Zhang T, Xu M J, Di Y P, Chen Y. Multiple-scale negative impacts of warming on ecosystem carbon use efficiency across the Tibetan Plateau grasslands. Global Ecology and Biogeography, 2021, 30(2): 398-413. DOI:10.1111/geb.13224 |
| [13] |
Li H Q, Zhang F W, Li Y N, Wang J B, Zhang L M, Zhao L, Cao G M, Zhao X Q, Du M Y. Seasonal and inter-annual variations in CO2 fluxes over 10 years in an alpine shrubland on the Qinghai-Tibetan Plateau, China. Agricultural and Forest Meteorology, 2016, 228-229: 95-103. DOI:10.1016/j.agrformet.2016.06.020 |
| [14] |
周文昌, 崔丽娟, 王义飞, 李伟, 康晓明. 若尔盖高原沼泽湿地CO2排放时空变化特征. 生态学报, 2021, 41(7): 2652-2662. |
| [15] |
Hao Y B, Cui X Y, Wang Y F, Mei X R, Kang X M, Wu N, Luo P, Zhu D. Predominance of precipitation and temperature controls on ecosystem CO2 exchange in Zoige alpine wetlands of southwest China. Wetlands, 2011, 31(2): 413-422. DOI:10.1007/s13157-011-0151-1 |
| [16] |
Chen X P, Wang G X, Huang K W, Hu Z Y, Song C L, Liang Y M, Wang J, Song X Y, Lin S. The effect of nitrogen deposition rather than warming on carbon flux in alpine meadows depends on precipitation variations. Ecological Engineering, 2017, 107: 183-191. DOI:10.1016/j.ecoleng.2017.07.018 |
| [17] |
Piao S L, Tan K, Nan H J, Ciais P, Fang J Y, Wang T, Vuichard N, Zhu B. Impacts of climate and CO2 changes on the vegetation growth and carbon balance of Qinghai-Tibetan grasslands over the past five decades. Global and Planetary Change, 2012, 98-99: 73-80. DOI:10.1016/j.gloplacha.2012.08.009 |
| [18] |
Kato T, Tang Y H, Gu S, Hirota M, Du M Y, Li Y N, Zhao X Q. Temperature and biomass influences on interannual changes in CO2 exchange in an alpine meadow on the Qinghai-Tibetan Plateau. Global Change Biology, 2006, 12(7): 1285-1298. DOI:10.1111/j.1365-2486.2006.01153.x |
| [19] |
Kang X M, Wang Y F, Chen H, Tian J Q, Cui X Y, Rui Y C, Zhong L, Kardol P, Hao Y B, Xiao X M. Modeling carbon fluxes using multi-temporal MODIS imagery and CO2 eddy flux tower data in Zoige alpine wetland, south-west China. Wetlands, 2014, 34(3): 603-618. DOI:10.1007/s13157-014-0529-y |
| [20] |
Zhang Y, Wang G X, Wang Y B. Changes in alpine wetland ecosystems of the Qinghai-Tibetan plateau from 1967 to 2004. Environmental Monitoring and Assessment, 2011, 180(1/4): 189-199. |
| [21] |
Song W M, Wang H, Wang G S, Chen L T, Jin Z N, Zhuang Q L, He J S. Methane emissions from an alpine wetland on the Tibetan Plateau: neglected but vital contribution of the nongrowing season. Journal of Geophysical research, 2015, 120(8): 1475-1490. DOI:10.1002/2015JG003043 |
| [22] |
李英年, 赵亮, 赵新全, 王勤学, 张法伟. 高寒湿地生态系统土壤有机物质补给及地-气CO2交换特征. 冰川冻土, 2007, 29(6): 940-946. DOI:10.3969/j.issn.1000-0240.2007.06.013 |
| [23] |
Zhao L, Li J, Xu S, Zhou H, Li Y, Gu S, Zhao X. Seasonal variations in carbon dioxide exchange in an alpine wetland meadow on the Qinghai-Tibetan Plateau. Biogeosciences, 2010, 7(4): 1207-1221. DOI:10.5194/bg-7-1207-2010 |
| [24] |
Zhang F W, Liu A H, Li Y N, Zhao L, Wang Q X, Du M Y. CO2 flux in alpine wetland ecosystem on the Qinghai-Tibetan Plateau, China. Acta Ecologica Sinica, 2008, 28(2): 453-462. DOI:10.1016/S1872-2032(08)60024-4 |
| [25] |
Cao S K, Cao G C, Chen K L, Han G Z, Liu Y, Yang Y F, Li X D. Characteristics of CO2, water vapor, and energy exchanges at a headwater wetland ecosystem of the Qinghai Lake. Canadian Journal of Soil Science, 2019, 99(3): 227-243. DOI:10.1139/cjss-2018-0104 |
| [26] |
李春, 何洪林, 刘敏, 苏文, 伏玉玲, 张雷明, 温学发, 于贵瑞. ChinaFLUX CO2通量数据处理系统与应用. 地球信息科学, 2008, 10(5): 557-565. DOI:10.3969/j.issn.1560-8999.2008.05.002 |
| [27] |
Webb E K, Pearman G I, Leuning R. Correction of flux measuREments for density effects due to heat and water vapour transfer. Quarterly Journal of the Royal Meteorological Society, 1980, 106(447): 85-100. DOI:10.1002/qj.49710644707 |
| [28] |
Li H Q, Zhang F W, Zhu J B, Guo X W, Li Y K, Lin L, Zhang L M, Yang Y S, Li Y N, Cao G M, Zhou H K, Du M Y. Precipitation rather than evapotranspiration determines the warm-season water supply in an alpine shrub and an alpine meadow. Agricultural and Forest Meteorology, 2021, 300: 108318. DOI:10.1016/j.agrformet.2021.108318 |
| [29] |
Yu G R, Zhang L M, Sun X M, Fu Y L, Wen X F, Wang Q F, Li S G, REn C Y, Song X, Liu Y F, Han S J, Yan J H. Environmental controls over carbon exchange of three forest ecosystems in eastern China. Global Change Biology, 2008, 14(11): 2555-2571. DOI:10.1111/j.1365-2486.2008.01663.x |
| [30] |
Peng F, You Q G, Xu M H, Guo J, Wang T, Xue X. Effects of warming and clipping on ecosystem carbon fluxes across two hydrologically contrasting years in an alpine meadow of the Qinghai-Tibet Plateau. PLoS One, 2014, 9(10): e109319. DOI:10.1371/journal.pone.0109319 |
| [31] |
Li H Q, Zhu J B, Zhang F W, He H D, Yang Y S, Li Y N, Cao G M, Zhou H K. Growth stage-dependant variability in water vapor and CO2exchanges over a humid alpine shrubland on the northeastern Qinghai-Tibetan plateau. Agricultural and Forest Meteorology, 2019, 268: 55-62. DOI:10.1016/j.agrformet.2019.01.013 |
| [32] |
DeLucia E H, Drake J E, Thomas R B, Gonzalez-Meler M. FoREst carbon use efficiency: is respiration a constant fraction of gross primary production?. Global Change Biology, 2007, 13(6): 1157-1167. DOI:10.1111/j.1365-2486.2007.01365.x |
| [33] |
De Deyn G B, Cornelissen J H C, Bardgett R D. Plant functional traits and soil carbon sequestration in contrasting biomes. Ecology Letter, 2008, 11(5): 516-531. DOI:10.1111/j.1461-0248.2008.01164.x |
| [34] |
Duan M, Li A D, Wu Y H, Zhao Z P, Peng C H, Deluca T H, Sun S Q. Differences of soil CO2 flux in two contrasting subalpine ecosystems on the eastern edge of the Qinghai-Tibetan Plateau: a four-year study. Atmospheric Environment, 2019, 198: 166-174. DOI:10.1016/j.atmosenv.2018.10.067 |
| [35] |
Chimner R A, Cooper D J. Influence of water table levels on CO2emissions in a colorado subalpine fen: an in situ microcosm study. Soil Biology and Biochemistry, 2003, 35(3): 345-351. DOI:10.1016/S0038-0717(02)00284-5 |
| [36] |
赵亮, 李英年, 赵新全, 徐世晓, 唐艳鸿, 于贵瑞, 古松, 杜明远, 王勤学. 青藏高原3种植被类型净生态系统CO2交换量的比较. 科学通报, 2005, 50(9): 926-932. DOI:10.3321/j.issn:0023-074X.2005.09.015 |