 2022, Vol. 42
2022, Vol. 42文章信息
- 陈青, 汪志如, 汤文超, 丁慧芳, 廖劲扬, 王亚芳, 王文娟
- CHEN Qing, WANG Zhiru, TANG Wenchao, DING Huifang, LIAO Jinyang, WANG Yafang, WANG Wenjuan
- 鄱阳湖周边稻田对白鹤的承载力
- Carrying capacity of Siberian cranes (Leucogeranus leucogeranus) in the rice fields around Poyang Lake
- 生态学报. 2022, 42(13): 5340-5347
- Acta Ecologica Sinica. 2022, 42(13): 5340-5347
- http://dx.doi.org/10.5846/stxb202108102197
-
文章历史
- 收稿日期: 2021-08-10
- 网络出版日期: 2022-03-18

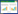




2. 江西省林业科学院, 南昌 330032;
3. 南昌大学空间科学与技术研究院, 南昌 330031;
4. 江西鄱阳湖湿地保护与恢复国家长期科研基地和江西鄱阳湖湿地生态系统国家定位观测研究站, 南昌 330031;
5. 南昌大学鄱阳湖环境与资源利用教育部重点实验室, 南昌 330031
2. Jiangxi Academy of Forestry, Nanchang 330032, China;
3. Institute of Space Science and Technology, Nanchang University, Nanchang 330031, China;
4. Jiangxi Poyang Lake Wetland Conservation and Restoration National Permanent Scientific Research Base, National Ecosystem Research Station of Jiangxi Poyang Lake Wetland, Nanchang 330031, China;
5. Key Laboratory of Poyang Lake Environment and Resource Utilization, Ministry of Education, Nanchang University, Nanchang 330031, China
承载力最初由牧场研究人员提出用于管理牲畜, 指一个特定区域在没有退化的情况下能够支持的牲畜数量[1-2]。后来承载力在野生动物管理中被运用, 考虑资源可用性以及捕食者的影响, 常常将其与逻辑生长曲线的最大理论值相联系[1, 3-4]。生态学家将承载力定义为在一定环境条件下所能支持的最大动物数量, 常常利用单一因素以及各种因素综合进行研究, 如气候因子、资源丰富度、捕食者、疾病因子和竞争者等[5-6]。对于迁徙鸟类来说, 环境承载力主要用于预测特定时间下, 特定地点的食物丰富度所能支持的最大鸟类生存的数量[7]。营养承载力和空间承载力是生态学研究中较为常见的两种估算特定种群的环境承载力的方法[8]。其中, 营养承载力的估算基础是目标种群所需的能量需求和所处生境所能提供的营养, 即在假设所有个体都相同并且在无干扰的情况下, 特定生境提供的可利用能量与个体某一时间段内平均消耗的总能量的比值[8-9]。空间承载力是以种群所处生境总格局为基础, 考虑食物密度、食物可获得性以及人为干扰等因素, 建立种群生存空间模型等方法来估计环境承载力[8]。环境承载力的评估对野生动物的有效保护和科学管理至关重要。目前, 野生动物环境承载力的研究主要集中在许多兽类和鸟类中, 例如马鹿(Cervus elaphus nelson)[10-11]、扭角羚(Budorcas taxicolor)[12]、沙丘鹤(Grus canadensis)[13]、丹顶鹤(G. japonensis)[14]和鸻鹬类[15]等。
白鹤(Leucogeranus leucogeranus)为IUCN极危物种, 国家Ⅰ级保护动物, 全球种群数量约为3500-4000只, 其中, 约98%的个体在鄱阳湖越冬[16]。随着长江中下游流域其它湖泊生态环境质量的退化, 鄱阳湖已成为流域内唯一适合白鹤生存的地方[17-18], 因此, 鄱阳湖的生态环境质量对于白鹤的生存而言至关重要。白鹤主要在浅水区取食, 对水域和沼泽有较强依赖性[19]。鄱阳湖优势沉水植物-苦草(Vallisneria spp.)冬芽是白鹤主要传统食物[20]。近年来, 由于夏季洪涝和秋季干旱频繁发生、水质下降、挖沙等因素的影响, 鄱阳湖沉水植被退化严重[21-24]。食物资源丰富度对鸟类的数量与分布影响较大[25-26]。鄱阳湖沉水植被的退化导致白鹤的觅食生境和食性发生变化[20, 27]。近年来, 大量白鹤离开自然生境, 前往稻田、藕田等人工生境觅食[28-29]。
鄱阳湖周边存在大面积的稻田, 秋季收割之后散落在稻田里的稻谷为白鹤等越冬鸟类提供了丰富的食物来源。近年来越来越多的鸟类前往鄱阳湖周边稻田中觅食, 包括灰鹤(G. grus)、白头鹤(G. monacha)和雁鸭类等[30-32]。稻田等人工生境已经成为白鹤等越冬候鸟重要的觅食生境, 在白鹤等鸟类的保护方面发挥的作用日益突出。然而, 目前鄱阳湖周边稻田对鸟类, 尤其是白鹤的承载力方面的研究却尚未见报道。相关研究可为白鹤等鸟类的保护和管理提供科学依据, 并且可以评估农业用地在白鹤保护中的作用。
为此, 本研究主要探讨鄱阳湖周边稻田对鸟类尤其是白鹤的承载力。已有研究表明白鹤在稻田里主要取食稻谷, 同时也取食少量的其它植物[33]。本文假设白鹤等越冬鸟类在稻田中仅以稻谷为食, 并且每日摄入的能量即为每日消耗的能量。本文首先调查鄱阳湖周边稻田散落稻谷的生物量, 测量了稻谷的营养成分含量。然后, 依靠遥感技术, 估算鄱阳湖周边稻田的总面积, 进而依据稻谷生物量和能量, 计算出稻田能提供的总能量。最后, 根据白鹤等以稻谷为食的常见越冬鸟类的数量占比、日能量消耗、越冬时长、以及稻田能提供的总能量, 计算出鄱阳湖周边稻田对所有鸟类以及白鹤的环境承载力。
1 研究区域鄱阳湖(115°48′-116°44′E, 28°25′-29°45′N)位于长江中下游南岸, 江西省北部, 是中国最大的淡水湖泊, 以生态和经济重要性以及剧烈的水位变化而闻名[28, 34]。作为东亚重要的候鸟越冬地, 鄱阳湖每年约有42万只水鸟在此越冬[35]。鄱阳湖水位主要受流域内降水的影响, 同时也受长江水位的影响, 流域内季节性降水驱使鄱阳湖呈现出明显的季节性水位波动[34]。4月至9月为丰水期, 鄱阳湖流域降水量增多, 水位升高, 洪泛平原被淹没形成一个面积超过3000 km2的单一湖泊;10月至次年3月为枯水期, 鄱阳湖水位下降, 湖泊面积缩小至1000 km2, 众多蝶形子湖泊出露[34]。丰水期的洪水给鄱阳湖带来了丰富的有机物和营养物质, 促进了生物的生长;枯水期水位下降后, 露出大面积的草地、浅水和泥滩等生境, 为水鸟提供了丰富多样的觅食生境。这种特殊的水文节律特征使鄱阳湖为越冬候鸟提供了丰富的食物资源和栖息生境[36]。
2 研究方法 2.1 稻谷生物量调查根据课题组以往的调查以及保护区工作人员的建议, 本研究选择有较多白鹤等越冬水鸟觅食的水稻田作为稻谷生物量调查样点。同时为了反映不同区域稻谷生物量的变化, 本研究在鄱阳湖各个不同方向上共设置了5个调查样点, 样点分别位于新建县恒湖农场(116°4′27.85″E, 28°59′52.47″N)、新建县朱港农场(116°10′12.76″E, 28°57′29.95″N)、南昌县五星垦殖场(116°18′29.55″E, 28°45′29.54″N)、余干县康山垦殖场(116°27′26.36″E, 28°50′4.14″N)和都昌县多宝乡(116°7′10.06″E, 29°22′43.81″N)(图 1)。

|
| 图 1 鄱阳湖周边稻田散落稻谷采样点分布图 Fig. 1 Distribution of rice sampling sites in rice fields around Poyang Lake |
为真实反映稻谷生物量情况, 本研究尽量在稻谷刚收割后以及白鹤等鸟类到达前采集稻谷样本。因此, 本研究于2020年11月到12月, 对每个样点进行散落稻谷生物量的调查。每个样点随机设置15个样方, 用镊子收集50 cm×50 cm范围内所有的稻谷颗粒, 装至密封袋内, 贴好标签。将采集好的稻谷样品带回实验室洗净, 放入烘箱中60℃烘至恒重, 称量每份样品的重量。根据样方的面积计算出稻谷生物量, 然后将所有样方的稻谷生物量求平均值, 得出鄱阳湖周边稻田的平均稻谷生物量。
2.2 稻谷营养成分测定分别使用酸水解法、蒽酮比色法、凯氏定氮法、酸碱水解法、灼烧减量法和索氏抽提法测定稻谷中淀粉、粗糖分、粗蛋白、粗纤维、粗灰分和粗脂肪的含量。采用营养计算公式, 根据测定的稻谷中的营养物质含量计算出单位重量稻谷含有的能量[37]:
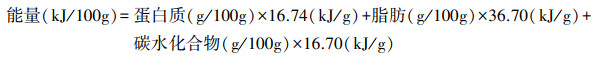
|
(1) |
觅食地与夜宿地的距离是影响鸟类觅食生境选择的重要因子[38]。鸟类为了减少飞行带来的能量损耗, 一般会选择离夜宿地较近的地方觅食。本研究将白鹤的适宜觅食范围设定为距离最近湖区10 km范围内, 该范围的确定主要是基于2方面原因。一方面, 课题组前期记录的白鹤稻田分布点数据显示, 白鹤的分布点离主湖区的最远距离为10 km。另一方面, Nilsson等[38]记录灰鹤的稻田觅食地离夜宿地的最远距离为11 km。人类干扰也是影响鸟类数量和分布的重要因子[39]。然而, 已有研究表明人为干扰对鄱阳湖稻田鸟类多样性影响较少, 白鹤数量甚至与道路距离、人工建筑和人为干扰呈现正相关[40], 因此本研究并未排除人工建筑和道路附近的稻田。
在中国资源卫星应用中心官网(http://www.cresda.com/CN/)获取2020年12月22日鄱阳湖区GF-1-WFV3 16 m分辨率遥感影像, 利用遥感影像处理软件ENVI(The Environment for Visualizing Images)进行辐射标定、大气校正、几何校正及影像拉伸(2%)等预处理, 降低影像目视解译过程的误差。利用线型感兴趣区域(region of interest, ROI)确定环湖区10 km的范围, 然后在该区域内建立一定数量的农田、建筑、道路、水域、林地、沙地等面型ROI作为训练样本, 保持各区域类型可分离阈值均为1.8以上。由于建筑与道路的分离阈值低于1.8, 难以准确区分, 因此将建筑与道路作为一个类参与训练。沙地与稻田的分离阈值也低于1.8, 考虑到稻田基本位于湖区外围, 沙地主要在湖区内部, 因此将整个湖区内部裁剪以降低沙地对稻田评估的影响。基于建立的ROI区, 利用最大似然法统计分类鄱阳湖周边的农田、建筑、水域和林地区域。最后使用混淆矩阵(confusion matrices)对影像分类结果进行精度评价, 并通过像元大小计算稻田面积。
2.4 稻田承载力估算将稻谷生物量、单位重量稻谷含有的能量以及稻田总面积三个因子相乘, 得出鄱阳湖周边稻田所能提供的总能量(T)。查阅文献, 获得鄱阳湖周边稻田水鸟种类和数量的数据。鄱阳湖周边稻田以稻谷为主要食物的鸟类主要包括6种, 即白鹤、灰鹤、豆雁(Anser fabalis)、鸿雁(A. cygnoid)、灰雁(A. anser)和小天鹅(Cygnus columbianus)[29, 40-41]。2016-2017年冬季和2017-2018年冬季在鄱阳湖7条稻田样线内记录到的鸟类平均数量和占比情况为:白鹤2482只(占比7.65%), 灰鹤6995只(占比21.56%), 豆雁8152只(占比25.13%), 鸿雁7033只(占比21.68%), 灰雁2518只(占比7.76%), 小天鹅5261只(占比16.22%)[31, 42]。
假设鄱阳湖周边稻田中的鸟类每日摄入的能量即为每日消耗的能量。参考Pearse等[13]的方法, 将基础代谢率、脂质生产能耗、自由生活能耗和觅食飞行能耗作为鸟类个体日能量消耗。将静息代谢率作为基础代谢率, 根据水禽的异速生长方程计算静息代谢率[43]:
鹤类:

|
(2) |
雁鸭类:
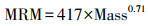
|
(3) |
其中, MRM为静息代谢率(kJ/day), Mass为体重(kg)。鸿雁体重约为3.55 kg, 豆雁体重约为2.83 kg, 灰雁体重约为2.94 kg, 小天鹅体重约为5.01 kg, 白鹤体重约为4.75 kg, 灰鹤体重约为4.35 kg[44-45]。
脂质生产能耗是鸟类产生脂肪所消耗的能量, 一般为基础代谢率的3倍[13]。自由生活能耗主要指在觅食地发生的所有行为所消耗的能量总和, 一般为基础代谢率的2倍[13]。觅食飞行能耗主要指清晨从夜宿地飞往稻田觅食, 傍晚从稻田飞回夜宿地所消耗的能量。关于觅食飞行能耗的计算, 先利用以下公式计算出单位时间飞行所需的能量(Pmet)[46]:

|
(4) |
以适宜觅食地离湖区的最远距离10 km作为鸟类的飞行距离。将飞行距离除以鸟类飞行速度, 得出飞行时间。其中鹤类的飞行速度为50 -60 km/h(取均值55.0 km/h)[47], 小天鹅飞行速度为25.5-32.8 km/h(取均值29.2 km/h)[48], 豆雁飞行速度为68 km/h [49], 鸿雁飞行速度为31 km/h[50], 灰雁飞行速度为47.9 km/h[51]。最后根据飞行时间和单位时间飞行所需的能量, 计算出觅食飞行消耗的总能量。
根据鸟类的日能量摄入和在鄱阳湖的越冬时长(11月-3月, 150天), 参考Hobbs等[10]的方法进行修改后, 得到以下公式计算鄱阳湖周边稻田的鸟类承载力:
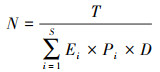
|
(5) |
其中, N为稻田能承载的鸟类总数量, T为稻田所能提供的总能量, Ei为物种i的日能量摄入, Pi为物种i的数量占所有鸟类总数量的比例, D为越冬时长, S为鸟类物种数(鄱阳湖周边稻田中以稻谷为主食的鸟类种数, 即6种)。将稻田承载的鸟类总数量乘以各种鸟类的数量占比, 即可得到稻田能承载的每种鸟类的数量。
3 研究结果 3.1 稻谷生物量和能量在此次调查中, 1号样点每个样方的稻谷平均干重为1.375 g, 2号样点每个样方的平均干重为1.890 g, 3号样点每个样方的平均干重为1.044 g, 4号样点每个样方的点平均干重为3.263 g, 5号样点每个样方的平均干重为0.546 g。最终得到稻田的平均生物量约为6.494 g/m2。测定两份稻谷的营养成分结果如表 1。利用公式(1)计算得到单位稻谷所含能量为1101.054 kJ/100 g。
| 编号 Number |
淀粉 Amylum/(g/100g) |
粗糖分 Crude sugar/(g/100g) |
粗蛋白 Crude protein/(g/100g) |
粗纤维 Crude fiber/(g/100g) |
粗灰分 Crude ash/(g/100g) |
粗脂肪 Crude fat/(g/100g) |
| 稻谷1 Rice 1 | 62.60 | 0.80 | 6.85 | 0.22 | 3.50 | 0.07 |
| 稻谷2 Rice 2 | 54.60 | 0.23 | 6.19 | 0.11 | 3.08 | 0.09 |
| 均值 Mean value | 58.60 | 0.52 | 6.52 | 0.12 | 3.29 | 0.08 |
利用ENVI 5.3软件中最大似然法模块, 对鄱阳湖周边稻田区域进行地物分类, 结果如图 2所示。分析结果显示鄱阳湖周边10 km范围内的稻田占据7751791个像元, 分类精度为99.81%。根据像元大小(16 m×16 m), 计算得出鄱阳湖周边10 km范围内的稻田总面积为1984.46 km2。

|
| 图 2 环鄱阳湖周边10 km范围内的稻田 Fig. 2 The rice fields with 10 km around Poyang Lake |
基于稻谷生物量、单位稻谷能量和稻田面积, 计算得到鄱阳湖周边稻田能提供的总能量T为1.42×1011 kJ。通过公式(2)(3)(4)计算得到白鹤的日能量摄入为8153.26 kJ, 灰鹤的日能量摄入为7639.35 kJ, 豆雁的日能量摄入为5307.35 kJ, 鸿雁的日能量摄入为6337.45 kJ, 灰雁的日能量摄入为5483.88 kJ, 小天鹅的日能量摄入为8116.95 kJ。基于文献中稻田的水鸟数量, 以及每种鸟类的日能量摄入, 根据公式(5)计算得到稻田能承载的鸟类总数量N为140860只, 其中稻田能承载的白鹤数量为10775只。
4 讨论以前白鹤主要在鄱阳湖的浅水和泥滩中挖掘苦草冬芽为食[52]。近年来, 受沉水植被退化和苦草冬芽锐减的影响, 白鹤的觅食生境和食性发生了变化。例如, 2010年, 夏季洪涝导致鄱阳湖苦草冬芽锐减, 白鹤首次出现在草滩上觅食, 其主要食物转变为下江委陵菜(Potentilla limprichtii)和老鸦瓣(Tulipa edulis)[22, 27]。2015-2016年冬季, 白鹤的觅食生境进一步改变, 大量白鹤离开浅水生境, 前往稻田、藕塘等人工生境觅食[29]。稻田中白鹤的食物组成也已发生变化, 农作物已成为白鹤的重要食物来源[20]。除白鹤外, 大量的鸿雁、小天鹅等以苦草冬芽为食的水鸟也转移至稻田中觅食[31]。因此, 稻田和藕塘在为白鹤等以苦草冬芽为食的鸟类保护方面发挥的作用日益突出。同时, 稻田也是灰鹤、豆雁等广谱食性鸟类的主要觅食生境[31]。与鄱阳湖类似, 受湿地生态环境质量退化的影响, 稻田已成为中国[53]及其它国家[54-55]水鸟的重要觅食生境。
本研究结果表明鄱阳湖周边10 km范围内的稻田总面积为1984.46 km2, 能承载的鸟类数量多达142860只, 其中能承载的白鹤数量多达10775只。能承载的白鹤数量远高于现有白鹤的总数量(3500-4000)[16], 这个结果表明鄱阳湖周边稻田能为白鹤等越冬水鸟提供较为充足的食物资源。
近些年, 为保护白鹤, 民间组织在南昌五星垦殖场的藕田建立了五星白鹤保护小区, 种植约66.67 hm2藕供白鹤食用。此外, 政府部门在余干县康山垦殖场插旗州赎买500余亩未收割稻田供鹤群食用。2020年冬季, 在五星白鹤保护小区记录到的白鹤数量最高值达到2900只, 占全球总数量的77%左右, 在余干插旗州的稻田记录到白鹤1096只, 而在其他稻田和藕塘并未记录到大群白鹤。以上调查结果表明虽然鄱阳湖周边稻田散落稻谷能承载大量白鹤, 但白鹤并未主要在已收割的稻田中取食, 而是集中在五星白鹤保护小区和余干插旗州这两个人为预留的、食物丰富度高的地方。鸟类集群的密度过大, 会增加疾病传播的风险[56]。考虑到鄱阳湖周边稻田具有较丰富的散落稻谷, 本文建议适当减少人为预留食物量, 让白鹤疏散至周边稻田, 以减少疾病传播的潜在风险。
此外, 应正确看待稻田等人工生境在白鹤保护中的作用, 它们只能作为白鹤在自然生境食物短缺时的避难所和临时觅食地, 不能替代自然生境[20, 28]。实际上相比稻田等人工生境, 白鹤更偏好自然生境, 一旦自然生境中具有较高的苦草冬芽丰富度, 白鹤便会返回自然生境取食。例如, 2015-2017年冬季, 苦草冬芽锐减导致大量白鹤前往稻田和藕田觅食, 2018年冬季苦草冬芽丰富度回升, 白鹤返回浅水生境取食[20, 28]。那么白鹤为何偏好自然生境?一般而言, 农作物含有的能量和营养成分与自然生境的食物类似, 甚至高于自然生境中的食物, 并且农作物具有更低的纤维素和灰分[55]。因此, 食物的营养含量可能不是影响白鹤取食的主要原因。鄱阳湖周边稻田的人为干扰强度较大, 这被认为是雁类受限于自然生境, 不能很好地利用人工生境中质量和丰富度高的农作物资源的重要原因[57-58]。较高的人为干扰也使得白鹤在藕田的警戒行为较自然生境中高出一倍[59]。因此, 人工生境中较强的人为干扰可能是白鹤偏好选择自然生境的主要原因。尽管本研究结果表明鄱阳湖周边稻田等人工生境能承载大量的白鹤, 但笔者呼吁应把恢复自然生境中的沉水植被放在首要位置, 以便为白鹤提供安全、高质量的觅食生境。
需要注意本研究存在一定的局限性。首先, 由于缺少鸟类的日能量摄入数据, 本文使用鸟类的日能量消耗作为日能量摄入, 并基于异速生长公式计算得出日能量消耗, 因此, 本文使用的日能量摄入数据并不准确, 这将导致计算结果存在一定偏差。其次, 文章假设稻田中所有的稻谷都能被鸟类利用。然而, 农民在稻田中散养的大量家禽会降低稻谷的丰富度[58], 鄱阳湖冬季阴雨连绵也会导致部分稻谷霉变, 进而减少稻谷的可利用性。此外, 本文未考虑鸟类取食时的放弃密度(giving-up density), 即当觅食地的食物密度低于某一阈值时, 鸟类需要耗费大量的能量寻找食物, 因此它们会放弃该觅食地, 转移至其它区域取食[60-61]。以上因素都可能导致本文计算出的鸟类承载力数量偏高。最后, 本文没有考虑人为干扰的影响, 没有排除居民区、道路等周边人为干扰强度大的稻田区域, 这也可能导致计算结果偏高。除以上因素外, 本研究未考虑不同稻谷品种和不同年份间稻谷丰富度的变化的影响。在后续的研究中, 应当调查稻田中主要觅食鸟类的日能量摄入, 排除人为干扰强度大的稻田区域, 以及考虑家禽取食、稻谷霉变、放弃密度、稻谷品种和稻谷产量的年际间变化等因素的影响, 从而增强研究结果的准确性。
| [1] |
Young C C. Defining the range: the development of carrying capacity in management practice. Journal of the History of Biology, 1998, 31(1): 61-83. DOI:10.1023/A:1004205522191 |
| [2] |
Price D. Carrying capacity reconsidered. Population and Environment, 1999, 21(1): 5-26. |
| [3] |
Mcleod S R. Is the concept of carrying capacity useful in variable environments?. Oikos, 1997, 79(3): 529-542. DOI:10.2307/3546897 |
| [4] |
Hixon M A. Carrying capacity//Jørgensen S E, Fath B D, eds. Encyclopedia of Ecology. Oxford: Elsevier Press, 2008: 528-530.
|
| [5] |
Firmansyah F, Umilia E, Yusuf M, Pratomoatmojo N A. Carrying capacity and environmental capacity analysis based on ecosystem services in Surabaya. IOP Conference Series: Earth and Environmental Science, 2020, 562: 012027. DOI:10.1088/1755-1315/562/1/012027 |
| [6] |
Liu Y G, Zhang J H, Wang S X, Wang Y, Zhao A L. Assessment of environmental carrying capacity using principal component analysis. Journal of Geoscience and Environment Protection, 2018, 6(3): 54-65. DOI:10.4236/gep.2018.63006 |
| [7] |
Goss-Custard J D, West A D. The concept of carrying capacity and shorebirds//Goss-Custard J D, Rufino R, Luis A, eds. Effect of Habitat Loss and Change on Waterbirds. London: The Stationery Office, 1997: 52-62.
|
| [8] |
Goss-Custard J D, Stillman R A, Caldow R W G, West A D, Guillemain M. Carrying capacity in overwintering birds: when are spatial models needed?. Journal of Applied Ecology, 2003, 40(1): 176-187. DOI:10.1046/j.1365-2664.2003.00785.x |
| [9] |
Alonso J C, Alonso J A, Bautista L M. Carrying capacity of staging areas and facultative migration extension in common cranes. Journal of Applied Ecology, 1994, 31(2): 212-222. DOI:10.2307/2404537 |
| [10] |
Hobbs N T, Baker D L, Ellis J E, Swift D M, Green R A. Energy- and nitrogen-based estimates of elk winter-range carrying capacity. The Journal of Wildlife Management, 1982, 46(1): 12-21. DOI:10.2307/3808403 |
| [11] |
张明海, 刘群秀. 黑龙江省完达山东部林区马鹿冬季环境容纳量估算. 兽类学报, 2008, 28(1): 56-64. DOI:10.3969/j.issn.1000-1050.2008.01.010 |
| [12] |
官天培, 谌利民, 符建荣, 宋延龄. 唐家河国家级自然保护区扭角羚环境容纳量. 四川林业科技, 2015, 36(3): 69-74. DOI:10.3969/j.issn.1003-5508.2015.03.014 |
| [13] |
Pearse A T, Krapu G L, Brandt D A, Kinzel P J. Changes in agriculture and abundance of snow geese affect carrying capacity of Sandhill cranes in Nebraska. Journal of Wildlife Management, 2010, 74(3): 479-488. DOI:10.2193/2008-539 |
| [14] |
董科, 吕士成, Healy T. 江苏盐城国家级珍禽自然保护区丹顶鹤的承载力. 生态学报, 2005, 25(10): 2608-2615. DOI:10.3321/j.issn:1000-0933.2005.10.022 |
| [15] |
Fonseca J, Basso E, Serrano D, Navedo J G. Effects of tidal cycles on shorebird distribution and foraging behaviour in a coastal tropical wetland: insights for carrying capacity assessment. Estuarine, Coastal and Shelf Science, 2017, 198: 279-287. DOI:10.1016/j.ecss.2017.09.016 |
| [16] |
BirdLife International. Species factsheet: Leucogeranus leucogeranus. [2021-06-15]. http://www.birdlife.org.
|
| [17] |
Harris J, Mirande C. A global overview of cranes: status, threats and conservation priorities. Chinese Birds, 2013, 4(3): 189-209. DOI:10.5122/cbirds.2013.0025 |
| [18] |
Wang W J, Fraser J D, Chen J K. Wintering waterbirds in the middle and lower Yangtze River floodplain: changes in abundance and distribution. Bird Conservation International, 2017, 27(2): 167-186. DOI:10.1017/S0959270915000398 |
| [19] |
Johnsgard P A. Cranes of the world: siberian crane (Bugeranus leucogeranus)//Johnsgard P A, ed. Cranes of the World. Bloomington: Indiana University Press, 1983: 129-139.
|
| [20] |
Hou J J, Li L, Wang Y F, Wang W J, Zhan H Y, Dai N H, Lu P. Influences of submerged plant collapse on diet composition, breadth, and overlap among four crane species at Poyang Lake, China. Frontiers in Zoology, 2021, 18(1): 24. DOI:10.1186/s12983-021-00411-2 |
| [21] |
Li Y K, Zhong Y F, Shao R Q, Yan C, Jin J F, Shan J H, Li F S, Ji W T, Bin L, Zhang X Y, Cao K Q, Shen J. Modified hydrological regime from the three gorges dam increases the risk of food shortages for wintering waterbirds in Poyang Lake. Global Ecology and Conservation, 2020, 24: e01286. DOI:10.1016/j.gecco.2020.e01286 |
| [22] |
Burnham J, Barzen J, Pidgeon A M, Sun B T, Wu J D, Liu G H, Jiang H X. Novel foraging by wintering Siberian cranes leucogeranus leucogeranus at China's Poyang Lake indicates broader changes in the ecosystem and raises new challenges for a critically endangered species. Bird Conservation International, 2017, 27(2): 204-223. DOI:10.1017/S0959270916000150 |
| [23] |
胡振鹏, 林玉茹. 鄱阳湖水生植被30年演变及其驱动因素分析. 长江流域资源与环境, 2019, 28(8): 1947-1955. |
| [24] |
简敏菲, 简美锋, 李玲玉, 汪斯琛, 余厚平, 余冠军. 鄱阳湖典型湿地沉水植物的分布格局及其水环境影响因子. 长江流域资源与环境, 2015, 24(5): 765-772. |
| [25] |
Percival S M, Evans P R. Brent geese Branta bernicla and Zostera; factors affecting the exploitation of a seasonally declining food resource. Ibis, 1997, 139(1): 121-128. |
| [26] |
Fox A D, Cao L, Zhang Y, Barter M, Zhao M J, Meng F J, Wang S L. Declines in the tuber-feeding waterbird guild at Shengjin Lake National Nature Reserve, China - a barometer of submerged macrophyte collapse. Aquatic Conservation: Marine and Freshwater Ecosystems, 2011, 21(1): 82-91. DOI:10.1002/aqc.1154 |
| [27] |
Jia Y F, Jiao S W, Zhang Y M, Zhou Y, Lei G C, Liu G H. Diet shift and its impact on foraging behavior of Siberian crane (Grus leucogeranus) in Poyang Lake. PLoS One, 2013, 8(6): e65843. DOI:10.1371/journal.pone.0065843 |
| [28] |
Hou J J, Liu Y F, Fraser J D, Li L, Zhao B, Lan Z C, Jin J F, Liu G H, Dai N H, Wang W J. Drivers of a habitat shift by critically endangered Siberian cranes: evidence from long-term data. Ecology and Evolution, 2020, 10(20): 11055-11068. DOI:10.1002/ece3.6720 |
| [29] |
王文娟, 王榄华, 侯谨谨. 人工生境已成为鄱阳湖越冬白鹤的重要觅食地. 野生动物学报, 2019, 40(1): 133-137. DOI:10.3969/j.issn.1000-0127.2019.01.020 |
| [30] |
于超, 周立志, 宋昀微. 人为干扰对越冬后期稻田中小天鹅(Cygnus columbianus)昼间能量消耗的影响. 湖泊科学, 2019, 31(1): 195-201. |
| [31] |
钟毅峰, 李言阔. 鄱阳湖越冬水鸟对稻田生境的利用. 中国鹤类通讯, 2018, 22(2): 2-4. |
| [32] |
蒋剑虹, 戴年华, 邵明勤, 黄志强, 卢萍. 鄱阳湖区稻田生境中灰鹤越冬行为的时间分配与觅食行为. 生态学报, 2015, 35(2): 270-279. |
| [33] |
侯谨谨, 王亚芳, 金斌松, 王榄华, 王文娟. 鄱阳湖越冬白鹤在农业用地的食物组成. 动物学杂志, 2019, 54(1): 15-21. |
| [34] |
Feng L, Hu C M, Chen X L, Cai X B, Tian L Q, Gan W X. Assessment of inundation changes of Poyang Lake using MODIS observations between 2000 and 2010. Remote Sensing of Environment, 2012, 121: 80-92. DOI:10.1016/j.rse.2012.01.014 |
| [35] |
Li Y K, Qian F W, Silbernagel J, Larson H. Community structure, abundance variation and population trends of waterbirds in relation to water level fluctuation in Poyang Lake. Journal of Great Lakes Research, 2019, 45(5): 976-985. DOI:10.1016/j.jglr.2019.08.002 |
| [36] |
Barzen J A. Studying and understanding wetland dynamics//Li F, Liu G, Wu J, Zeng N, Harris J, Jin J, eds. Ecological Study of Wetlands and Waterbirds at Poyang Lake. Beijing: Popular Science Press, 2012: 256-266.
|
| [37] |
胡秋红, 谢玮. 食品营养与卫生(第二版). 北京: 北京理工大学出版社, 2017: 25-27.
|
| [38] |
Nilsson L, Persson J, Bunnefeld N, Månsson J. Central place foraging in a human‐dominated landscape: how do common cranes select feeding sites?. Journal of Avian Biology, 2020, 51(6): e02487. DOI:10.1111/jav.02487 |
| [39] |
Zhang Y, Fox A D, Cao L, Jia Q, Lu C H, Prins H H T, de Boer W F. Effects of ecological and anthropogenic factors on waterbird abundance at a ramsar site in the Yangtze River floodplain. Ambio, 2019, 48(3): 293-303. DOI:10.1007/s13280-018-1076-1 |
| [40] |
钟毅峰. 鄱阳湖水稻田鸟类多样性及其生境选择研究[D]. 南昌: 江西师范大学, 2020.
|
| [41] |
何文韵, 邵明勤, 植毅进, 戴年华, 卢萍, 涂小云. 鄱阳湖三个垦殖场的水鸟多样性. 生态学杂志, 2019, 38(9): 2765-2771. |
| [42] |
李言阔, 钟毅峰, 邵瑞清, 魏振华, 张娜, 周鸭仙. 鄱阳湖水稻田越冬水鸟多样性及越冬行为调查简报. 中国鹤类通讯, 2017, 21(2): 14-17. |
| [43] |
Miller M R, Eadie J M. The allometric relationship between resting metabolic rate and body mass in wild waterfowl (Anatidae) and an application to estimation of winter habitat requirements. The Condor, 2006, 108(1): 166-177. DOI:10.1093/condor/108.1.166 |
| [44] |
王岐山, 马鸣, 高育仁. 中国动物志: 鸟纲(第五卷) 鹤形目鸻形目鸥形目. 北京: 科学出版社, 1979: 28-41.
|
| [45] |
郑作新, 张荫荪, 冼耀华, 卢汰春, 谭耀匡, 龙泽虞, 王子玉, 唐蟾珠, 周福璋, 刘长江, 刘如荀, 贝天祥, 丁文宁, 张坦心. 中国动物志: 鸟纲(第二卷) 雁形目. 北京: 科学出版社, 2006: 26-47.
|
| [46] |
Norberg U M. Energetics of flight//Carey C, ed. Avian Energetics and Nutritional Ecology. Boston: Springer, 1996: 199-249.
|
| [47] |
李秀明. 白鹤(Grus leucogranus)迁徙路线及中途停歇地的卫星跟踪研究[D]. 北京: 中国林业科学研究院, 2016.
|
| [48] |
黄田, 徐正刚, 彭姣, 赵运林. 基于卫星跟踪的洞庭湖越冬小天鹅迁徙路线研究. 四川动物, 2018, 37(4): 361-372. |
| [49] |
Skyllberg U, Nousiainen I, Hansson P, Bernhardtson P, Andersson Ö, Nordlund M. Spring migration of the Taiga Bean Goose Anser f. fabalis along the "Western Flyway" in northern Sweden: numbers in 2003-2008 and timing in comparison with the "Central Flyway" in Finland. Ornis Svecica, 2009, 19(4): 199-214. |
| [50] |
Batbayar N, Takekawa J Y, Newman S H, Prosser D J, Natsagdorj T, Xiao X M. Migration strategies of Swan Geese Anser cygnoides from northeast Mongolia. Wildfowl, 2011, 61: 90-109. |
| [51] |
Li X H, Wang X, Fang L, Batbayar N, Natsagdorj T, Davaasuren B, Damba I, Xu Z G, Cao L, Fox A D. Annual migratory patterns of Far East Greylag Geese (Anser anser rubrirostris) revealed by GPS tracking. Integrative Zoology, 2020, 15(3): 213-223. DOI:10.1111/1749-4877.12414 |
| [52] |
吴建东, 李凤山, Burnham J. 鄱阳湖沙湖越冬白鹤的数量分布及其与食物和水深的关系. 湿地科学, 2013, 11(3): 305-312. DOI:10.3969/j.issn.1672-5948.2013.03.001 |
| [53] |
Wood C, Qiao Y, Li P, Ding P, Lu B Z, Xi Y M. Implications of rice agriculture for wild birds in China. Waterbirds, 2010, 33(sp1): 30-43. DOI:10.1675/063.033.s103 |
| [54] |
Czech H A, Parsons K C. Agricultural wetlands and waterbirds: a review. Waterbirds: The International Journal of Waterbird Biology, 2002, 25(sp2): 56-65. |
| [55] |
Fox A D, Abraham K F. Why geese benefit from the transition from natural vegetation to agriculture. Ambio, 2017, 46(S2): 188-197. |
| [56] |
Hill S C, Lee Y J, Song B M, Kang H M, Lee E K, Hanna A, Gilbert M, Brown I H, Pybus O G. Wild waterfowl migration and domestic duck density shape the epidemiology of highly pathogenic H5N8 influenza in the Republic of Korea. Infection, Genetics and Evolution, 2015, 34: 267-277. |
| [57] |
Zhao Q S, Wang X, Cao L, Fox A D. Why Chinese wintering geese hesitate to exploit farmland. Ibis, 2018, 160(3): 703-705. DOI:10.1111/ibi.12605 |
| [58] |
Yu H, Wang X, Cao L, Zhang L, Jia Q, Lee H, Xu Z G, Liu G H, Xu W B, Hu B H, Fox A D. Are declining populations of wild geese in China 'prisoners' of their natural habitats?. Current Biology, 2017, 27(10): R376-R377. DOI:10.1016/j.cub.2017.04.037 |
| [59] |
邵明勤, 龚浩林, 戴年华, 植毅进, 徐宁, 卢萍. 鄱阳湖围垦区藕塘越冬白鹤的时间分配与行为节律. 生态学报, 2018, 38(14): 5206-5212. |
| [60] |
Nolet B A, Langevoord O, Bevan R M, Engelaar K R, Klaassen M, Mulder R J W, Van Dijk S. Spatial variation in tuber depletion by swans explained by differences in net intake rates. Ecology, 2001, 82(6): 1655-1667. |
| [61] |
Sponberg A F, Lodge D M. Seasonal belowground herbivory and a density refuge from waterfowl herbivory for Vallisneria americana. Ecology, 2005, 86(8): 2127-2134. DOI:10.1890/04-1335 |