 2022, Vol. 42
2022, Vol. 42文章信息
- 熊千志, 杜恩在, 薛峰, 李文卿, 周子建, 赵守栋, 江源
- XIONG Qianzhi, DU Enzai, XUE Feng, LI Wenqing, ZHOU Zijian, ZHAO Shoudong, JIANG Yuan
- 塞罕坝地区人工针叶林径向生长对水热条件的响应
- Response of radial growth of artificial coniferous forest to hydrothermal conditions in Saihanba Area
- 生态学报. 2022, 42(13): 5371-5380
- Acta Ecologica Sinica. 2022, 42(13): 5371-5380
- http://dx.doi.org/10.5846/stxb202107071814
-
文章历史
- 收稿日期: 2021-07-07
- 网络出版日期: 2022-03-18





2. 北京师范大学地理科学学部, 北京 100875
2. Faculty of Geographical Science, Beijing Normal University, Beijing 100875, China
水热条件是影响树木生长的重要生态因子。早在1941年, Glock就系统地阐述了树木年轮与气温和降水之间的关系, 并提出了气候影响树木生长的机理[1]。树木年轮宽度数据具有定年精确、连续性好、分辨率高、易获取等优点[2], 并能够通过其所反映的树木生长特征来推断生长与气候关系[3], 因此不仅成为重建古气候的重要代用指标, 而且也为研究树木生长与水热因子的相互关系等提供了有效的科学手段[4-7]。近百年来全球变暖明显, 树木年轮数据在研究树木生长对气候变化的响应方面发挥了重要作用[8]。
塞罕坝地区历史上曾经是皇家狩猎场所, 分布着一定面积的森林植被[9], 但由于历史上的人为破坏和气候变化, 森林植被几乎消失殆尽。自20世纪60年代以来该地区开始营造以华北落叶松(Larix principis-rupprechtii)和樟子松(Pinus sylvestris var. mongolica)为优势种的人工针叶林, 经过多年坚持不懈的努力, 现已形成亚洲面积最大的人工林, 发挥着阻挡北部浑善达克沙地南侵、调节当地气候、涵养水源、保障农业生产等重要生态功能。然而, 塞罕坝地区地处农牧交错带, 寒冷干旱的大陆性季风气候使树木的生长受到较多限制。之前对塞罕坝围场地区森林与气候关系的研究表明, 油松(Pinus tabuliformis Carr)径向生长对当地5、6月的平均气温的负响应显著[10];塞罕坝围场木兰林区华北落叶松的生长也受区域干旱限制[11];此外, 有关毛乌素沙地地区和塞罕坝地区樟子松径向生长的对比研究表明, 塞罕坝地区7、8月干旱和春季的快速升温是限制当地樟子松生长的重要因素[12]。综上所述, 之前的研究表明降水不足以及因春季、夏季高温所导致的干旱对塞罕坝地区林木生长产生了不利影响。
然而, 纬度较高的塞罕坝地区存在季节性冻土[13], 而冻土地区的冻融过程以及与之相联系的土壤水势变化, 对树木生长有着特殊影响[14-17], 故长期以来也被认为是影响人工林建植的一个不利因素[18]。之前的相关研究集中关注了生长—气候关系, 其结果显示气温和降水导致的水亏缺是树木生长的不利因素。有关季节性冻土地区的地温条件是否对树木生长产生影响这一问题, 尚缺少深入探讨。此外, 塞罕坝地区人工林中的优势种华北落叶松和樟子松分属两类不同生活型, 二者对上述影响本区域人工林建设的不利因素是否具有相似的响应特点, 是关系到本地区人工林是否能够长期稳定的重要问题。
针对以上科学问题, 本文拟采用树木年轮生态学方法开展研究, 以期通过分析轮宽指数与气温、地表温度、降水量和饱和水汽压差等气候要素的相关关系, 揭示两个树种的径向生长—水热因子关系特征, 进而探讨两树种在气候变化背景下对塞罕坝地区生态条件的适应能力, 为建设更加稳定的人工林植被提供基础科学依据。
1 研究区域及研究方法 1.1 研究区概况塞罕坝林场地处内蒙古高原和河北北部间山盆地的交汇区域(图 1), 海拔1010—1939.6 m, 总面积约930 km2。高原地形使位于中纬度的该地区在气候上冬季漫长寒冷, 夏季短暂凉爽。研究区年平均气温为3.1℃, 极端最高气温29.4℃, 极端最低气温-25.6℃;年平均地表温度5.8℃;年均降水量仅378.2 mm, 其中6—9月的降水量占全年的77.5%(图 2);全年平均相对湿度59.6%, 夏季相对湿度可超70%。自1962年建场以来, 塞罕坝地区开始了大面积人工造林, 造林的主要针叶树种为华北落叶松和樟子松。1962年, 林场拥有130 km2的天然次生林, 2017年时, 林场的林地面积已达740 km2[19], 森林覆盖率由14%增长到80%。

|
| 图 1 研究区概况和采样点、气象站位置 Fig. 1 General situation of study area and location of tree-ring sampling sites and meteorology stations |

|
| 图 2 多伦气象站1988—2016年气温、地温、降水量和VPD年际变化趋势与气温和降水年内变化趋势 Fig. 2 Annual trends of air temperature, surface temperature, precipitation and vapor pressure deficit and monthly variation trend of temperature and precipitation from 1988 to 2016 in Duolun County Meteorology Station ATmax:平均最高气温;ATmean:平均气温;ATmin:平均最低气温;STmax:平均最高地温;STmean:平均地温;STmin:平均最低地温;PR:降水量;VPD:饱和水气压差 |
2016年8月, 在塞罕坝木兰围场生境相似的两个人工林样地, 分别采集了华北落叶松和樟子松树轮样本。采样时, 选择生长良好的树木, 在树木胸径高处分别在顺等高线方向和垂直坡面方向分别钻取树木样芯。每棵树采集四个样芯, 每个树种采样25棵树共100个可用于分析的样芯, 之后对采样过程中损坏的样芯予以剔除。采样信息如表 1。
| 树种 Tree specie |
纬度 Latitude /N |
经度 Longitude /E |
样本量(树/芯) Sample size(tree/core) |
海拔 Altitude/m |
| 华北落叶松Larix principis-rupprechtii | 42 °24′33 ″ | 117 °19′9 ″ | 25/100 | 1590 |
| 樟子松Pinus sylvestris var. mongolica | 42 °23′35 ″ | 117 °14′59 ″ | 25/98 | 1480 |
样品带回实验室后依照Stokes等的方法进行样品的预处理[20]。将样品晾干、固定后进行打磨, 并在放大镜下进行交叉定年。使用Lintab年轮测量仪在0.01 mm的精度下测量轮宽, 之后使用COFECHA程序对测得的序列进行校正[21]。处理后的年轮序列使用ARSTAN程序建立年表, 并采用样条函数法进行拟合, 去除树木生长对树轮轮宽的影响。最终建立塞罕坝华北落叶松和樟子松的标准化年表(STD)、差值年表(RES)和自回归年表(ARS)。由于标准年表包含更多的低频信息, 且剔除了非气候信号和生长趋势[22], 因此本文采用标准年表进行后续分析, 以揭示两个树种的生长-水热因子关系, 以及对气候变化的响应。
1.3 气象数据收集距离采样点较近的气象站有围场站、赤峰站和多伦站, 鉴于围场和赤峰气象站曾分别于2009、1993和2011年有过迁站历史[23, 24], 本研究采用多伦气象站的数据进行分析。从中国气象科学数据共享服务网(http://cdc.cma.gov.cn)下载多伦站1988—2016年的月平均气温、月平均最高气温、月平均最低气温、月降水量、月平均相对湿度、日平均0 cm地温、日最高0 cm地温和日最低0 cm地温的连续气象数据, 通过进一步计算得到月平均0 cm地温(简称月平均地温)、月平均最高0 cm地温(简称月平均最高地温)、月平均最低0 cm地温(简称月平均最低地温)和月平均饱和水气压差(VPD)。
VPD计算公式如下:
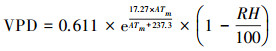
|
式中, VPD为饱和水气压差;ATm为月平均气温;RH为月平均相对湿度。
1.4 数据分析采用皮尔逊相关分析径向生长对水热因子的响应关系。由于树木的径向生长不仅受当年, 而且受前一年水热因子的影响[25], 本研究选择上年6月到当年9月的月降水量、月均饱和水气压差、月平均气温、月平均最高、最低气温和月平均地温、月平均最高和最低地温, 与两树种的标准年表数据进行相关分析。在以上分析的基础上, 进而采用滑动相关分析评估径向生长与水热因子间关系的时间变化特点[26]。生长—水热因子关系呈现正相关, 表明该因子数值高时对生长会产生促进作用, 呈现负相关则表明该因子数值增高时有可能对生长产生不利影响[27]。
为了避免树木幼年生长数据的影响, 采用了生长相对稳定时期的年表数据进行分析, 年表数据的公共时段区间为1988—2016年。
2 结果与分析 2.1 气候变化特征根据位于42.25° N, 116.22° E, 海拔1211 m处的多伦气象站数据, 本地区的水热条件在过去29年发生了一定程度的变化(图 2)。从1988—2016年, 年平均气温、年平均最高和最低气温分别以0.2℃/10a、0.21℃/10a和0.07℃/10a的速率上升, 0cm深度的平均地温、平均最低和最高地温分别以0.89℃/10a、1.88℃/10a的速率上升(P≤0.01)和0.09℃/10a的速率下降;年平均饱和水气压差(VPD)以0.27hpa/10a的速率显著上升(P≤0.01), 年降水量变化趋势不显著。在本文获得的树木年轮数据所显示的共同时间段内(1988—2016年), 仅0cm深度的年平均和年平均最低地温和空气VPD分别表现出以0.89℃/10a、1.88℃/10a和0.27 hpa/10a的速率升高, 其余参数未表现出显著变化。
对月数据的多年变化进行分析的结果(图 3)表明, 从1988—2016年, 3、5、7和8月VPD显著上升(P≤0.05), 其中8月VPD极显著上升(P≤0.01);5月平均气温和6月平均最低气温显著上升(P≤0.05);1、3和8—12月的平均地温显著上升(P≤0.05), 其中1、9、11和12月的平均地温极显著上升(P≤0.01);所有月份平均最低地温都呈显著上升趋势(P≤0.05), 除7和8月外, 都表现出极显著上升趋势(P≤0.01)。月数据的多年变化趋势与年数据的变化趋势一致。总体来看, 本区域的气候表现出向暖干化发展的特点。

|
| 图 3 多伦气象站月气候因子年际变化趋势 Fig. 3 Annual trends of monthly climatic factors in Duolun County Meteorology Station |
华北落叶松和樟子松的树轮宽度年表参数如表 2所示。其中, 华北落叶松年表的平均敏感度(MS)为0.32, 樟子松平均敏感度(MS)为0.195, 这表明华北落叶松的年表具有相对更强的高频信号。两个树种年表的一阶自相关系数(AC)较低, 表明其受前一年生长的影响均相对较小[22]。两个年表所有样芯的平均相关系数(Rtot)数值较高, 表明不同样芯间的轮宽变化一致性较强。另外, 两年表的第一特征向量百分比(PCI)都较高, 表明年表包含了较多的环境信息。其中, 华北落叶松的信噪比(SNR)较高, 而樟子松信噪比(SNR)相对偏低。总的来说, 两个树种的样本总体代表性(EPS)均超过了90%。
| 标准年表 Standard chronology |
华北落叶松 Larix principis-rupprechtii |
樟子松 Pinus sylvestris var. mongolica |
|
| 样本量(树/芯)Sample number | 25/100 | 25/98 | |
| 序列长度Period | 1967—2016 | 1978—2016 | |
| 平均敏感度Mean sensitivity (MS) | 0.32 | 0.195 | |
| 一阶自相关系数First-order auto-correlation (AC) | 0.227 | 0.19 | |
| 公共时段区间Common intervals | 1988—2016 | 1988—2016 | |
| 公共区间统计量 Common interval parameters |
所有样芯平均相关系数 Mean inter-series correlations (Rtot) |
0.678 | 0.665 |
| 树间平均相关系数 Mean among-trees correlations (Rbt) |
0.672 | 0.526 | |
| 树内平均相关系数 Mean within-trees correlations (Rwt) |
0.871 | 0.823 | |
| 第一特征向量百分比 Variance in first eigenvector (PC1) |
0.688 | 0.726 | |
| 标准差 Standard deviation (SD) |
0.123 | 0.164 | |
| 信噪比 Signal-to-noise ratio (SNR) |
175.111 | 11.913 | |
| 样本总体代表性 Expressed population signal (EPS) |
0.994 | 0.923 | |
华北落叶松标准年表与各气候因子的相关分析显示(图 4):(1)气温方面, 华北落叶松的径向生长与当年6、7月的平均气温和平均最高气温, 以及上年6、9月的平均最高气温均呈显著负相关。(2)地温方面, 其径向生长与上年6、9月, 当年6、7月的平均地温和平均最高地温呈显著负相关;而与当年1月的平均地温和平均最低地温呈显著正相关。(3)降水方面, 华北落叶松的径向生长与上年6月、当年2、6、7月的降水均呈显著正相关, 而与8月降水呈显著负相关。(4)VPD方面, 径向生长与上年6、9月以及当年6、7月的VPD均呈显著负相关。

|
| 图 4 两树种标准年表与月气候因子相关分析结果 Fig. 4 Correlation coefficient between chronologies and monthly climatic factors |
樟子松标准年表与各气候因子的相关分析(图 4)表明:(1)气温方面, 其径向生长与当年6月的平均气温, 当年7月的平均最高气温均呈显著负相关;与当年8月的平均气温呈显著正相关。(2)地温方面, 径向生长与当年7月的平均最高地温呈显著负相关;与当年8月的平均地温和上年12月、当年5、7、8月的平均最低地温呈显著正相关。(3)降水方面, 与上年和当年8月降水均呈显著负相关;与当年7月降水呈显著正相关。(4)VPD方面, 与当年7月的VPD呈显著负相关。
2.4 径向生长受水热因子影响的时间变化特征在上述相关分析的基础上, 我们对其中的显著相关月份中的生长-水热因子关系进行滑动相关分析, 以揭示其关系随时间的变化特征。参考前人研究[10], 选择11年进行滑动相关, 对两树种进行对比分析。
华北落叶松的滑动相关分析结果(图 5)显示:除平均最低地温和平均最低气温之外, 径向生长与温度因子和VPD的关系在整个时期内以负相关为主, 且以当年7月的负相关最为典型, 与7月平均最低地温的关系也表现出负相关为主的特点;径向生长与温度因子正相关为主的关系, 表现在与9月平均最低气温、1月平均地温、当年1月、5月和8月的平均最低地温;与降水的关系, 除8月份呈现负相关之外, 总体表现为正相关关系。

|
| 图 5 华北落叶松与各气候因子显著相关月份的滑动相关系数 Fig. 5 Moving correlation coefficients between significantly correlated monthly climatic factors and chronologies of Larix principis-rupprechtii c:前一年;黑色圆圈表示显著相关P < 0.05 |
樟子松的滑动相关分析结果(图 6)显示:除平均最低地温之外, 径向生长与大部分温度因子和VPD主要在当年6月和7月呈负相关, 与最低平均气温在当年9月呈负相关;与温度因子呈正相关为主的关系, 表现在与8月的平均气温和平均地温, 以及几乎全部参与分析月份的平均最低地温;与降水的关系呈现出在2月、6月和7月以正相关为主, 8月则以负相关为主。

|
| 图 6 樟子松与各气候因子显著相关月份的滑动相关系数 Fig. 6 Moving correlation coefficients between significantly correlated monthly climatic factors and chronologies of Pinus sylvestris var. mongolica c:前一年;黑色圆圈表示显著相关P < 0.05 |
树木生长与水热因子的关系, 既与水热因子时空差异有关, 也与树木本身的生理生态特性相关[28]。本文所研究的华北落叶松和樟子松分别属于落叶针叶树和常绿针叶树两类不同生活型的树种。来自于树木生理方面的研究结果表明, 落叶松较之于常绿针叶树而言, 不仅每年的光合作用产物形成和分配受长叶和落叶影响, 而且其在水分平衡方面同时具有蒸腾丢失水分和根系吸收水分能力较强的特征[29]。
3.1 两树种径向生长对水热条件响应的相似性两树种径向生长对气候因子的响应具有一定的相似性, 表现在两树种径向生长都受到夏季6、7月高温干旱的限制、8月过多降水的限制(图 4)。夏季6、7月份是华北地区树木径向生长最旺盛的时期[30, 31], 夏季的高温会导致植物过强的蒸腾, 气孔关闭, 植物光合作用减弱, 从而形成窄轮[5, 32, 33]。而树木径向生长与8月降水的负相关, 可能是因当地8月的充盈降水(图 1)导致辐射量不足, 树木的营养积累减少, 形成窄轮, 甚至影响到下一年的生长[34]。这种由于高温导致的干旱影响也可以通过两个树种径向生长与5、6、7月降水量正相关, 以及与6、7月VPD的负相关得以佐证。与气候条件的这种响应关系, 在整个研究时段上表现得相对稳定。
地温方面, 在生长旺盛时期的6、7月, 平均地温和最高平均地温的影响与气温的影响相似, 呈现与径向生长的负相关, 即较高的温度会通过引起水分土壤水分亏缺而限制径向生长。上年12月、当年1月和5月份平均地温和平均最低地温与两树种的径向生长更多地表现为正相关, 这很可能与研究区存在季节性冻土有关。塞罕坝地处中纬度稍偏北位置, 加之区内整体海拔较高, 是华北地区季节性冻土的分布地区[13]。冬季较低的地温将有可能导致冻土对树木根部的伤害[35]。而5月份作为树木生长过程中形成层开始活动的重要时期[36], 快速上升的气温加速了树木的蒸腾作用, 而此时较低的地温却限制了树木根系的水分吸收过程, 导致树木经受生理干旱, 对径向生长产生不利影响。
3.2 两树种径向生长对水热条件响应的差异气温方面, 华北落叶松对9月最低平均气温表现出正相关特征, 樟子松表现出负相关特征;两个树种对8月份的平均气温的响应却表现出与此相反的特点;地温方面, 较为突出的差异表现在8月的平均地温与落叶松径向生长表现为负相关, 而与樟子松呈现出稳定的正相关。
两个树种径向生长在秋季表现出来的对水热条件响应的差异, 显现出二者在生理生态方面的差异。水分生理生态方面的研究表明[37], 落叶松的蒸腾失水不仅对大气VPD变化较为敏感, 而且由此形成的蒸腾拉力导致的根系吸水力也较强。相对而言, 樟子松的针叶具有较厚的角质层且气孔深陷, 其水分平衡趋向于采取保守的策略, 蒸腾对VPD的敏感性不高, 因此与蒸腾拉力相关联的吸水力也相对偏弱。两个树种在水分利用方面的这种差异, 导致了落叶松对秋季(8月)平均地温的关系具有负相关特点, 即如果土温偏高, 蒸散量大, 土壤水分含量降低, 会导致生长受到抑制。对于樟子松而言, 与蒸腾拉力相联系的吸水力偏小, 如果地温在秋季降低幅度较大, 则容易对该树种产生生理干旱, 进而抑制其生长;而此时如果气温偏高不仅会因蒸腾较强而使水分亏缺程度增强, 而且会因较高的呼吸速率而消耗更多的光合产物, 从而使径向生长受到更强抑制。从对气温的响应差异看, 代表秋季的夜间温度的最低平均地温不利于华北落叶松的径向生长, 其原因很可能是因为低温加速落叶松的落叶使生长提前结束。
3.3 区域增温对两树种径向生长的可能影响本研究分析了1987以来年内各月份水热参数的时间的变化趋势, 通过分析一方面可以看出, 生长季前期(3—5月)和生长季后期(7—8月)VPD呈现增强的趋势, 秋冬季节(前一年11—12月, 当年1—3月)的平均地温呈现升高趋势, 以及平均最低地温普遍出现增高趋势(图 3)。另一方面本文的数据分析结果表明, 樟子松的径向生长与平均最低地温基本呈现正相关关系, 与平均地温在8月份呈现正相关关系, 在当年1月和前一年的夏秋季节的一些时期也呈现出正相关特征。因此, 从多年变化最普遍的平均地温和平均最低地温的增温趋势看, 如果未来保持这种特点不变, 樟子松的生长可能获得较大的促进, 相对于华北落叶松其竞争力会相对增强。由此, 也可推断, 常绿的樟子松是未来一段时期内更适合于在塞罕坝地区进行植树造林的地方性树种。
4 结论(1) 塞罕坝林场华北落叶松树轮年表和樟子松树轮年表的样本总体代表性(EPS)均超过0.9, 是研究树木生长对气候要素响应的可靠材料。
(2) 塞罕坝地区人工林径向生长均表现出主要受到当地夏季的高温干旱胁迫和深秋至冬季偏低地温的不利影响。由于两树种对环境适应特征的差异, 相比较而言, 华北落叶松受干旱胁迫强于樟子松, 而樟子松受较低地温的胁迫强于华北落叶松。
(3) 在塞罕坝区域增温的背景下, 樟子松的稳定性可能较高, 与华北落叶松相比或许是更适合于作为塞罕坝人工林营造的树种。
| [1] |
Glock W S. Growth rings and climate. The Botanical Review, 1941, 7(12): 649. DOI:10.1007/BF02872439 |
| [2] |
邵雪梅. 树轮年代学的若干进展. 第四纪研究, 1997, 17(3): 265-271. DOI:10.3321/j.issn:1001-7410.1997.03.009 |
| [3] |
Zhang Q B, Hebda R J. Variation in radial growth patterns of Pseudotsuga menziesii on the central coast of British Columbia, Canada. Canadian Journal of Forest Research, 2004, 34(9): 1946-1954. DOI:10.1139/x04-078 |
| [4] |
王振威, 李强, 刘禹, 杨银科, 任萌, 崔琳琳. 树轮记录的吕梁山北部过去175年来帕尔默干旱指数变化. 地球环境学报, 2020, 11(1): 72-80. |
| [5] |
姜盛夏, 袁玉江, 喻树龙, 尚华明, 张瑞波, 秦莉, 张合理, 张同文. 阿尔泰山西伯利亚云杉多种树轮宽度年表研制及其气候响应研究. 地球环境学报, 2020, 11(6): 616-628. |
| [6] |
李淑娟, 喻树龙, 尚华明, 袁玉江, 姜盛夏, 刘艳, 范煜婷, 牛军强. 新疆西天山不同海拔雪岭云杉树轮宽度年表特征及其气候响应分析. 沙漠与绿洲气象, 2017, 11(1): 50-57. |
| [7] |
Aragão J R V, Groenendijk P, Lisi C S. Dendrochronological potential of four neotropical dry-forest tree species: climate-growth correlations in northeast Brazil. Dendrochronologia, 2019, 53: 5-16. DOI:10.1016/j.dendro.2018.10.011 |
| [8] |
Depardieu C, Girardin M P, Nadeau S, Lenz P, Bousquet J, Isabel N. Adaptive genetic variation to drought in a widely distributed conifer suggests a potential for increasing forest resilience in a drying climate. New Phytologist, 2020, 227(2): 427-439. DOI:10.1111/nph.16551 |
| [9] |
姚伟强, 李巍, 刘亚春. 英雄创业越千秋茫茫荒原变绿洲——河北省塞罕坝机械林场三代人55年艰苦奋斗造林纪实. 河北林业, 2017(6): 22-27. |
| [10] |
崔明星, 何兴元, 陈玮, 陈振举, 周长虹, 吴涛. 河北木兰围场油松年轮生态学的初步研究. 应用生态学报, 2008, 19(11): 2339-2345. |
| [11] |
郭敬丽, 丁旭光, 巩建新, 刘楠. 木兰林区华北落叶松生长与气候因子的关系. 河北林果研究, 2015, 30(1): 21-25. |
| [12] |
雷帅, 张劲松, 孟平, 何子兵, 程顺, 孙守家. 中国北部不同地点樟子松人工林径向生长对气候响应的差异. 生态学报, 2020, 40(13): 4479-4492. |
| [13] |
杨帅, 侯奇奇, 耿雪莹, 高宇俊, 刘璇, 周鹏. 1974-2016年河北省最大冻土深度及其与温度的关系. 干旱气象, 2020, 38(3): 380-387. |
| [14] |
常永兴, 陈振举, 张先亮, 白学平, 赵学鹏, 李俊霞, 陆旭. 气候变暖下大兴安岭落叶松径向生长对温度的响应. 植物生态学报, 2017, 41(3): 279-289. |
| [15] |
张轩文, 杨丽, 刘晓宏, 张秋良, 王文志, 曾小敏, 吴国菊. 大兴安岭北部多年冻土区落叶松和樟子松生长的气候响应差异研究. 冰川冻土, 2017, 39(1): 165-174. |
| [16] |
Sugimoto A, Yanagisawa N, Naito D, Fujita N, Maximov T C. Importance of permafrost as a source of water for plants in East Siberian taiga. Ecological Research, 2002, 17(4): 493-503. DOI:10.1046/j.1440-1703.2002.00506.x |
| [17] |
杜子银, 蔡延江, 王小丹, 鄢燕, 鲁旭阳, 刘淑珍. 土壤冻融作用对植物生理生态影响研究进展. 中国生态农业学报, 2014, 22(1): 1-9. |
| [18] |
Mellander P E, Bishop K, Lundmark T. The influence of soil temperature on transpiration: a plot scale manipulation in a young Scots pine stand. Forest Ecology and Management, 2004, 195(1/2): 15-28. |
| [19] |
常伟强. 塞罕坝机械林场森林资源动态变化分析. 林业资源管理, 2018(6): 13-17. |
| [20] |
Stokes M A, Smiley T L. An Introduction to Tree-Ring Dating. Tucson: The University of Arizona Press, 1996.
|
| [21] |
Holmes R L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bulletin, 1983, 43: 69-78. |
| [22] |
Fritts H C. Tree Rings and Climate. New York: Academic Press, 1976.
|
| [23] |
马凤莲, 刘园园, 周士茹, 朱环娟, 董学友. 从对比观测资料看城镇化对气象要素的影响——以围场国家基本气象站为例. 干旱气象, 2011, 29(2): 205-210. DOI:10.3969/j.issn.1006-7639.2011.02.013 |
| [24] |
程玉琴, 尤莉, 马小林, 张少文, 訾耀海, 于春艳. 赤峰基准气象站两次迁址对温度的影响及订正. 干旱区资源与环境, 2015, 29(4): 125-130. |
| [25] |
尹训钢, 吴祥定. 华山松树木年轮对气候响应的模拟分析. 应用气象学报, 1995, 6(3): 257-264. |
| [26] |
张文涛, 江源, 王明昌, 张凌楠, 董满宇. 芦芽山阳坡不同海拔华北落叶松径向生长对气候变化的响应. 生态学报, 2015, 35(19): 6481-6488. |
| [27] |
吴祥定, 邵雪梅. 采用树轮宽度资料分析气候变化对树木生长量影响的尝试. 地理学报, 1996, 51(S1): 92-101. DOI:10.11821/xb1996S1010 |
| [28] |
Li W Q, Jiang Y, Dong M Y, Du E Z, Wu F, Zhao S D, Xu H. Species-specific growth-climate responses of Dahurian larch (Larix gmelinii) and Mongolian pine (Pinus sylvestris var. mongolica) in the Greater Khingan Range, Northeast China. Dendrochronologia, 2021, 65: 125803. DOI:10.1016/j.dendro.2020.125803 |
| [29] |
Urban J, Rubtsov A V, Urban A V, Shashkin A V, Benkova V E. Canopy transpiration of a Larix sibirica and Pinus sylvestris forest in Central Siberia. Agricultural and Forest Meteorology, 2019, 271: 64-72. DOI:10.1016/j.agrformet.2019.02.038 |
| [30] |
董满宇, 江源, 王明昌, 张文涛, 杨浩春. 芦芽山林线华北落叶松径向变化季节特征. 生态学报, 2012, 32(23): 7430-7439. |
| [31] |
季倩雯, 郑成洋, 张磊, 曾发旭. 河北塞罕坝樟子松径向生长动态变化及其与气象因子的关系. 植物生态学报, 2020, 44(3): 257-265. |
| [32] |
王婷, 于丹, 李江风, 马克平. 树木年轮宽度与气候变化关系研究进展. 植物生态学报, 2003, 27(1): 23-33. DOI:10.3321/j.issn:1005-264X.2003.01.004 |
| [33] |
宋来萍, 刘礴霏, 王玉华, 高敬泽. 呼伦贝尔沙地不同树龄樟子松对气候的响应. 南京林业大学学报: 自然科学版, 2020, 44(2): 159-164. |
| [34] |
吴祥定. 树木年轮与气候变化. 北京: 气象出版社, 1990.
|
| [35] |
袁玉江, 李江风. 天山乌鲁木齐河源450a冬季温度序列的重建与分析. 冰川冻土, 1999, 21(1): 64-70. |
| [36] |
Ding X Y, Jiang Y, Xue F, Zhang Y P, Wang M C, Kang M Y, Xu H. Intra-annual growth dynamics of Picea meyeri needles, shoots, and stems on Luya Mountain, North-Central China. Trees, 2021, 35(2): 637-648. DOI:10.1007/s00468-020-02065-9 |
| [37] |
Oberhuber W, Kofler W, Schuster R, Wieser G. Environmental effects on stem water deficit in co-occurring conifers exposed to soil dryness. International Journal of Biometeorology, 2015, 59(4): 417-426. DOI:10.1007/s00484-014-0853-1 |