 2022, Vol. 42
2022, Vol. 42文章信息
- 李敏敏, 刘鹏程, 孔维民, 马方莲, 李帅锋, 王萌
- LI Minmin, LIU Pengcheng, KONG Weimin, MA Fanglian, LI Shuaifeng, WANG Meng
- 濒危植物澜沧黄杉种群结构及动态特征
- Population structure and dynamic characteristics of the endangered Pseudotsuga forrestiiCraib
- 生态学报. 2022, 42(13): 5504-5515
- Acta Ecologica Sinica. 2022, 42(13): 5504-5515
- http://dx.doi.org/10.5846/stxb202001100087
-
文章历史
- 收稿日期: 2020-01-10
- 网络出版日期: 2022-03-22





2. 中国林业科学研究院高原林业研究所, 昆明 650224;
3. 上海同济城市规划设计研究院有限公司(城市空间与生态规划研究中心), 上海 200092
2. Institute of Highland Forest Science, Chinese Academy of Forestry, Kunming 650224, China;
3. Shanghai Tongji Urban Planning and Design Institute (Urban Space and Ecological Planning Institute), Shanghai 200092, China
植物生态学研究的核心问题包括了植物种群的结构特点及时空变化趋势[1-2]。种群结构作为种群最基本的特征, 反映了种群内不同个体的数量及分布情况。而种群的数量动态变化不仅可以揭示种群生态学与生境的关系, 还可以预测种群未来发展趋势, 这对于科学合理进行种群的保护和管理(恢复)具有重要的理论指导意义[3-4]。静态生命表和存活曲线是进行种群统计及研究种群数量动态变化的重要工具, 通过编制静态生命表和绘制生存曲线不仅能揭示种群的结构现状[5], 利用其中得出的生存率、死亡率等重要参数结合种群动态量化方法和时间序列分析方法还可以反映种群的受干扰情况, 进一步还可预测种群的未来发展趋势。目前, 这些分析方法被广泛应用于植物种群相关研究中[6-8]。
澜沧黄杉(Pseudotsuga forrestii), 松科黄杉属, 我国特有种, 国家Ⅱ级保护植物[9], 《中国植物红皮书》(第一册)将澜沧黄杉定为“渐危种(V)”[10], 该物种主要分布于云南西北部、西藏东南部及四川南部高山地带区域[11]。黄杉属是裸子植物中“东亚-北美西部间断分布”的典型例证, 澜沧黄杉是黄杉属分布最西, 垂直分布较高的种类[12-13], 对研究植物区系地理和松科分类进化具有较高的学术价值。近年来, 由于人类活动的加剧、生境的恶化, 澜沧黄杉野外种群面临重大威胁。以往有学者研究了澜沧黄杉的植物地理学[12]、资源分布及保护利用[14-15]等, 但针对澜沧黄杉种群结构及种群动态变化还未见专门报道。本文基于对澜沧黄杉种群的野外调查数据, 通过种群静态生命表、时间序列预测以及生存分析, 旨在揭示澜沧黄杉种群的生存现状及动态趋势, 为澜沧黄杉的保护与恢复提供基础资料和科学依据。
1 研究区概况研究区位于云南省迪庆藏族自治州德钦县, 为三江并流地带, 地理位置介于98°42′-99°17′ E, 27°38′-28°36′ N之间, 属于亚热带与温带季风高原山地气候, 年平均气温15.3 ℃, 年平均降雨量938 mm。本次调查的澜沧黄杉种群分布在海拔2300-3300 m的高山区域, 土壤为棕壤, 该区域内的植被以针叶林为主, 与澜沧黄杉伴生的主要乔木树种有丽江云杉Picea likiangensis、油麦吊云杉Picea brachytyla var.complanata、长苞冷杉Abies georgei、川滇冷杉Abies forrestii、云南黄果冷杉Abies ernestii var. salouenensis、云南松Pinus yunnanensis、青榨槭Acer davidii、云南铁杉Tsuga dumosa、华山松Pinus armandi、冠萼花楸Sotbus coronata、川滇高山栎Quercus aquifolioides、槲栎Quercus aliena、滇青冈Cyclobalanopsis glaucoides、山杨Populus davidiana var. davidiana等, 林下灌木主要有马缨花Rhododendron delavayi、长圆叶梾木Cornus oblonga、长苞小檗Berberis jamesiana、大白花杜鹃 Rhododendron decorum、高丛珍珠梅Sorbaria arborea、黑穗箭竹Fargesia melanostachys等, 草本主要有白顶早熟禾Poa acroleuca、野雉尾金粉蕨Onychium japonicum、茅叶荩草Arthraxon prionodes、西南野古草Arundinella hookeri、线纹香茶菜Rabdosia lophanthoides、青蒿Artemisia carvifolia、菜蕨Callipteris esculenta等。
2 研究方法 2.1 样方设置及调查方法2014年9月-11月, 对集中分布有澜沧黄杉的群落进行调查, 共设置30个20m×20m的样方, 分别调查样方内乔木种类、个体数、高度、胸径、盖度、冠幅等指标, 并记录了样方的海拔、坡向、坡度、坡位、森林郁闭度等环境因子及人为干扰情况(表 1)。
| 样方号 Plot code |
海拔 Altitude/m |
坡向 Slope aspect |
坡度 Gradient/(°) |
坡位 Slope position |
郁闭度 Canopy density |
人为干扰 Anthropogenic disturbance |
| 1 | 3102 | E | 30 | 中 | 0.60 | 砍伐、放牧 |
| 2 | 3043 | N | 20 | 中 | 0.65 | 砍伐、放牧、开荒 |
| 3 | 3043 | N | 25 | 中 | 0.70 | 砍伐、放牧 |
| 4 | 2530 | E | 20 | 中 | 0.75 | 放牧 |
| 5 | 2554 | EN | 25 | 上 | 0.65 | 放牧 |
| 6 | 2918 | N | 26 | 下 | 0.70 | 放牧 |
| 7 | 3002 | N | 20 | 中 | 0.70 | 放牧 |
| 8 | 3056 | N | 25 | 中 | 0.70 | 放牧、开荒 |
| 9 | 2403 | E | 20 | 中 | 0.75 | 放牧 |
| 10 | 2418 | E | 20 | 下 | 0.70 | 放牧 |
| 11 | 2555 | S | 20 | 下 | 0.40 | 砍伐、放牧 |
| 12 | 2562 | N | 40 | 中 | 0.60 | 砍伐、放牧 |
| 13 | 3101 | W | 25 | 中 | 0.70 | 砍伐、放牧 |
| 14 | 3072 | W | 30 | 下 | 0.70 | 砍伐、放牧 |
| 15 | 2610 | EN | 25 | 下 | 0.50 | 砍伐、放牧 |
| 16 | 2718 | EN | 30 | 中 | 0.70 | 无 |
| 17 | 2972 | N | 27 | 中 | 0.50 | 放牧 |
| 18 | 3282 | N | 30 | 中 | 0.65 | 无 |
| 19 | 2663 | WN | 25 | 下 | 0.65 | 砍伐、放牧 |
| 20 | 2578 | N | 25 | 下 | 0.60 | 砍伐、放牧 |
| 21 | 2458 | EN | 25 | 下 | 0.60 | 砍伐、放牧 |
| 22 | 2640 | N | 25 | 下 | 0.60 | 砍伐、放牧 |
| 23 | 2645 | N | 25 | 下 | 0.70 | 砍伐、放牧 |
| 24 | 2616 | N | 30 | 下 | 0.50 | 砍伐、放牧 |
| 25 | 2617 | N | 40 | 中 | 0.70 | 砍伐、放牧 |
| 26 | 2624 | N | 35 | 中 | 0.65 | 砍伐、放牧 |
| 27 | 2661 | EN | 35 | 中 | 0.75 | 砍伐、放牧 |
| 28 | 2781 | ES | 35 | 中 | 0.75 | 无 |
| 29 | 2626 | N | 25 | 中 | 0.65 | 砍伐、放牧 |
| 30 | 2622 | N | 30 | 下 | 0.50 | 砍伐、放牧 |
重要值是确定植物群落类型的重要依据, 本文分别计算每个样方中乔木树种的重要值, 依据重要值排名第一的乔木物种来命名澜沧黄杉种群所在的森林群落类型。计算公式:重要值=(相对密度+相对高度+相对显著度)/3×100 [16]。其中:相对密度=某个种的个体数/全部植物的个体数;相对高度=某个种的高度/所有种高度之和×100;相对显著度=某个种的胸高断面积/样方中全部个体胸高断面积总和×100。
2.3 种群结构的划分对于物种数量稀少又无解析木资料的情况下, 主要采用“空间代替时间”的方法, 采用径级结构代替年龄结构的分析方法进行种群结构特征分析[17-18]。本文将澜沧黄杉按胸径(DBH)大小划分为10个龄级:I级(DHB < 2cm)、II级(2cm≤DHB < 5 cm)、Ⅲ-Ⅸ级(采用上限排外法每隔10cm划分为1级)、Ⅹ级(DHB≥75cm)。并对不同群落类型中澜沧黄杉种群各龄级植株个体数进行统计, 绘制种群结构图。
2.4 种群动态量化分析参考陈晓德[19]植物种群与群落结构动态量化分析方法, 定量描述不同群落类型澜沧黄杉种群动态, 计算公式如下:
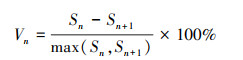
|
(1) |
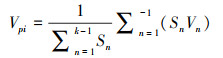
|
(2) |
式(1)中, Vn为种群内各龄级间个体数量的变化动态;式(2)中Vpi为整个种群的数量变化动态指数(忽略外部干扰), Sn、Sn+1分别为第n和n+1级种群个体数, k为种群年龄级数量;而当考虑未来的外部干扰时, Vpi还与年龄级数量(k)以及各龄级内的个体数(Sn)相关, 可进一步将式(2)修正为:
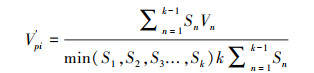
|
(3) |
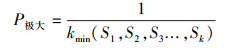
|
(4) |
式中, Vn、Vpi、V′pi取正、负、0值时分别反映种群个体数量的增长、衰退和稳定的结构动态关系。仅当P(种群对外界干扰所承担的风险概率)取值为最大时才会对种群动态V′pi构成最大的影响。
2.5 静态生命表的编制及存活曲线研究区域内的澜沧黄杉种群为天然林, 种群多世代重叠, 调查数据不能完全满足静态生命表的3个假设[20], 而且在调查中存在系统误差, 在生命表编制中会出现死亡率为负值的情况, 故而采用匀滑技术进行处理[21]。本文分别对不同群落类型下澜沧黄杉各龄级的个体数进行统计, 按照静态生命表编制方法进行编制[22], 并绘制存活曲线。
为确定澜沧黄杉属于Deevey-Ⅱ型还是Deevey-Ⅲ型, 本文采用Hett和Loucks[23]提出的数学模型对其检验, 即用指数方程Nx=N0e-bx和幂函数方程Nx=N0x-b分别描述Deevey-Ⅱ、Deevey-Ⅲ型存活曲线。式中Nx、N0分别代表x龄级内存活数和种群形成初期的个体数, b为死亡率。
2.6 生存分析为了更好地揭示澜沧黄杉种群的动态变化规律, 应用生存分析中的种群生存率函数Sx、种群累计死亡率函数Fx、种群死亡密度函数fx以及种群危险率函数λx来分析澜沧黄杉种群的生存现状, 参考杨凤翔等[24]的计算方法。公式如下:

|
(5) |
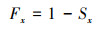
|
(6) |
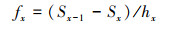
|
(7) |

|
(8) |
式中, Px为存活频率, hx为区间长度, qx为死亡频率。根据以上函数绘制生存率曲线、积累死亡率曲线、死亡密度曲线和危险率曲线。
2.7 时间序列预测时间序列分析是研究种群未来年龄结构和发展趋势的理想手段[25], 移动平均法作为时间序列预测中的常用方法, 在预测种群动态方面已经得到广泛应用, 本研究按照一次移动平均法[26-27]对澜沧黄杉种群年龄结构进行模拟和预测, 计算公式如下:
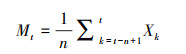
|
(9) |
式中, n为需要预测的时间, t代表龄级, Xk为当前k龄级种群存活数, Mt表示经过未来n年时t龄级的种群存活数。应用以上模型, 以澜沧黄杉种群总和各径级植株株数为原始数据, 按照一次平均推移法计算。本文对经过2、4、6、8径级时间后的澜沧黄杉种群各径级的个体数量进行预测。
3 结果与分析 3.1 种群的结构特征 3.1.1 群落类型划分30个样方乔木树种的重要值计算结果见表 2。样方1-10、样方11-20、样方21-30中重要值排名第一位的乔木树种分别为澜沧黄杉、冷杉属(长苞冷杉和川滇冷杉)、云南松, 因此, 将澜沧黄杉种群确定为三种不同的森林群落类型, 即澜沧黄杉林、冷杉林和云南松林。
| 样方号 Plot code |
重要值排名Important values ranking | ||||
| 1 | 2 | 3 | 4 | 5 | |
| 1 | 澜沧黄杉(63.58) | 云南铁杉(13.76) | 华山松(8.85) | 油麦吊云杉(7.52) | 云南松(6.29) |
| 2 | 澜沧黄杉(74.08) | 云南铁杉(10.64) | 华山松(7.5) | 云南黄果冷杉(4.71) | 云南松(3.08) |
| 3 | 澜沧黄杉(71.95) | 云南铁杉(10.14) | 华山松(6.99) | 油麦吊云杉(3.56) | 五裂槭(3.93) |
| 4 | 澜沧黄杉(68.18) | 云南铁杉(12.85) | 云南松(4.84) | 华山松(4.66) | 五裂槭(4.66) |
| 5 | 澜沧黄杉(71.11) | 云南铁杉(11.46) | 华山松(8.72) | 青榨槭(4.56) | 香白蜡树(4.15) Fraxinus suaveolens |
| 6 | 澜沧黄杉(89.88) | 丽江云杉(4.11) | 华山松(3.22) | 川滇高山栎(2.79) | - |
| 7 | 澜沧黄杉(92.97) | 华山松(2.65) | 微毛樱桃(2.3) Cerasus clarofolia |
川滇高山栎(2.07) | - |
| 8 | 澜沧黄杉(64.53) | 云南铁杉(17.06) | 丽江云杉(9.79) | 五裂槭(4.48) | 华山松(4.13) |
| 9 | 澜沧黄杉(63.68) | 华山松(15.39) | 云南松(8.98) | 云南铁杉(8.06) | 川滇高山栎(3.89) |
| 10 | 澜沧黄杉(68.35) | 华山松(11.33) | 云南松(7.5) | 五裂槭(6.92) | 冠萼花楸(2.91) |
| 11 | 长苞冷杉(35.77) | 云南铁杉(23.05) | 澜沧黄杉(17.87) | 喜马拉雅红豆杉(6.19) | 绿叶润楠(4.38) Machilus viridis |
| 12 | 长苞冷杉(37.76) | 澜沧黄杉(34.23) | 槲栎(9.66) | 华山松(6.25) | 云南松(3.26) |
| 13 | 川滇冷杉(41.66) | 澜沧黄杉(34.34) | 青榨槭(8.04) | 山杨(3.92) | 锐齿花楸(3.02) Sorbus arguta |
| 14 | 长苞冷杉(49.83) | 澜沧黄杉(35.71) | 华山松(9.19) | 山杨(3.09) | 华椴(2.17) Tilia chinensis var. chinensis |
| 15 | 长苞冷杉(43.18) | 澜沧黄杉(37.62) | 槲栎(8.82) | 华山松(5.08) | 五裂槭(2.81) |
| 16 | 长苞冷杉(34.47) | 澜沧黄杉(27) | 云南铁杉(19.85) | 青榨槭(7.33) | 山杨(5.00) |
| 17 | 长苞冷杉(32.34) | 丽江云杉(22.57) | 川滇高山栎(20.61) | 澜沧黄杉(20.34) | 青榨槭(2.32) |
| 18 | 长苞冷杉(27.34) | 澜沧黄杉(26.33) | 云南铁杉(20.5) | 华山松(13.12) | 川滇冷杉(5.78) |
| 19 | 川滇冷杉(38.95) | 澜沧黄杉(36.24) | 青榨槭(12.32) | 山杨(4.22) | 五裂槭(2.35) |
| 20 | 长苞冷杉(36.5) | 澜沧黄杉(32.18) | 青榨槭(22.53) | 华山松(5.21) | 槲栎(2.13) |
| 21 | 云南松(42.33) | 华山松(22.12) | 五裂槭(18.34) | 澜沧黄杉(15.32) | 喜马拉雅红豆杉(1.89) Taxus wallichiana |
| 22 | 云南松(38.74) | 澜沧黄杉(30.52) | 青榨槭(13.22) | 川滇冷杉(7.94) | 山杨(3.75) |
| 23 | 云南松(36.22) | 青榨槭(22.75) | 华山松(20.32) | 澜沧黄杉(17.88) | 五裂槭(1.72) |
| 24 | 云南松(43.24) | 澜沧黄杉(32.89) | 长苞冷杉(12.15) | 华山松(6.24) | 五裂槭(2.48) |
| 25 | 云南松(37.15) | 澜沧黄杉(33.28) | 华山松(17.13) | 青榨槭(5.78) | 高盆樱桃(3.68) Cerasus cerasoides |
| 26 | 云南松(33.55) | 槲栎(25.7) | 华山松(20.32) | 澜沧黄杉(15.88) | 滇青冈(4.55) |
| 27 | 云南松(32.22) | 川滇冷杉(27.88) | 澜沧黄杉(25.65) | 华山松(8.23) | 青榨槭(3.82) |
| 28 | 云南松(43.15) | 槲栎(22.23) | 云南铁杉(13.59) | 华山松(10.78) | 澜沧黄杉(7.89) |
| 29 | 云南松(37.88) | 澜沧黄杉(32.54) | 华山松(15.84) | 川滇冷杉(6.82) | 青榨槭(3.87) |
| 30 | 云南松(33.65) | 澜沧黄杉(31.32) | 川滇冷杉(26.53) | 云南铁杉(3.68) | 华山松(2.32) |
澜沧黄杉种群的龄级结构如图 1所示, 不同群落类型各龄级个体数存在明显的差异, 种群结构均呈不规则金字塔型。澜沧黄杉林、云南松林和冷杉林中第Ⅰ龄级和第Ⅱ龄级个体数分别占总个体数的36.11%、38.19%、31.73%, 第Ⅲ龄级至第Ⅵ龄级个体数分别占总个体数的55.09%、49.45%、43.27%, 第Ⅶ龄级至第Ⅹ龄级分别占总个体数的8.80%、12.36%、25%。这表明不同群落类型下澜沧黄杉种群均以中龄个体占优势, 幼龄个体和老龄个体少, 云南松林和冷杉林中第Ⅰ龄级个体数相对澜沧黄杉林中少, 冷杉林中的老龄个体相对于另外两种生境多。

|
| 图 1 不同群落类型下澜沧黄杉的龄级结构 Fig. 1 The age structure of P. forrestii popultion in different community types |
通过量化计算方法分析澜沧黄杉的种群动态, 结果如表 3所示。澜沧黄杉林中仅V2小于0, 整体呈现增长的结构动态关系;云冷杉林中V1、V3、V5、V9均小于0, 云南松林中V1、V5、V9均小于0, 这两种生境均呈现“衰退-增长-衰退”波动的结构动态关系。通过计算动态变化指数Vpi和V′pi, 三种群落类型下澜沧黄杉的总体数量动态变化指数V′pi均小于Vpi, 但都大于0, 其中V′pi更加趋于0, 说明澜沧黄杉种群均表现为增长型, 但种群结构增长性低, 趋近于稳定型。随机干扰风险极大值(P极大)分别为0.05(澜沧黄杉林)、0.05(冷杉林)、0.1(云南松林), 表明澜沧黄杉种群均对外界干扰具有较高的敏感性。
| 种群动态指数级 Dynamic index |
动态指数Dynamic index value/% | ||
| 澜沧黄杉林 P. forrestii community |
冷杉林 A.spp community |
云南松林 P. yunnanensis community |
|
| V1 | 9.76 | -62.5 | -69.23 |
| V2 | -15.91 | 66.67 | 42.31 |
| V3 | 22.73 | -27.27 | 20 |
| V4 | 32.35 | 18.18 | 50 |
| V5 | 21.74 | -47.06 | -45.45 |
| V6 | 44.44 | 11.76 | 45.45 |
| V7 | 50 | 80 | 66.67 |
| V8 | 60 | 33.33 | 50 |
| V9 | 0 | -66.67 | -50 |
| Vpi | 7.61 | 38.6 | 33.25 |
| V′pi | 0.38 | 1.93 | 3.33 |
静态生命表分析结果如表 4所示, 随着年龄增加, 澜沧黄杉种群的存活数(lx)逐渐减小, 个体期望寿命值(ex)也逐渐的降低, 寿命期望值在三种群落类型中均为第Ⅰ径级中最高, 表明种群在第Ⅰ径级即幼苗阶段具有较高的生命质量。
| 群落类型 Comunity |
龄级 Age stage |
胸径/cm Interva |
ax | lx | dx | qx | Lx | Tx | kx | ex | lnlx |
| 澜沧黄杉林 | Ⅰ | DHB < 2cm | 41 | 1000 | 73 | 0.07 | 964 | 4749 | 0.08 | 4.74 | 6.91 |
| P. forrestii community | Ⅱ | 2cm≤DHB < 5 cm | 38 | 927 | 49 | 0.05 | 903 | 3785 | 0.05 | 4.08 | 6.83 |
| Ⅲ | 5 cm≤DHB < 15 cm | 36 | 878 | 73 | 0.08 | 842 | 2882 | 0.09 | 3.28 | 6.78 | |
| Ⅳ | 15 cm≤DHB < 25 cm | 33 | 805 | 49 | 0.06 | 781 | 2040 | 0.06 | 2.53 | 6.69 | |
| Ⅴ | 25 cm≤DHB < 35 cm | 31 | 756 | 439 | 0.58 | 537 | 1259 | 0.87 | 1.66 | 6.63 | |
| Ⅵ | 35 cm≤DHB < 45 cm | 13 | 317 | 73 | 0.23 | 281 | 722 | 0.26 | 2.27 | 5.76 | |
| Ⅶ | 45cm≤DHB < 55 cm | 10 | 244 | 73 | 0.3 | 208 | 441 | 0.36 | 1.8 | 5.5 | |
| Ⅷ | 55 cm≤DHB < 65 cm | 7 | 171 | 49 | 0.29 | 147 | 233 | 0.34 | 1.36 | 5.14 | |
| Ⅸ | 65 cm≤DHB < 75 cm | 5 | 122 | 73 | 0.6 | 86 | 86 | 0.91 | 0.7 | 4.8 | |
| Ⅹ | DHB≥75cm | 2 | 49 | - | - | - | - | - | - | 3.89 | |
| 冷杉林 | Ⅰ | DHB < 2cm | 21 | 1000 | 143 | 0.14 | 929 | 4408 | 0.16 | 4.4 | 6.91 |
| A. spp community | Ⅱ | 2cm≤DHB < 5 cm | 18 | 857 | 143 | 0.17 | 786 | 3479 | 0.18 | 4.06 | 6.75 |
| Ⅲ | 5 cm≤DHB < 15 cm | 15 | 714 | 95 | 0.13 | 667 | 2693 | 0.14 | 3.77 | 6.57 | |
| Ⅳ | 15 cm≤DHB < 25 cm | 13 | 619 | 95 | 0.15 | 572 | 2026 | 0.17 | 3.27 | 6.43 | |
| Ⅴ | 25 cm≤DHB < 35 cm | 11 | 524 | 95 | 0.18 | 477 | 1454 | 0.2 | 2.77 | 6.26 | |
| Ⅵ | 35 cm≤DHB < 45 cm | 9 | 429 | 143 | 0.33 | 358 | 977 | 0.4 | 2.28 | 6.06 | |
| Ⅶ | 45cm≤DHB < 55 cm | 6 | 286 | 48 | 0.17 | 262 | 619 | 0.19 | 2.16 | 5.66 | |
| Ⅷ | 55 cm≤DHB < 65 cm | 5 | 238 | 48 | 0.2 | 214 | 357 | 0.22 | 1.5 | 5.47 | |
| Ⅸ | 65 cm≤DHB < 75 cm | 4 | 190 | 95 | 0.5 | 143 | 143 | 0.7 | 0.75 | 5.25 | |
| Ⅹ | DHB≥75cm | 2 | 95 | - | - | - | - | - | - | 4.55 | |
| 云南松林 | Ⅰ | DHB < 2cm | 20 | 1000 | 150 | 0.15 | 925 | 3925 | 0.16 | 3.93 | 6.91 |
| P. yunnanensis | Ⅱ | 2cm≤DHB < 5 cm | 17 | 850 | 150 | 0.18 | 775 | 3000 | 0.2 | 3.53 | 6.75 |
| community | Ⅲ | 5 cm≤DHB < 15 cm | 14 | 700 | 100 | 0.14 | 650 | 2225 | 0.15 | 3.18 | 6.55 |
| Ⅳ | 15 cm≤DHB < 25 cm | 12 | 600 | 150 | 0.25 | 525 | 1575 | 0.29 | 2.63 | 6.4 | |
| Ⅴ | 25 cm≤DHB < 35 cm | 9 | 450 | 150 | 0.33 | 375 | 1050 | 0.41 | 2.33 | 6.11 | |
| Ⅵ | 35 cm≤DHB < 45 cm | 6 | 300 | 50 | 0.17 | 275 | 675 | 0.18 | 2.25 | 5.7 | |
| Ⅶ | 45cm≤DHB <55 cm | 5 | 250 | 100 | 0.4 | 200 | 400 | 0.51 | 1.6 | 5.52 | |
| Ⅷ | 55 cm≤DHB <65 cm | 3 | 150 | 50 | 0.33 | 125 | 200 | 0.4 | 1.33 | 5.01 | |
| Ⅸ | 65 cm≤DHB < 75 cm | 2 | 100 | 50 | 0.5 | 75 | 75 | 0.7 | 0.75 | 4.61 | |
| Ⅹ | DHB≥75cm | 1 | 50 | - | - | - | - | - | - | 3.91 | |
| ax:在x年龄级内存活个体数匀滑处理后的个体数Number of individuals after smoothing; lx: 在x年龄级开始时标准化存活的个体数Proportion organidms surviving from the beginning of the life table to age x; dx: x年龄级到x+1年龄级间的标准化死亡数Number of dead organisms from age to x+1; qx: 从x年龄级到x+1年龄级间隔期内死亡率Mortality rate age x to x+1; Lx: 从x年龄级到x+1龄级间平均存活的个体数Number of survivors from age x to x+1; Tx : 从x年龄级到超过x年龄级的个体总数Total number of alive organisms from x to x+1; Kx: 各年龄组的致死力Elimination rate; ex:进入x龄级个体的期望寿命Life expectation of individual in age x class; lnlx: lx的自然对数The natural logarithm of lx; 破折号(-)表示由于x为最后一个龄级而无法计算的数据; 龄级同图 1 | |||||||||||
本研究所作存活曲线是以径级相对龄级为横坐标, 以lnlx为纵坐标, 如图 2所示, 根据Deevey[28]的划分, 存活曲线可以分为三种类型, Ⅰ型是凸曲线, 该类型的种群绝大多数能活到该物种的年龄, 早期死亡率低, 但活到一定的生理年龄时, 基本上全部死亡;Ⅱ型是直线型, 也称对角线型, 该类型的种群各龄级的死亡率基本相同;Ⅲ型是凹曲线, 早期死亡率较高, 一旦活到某个年龄, 死亡率就降低。采用Hett和Loucks提出的数学模型对其检验, 结果如表 5所示, 三种群落类型下两种模型的拟合结果均达到了显著水平(P<0.05), 指数函数的R2均大于幂函数, 说明指数函数模型拟合最优, 因此断定种群的存活曲线均趋近于Deevey-Ⅱ型, 表明种群各龄级的死亡率基本相同。

|
| 图 2 不同群落类型下澜沧黄杉种群的存活曲线 Fig. 2 Survival curve of P.forrestii population in different community types |
| 群落类型Population type | 方程Equation | R2 | F | P |
| 澜沧黄杉林P. forrestii community | N=8.029e-0.059x | 0.87 | 51.68 | 0.000 |
| N=7.945x-0.208 | 0.63 | 13.63 | 0.006 | |
| 冷杉林A. Spp community | N=7.472e-0.042x | 0.92 | 94.72 | 0.000 |
| N=7.487x-0.152 | 0.73 | 21.52 | 0.002 | |
| 云南松林P. yunnanensis community | N=7.812e-0.058x | 0.92 | 88.31 | 0.000 |
| N=7.801x-0.212 | 0.71 | 19.3 | 0.002 |
不同群落类型澜沧黄杉的生存率和累积死亡率曲线如图 3所示。生存率和死亡率二者互补, 随着龄级的增加, 澜沧黄杉种群的生存率逐渐下降, 而累积死亡率逐渐上升。不同群落类型澜沧黄杉种群的生存率和累积死亡率分别在4.6龄级(澜沧黄杉林)、4.2龄级(冷杉林)和3.8龄级(云南松林)达到平衡, 反映出在云南松林中的种群较其它群落类型更早进入衰退期。

|
| 图 3 不同群落类型下澜沧黄杉种群生存率和累积死亡率 Fig. 3 Survival rate and cumulative mortality rate of P.forristii population in different community types |
不同群落类型下澜沧黄杉种群死亡密度和危险率如图 4所示, 三种群落类型下澜沧黄杉种群的死亡密度变化趋势基本一致, 随着年龄的增加, 种群死亡密度下降, 澜沧黄杉林种群第Ⅰ龄级、第V龄级具有死亡高峰期, 第Ⅵ龄级后趋向平缓, 冷杉林和云南松林种群第Ⅰ龄级具有死亡高峰期, 第Ⅲ龄级后趋向平缓;危险率曲线整体波动较大, 澜沧黄杉林中第Ⅰ龄级至第IV龄级、冷杉林和云南松林中的第Ⅰ龄级至III龄级均呈下降趋势, 进入中龄以后, 个体死亡率相对稳定, 随着龄级的增加, 个体死亡危险率增加。表明澜沧黄杉种群从幼龄到中龄的过渡期生命状态稳定性差, 死亡的可能性较大, 进入老龄以后, 种群的死亡主要由衰老死亡构成, 比较可知, 云南松林澜沧黄杉种群危险率曲线波动起伏更大, 表明其受环境影响更明显。

|
| 图 4 不同群落类型下澜沧黄杉种群死亡密度和危险率 Fig. 4 Mortality dengsity and hazard rate of P.forristii population in different community type |
时间序列分析结果如表 6所示, 在经历未来2、4、6、8龄级的时间之后, 种群初期个体数量减少, 澜沧黄杉林种群从Ⅳ龄级开始个体数量逐渐增加, 云南松林和冷杉林在第Ⅲ龄级开始种群个体数量逐渐增加, 但在中龄时期种群个体数量均呈不规则的波动。综合整个种群的发展趋势来看, 澜沧黄杉种群在一定时间内, 仍然可以维持更新, 保持群落增长, 但由于老龄个体的增加, 未来仍然会出现不同程度的衰退, 甚至濒危。
| 龄级 Age class |
澜沧黄杉林 P.forristii community |
冷杉林 A. spp community |
云南松林 P. yunnanensis community |
||||||||||||||
| 原始数据 Primary data |
M2(1) | M4(1) | M6(1) | M8(1) | 原始数据 Primary data |
M2(1) | M4(1) | M6(1) | M8(1) | 原始数据 Primary data |
M2(1) | M4(1) | M6(1) | M8(1) | |||
| Ⅰ | 41 | 9 | 8 | ||||||||||||||
| Ⅱ | 37 | 39 | 24 | 17 | 26 | 17 | |||||||||||
| Ⅲ | 44 | 41 | 8 | 16 | 15 | 21 | |||||||||||
| Ⅳ | 34 | 39 | 39 | 11 | 10 | 13 | 12 | 14 | 15 | ||||||||
| Ⅴ | 23 | 29 | 35 | 9 | 10 | 13 | 6 | 9 | 15 | ||||||||
| Ⅵ | 18 | 21 | 30 | 33 | 17 | 13 | 11 | 13 | 11 | 9 | 11 | 13 | |||||
| Ⅶ | 10 | 14 | 21 | 28 | 15 | 16 | 13 | 14 | 6 | 9 | 9 | 13 | |||||
| Ⅷ | 5 | 8 | 14 | 22 | 27 | 3 | 9 | 11 | 11 | 12 | 2 | 4 | 6 | 9 | 11 | ||
| Ⅸ | 2 | 4 | 9 | 15 | 22 | 2 | 3 | 9 | 10 | 11 | 1 | 2 | 5 | 6 | 10 | ||
| Ⅹ | 2 | 2 | 5 | 10 | 17 | 6 | 4 | 7 | 9 | 9 | 2 | 2 | 3 | 5 | 7 | ||
| (1)表示一次移动平均;M2, M4, M6, M8分别表示对经历2, 4, 6, 8龄级时间后澜沧黄杉的种群大小的预测; 原始数据为野外的实际调查数据 | |||||||||||||||||
本研究澜沧黄杉种群的栖息地植被类型为澜沧黄杉林、冷杉林与云南松林, 年龄结构均呈不规则金字塔型, 动态指数结果显示三种群落类型下种群均为增长型, 趋近于稳定, 虽然澜沧黄杉种群暂时能适应不同的生存环境, 但由于种群I龄级个体数较少, 幼龄个体无法补充中老龄个体的发展, 这种增长可能难以维持, 当生存环境受到干扰后, 种群极易衰退, 随机干扰风险极大值也显示种群对生存环境具有较高的敏感性, 这与裸果木和沙拐枣群落结构较为相似[29-30]。从野外调查可知, 澜沧黄杉分布相对狭窄, 喜阴坡、半阴坡或“V”型河谷等湿润、荫蔽的生长环境, 种子所到达的生境, 决定它能否成功更新, 生境中的土壤、水分、光隙和植物邻居等因子也限制着幼苗的生长机会[31], 澜沧黄杉种子带翅, 成熟后随风散布, 当种子扩散到干燥、土壤瘠薄的环境中时可能不利于植物种群更新, 种子难以萌发或萌发后其幼苗可能不适宜环境, 导致种群幼龄个体少, 形成种群发展的瓶颈。
4.2 澜沧黄杉种群动态趋势不同时期由于栖息地环境特征、人为干扰情况的不同, 物种种群调查的结果会存在差异, 但在难以获得澜沧黄杉动态生命表数据的情况下, 进行静态生命表的编制对于预测种群动态变化具有重要参考价值。静态生命表是用同一时期收集的种群所有个体的径级编制而成, 反映了多个时代重叠的年龄动态历程中的一个特定时间, 而不是对这一种群全部生活史的追踪[32]。静态生命表显示, 澜沧黄杉种群的存活曲线均趋向于Deevey-Ⅱ型, 说明种群有相对稳定的死亡率, 种群数量动态分析显示澜沧黄杉种群均为增长型, 趋近于稳定, 分析其原因, 可能是由于种群现存中龄个体数量较多, 一定的时间内种群仍然可以维持更新, 保持群落的增长, 时间序列预测也证明了这一结论, 这与羽叶丁香的种群特征较为相似[33]。
通过存活曲线可以看出澜沧黄杉的第Ⅰ-Ⅳ龄级在冷杉林和云南松林中下降幅度更大, 种群的生存率和累积死亡率反映出云南松林种群较早进入衰退期, 冷杉林次之, 同时生存分析中的死亡密度和危险率曲线显示澜沧黄杉种群在幼龄期存在较高的死亡风险, 这种死亡风险在云南松林和冷杉林中表现更为突出, 因此, 云南松林和冷杉林种群在中幼龄阶段受环境影响更明显, 这种明显的环境影响可能是由于中幼龄阶段种群与其他物种争夺生存资源和空间而导致的。有研究表明, 幼苗阶段是森林更新过程中最重要和最敏感的阶段[34-35], 有限的资源和空间往往成为种群更新的限制因子[36], 在云南松林和冷杉林中, 澜沧黄杉种群的中幼龄个体不仅存在个体竞争, 还存在与其他物种生存资源和空间的竞争, 因而受环境影响更明显, 同时, 也有研究发现不同植被类型对于植物幼苗生长的适宜度存在差异[31], 本研究也同样发现在不同的植被类型下澜沧黄杉种群幼苗死亡率不同, 云南松林和冷杉林中种群幼龄阶段的死亡率更高, 因此, 澜沧黄杉林更适宜种群的生长。
4.3 澜沧黄杉种群的保护和恢复对策导致物种濒危的因素有很多, 如种子的活力、休眠期、发芽率、种群的光合、水分、生理、生态适应方式、生态位、生殖特征等[37], 本文发现目前澜沧黄杉种群濒危的原因在于分布范围狭窄, 幼龄个体不足, 幼龄期植株的死亡率较高。但是否还存在其他的致危因素, 还有待进一步研究, 因此, 未来还需通过不断加大科研力度等关键措施, 以此来快速保护和恢复澜沧黄杉天然种群。种群天然更新及生长繁殖机制复杂, 除与自身的生理生态状况有关之外, 人为干扰也是其重要因素[38], 通过野外调查可知, 调查区域的样方16、18、28处于保护区内, 澜沧黄杉种群受到了较好的保护, 而保护区以外的种群栖息地受到放牧、开荒等严重的人为干扰, 对种群生存产生了较大威胁, 严重影响了种群的生存。另外, 澜沧黄杉因树形高大、笔直, 被当地居民采伐后用于房屋建造、围栏、养蜂、薪柴等, 使其种群遭到破坏。要加强对澜沧黄杉的保护, 关键是杜绝人类活动对生境的破坏, 禁止采伐, 在加大科学研究力度的同时要不断加强对澜沧黄杉的保护宣传力度, 扩大完善相应自然保护区的建立, 促使其种群恢复正常更新。
| [1] |
Gurevitch J, Scheiner SM, Fox GA. The Ecology of plants. Sunderland, Massachusetts, USA: Sinauer Associates, Inc., 2002.
|
| [2] |
金慧, 赵莹, 刘丽杰, 贾翔, 代玉红, 秦立武, 王超, 尹航. 长白山区濒危植物朝鲜崖柏种群数量特征及动态. 应用生态学报, 2019, 30(5): 1563-1570. |
| [3] |
Röder D, Kiehl K. Population structure and population dynamic of Pulsatilla patens (L.) Mill. in relation to vegetation characteristics. Flora-Morphology, Distribution, Functional Ecology of Plants, 2006, 201(6): 499-507. DOI:10.1016/j.flora.2005.11.001 |
| [4] |
吴承祯, 洪伟, 谢金寿, 吴继林. 珍稀濒危植物长苞铁杉种群生命表分析. 应用生态学报, 2000, 11(3): 333-336. DOI:10.3321/j.issn:1001-9332.2000.03.003 |
| [5] |
Hercombe PA. Tree life tables. Bioscience, 1987, 37(8): 557-568. DOI:10.2307/1310666 |
| [6] |
胡刚, 黎洁, 覃盈盈, 胡宝清, 刘熊, 张忠华. 广西北仑河口红树植物种群结构与动态特征. 生态学报, 2018, 38(9): 3022-3034. |
| [7] |
李翔, 侯璐, 李双喜, 李颖岳. 濒危树种庙台槭种群数量特征及动态分析. 植物科学学报, 2018, 36(4): 524-533. |
| [8] |
刘方炎, 李昆, 廖声熙, 崔永忠. 濒危植物翠柏的个体生长动态及种群结构与种内竞争. 林业科学, 2010, 46(10): 23-28. DOI:10.11707/j.1001-7488.20101004 |
| [9] |
云南省林业厅, 云南省林业科学院, 国家林业局云南珍稀濒特森林植物保护和繁育实验室. 云南国家重点保护野生植物. 昆明: 云南科技出版社, 2005: 119-120.
|
| [10] |
傅立国. 中国植物红皮书--稀有濒危植物(第一册). 北京: 科学出版社, 1992: 14-118.
|
| [11] |
中国科学院中国植物志编辑委员会. 中国植物志. 北京: 科学出版社, 1991.
|
| [12] |
吴征镒. 中国种子植物属的分布区类型. 云南植物研究, 1991(S4): 135-139. |
| [13] |
李楠. 黄杉属植物地理学的研究. 植物研究, 1993, 13(4): 404-411. |
| [14] |
孙卫邦, 孔繁才, 周元, 李崇仁. 黄杉属植物在云南的资源现状及保护利用研究. 广西植物, 2003, 23(1): 15-18. DOI:10.3969/j.issn.1000-3142.2003.01.004 |
| [15] |
孔维民, 刘鹏程, 马方莲, 李敏敏. 澜沧黄杉在迪庆州的分布现状与保护建议. 林业调查规划, 2018, 43(4): 89-91, 103-103. DOI:10.3969/j.issn.1671-3168.2018.04.020 |
| [16] |
Curtis J T, McIntosh R P. An upland forest continuum in the Prairie-Forest Border Region of Wisconsin. Ecology, 1951, 32(3): 476-496. DOI:10.2307/1931725 |
| [17] |
Dimobe K, Wala K, Dourma M, Kiki M, Woegan Y, Folega F, Batawila K, Akpagana K. Disturbance and population structure of plant communities in the wildlife reserve of Oti-Mandouri in Togo (West Africa). Annual Research & Review in Biology, 2014, 4(15): 2501-2516. |
| [18] |
王魏, 刘灿然, 马克平, 于顺利. 东灵山两个落叶阔叶林中辽东栎种群结构和动态. 植物学报, 1999, 41(4): 425-432. |
| [19] |
陈晓德. 植物种群与群落结构动态量化分析方法研究. 生态学报, 1998, 18(2): 214-217. DOI:10.3321/j.issn:1000-0933.1998.02.017 |
| [20] |
于大炮, 周莉, 董百丽, 代力民, 王庆礼. 长白山北坡岳桦种群结构及动态分析. 生态学杂志, 2004, 23(5): 30-34. DOI:10.3321/j.issn:1000-4890.2004.05.006 |
| [21] |
江洪. 云杉种群生态学. 北京: 中国林业出版社, 1992: 8-40.
|
| [22] |
孙儒泳, 李博. 普通生态学. 北京: 高等教育出版社, 1993.
|
| [23] |
Hett J M, Loucks O L. Age structure. models of balsam fir and eastern hemlock. Journal of Ecology, 1976, 64(3): 1029-1044. DOI:10.2307/2258822 |
| [24] |
杨凤翔, 王顺庆, 徐海根, 李邦庆. 生存分析理论及其在研究生命表中的应用. 生态学报, 1991, 11(2): 153-158. DOI:10.3321/j.issn:1001-9332.1991.02.013 |
| [25] |
卢杰, 郭其强, 郑维列, 徐阿生. 藏东南高山松种群结构及动态特征. 林业科学, 2013, 49(8): 154-160. |
| [26] |
肖宜安, 何平, 李晓红, 邓洪平. 濒危植物长柄双花木自然种群数量动态. 植物生态学报, 2004, 28(2): 252-257. DOI:10.3321/j.issn:1005-264X.2004.02.017 |
| [27] |
杨彪, 张全建, 龚旭, 段晨松, 张远彬. 雅砻江冬麻豆(Salweenia bouffordiana)种群结构与动态特征. 生态学报, 2020, 40(4): 1184-1194. |
| [28] |
Deevey E S Jr. Life tables for natural populations of animals. The Quarterly Review of Biology, 1947, 22(4): 283-314. DOI:10.1086/395888 |
| [29] |
王立龙, 王亮, 张丽芳, 刘玉洋, 徐世健. 不同生境下濒危植物裸果木种群结构及动态特征. 植物生态学报, 2015, 39(10): 980-989. DOI:10.17521/cjpe.2015.0095 |
| [30] |
解婷婷, 苏培玺, 周紫鹃, 张海娜, 李善家. 荒漠绿洲过渡带沙拐枣种群结构及动态特征. 生态学报, 2014, 34(15): 4272-4279. |
| [31] |
李宁, 白冰, 鲁长虎. 植物种群更新限制—从种子生产到幼树建成. 生态学报, 2011, 31(21): 6624-6632. |
| [32] |
张志祥, 刘鹏, 蔡妙珍, 康华靖, 廖承川, 刘春生, 楼中华. 九龙山珍稀濒危植物南方铁杉种群数量动态. 植物生态学报, 2008, 32(5): 1146-1156. DOI:10.3773/j.issn.1005-264x.2008.05.019 |
| [33] |
姜在民, 和子森, 宿昊, 赵涵, 蔡靖. 濒危植物羽叶丁香种群结构与动态特征. 生态学报, 2018, 38(7): 2471-2480. |
| [34] |
张婕, 上官铁梁, 段毅豪, 郭微, 刘卫华, 郭东罡. 灵空山辽东栎种群年龄结构与动态. 应用生态学报, 2014, 25(11): 3125-3130. |
| [35] |
Silvertown J W. Introduction to Plant Population Ecology. London: Longman Group United Kingdom, 1982.
|
| [36] |
项小燕, 吴甘霖, 段仁燕, 王志高, 张中信, 王广艳, 张小平. 大别山五针松种群结构及动态研究. 长江流域资源与环境, 2016, 25(1): 55-62. DOI:10.11870/cjlyzyyhj201601007 |
| [37] |
张文辉, 祖元刚, 刘国彬. 十种濒危植物的种群生态学特征及致危因素分析. 生态学报, 2002, 22(9): 1512-1520. DOI:10.3321/j.issn:1000-0933.2002.09.021 |
| [38] |
Vacek Z, Vacek S, Podrázský V, Bílek L, Štefančík I, Moser W K, Bulušek D, Král J, Remeš J, Králíček I. Effect of tree layer and microsite on the variability of natural regeneration in autochthonous beech forests. Polish Journal of Ecology, 2015, 63(2): 233-246. DOI:10.3161/15052249PJE2015.63.2.007 |