 2022, Vol. 42
2022, Vol. 42文章信息
- 陈彦云, 夏皖豫, 赵辉, 曾明
- CHEN Yanyun, XIA Wanyu, ZHAO Hui, ZENG Ming
- 粉垄耕作对耕地土壤酶活性、微生物群落结构和功能多样性的影响
- Effects of deep vertical rotary tillage on soil enzyme activity, microbial community structure and functional diversity of cultivated land
- 生态学报. 2022, 42(12): 5009-5021
- Acta Ecologica Sinica. 2022, 42(12): 5009-5021
- http://dx.doi.org/10.5846/stxb202106241676
-
文章历史
- 收稿日期: 2021-06-24
- 修订日期: 2021-11-17





2. 宁夏大学生态环境学院, 银川 750021;
3. 宁夏科技特派员创业指导服务中心, 银川 750021
2. School of Ecological Environment, Ningxia University, Yinchuan 750021, China;
3. Ningxia Science and Technology Commissioner Entrepreneurship Guidance Service Center, Yinchuan 750021, China
土壤微生物是生态系统的重要组成部分, 也是土壤生态系统的核心, 直接或间接参与调节土壤养分循环、能量流动、有机质转换、土壤肥力形成、污染物的降解及环境净化等, 特别在维持生物多样性、改善土壤碳固存、生态系统功能和自然和农业生态系统中的养分循环方面发挥着重要作用[1-3]。土壤微生物组成的质与量的变化是土壤健康状况的重要敏感指示, 土壤微生物多样性可以定义为生命的丰富度, 代表着微生物群落的稳定性, 也反映土壤生态机制及土壤胁迫对微生物群落的影响, 因此对土壤微生物群落结构和功能的研究一直是土壤与植物营养学的研究热点[4-6]。研究表明, 土壤微生物群落受到各种因素的影响, 包括耕作方式、气候、施肥和pH, 其中耕作方式是对土壤微生物的影响尤为关键[7-10]。
玉米(Zea mays L.)是禾本科玉蜀黍属一年生草本植物, 是重要的粮食作物和饲料作物, 也是全世界总产量最高的农作物。耕作方式一直是提高土壤肥力和提高作物生产力的最重要的农业实践之一, 大量研究表明不同耕作方式对土壤特性和作物产量的影响[11-14]。宁夏引黄灌区的玉米种植大多采用传统耕作。然而, 传统耕作导致水土流失加剧, 环境污染加剧, 土壤退化, 犁盘紧凑且封闭, 阻碍了深层土层中土壤水、肥和热量的循环影响生态系统功能[15-18]。粉垄耕作是一种集深松、深耕、旋耕、垂直耕作优点于一体的深耕技术[19]。它由配备六个立式螺旋钻的新型强力机器进行, 可以在不扰乱土壤层和生产新的硬质犁的情况下打碎压实的犁盘并松动土层[20]。有研究表明, 粉垄耕作模式对农田土壤质量、蓄水能力、作物产量、根系活力以及土壤微生物生物量和微生物多样性等方面的有益作用更强[19, 21-24]。近年来, 粉垄耕作技术在农业生产方面取得了巨大成功。然而, 目前尚不清楚粉垄耕作对耕地土壤微生物群落和土壤生态系统功能的影响。
本研究利用高通量16S rRNA和ITS基因Illumina测序和Biolog-ECO方法, 系统地研究了粉垄耕作对土壤中微生物群落结构和土壤微生物功能多样性的影响。本研究的目的是:(1)探寻不同耕作方式下耕地土壤酶活性、土壤微生物多样性和群落结构的差异;(2)分析不同耕作方式下土壤微生物群落功能多样性及功能基团;(3)粉垄耕作如何影响土壤微生物结构、酶活性与微生物功能多样性间的相关性, 使玉米产量得到提升。本研究为粉垄耕作技术在宁夏引黄灌区作物栽培中的应用发展提供了科学理论依据和农业实践理论支撑。
1 材料与方法 1.1 试验样地试验于2019年4月-10月在宁夏回族自治区石嘴山市平罗县头闸村(106°63′N、38°95′E)进行, 属温带大陆性气候, 海拔1091 m, 日照充足、昼夜温差大、干旱少雨、年蒸发量1755 mm, 平均风速2-3 m/s, 无霜期171 d, 年降雨量为184 mm, 年均气温8.21℃。试验区土壤为黏土, 土壤状况为全磷0.86 g/kg, 全氮1.10 g/kg, 碱解氮76.92 mg/kg, 速效磷34.69 mg/kg, 速效钾214.03 mg/kg, 有机碳11.42 mg/kg, 总盐0.67 g/kg, 含水量12.38%, pH值为8.5。
1.2 试验处理与土样采集采用大区试验, 试验面积为900 m2 (60 m×15 m), 设置3个处理分别为传统耕作深度20 cm(CK)、粉垄耕作深度35 cm(FL1)、粉垄耕作深度50 cm(FL2)。玉米出苗后将各处理的样地平均分为4个67.5 m2(15 m×4.5 m)的小区, 即为每个处理的4次重复, 每个小区之间设1 m宽走道, 重复小区间设走道, 宽1 m, 四周设置保护行, 宽3 m。
供试玉米品种为“迪卡5号”, 采用吴忠市伊禾农机作业服务有限公司提供的悬挂式粉垄机作业, 全面深松垂直旋磨粉碎土壤, 粉垄后镇压。传统耕作采用拖拉机犁耙整地。2019年5月3日整地, 5月16日播种, 10月18日收获。播种密度为82500株/hm2, 肥料为控释肥0.12 kg/m2(N-P-K:30-12-5), 作为底肥一次性施入, 各处理田间管理均一致。土壤采集时间为10月18日, 以五点取样法在0-20 cm, 20-40 cm耕作层取土样, 每个土壤3个平行样品。将土壤样品装入无菌袋中, 一部分鲜土过2 mm筛后4℃保存, 用于土壤酶活性及Biology-ECO测定, 另一部分鲜土过1 mm筛-80℃保存, 用于土壤微生物群落结构的测定。
1.3 土壤酶活性的测定脲酶用靛酚蓝比色法测定, 以24 h后1 g土壤NH3-N的毫克数表示;土壤碱性磷酸酶采用磷酸苯二钠比色法, 以1 h后1g土壤中对硝基苯酚的毫克数表示;转化酶用3, 5-二硝基水杨酸比色法, 以24 h后1 g土壤葡萄糖的毫克数表示[25]。
1.4 土壤微生物功能多样性的测定土壤微生物功能多样性用Biolog方法进行测定[26, 27]:称取5.0 g土壤样品加入装有45 mL 0.85%无菌生理盐水的三角瓶中, 25℃ 150 r/min振荡30 min, 冰浴1 min, 后静置30 min, 获得土壤样品的微生物悬浮液。将土壤悬液梯度稀释为10-3 g/mL, 在超净工作台中用移液枪将制备好的土壤悬液接种至Biolog微平板的各孔中, 每孔150 μL, 盖好盖子置于25℃培养箱中暗培养7 d, 每隔24 h用酶标仪读取在590 nm与750 nm波长的数值。
Shannon指数(H)用于评估物种丰富度, 计算公式如下:
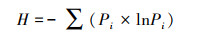
|
(1) |
式中, Pi为第i孔的相对吸光值与整个平板相对吸光值总和的比率。
McIntosh指数(U)用于评估群落均一度:
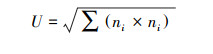
|
(2) |
式中, ni为第i孔的相对吸光值。
Simpson指数(D)用于评估常见种优势度的指数:
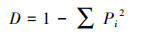
|
(3) |
式中, Pi为第i孔的相对吸光值与整个平板相对吸光值总和的比率。
1.5 DNA抽提和PCR扩增各土层FL1、FL2和CK一共18个土壤样品进行IlluminaMiSeq测序。根据E.Z.N.A.® soil DNA kit (Omega Bio-tek, Norcross, GA, U.S.)说明书进行微生物总DNA抽提, 使用1%的琼脂糖凝胶电泳检测DNA的提取质量, 使用NanoDrop2000测定DNA浓度和纯度;细菌引物采用338F (5′-ACTCCTACGGGAGGCA GCAG-3′)和806R (5′-GGACTACHVGGGTWTCTAAT-3′), 真菌引物采用ITS1F (5′-CTTGGTCATTTAGAGGAA GTAA-3′)和ITS2R (5′-GCTGCGTTCTTCATCGATGC-3′), 扩增程序如下:95℃预变性3 min, 27个循环(95℃变性30s, 55℃退火30s, 72℃延伸30s), 然后72℃稳定延伸10min, 最后在4℃进行保存(PCR仪:ABI GeneAmp® 9700型)。PCR反应体系为:5×TransStart FastPfu缓冲液4 μL, 2.5mmol·L-1 dNTPs 2μL, 上游引物(5μmol·L-1)0.8 μL, 下游引物(5μmol·L-1)0.8 μL, TransStart FastPfu DNA聚合酶0.4 μL, 模板DNA 10 ng, 补足至20 μL。
1.6 Illumina Miseq测序和测序数据处理将同一样本的PCR产物混合后使用2%琼脂糖凝胶回收PCR产物, 利用AxyPrep DNA Gel Extraction Kit (Axygen Biosciences, Union City, CA, USA)进行回收产物纯化, 2%琼脂糖凝胶电泳检测, 并用QuantusTM Fluorometer (Promega, USA)对回收产物进行检测定量。使用NEXTFLEX Rapid DNA-Seq Kit进行建库。利用Illumina公司的Miseq PE300平台进行测序(上海美吉生物医药科技有限公司)。
使用fastp[28]软件对原始测序序列进行质控, 使用FLASH[29]软件进行拼接。使用UPARSE软件, 根据97%[30, 31]的相似度对序列进行OTU聚类并剔除嵌合体。利用RDP classifier[32]对每条序列进行物种分类注释, 细菌比对Silva 16S rRNA数据库, 真菌比对UNITE ITS数据库, 设置比对阈值为70%。功能注释分别基于Tax4Fun和FunGuild用于细菌和真菌。
1.7 数据分析方差分析(ANOVA)采用SPSS 18.0分析软件(SPSS Inc., 美国)和Tukey检验, 在P=0.05显着性水平上进行。结构方程模型(SEM)用于评估假设的潜在因素, 使用IBM SPSS Amos 24进行分析。使用Microsoft Excel 2016, Origin 2016和R软件绘图。
2 结果Results 2.1 粉垄耕作对土壤酶活性及玉米产量的影响表 1列出了FL1、FL2和CK处理下不同土壤深度的土壤脲酶、碱性磷酸酶和转化酶活性。在不同处理下, 0-20 cm深度的3种酶活性高于20-40 cm深度。0-20 cm土层中, FL1处理的土壤脲酶活性显著高于CK处理、碱性磷酸酶和转化酶活性显著高于FL2和CK处理。20-40 cm土层中, FL1和FL2处理的脲酶活性比CK处理提高了28.80%和4.45%, 碱性磷酸酶活性比CK处理提高了30.95%和16.67%, FL1处理土壤转化酶比CK处理提高了25.61%, 但各处理间差异均没达到显著水平(P>0.05)。FL1、FL2处理产量分别为8.58 t/hm2和8.38 t/hm2, 与对照处理相比提高了37.9%和34.7%(P < 0.05)。
| 处理 Treatment |
脲酶 Urease/ (mg g-1 24h-1) |
碱性磷酸酶 Alkaline phosphatase/ (mg g-1 h-1) |
转化酶 Invertase/ (mg g-1 24h-1) |
产量 Yield/ (t/hm2) |
| CK-1 | 40.49±3.10b | 0.71±0.10b | 14.80±0.90b | 6.22±0.58b |
| FL1-1 | 50.08±4.70a | 0.81±0.06a | 18.41±1.13a | 8.58±0.45a |
| FL2-1 | 42.43±5.56ab | 0.69±0.00b | 14.48±0.20b | 8.38±0.66a |
| CK-2 | 27.40±0.40a | 0.42±0.06a | 4.53±0.95a | - |
| FL1-2 | 35.29±7.75a | 0.55±0.06a | 5.69±0.15a | - |
| FL2-2 | 28.62±2.60a | 0.49±0.10a | 4.36±1.70a | - |
| CK: 传统耕作深度20 cm traditional tillage with a depth of 20 cm;FL1: 粉垄耕作深度35 cm deep vertical rotary tillage with a depth of 35 cm;FL2: 粉垄耕作深度50 cm deep vertical rotary tillage with a depth of 50 cm;-1、-2分别表示0-20 cm土层、20-40 cm土层; 表中数据为平均值±标准差(n=3) | ||||
本研究中, 通过高通量测序技术检测所有土壤样品的微生物, 获得了30730个OTUs, 包括26668个细菌OTUs和4062个真菌OTUs。对各处理平均丰度大于1的所有土壤细菌和真菌OTUs进行了Flower plot分析。对于土壤细菌OTUs, 在0-20 cm和20-40 cm的FL1、FL2和CK处理共有2296个相同OTUs。0-20 cm土层中FL1、FL2和CK处理特有细菌OTUs分别是81、206和72个, 20-40 cm土层中FL1、FL2和CK处理特有细菌OTUs分别是135、83和194个(图 1)。对于土壤真菌OTUs, 在0-20 cm和20-40 cm的FL1、FL2和CK处理共有211个相同OTUs。0-20 cm土层中FL1、FL2和CK处理特有真菌OTUs分别是129、85和102个, 20-40 cm土层中FL1、FL2和CK处理特有真菌OTUs分别是67、129和88个(图 1)。在门水平上分析了样本中的细菌和真菌分布(图 1)。不同土层各样本中的细菌在门水平排名前10的物种组成相同(图 1)。不同土层各处理间优势菌门为变形杆菌、放线菌门、绿弯菌门和酸杆菌门, 达78.85%-81.83%。0-20 cm土层中的FL1、FL2和CK处理的细菌门差异不显著(P>0.05)。

|
| 图 1 不同处理下土壤微生物群落组成及花瓣图 Fig. 1 Soil microbial community composition and flower plot under different treatments 细菌相对值:Proteobacteria变形杆菌;Actinobacteria放线菌门;Chloroflexi绿弯菌门;Acidobacteria酸杆菌门;Gemmatimonadetes芽单胞菌门;Firmicutes厚壁菌门;Bacteroidetes拟杆菌门;Planctomycetes浮霉菌门;Nitrospirae硝化螺旋菌门;真菌相对值:Ascomycota子囊菌门;Basidiomycota担子菌门;Mortierellomycota被孢霉门;unclassified_k__Fungi未分类_k__真菌 |
20-40 cm土层中, FL1处理的Rokubacteria与FL2和CK处理差异显著(P < 0.05), FL2的变形杆菌门与FL1处理差异显著(P < 0.05)。在真菌在不同土层各样本中, 对丰度小于1%的物种归类为others。0-20 cm土层中, FL1处理的unclassified_k__Fungi与FL2处理差异显著(P < 0.05)。20-40 cm土层中, FL1处理的子囊菌门与FL2处理差异显著(P < 0.05)(图 1)。
在这些OTUs的基础上, 进一步分析了物种丰富度指数和多样性指数(表 2)。总的来说, 在0-20 cm土层中, FL2与CK处理的细菌Ace、Chao和香农指数差异显著(P < 0.05);FL1与CK处理的真菌Ace指数、Chao指数和辛普森指数差异显著(P < 0.05), FL1与FL2处理间Ace指数、Chao指数、香农指数和辛普森指数差异显著(P < 0.05)。20-40 cm土层中, FL1与FL2处理的细菌香农指数差异显著(P < 0.05), FL2与CK、FL1处理的细菌辛普森指数差异显著(P < 0.05);FL1与CK处理间真菌辛普森指数差异显著(P < 0.05)。
| 处理 Treatment |
丰富度指数Richness estimator | 多样性指数Diversity index | ||||
| Ace指数 Ace index |
Chao指数 Chao index |
Shannon指数 Shannon index H′ |
Simpson指数 Simpson index D′ |
|||
| 细菌Bacteria | CK-1 | 3679±237b | 3691±305b | 6.52±0.03b | 0.37±0.00a | |
| FL1-1 | 3786±107ab | 3781±70ab | 6.57±0.09ab | 0.36±0.00a | ||
| FL2-1 | 4077±76a | 4127±119a | 6.67±0.02a | 0.30±0.00a | ||
| CK-2 | 4245±217a | 4230±160a | 6.72±0.05ab | 0.32±0.00a | ||
| FL1-2 | 4309±207a | 4292±234a | 6.67±0.09b | 0.33±0.00a | ||
| FL2-2 | 4282±60a | 4256±43a | 6.81±0.04a | 0.27±0.00b | ||
| 真菌Fungi | CK-1 | 400±1b | 417±48b | 3.97±0.15ab | 3.68±0.49c | |
| FL1-1 | 479±35a | 496±25a | 4.26±0.20a | 8.40±1.72a | ||
| FL2-1 | 423±23b | 437±6b | 3.78±0.04b | 5.88±0.53b | ||
| CK-2 | 390±5a | 353±74a | 3.80±0.19a | 4.44±0.47b | ||
| FL1-2 | 404±53a | 379±77a | 3.75±0.27a | 8.63±1.49a | ||
| FL2-2 | 374±62a | 374±65a | 3.92±0.05a | 5.98±1.22b | ||
本研究中以培养96 h获得的数据对土壤微生物功能多样性指数进行分析(表 3)。0-20 cm土层中, FL1处理的香农指数与FL2处理相比差异显著(P < 0.05), 麦金托什指数中, FL1处理与FL2、CK处理相比差异显著(P < 0.05)。20-40 cm土层中, FL2处理香农指数与FL1、CK处理相比差异显著(P <0.05), 辛普森指数FL2与CK处理之间差异显著(P <0.05)。
| 处理 Treatment |
Shannon指数 Shannon index (H) |
麦金托什指数 McIntosh index (U) |
Simpson指数 Simpson index (D) |
处理 Treatment |
Shannon指数 Shannon index (H) |
麦金托什指数 McIntosh index (U) |
Simpson指数 Simpson index (D) |
|
| CK-1 | 2.72±0.00b | 3.27±0.15b | 0.93±0.01a | CK-2 | 2.81±0.10b | 3.55±0.65a | 0.93±0.01b | |
| FL1-1 | 2.92±0.10a | 3.92±0.20a | 0.93±0.02ab | FL1-2 | 2.89±0.00b | 3.96±0.35a | 0.93±0.01ab | |
| FL2-1 | 2.81±0.00ab | 3.37±0.25b | 0.90±0.01b | FL2-2 | 3.06±0.06a | 3.92±0.55a | 0.95±0.00a |
对不同土层各处理间土壤微生物群落的碳源利用情况进行分析时发现, 从土壤微生物群落利用百分比来看, 以利用羧酸类、聚合物类、氨基酸类以及糖类为主, 胺类和双亲化合物类的利用较低(图 2)。在0-20 cm土层中, FL1、FL2和CK处理利用羧酸类碳源差异显著(P < 0.05)。在20-40 cm土层中, FL1与FL2处理对胺类碳源的利用差异显著(P < 0.05)(图 2)。

|
| 图 2 不同处理下土壤微生物对6种碳源的相对利用 Fig. 2 The relative utilization of soil microorganisms to 6 carbon sources under different treatments 不同小写表示不同处理间差异显著(P<0.05) |
主成分分析结果表明, PC1和PC2的贡献率分别为27.7%和16.6%。影响PC1的碳源主要有糖类、氨基酸类、羧酸类和聚合物类;影响PC2的碳源主要有羧酸类和糖类。0-20 cm土层中, FL1和CK处理位于PC2正端, 受PC2上的碳源影响较大, 相似度较高, 碳源利用差异较小;FL2处理在第三象限, 与FL1和CK距离过大, 碳源利用差异较大(图 3)。20-40 cm土层中, CK位于第四象限, 且FL1、FL2和CK处理相对分离, 说明各处理间碳源利用有差异(图 3)。

|
| 图 3 不同处理下土壤微生物群落碳源利用主成分分析 Fig. 3 Principal component analysis of carbon source utilization of soil microbial communities under different treatments |
使用Tax4Fun进行潜在的微生物KEGG功能预测, 代谢作用和环境信息处理是主要一级代谢通路, 平均占比分别为63.67%和17.29%(图 4)。在二级代谢通路上总共注释了41个特征, 图 5显示了前15个细菌的相对功能基团丰度。在0-20 cm和20-40 cm土层中, FL1、FL2和CK处理的主要细菌功能基团的功能注释为:氨基酸代谢、碳水化合物代谢、膜运输、信号传递、萜类化合物和聚酮化合物的代谢、能量代谢(图 4)。FL1、FL2和CK处理在0-20 cm土层的辅因子和维生素的代谢差异显著(P < 0.05)。在20-40 cm土层中, FL1、FL2的碳水化合物代谢显著高于CK处理(P < 0.05), FL1的氨基酸代谢、辅因子和维生素的代谢和膜运输显著高于CK处理(P < 0.05), FL2的信号传递、能量代谢显著高于CK处理(P < 0.05)。

|
| 图 4 不同处理下不同土层主要土壤细菌的功能结构(二级代谢通路) Fig. 4 Functional structure of main soil bacteria in different soil layers under different treatments (Pathway level 2) |

|
| 图 5 不同处理下不同土层主要土壤真菌的功能结构(二级代谢通路) Fig. 5 Functional structure of main soil fungi in different soil layers under different treatments (Pathway level 2) |
使用FUNGuild对真菌进行功能Guild注释(图 5)。按真菌的营养类型主要分为9种功能类型:病原-腐生-共生菌、腐生菌、病菌、腐生-共生菌、病原-腐生-共生菌、病菌-腐生菌、共生菌和病菌-共生菌。在0-20 cm土层中, FL1的病菌-腐生菌、病菌-共生菌、腐生菌和腐生-共生菌显著高于CK(P < 0.05), FL2的共生菌和腐生-共生菌显著高于CK(P < 0.05)。在20-40 cm土层中, FL1的病菌-腐生菌-共生菌显著高于CK(P < 0.05), FL2的病菌-腐生菌和共生菌显著高于CK(P < 0.05)(图 5)。
2.5 影响微生物功能多样性的生态因素及相关性分析结构方程模型(SEM)检测控制因素对土壤微生物功能多样性有直接和间接的影响(图 6)。耕作方式的直接效应能解释土壤细菌群落多样性总方差的72%(图 6)。土壤细菌群落多样性能够解释土壤真菌群落多样性总方差的11%(图 6)。耕作方式和土壤微生物(细菌、真菌)群落多样性能解释土壤酶活性总方差的96% (图 6)。耕作方式、土壤微生物群落多样性和土壤酶活性能解释土壤微生物功能多样性总方差的69% (图 6)。

|
| 图 6 耕作方法、土壤酶活性、细菌和真菌多样性指数和土壤微生物功能多样性的结构方程模型(SEM) Fig. 6 Structural equation model (SEM) of farming methods, soil enzyme activity, bacterial and fungal diversity index and soil microbial functional diversity 箭头的粗细表示标准化路径系数的大小; R2值代表每个内生变量解释方差的比例;GFI:拟合优度指数;RMSEA:近似的均方根误差; *P < 0.05, **P < 0.01, ***P < 0.001 |
如表 4所示, 玉米产量与土壤脲酶、转化酶、土壤微生物香农指数(H)、麦金托什指数(U)和细菌群落香农指数(H′)显著正相关(P < 0.05); 三种土壤酶(Ure、Inv、Alp)均与土壤微生物辛普森指数(D)、细菌群落多样性指数(Ace、Chao、H′、D′)和真菌群落多样性指数(Ace、Chao)显著正相关(P < 0.05);土壤微生物香农指数(H)与细菌群落多样性指数(Ace、Chao、H′、D′)和真菌群落多样性指数(H′、D′)显著正相关(P < 0.05)。
| Ure | Inv | Alk | H | U | D | 细菌Bacteria | ||||
| Ace | Chao | H′ | D′ | |||||||
| 产量Yield | 0.498* | 0.501* | 0.270 | 0.600** | 0.469* | 0.113 | 0.222 | 0.220 | 0.547* | 0.021 |
| Ure | 0.870** | 0.893** | 0.299 | -0.034 | 0.716** | 0.654** | 0.614** | 0.731** | 0.740** | |
| Inv | 0.912** | 0.216 | -0.204 | 0.862** | 0.790** | 0.734** | 0.744** | 0.945** | ||
| Alk | 0.199 | -0.087 | 0.753** | 0.695** | 0.662** | 0.671** | 0.817** | |||
| H | 0.177 | -0.179 | 0.665** | 0.620** | 0.663** | 0.863** | ||||
| U | 0.368 | 0.232 | 0.158 | 0.284 | 0.319 | |||||
| D | 0.161 | 0.210 | 0.126 | -0.191 | ||||||
| 真菌Fungi | ||||||||||
| Ace | Chao | H′ | D′ | |||||||
| 产量Yield | 0.113 | 0.0782 | -0.074 | 0.356 | ||||||
| Ure | 0.759** | 0.781** | 0.063 | 0.036 | ||||||
| Inv | 0.706** | 0.731** | 0.088 | -0.031 | ||||||
| Alk | 0.840** | 0.857** | 0.081 | -0.050 | ||||||
| H | 0.053 | 0.024 | 0.678** | 0.664** | ||||||
| U | -0.050 | -0.048 | -0.143 | 0.240 | ||||||
| D | -0.334 | 0.331 | -0.113 | 0.213 | ||||||
| Ure: 脲酶Urease; Inv: 转化酶Invertase; Alk: 碱性磷酸酶Alkaline phosphatase; H: Shannon指数Shannon index; U: McIntosh指数McIntosh index; D: 优势度指数Simpson index; Ace: Ace指数Ace index; Chao: Chao指数Chao index; H′: Shannon指数Shannon index; D′: Simpson指数Simpson index; * P<0.05; ** P<0.01 | ||||||||||
土壤微生物群落活动和组成受到农业耕作方式影响[33-35]。粉垄耕作较常规耕作有效提高土壤微生物数量, 使土壤中的氨化细菌、固氮菌、无机磷细菌和钾细菌的数量高于旋耕耕作处理和深翻耕作处理[24]。杨博[36]研究发现, 粉垄耕作为深层土壤微生物的生存提供了有利条件, 0-20 cm和20-40 cm土层土壤微生物总菌落数较传统耕作显著提升。本研究中, 细菌的OTUs数量明显高于真菌的OTUs, 表明土壤细菌占土壤微生物总数量的比例较大。本研究发现, 粉垄耕作与传统耕作相比改变了细菌的多样性和群落组成(图 1, 表 2)。
造成的原因可能是, 粉垄耕作提高了耕地土壤耕层碳存储, 降低了土壤紧实度, 促进了土壤大团聚体的形成[37, 38]。土壤团聚结构的动态变化又反馈控制土壤微生物活动和土壤有机质组分的分异作用, 使土壤有机质组分-土壤团聚结构-微生物群落结构的动态变化之间存在耦合作用[39], 从而改变了细菌群落结构和多样性。此外, 由于细菌提供信号分子或将介质化合物转化为真菌消耗的物质促进了真菌的生长, 提高了真菌的定殖率, 甚至促进真菌的完整生命周期[40-42]。这些结果表明, 真菌群落间接地受到了粉垄耕作方式的影响, 导致真菌群落结构、物种丰富度指数和多样性指数较传统耕作有所提升(图 1, 表 2)。粉垄耕作提升了耕地土壤的微生物群落多样性, 使参与土壤养分转化的功能型微生物增加, 为维系耕地土壤微生态环境的稳定发挥了重要作用。
3.2 控制因素对土壤酶活性的影响研究发现, 耕作方式和土壤酶活性显著正相关(图 6)。土壤中的氮、磷转化以及碳水化合物的代谢和运输贮藏与脲酶、转化酶和碱性磷酸酶密切相关[43, 44]。作物根系可以产生酶进行养分吸收[45], 粉垄耕作通过物理方式改善作物土壤环境, 使作物根系具有活力, 向下伸展[46], 释放出更多的酶来加强与土壤微生境养分的互换[24], 使粉垄耕作比传统耕作具有更高的土壤酶活性。土壤中的酶不仅来源于作物根系, 还来源于土壤微生物的代谢活动[47]。异养型细菌为了自身发展, 会释放出相应功能的酶, 来参与土壤养分的循环[48]。王丹等[49]研究发现, 放线菌门(Actinobacteriota)可以降解土壤中各种不溶性有机物质以供细胞代谢所需的各种营养, 与水解酶的活性有显著相关。拟杆菌门(Bacteroidota)是具有溶磷作用的富营养菌, 参与养分代谢, 与脲酶、转化酶和碱性磷酸酶呈正相关[50]。真菌群落的发展使养分循环加快, 能量代谢有关的真菌和参与养分循环土壤酶高度相关[51], 也进一步证明土壤酶活性的高低受土壤真菌的影响。传统耕作不利于作物表层根系发育和土壤微生物活动[52], 导致土壤中酶活性低。粉垄耕作通过改善耕地土壤物理结构, 促进了作物根系的生长和土壤微生物自身发展, 并释放出相应的酶, 加快了土壤养分的循环, 使粉垄耕作下的土壤酶活性高于传统耕作(表 1), 并且脲酶和转化酶活性与产量显著正相关(表 4), 粉垄耕作处理的玉米产量得到提升。
3.3 控制因素对功能多样性的影响已有研究表明, 根系的生长可以释放酶和碳源[53], 从而影响分解碳源的微生物, 有利于土壤微生物多样性的增加[54, 55]。粉垄耕作与传统耕作之间微生物群落结构存在差异, 粉垄耕作深度35 cm显著提升了0-20 cm土层的功能多样性, 粉垄耕作深度50 cm显著提升了20-40 cm土层微生物功能多样性。造成的原因可能是, 粉垄耕作可以促进根系生长, 使根系释放出更多的碳源, 从而提高了微生物功能多样性。有研究表明, 患病根系增加了羧酸类和胺类碳源的释放, 从而提升了土壤微生物对羧酸类和胺类碳源的利用[56, 57]。本研究得出, 在0-20 cm土层中粉垄耕作对羧酸类碳源的利用率显著降低;在20-40 cm土层中, 粉垄耕作深度35 cm和粉垄耕作深度50 cm相比显著降低了对胺类碳源的利用比例。说明粉垄耕作可以减少作物根系患病的几率, 患病根系向周围分泌羧酸类和胺类碳源减弱, 使微生物利用碳源的能力发生改变。
土壤微生物细菌和真菌群落多样性可以直接影响微生物功能多样性(图 6)。已有研究表明, 土壤中微生物群落结构和碳源代谢紧密相关[58]。土壤中拥有代谢、遗传信息处理、有机系统3类功能基因的共养生物(copiotrophs、R-strategists)和寡养生物(oligotrophs、K-strategists)相互作用, 促进了细菌群落从土壤中吸收营养物质和溶解铁和小分子等来加速自身生长, 使土壤细菌代谢旺盛, 提高了细菌功能群落多样性[59-61]。真菌功能预测的腐生营养型为最主要的营养型, 这可能与子囊菌门为优势菌门有关。子囊菌门大多为腐生菌, 是土壤中重要的分解者[62], 可以分解难降解的有机质, 在养分循环方面起着重要作用[63]。另外, 含有共生营养型的丛枝菌根真菌(AMF)与宿主植物建立共生关系后, 不仅显著影响植物生长, 还能引起根系分泌物的变化[64, 65]。AMF与植物共生可以影响土壤细菌群落结构和功能多样性[66]。本研究得出, 粉垄耕作下细菌功能预测的一级代谢通路代谢作用和环境信息处理比传统耕作显著增加(图 4), 真菌的功能预测出的营养类型也比传统耕作显著提高(图 5)。原因可能是, 粉垄耕作可以增加了细菌的功能基团, 细菌增强了养分的摄取能力, 功能代谢得到提升。真菌不同营养类型的功能基团增加, 提升了自己参与碳源代谢的能力。真菌不仅可以分解出细菌利用的碳源, 而且间接促进根系向土壤释放碳源[67]。细菌真菌相互作用, 使土壤中代谢底物变得丰富多样, 土壤微生物功能多样性提升。此外, 粉垄耕作下的病菌-腐生菌、病菌-共生菌、腐生菌、腐生-共生菌和病菌-腐生菌-共生菌等功能基团不同程度下均高于传统耕作处理, 可能的原因是, 粉垄耕作处理显著改变了真菌群落结构, 真菌数量增多, 导致相应功能的功能基团也增多。
土壤酶活性作为微生物活性的指示物, 与微生物功能多样性显著相关[68-71]。段益莉等[72]研究表明, 土壤酶活性的高低不仅可以反映出有机物质的水解程度, 还能反映出对碳源利用能力的高低。本研究也发现, 土壤酶活性显著影响了微生物功能多样性(图 6), 表明粉垄耕作可以通过提升酶活性间接的使土壤微生物功能多样性提高, 使土壤微生物群落利用养分的能力加强, 群落功能结构完善, 增加了土壤生态系统的稳定性, 提升了玉米的产量。
4 结论本文研究了粉垄耕作对玉米耕地土壤酶活性、微生物群落结构和功能代谢的影响, 研究表明:粉垄耕作可以显著改善土壤细菌、真菌群落结构, 增加参与养分代谢的功能基团, 提高土壤微生物群落多样性, 并增加了玉米产量;粉垄耕作提升了参与养分循环的土壤脲酶、转化酶和碱性磷酸酶的活性;粉垄耕作技术促进了土壤微生物养分代谢的能力, 使土壤微生物总体的功能多样性得到提升;从土壤微生物群落多样性、功能多样性、酶活性和玉米产量可以看出, 粉垄耕作深度35 cm处理优于粉垄耕作深度50 cm。总的来说, 从土壤微生态的角度可以初步得出粉垄耕作技术可以促进了养分循环, 使土壤耕地质量得到提升, 增加了农作物的产量。本研究为粉垄耕作技术在中国西部干旱半干旱地区作物栽培的应用发展提供了科学依据。
耕作方式对耕地质量的影响是一个周期较长的过程。因此, 随着长期粉垄耕作的使用, 耕地物理结构和土壤微生态也会发生着变化。本团队仅研究了一年的粉垄耕作不同深度对耕地土壤酶活性、微生物群落结构和功能多样性的影响, 未来会在此试验田进行长期工作, 今后会对土壤微生态的变化进行长期检测和跟踪研究, 并进一步增加关于土壤团聚体、土壤碳含量以及土壤病害等研究内容。
| [1] |
Waldrop M P, Balser T C, Firestone M K. Linking microbial community composition to function in a tropical soil. Soil Biology and Biochemistry, 2000, 32(13): 1837-1846. DOI:10.1016/S0038-0717(00)00157-7 |
| [2] |
Hinsinger P, Bengough A G, Vetterlein D, Young I M. Rhizosphere: biophysics, biogeochemistry and ecological relevance. Plant and Soil, 2009, 321(1/2): 117-152. |
| [3] |
王理德, 姚拓, 王方琳, 魏林源, 郭春秀, 吴春荣, 李发明. 石羊河下游退耕地土壤微生物变化及土壤酶活性. 生态学报, 2016, 36(15): 4769-4779. |
| [4] |
刘红梅, 张贵龙, 皇甫超河, 杨殿林, 赵建宁. 不同耕作方式对农田土壤微生物功能多样性的影响. 农业科学, 2016, 6(1): 1-9. |
| [5] |
Gu Y F, Zhang X P, Tu S H, Lindström K. Soil microbial biomass, crop yields, and bacterial community structure as affected by long-term fertilizer treatments under wheat-rice cropping. European Journal of Soil Biology, 2009, 45(3): 239-246. DOI:10.1016/j.ejsobi.2009.02.005 |
| [6] |
Kaschuk G, Alberton O, Hungria M. Three decades of soil microbial biomass studies in Brazilian ecosystems: lessons learned about soil quality and indications for improving sustainability. Soil Biology and Biochemistry, 2010, 42(1): 1-13. DOI:10.1016/j.soilbio.2009.08.020 |
| [7] |
Jat H S, Choudhary M, Datta A, Yadav A K, Meena M D, Devi R, Gathala M K, Jat M L, McDonald A, Sharma P C. Temporal changes in soil microbial properties and nutrient dynamics under climate smart agriculture practices. Soil and Tillage Research, 2020, 199: 104595. DOI:10.1016/j.still.2020.104595 |
| [8] |
Essel E, Xie J H, Deng C C, Peng Z K, Wang J B, Shen J C, Xie J H, Coulter J A, Li L L. Bacterial and fungal diversity in rhizosphere and bulk soil under different long-term tillage and cereal/legume rotation. Soil and Tillage Research, 2019, 194: 104302. DOI:10.1016/j.still.2019.104302 |
| [9] |
Wang Z T, Li Y Z, Li T, Zhao D Q, Liao Y C. Tillage practices with different soil disturbance shape the rhizosphere bacterial community throughout crop growth. Soil and Tillage Research, 2020, 197: 104501. DOI:10.1016/j.still.2019.104501 |
| [10] |
Yang W H, Zhang X T, Wu L Q, Rensing C, Xing S H. Short-term application of magnesium fertilizer affected soil microbial biomass, activity, and community structure. Journal of Soil Science and Plant Nutrition, 2021, 21(1): 675-689. DOI:10.1007/s42729-020-00392-x |
| [11] |
Licht M A, Parvej M R, Wright E E. Corn yield response to row spacing and plant population in Iowa. Crop, Forage & Turfgrass Management, 2019, 5(1): 1-7. |
| [12] |
Afzalinia S, Khosravani A, Javadi A, Mohammadi D, Alavimanesh S M. Effect of tillage and planting methods on the soil properties, grain drill performance, and wheat yield. Journal of Agricultural Science and Technology A, 2012, 2(4): 537-543. |
| [13] |
Anand S R, Rajkumar R H, Vishwanatha J, Karegoudar A V. Evaluation of different alternative crops for saline vertisols under different tillage methods in rice fallows in tungabhadra command area of Karnataka. Journal of Progressive Agriculture, 2018, 9(2): 18-22. |
| [14] |
Zhai L C, Xu P, Zhang Z B, Li S K, Xie R Z, Zhai L F, Wei B H. Effects of deep vertical rotary tillage on dry matter accumulation and grain yield of summer maize in the Huang-Huai-Hai Plain of China. Soil and Tillage Research, 2017, 170: 167-174. DOI:10.1016/j.still.2017.03.013 |
| [15] |
Montgomery D R. Soil erosion and agricultural sustainability. Proceedings of the National Academy of Sciences of the United States of America, 2007, 104(33): 13268-13272. DOI:10.1073/pnas.0611508104 |
| [16] |
Choudhury S G, Srivastava S, Singh R, Chaudhari S K, Sharma D K, Singh S K, Sarkar D. Tillage and residue management effects on soil aggregation, organic carbon dynamics and yield attribute in rice-wheat cropping system under reclaimed sodic soil. Soil and Tillage Research, 2014, 136: 76-83. DOI:10.1016/j.still.2013.10.001 |
| [17] |
Zhang Y J, Wang S L, Wang H, Ning F, Zhang Y H, Dong Z Y, Wen P F, Wang R, Wang X L, Li J. The effects of rotating conservation tillage with conventional tillage on soil properties and grain yields in winter wheat-spring maize rotations. Agricultural and Forest Meteorology, 2018, 263: 107-117. DOI:10.1016/j.agrformet.2018.08.012 |
| [18] |
Li X Z, Wei B H, Xu X L, Zhou J. Effect of deep vertical rotary tillage on soil properties and sugarcane biomass in rainfed dry-land regions of southern China. Sustainability, 2020, 12(23): 10199. DOI:10.3390/su122310199 |
| [19] |
Wu F, Zhai L C, Xu P, Zhang Z B, Baillo E H, Tolosa L N, Kimotho R N, Jia X L, Guo H Q. Effects of deep vertical rotary tillage on the grain yield and resource use efficiency of winter wheat in the Huang-Huai-Hai Plain of China. Journal of Integrative Agriculture, 2021, 20(2): 593-605. DOI:10.1016/S2095-3119(20)63405-0 |
| [20] |
Wei B H. Fenlong cultivation-the fourth set of farming methods invented in China. Agricultural Science & Technology, 2017, 18(11): 2045-2048, 2052-2052. |
| [21] |
李浩, 黄金玲, 李志刚, 韦本辉, 陈晓茹, 韩世健, 梁晓莹, 李素丽. 粉垄耕作提高土壤养分有效性并促进甘蔗维管组织发育和养分吸收. 植物营养与肥料学报, 2021, 27(2): 204-214. |
| [22] |
郑佳舜, 胡钧铭, 韦翔华, 韦燕燕, 苏世鸣, 李婷婷, 夏旭, 俞月凤, 张俊辉. 绿肥压青对粉垄稻田土壤微生物量碳和有机碳累积矿化量的影响. 中国生态农业学报(中英文), 2021, 29(4): 691-703. |
| [23] |
周佳, 周灵芝, 劳承英, 申章佑, 李艳英, 胡泊, 黄渝岚, 韦本辉. 短期不同耕作方式对水稻根际土壤细菌群落结构多样性的影响. 南方农业学报, 2020, 51(10): 2401-2411. DOI:10.3969/j.issn.2095-1191.2020.10.011 |
| [24] |
黎佐生, 蒋代华, 韦本辉. 粉垄耕作对宿根蔗地根际微生物及酶活性的影响. 新农业, 2020(7): 45-47. |
| [25] |
关松荫. 土壤酶及其研究法. 北京: 农业出版社, 1986, 274-338. |
| [26] |
Hadwin A K M, Del Rio L F, Pinto L J, Painter M, Routledge R, Moore M M. Microbial communities in wetlands of the Athabasca oil sands: genetic and metabolic characterization. FEMS Microbiology Ecology, 2006, 55(1): 68-78. DOI:10.1111/j.1574-6941.2005.00009.x |
| [27] |
Yin C Y, Yan Z Z. Variations of soil bacterial diversity and metabolic function with tidal flat elevation gradient in an artificial mangrove wetland. Science of the Total Environment, 2020, 718: 137385. DOI:10.1016/j.scitotenv.2020.137385 |
| [28] |
Chen S F, Zhou Y Q, Chen Y R, Gu J. Fastp: an ultra-fast all-in-one FASTQ preprocessor. Bioinformatics, 2018, 34(17): i884-i890. DOI:10.1093/bioinformatics/bty560 |
| [29] |
Magoč T, Salzberg S L. FLASH: fast length adjustment of short reads to improve genome assemblies. Bioinformatics, 2011, 27(21): 2957-2963. DOI:10.1093/bioinformatics/btr507 |
| [30] |
Stackebrand E, Goebel B M. Taxonomic note: a place for DNA-DNA reassociation and 16S rRNA sequence analysis in the present species definition in Bacteriology. International Journal of Systematic and Evolutionary Microblogy, 1994, 44(4): 846-849. DOI:10.1099/00207713-44-4-846 |
| [31] |
Edgar R C. UPARSE: highly accurate OTU sequences from microbial amplicon reads. Nature Methods, 2013, 10(10): 996-998. DOI:10.1038/nmeth.2604 |
| [32] |
Wang Q, Garrity G M, Tiedje J M, Cole J R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Applied and Environmental Microbiology, 2007, 73(16): 5261-5267. DOI:10.1128/AEM.00062-07 |
| [33] |
Bardgett R D, Jones A C, Jones D L, Kemmitt S J, Cook R, Hobbs P J. Soil microbial community patterns related to the history and intensity of grazing in sub-montane ecosystems. Soil Biology and Biochemistry, 2001, 33(12/13): 1653-1664. |
| [34] |
Nannipieri P, Ascher J, Ceccherini M T, Landi L, Pietramellara G, Renella G. Microbial diversity and soil functions. European Journal of Soil Science, 2003, 54(4): 655-670. DOI:10.1046/j.1351-0754.2003.0556.x |
| [35] |
Griffiths B S, Philippot L. Insights into the resistance and resilience of the soil microbial community. FEMS Microbiology Reviews, 2013, 37(2): 112-129. DOI:10.1111/j.1574-6976.2012.00343.x |
| [36] |
杨博. 粉垄耕作对盐碱地土壤性质及耐盐碱高粱生长的影响研究[D]. 呼和浩特: 内蒙古农业大学, 2020.
|
| [37] |
陈仕林, 胡钧铭, 黄忠华, 李婷婷, 郑佳舜, 黄俞铭, 罗维钢, 何铁光, 韦翔华. 粉垄耕作对平地和坡耕地蔗田土壤有机碳矿化和结构的影响. 中国农业气象, 2020, 41(5): 299-307. DOI:10.3969/j.issn.1000-6362.2020.05.004 |
| [38] |
王世佳, 蒋代华, 朱文国, 张蓉蓉, 李军伟, 韦本辉. 粉垄耕作对农田赤红壤团聚体结构的影响. 土壤学报, 2020, 57(2): 326-335. |
| [39] |
Chenu C, Sotzky G. Interactions between microorganisms and soil particles: an overview//Huang P M, Bollag J M, Senesi N, eds. Interactions Between Soil Particles and Microorganisms. New York: John Wiley and Sons Inc., 2002: 3-40.
|
| [40] |
Hildebrandt U, Ouziad F, Marner F J, Bothe H. The bacterium Paenibacillus validus stimulates growth of the arbuscular mycorrhizal fungus Glomus intraradices up to the formation of fertile spores. FEMS Microbiology Letters, 2006, 254(2): 258-267. DOI:10.1111/j.1574-6968.2005.00027.x |
| [41] |
Visen A, Bohra M, Singh P N, Srivastava P C, Kumar S, Sharma A K, Chakraborty B. Two pseudomonad strains facilitate AMF mycorrhization of litchi (Litchi chinensis Sonn.) and improving phosphorus uptake. Rhizosphere, 2017, 3: 196-202. DOI:10.1016/j.rhisph.2017.04.006 |
| [42] |
Deveau A, Palin B, Delaruelle C, Peter M, Kohler A, Pierrat J C, Sarniguet A, Garbaye J, Martin F, Frey-Klett P. The mycorrhiza helper Pseudomonas fluorescens BBc6R8 has a specific priming effect on the growth, morphology and gene expression of the ectomycorrhizal fungus Laccaria bicolor S238N. New Phytologist, 2007, 175(4): 743-755. DOI:10.1111/j.1469-8137.2007.02148.x |
| [43] |
Qin X, Liu Y T, Huang Q Q, Zhao L J, Xu Y M. Effects of sepiolite and biochar on enzyme activity of soil contaminated by Cd and Atrazine. Bulletin of Environmental Contamination and Toxicology, 2020, 104(5): 642-648. DOI:10.1007/s00128-020-02833-w |
| [44] |
Singh D X, Kumar S. Nitrate reductase, arginine deaminase, urease and dehydrogenase activities in natural soil (ridges with forest) and in cotton soil after acetamiprid treatments. Chemosphere, 2008, 71(3): 412-418. DOI:10.1016/j.chemosphere.2007.11.005 |
| [45] |
Tang M Y, Wang Q, Chen L, Zhang X L, Zhang Z Q, Lü R H, Liang T F. On the growth and physiological characteristics of rice under smashing ridge tillage. Agricultural Science & Technology, 2016, 17(2): 334-337. |
| [46] |
胡钧铭, 陈胜男, 韦翔华, 夏旭, 韦本辉. 耕作对健康耕层结构的影响及发展趋势. 农业资源与环境学报, 2018, 35(2): 95-103. |
| [47] |
战秀梅, 李秀龙, 韩晓日, 李亭亭, 杨劲峰, 刘小虎. 深耕及秸秆还田对春玉米产量、花后碳氮积累及根系特征的影响. 沈阳农业大学学报, 2012, 43(4): 461-466. DOI:10.3969/j.issn.1000-1700.2012.04.013 |
| [48] |
王文晓, 李小伟, 黄文广, 杨君珑. 蒙古沙冬青根际土壤细菌群落组成及多样性与生态因子相关性研究. 生态学报, 2020, 40(23): 8660-8671. |
| [49] |
王丹, 赵亚光, 马蕊, 杨鹏, 张成, 周东姣, 孙福新, 张凤华. 微生物菌肥对盐碱地枸杞土壤改良及细菌群落的影响. 农业生物技术学报, 2020, 28(8): 1499-1510. |
| [50] |
Fierer N, Lauber C L, Ramirez K S, Zaneveld J, Bradford M A, Knight R. Comparative metagenomic, phylogenetic and physiological analyses of soil microbial communities across nitrogen gradients. The ISME Journal, 2012, 6(5): 1007-1017. DOI:10.1038/ismej.2011.159 |
| [51] |
李鹏飞, 张兴昌, 郝明德, 崔勇兴, 张燕江, 朱世雷. 植被恢复对黄土高原矿区重构土壤理化性质、酶活性以及真菌群落的影响. 水土保持通报, 2019, 39(5): 1-7. |
| [52] |
邓小华, 王新月, 杨红武, 刘勇军, 邓永晟, 周米良, 张明发, 赵炯平, 李奇, 王卫民, 陈金, 粟戈璇. 粉垄耕作深度对烤烟生长和物质积累及烟叶产质量的影响. 中国烟草科学, 2020, 41(5): 28-35. |
| [53] |
李骁, 王迎春. 土壤微生物多样性与植物多样性. 内蒙古大学学报: 自然科学版, 2006, 37(6): 708-713. DOI:10.3969/j.issn.1000-1638.2006.06.024 |
| [54] |
Lorenz K, Preston C M, Krumrei S, Feger K H. Decomposition of needle/leaf litter from scots pine, black cherry, common oak and European beech at a conurbation forest site. European Journal of Forest Research, 2004, 123(3): 177-188. DOI:10.1007/s10342-004-0025-7 |
| [55] |
Pengthamkeerati P, Motavalli P P, Kremer R J. Soil microbial activity and functional diversity changed by compaction, poultry litter and cropping in a claypan soil. Applied Soil Ecology, 2011, 48(1): 71-80. DOI:10.1016/j.apsoil.2011.01.005 |
| [56] |
张丽娟, 茆军, 张志东, 谢玉清. 新疆大蒜根腐型病害根际土壤微生物群落多样性初探. 新疆农业科学, 2013, 50(11): 2109-2117. |
| [57] |
吴照祥, 郝志鹏, 陈永亮, 曾燕, 郭兰萍, 黄璐琦, 王勇, 陈保冬. 三七根腐病株根际土壤真菌群落组成与碳源利用特征研究. 菌物学报, 2015, 34(1): 65-74. |
| [58] |
Kaiser K, Wemheuer B, Korolkow V, Wemheuer F, Nacke H, Schöning I, Schrumpf M, Daniel R. Driving forces of soil bacterial community structure, diversity, and function in temperate grasslands and forests. Scientific Reports, 2016, 6: 33696. DOI:10.1038/srep33696 |
| [59] |
Wang Y J, Liu L, Tian Y L, Wu X P, Yang J F, Luo Y, Li H K, Awasthi M K, Zhao Z Y. Temporal and spatial variation of soil microorganisms and nutrient under white clover cover. Soil and Tillage Research, 2020, 202: 104666. DOI:10.1016/j.still.2020.104666 |
| [60] |
Wu Z X, Hao Z P, Sun Y Q, Guo L P, Huang L Q, Zeng Y, Wang Y, Yang L, Chen B D. Comparison on the structure and function of the rhizosphere microbial community between healthy and root-rot Panax notoginseng. Applied Soil Ecology, 2016, 107: 99-107. DOI:10.1016/j.apsoil.2016.05.017 |
| [61] |
Tolli J, King G M. Diversity and structure of bacterial chemolithotrophic communities in pine forest and agroecosystem soils. Applied and Environmental Microbiology, 2005, 71(12): 8411-8418. DOI:10.1128/AEM.71.12.8411-8418.2005 |
| [62] |
任玉连, 范方喜, 彭淑娴, 陆梅. 纳帕海沼泽化草甸不同季节土壤真菌群落结构与理化性质的关系. 中国农学通报, 2018, 34(29): 69-75. DOI:10.11924/j.issn.1000-6850.casb17080110 |
| [63] |
Beimforde C, Feldberg K, Nylinder S, Rikkinen J, Tuovila H, Dörfelt H, Gube M, Jackson D J, Reitner J, Seyfullah L J, Schmidt A R. Estimating the Phanerozoic history of the Ascomycota lineages: combining fossil and molecular data. Molecular Phylogenetics and Evolution, 2014, 78: 386-398. DOI:10.1016/j.ympev.2014.04.024 |
| [64] |
Igiehon N O, Babalola O O. Biofertilizers and sustainable agriculture: exploring arbuscular mycorrhizal fungi. Applied Microbiology and Biotechnology, 2017, 101(12): 4871-4881. DOI:10.1007/s00253-017-8344-z |
| [65] |
吴金平, 顾玉成. 丛枝菌根真菌(AMF)在土壤修复中的生态应用. 中国农学通报, 2009, 25(7): 243-245. |
| [66] |
屈明华, 俞元春, 李生, 张金池. 丛枝菌根真菌对矿质养分活化作用研究进展. 浙江农林大学学报, 2019, 36(2): 394-405. |
| [67] |
Toljander J F, Lindahl B D, Paul L R, Elfstrand M, Finlay R D. Influence of arbuscular mycorrhizal mycelial exudates on soil bacterial growth and community structure. FEMS Microbiology Ecology, 2007, 61(2): 295-304. DOI:10.1111/j.1574-6941.2007.00337.x |
| [68] |
李东坡, 武志杰, 陈利军. 有机农业施肥方式对土壤微生物活性的影响研究. 中国生态农业学报, 2005, 13(2): 99-101. |
| [69] |
张逸飞, 钟文辉, 李忠佩, 蔡祖聪. 长期不同施肥处理对红壤水稻土酶活性及微生物群落功能多样性的影响. 生态与农村环境学报, 2006, 22(4): 39-44. DOI:10.3969/j.issn.1673-4831.2006.04.008 |
| [70] |
李忠佩, 吴晓晨, 陈碧云. 不同利用方式下土壤有机碳转化及微生物群落功能多样性变化. 中国农业科学, 2007, 40(8): 1712-1721. DOI:10.3321/j.issn:0578-1752.2007.08.017 |
| [71] |
刘淑娟, 张伟, 王克林, 舒世燕, 何寻阳, 杨珊, 潘复静. 桂西北喀斯特峰丛洼地不同植被演替阶段的土壤脲酶活性. 生态学报, 2011, 31(19): 5789-5796. |
| [72] |
段益莉, 李继侠, 江强, 赵鸿宇. 长白山东坡不同海拔落叶松土壤微生物碳代谢及酶活性研究. 生态环境学报, 2019, 28(4): 652-660. |