 2022, Vol. 42
2022, Vol. 42文章信息
- 杨大星, 杨娟, 李灿
- YANG Daxing, YANG Juan, LI Can
- 喀斯特洞穴小型土壤节肢动物多样性
- Diversity of cave soil microarthropod in karst region
- 生态学报. 2022, 42(12): 5089-5102
- Acta Ecologica Sinica. 2022, 42(12): 5089-5102
- http://dx.doi.org/10.5846/stxb202008122104
-
文章历史
- 收稿日期: 2020-08-12
- 网络出版日期: 2022-02-11





贵州是中国乃至世界喀斯特分布面积最大片区的中心, 发育着一套最典型、显著的喀斯特洞穴, 洞穴结构复杂、类型多样[1], 洞穴具有植被稀少, 有机质贫乏, 湿度高, 温度恒定等特征, 环境条件与常见的生态系统类型明显不同, 是研究喀斯特生态过程的理想场所[2], 贵州省因目前发现的岩溶洞穴最多也成为了洞穴生态系统研究的首选区域[3]。土壤节肢动物可通过取食作用和非取食作用发挥其生态功能, 进而改善土壤土壤结构、促进有机质分解和养分循环[4-7]。不仅如此, 土壤节肢动物还能指征环境变化, 其敏感性甚至优于植被和土壤理化性质对环境变化的反应[8-9]。所以研究洞穴土壤节肢动物, 对于深入认识喀斯特地下生态过程, 揭示土壤节肢动物在特殊环境下的响应机制具有重要意义。
近年来, 科研工作者在喀斯特地区陆续开展了一系列土壤动物生态学研究, 但主要以森林[10-13]、农田[14-16]和草地[17]等常见的生态系统类型为主, 洞穴少有涉及[18-20], 这并不利于全面认识喀斯特地区的生态过程。因此, 本研究采用生态学相关原理及方法对洞穴土壤节肢动物进行研究, 旨在弄清洞穴土壤节肢动物群落结构特征及其与环境因子的相互作用关系, 为喀斯特地区洞穴生物多样性保护提供科学资料。
1 材料与方法 1.1 研究区概况研究区位于贵州省南部的黔南州, 地理坐标介于106°12′—108°18′E, 25°04′—27°29′N之间。地势北高南低, 海拔在242—1961 m之间, 平均海拔高度997 m。气候属亚热带季风气候, 无霜期平均为292 d, 区内大部分地区日照度在30%左右, 年平均气温13.6—19.6 ℃, 温度自北向南、自西向东逐渐递增, 年降水多在1200 mm以上。区内共有各种植物170科, 744属, 2000余种, 主要树种有马尾松(Pinus massoniana)、杉木(Cunninghamia lanceolat)、栲树(Castanopsis fargesii)、水青冈(Fagus longipetiolata)、栓皮栎(Quercus variabilis)、光皮桦(Betula luminifera)和枫香(Liquidambar formosana)等。土壤类型主要有赤红壤土、红壤土、黄壤土、红色石和石灰土等。
1.2 研究方法 1.2.1 样地设置于2018年5月(春季)、7月(夏季)、10月(秋季)和2019年1月(冬季)在黔南州选取无干扰或干扰较小洞穴作为实验样地, 以自然林为对照进行小型土壤节肢动物研究。洞穴和自然林分别设置4个和3个重复样地, 同类型样地间距离在100 m以上。根据各洞段光照强度的不同将洞穴划分为有光带(光照度10 lx以上)、弱光带(光照度0.1—10 lx)和黑暗带(光照度0—0.1 lx)[21]进行调查, 使用泰仕tes1334a数字照度计测定光照强度。
1.2.2 调查和分类鉴定除洞穴3有光带和黑暗带面积较小, 仅设置1个样方, 弱光带设置3个样方外, 其余洞穴在有光带、弱光带和黑暗带分别随机设置3个样方, 每个自然林设置5个样方进行调查, 同一生境样方间距不小于5 m, 取样面积为5 cm×5 cm, 每个样方取样2份, 分别用于小型土壤节肢动物分离和理化指标测定。由于大多数洞穴土层较薄, 因此小型土壤节肢动物和土壤理化性质分析用的土样取样深度均为5 cm, 为了便于平行比较, 自然林小型土壤节肢动物和土壤理化性质分析用的土样取样深度也为5 cm。
小型土壤节肢动物在室内使用Tullgren法24 h分离获取, 获得的小型土壤节肢动物使用《中国土壤动物检索图鉴》[22]、《The Oribatid Mites Genera of the World》[23]和《A Manual of Acarology (Third edition)》[24]等工具书进行分类鉴定, 一般鉴定到科, 并统计个体数量。分类单元主要使用《中国土壤动物检索图鉴》[22]的分类系统。土壤理化性质的分析根据《土壤农业化学分析法》[25]完成:pH值采用电位法(水土比2.5∶1);有机质采用高温外热重铬酸钾氧化-容量法;全氮采用凯氏定氮法;全磷采用钼锑抗比色法;全钾采用火焰分光光度法;有效氮采用FeSO4-Zn还原- 碱解扩散法;有效磷采用钼锑抗比色法;有效钾采用火焰分光光度法。
1.2.3 数据分析处理采用Shannon-Wiener多样性指数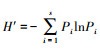
将同一季节每个洞穴或自然林所有样方的数据进行平均后再进行分析, 即洞穴4个重复, 自然林3个重复。数据进行log(x+1)转换降低非正态性后使用重复测量方差分析(Repeated measures ANOVA)检验生境和取样季节及其交互作用对小型土壤节肢动物的影响, 若差异显著则用LSD法(方差齐性)和Tamhane′s T2(M)法(方差不齐)进行多重比较。使用单因素方差分析(One-way ANOVA)对洞穴和自然林小型土壤节肢动物进行季节间的差异性检验。使用独立性T检验分析不同生境土壤理化性质的差异。使用Pearson系数(双尾检验) 进行相关性分析。
使用主成分分析(Principal component analysis, PCA)对小型土壤节肢动物群落进行排序, 以查明空间变化对小型土壤节肢动物群落结构的影响, 为减少排序图的复杂性, 仅使用密度百分比大于1%的类群进行PCA排序分析。
按生境将4个季节的小型土壤节肢动物数据进行平均后再使用冗余分析(RDA)确定小型土壤节肢动物多样性与环境因子之间的关系。为降低共线性的影响, 通过R语言vegan程序包中的函数vif.cca检验每个环境因子的VIF值, 若只有1个环境因子VIF值超过10, 则直接删除后进行RDA分析, 若VIF值超过10的环境因子≥2个, 则按VIF值由大到小逐个删除直至每个环境因子VIF值均低于10后再进行RDA分析。最终, 筛选出pH值、有机质(SOM)、有效钾(AK)和有效磷(AP)进行RDA分析。
优势类群的划分:个体密度占总密度10.0%以上者为优势类群, 占1.0%—10.0%者为常见类群, 不足1.0%者为稀有类群[9]。
以上数据的分析和作图在Excel 2010、Origin 2016和SPSS 16.0上完成, 使用R语言[26]进行PCA分析和RDA分析并实现可视化。
2 结果与分析 2.1 小型土壤节肢动物群落组成调查获得小型土壤节肢动物2399个, 隶属7纲15目121科(表 2)。其中优势类群为等节跳科(Isotomidae), 占总密度的11.58%;常见类群为杆棱甲螨科(Mochlozetidae)、懒甲螨总科(Nothroidea)和若甲螨科(Oribatulidae)等25类, 占总密度的66.82%;稀有类群为同翅目(Homoptera)、单翼甲螨科(Haplozetidae)和粪蚊科(Scatopsidae)等109类, 占总密度的21.60%。
| 样地 Plot |
地理位置 Location |
海拔/m Elevation |
土壤类型及特征 Soil types and characterization |
主要植被 Dominant vegetation species |
| 洞穴1 Cave 1 |
惠水县高石墙村(106°38′46″E, 26°0′6″N) | 1223 | 黄棕壤, 弱光带和黑暗带夏季和秋季土壤湿度较大且质地较硬 | 有光带有少量苔藓(Bryophytes)、杂草(Weeds)和蕨类(Ferns) |
| 洞穴2 Cave 2 |
惠水县高石墙村(106°38′32″E, 25°57′28″N) | 1153 | 碱性石灰土, 弱光带和黑暗带夏季和秋季土壤湿度较大 | 无植物 |
| 洞穴3 Cave 3 |
惠水县双坪村(106°35′27″E, 26°08′44″N) | 1119 | 黄棕壤, 弱光带和黑暗带夏季和秋季土壤湿度较大且土壤质地较硬 | 无植物 |
| 洞穴4 Cave 4 |
长顺县板床村(106°28′4″E, 25°58′8″N) | 1111 | 黄棕壤, 弱光带和黑暗带夏季和秋季土壤湿度较大且土壤质地较硬 | 有光带有少量杂草 |
| 自然林1 Natural forest 1 |
茂兰国家级自然保护区(107°56′59″E, 25°17′1″N) | 950 | 黑色石灰土, 土壤较松软 | 圆果化香(platycarya longipes)、青冈栎(Cyclobalanopsis glauca)、樟叶槭(Acer cinnamomifolium)和翠云草(Selaginella uncinata)等。林龄60年 |
| 自然林2 Natural forest 2 |
茂兰国家级自然保护区(107°56′59″E, 25°16′56″N) | 848 | 黑色石灰土, 土壤较松软 | 圆果化香、樟叶槭、掌叶木(Handeliodendron bodinieri)、朴树(Celtis sinensis)、香叶树(Lindera communis)等。林龄80年 |
| 自然林3 Natural forest 3 |
茂兰国家级自然保护区(107°56′43″E, 25°17′5″N) | 943 | 黑色石灰土, 土壤较松软 | 圆果化香、樟叶槭、黄梨木(Boniodendron minus)、掌叶木、香叶树等。林龄60年 |
| 洞穴Cave | 自然林Natural forest | 平均Average | ||||||
| 密度 Density/ (个/m2) |
百分比 Percent/% |
密度 Density/ (个/m2) |
百分比 Percent/% |
密度 Density/ (个/m2) |
百分比 Percent /% |
|||
| 螨类Mites(70.55%) | ||||||||
| 杆棱甲螨科Mochlozetidae | 25.00 | 2.25 | 1033.33 | 7.59 | 529.17 | 7.18 | ||
| 懒甲螨总科Nothroidea | 6.25 | 0.56 | 840.00 | 6.17 | 423.13 | 5.74 | ||
| 若甲螨科Oribatulidae | 3.13 | 0.28 | 826.67 | 6.07 | 414.90 | 5.63 | ||
| 盖头甲螨科Tectocepheidae | 21.88 | 1.97 | 706.67 | 5.19 | 364.27 | 4.95 | ||
| 缝甲螨科Hypochthoniidae | 3.13 | 0.28 | 473.33 | 3.48 | 238.23 | 3.23 | ||
| 盾珠甲螨科Suctobelbidae | 6.25 | 0.56 | 466.67 | 3.43 | 236.46 | 3.21 | ||
| 厚厉螨科Pachylaelapidae | 50.00 | 4.49 | 420.00 | 3.08 | 235.00 | 3.19 | ||
| 尾足螨科Uropodidae | 466.67 | 3.43 | 233.33 | 3.17 | ||||
| 矮汉甲螨科Nanhermanniidae | 3.13 | 0.28 | 446.67 | 3.28 | 224.90 | 3.05 | ||
| 奥甲螨科Oppiidae | 231.25 | 20.79 | 200.00 | 1.47 | 215.63 | 2.93 | ||
| 上罗甲螨科Epilohmanniidae | 3.13 | 0.28 | 313.33 | 2.30 | 158.23 | 2.15 | ||
| 阿斯甲螨科Astegistidae | 18.75 | 1.69 | 273.33 | 2.01 | 146.04 | 1.98 | ||
| 厉螨科Laelapidae | 15.63 | 1.40 | 253.33 | 1.86 | 134.48 | 1.83 | ||
| 囊螨科Ascidae | 240.00 | 1.76 | 120.00 | 1.63 | ||||
| 盾螨科Scutacaridae | 233.33 | 1.71 | 116.67 | 1.58 | ||||
| 剑甲螨科Gustaviidae | 173.33 | 1.27 | 86.67 | 1.18 | ||||
| 大翼甲螨科Galumnidae | 153.33 | 1.13 | 76.67 | 1.04 | ||||
| 毛跳甲螨科Zetomotrichidae | 153.33 | 1.13 | 76.67 | 1.04 | ||||
| 美绥螨科Ameroseiidae | 3.13 | 0.28 | 146.67 | 1.08 | 74.90 | 1.02 | ||
| 单翼甲螨科Haplozetidae | 9.38 | 0.84 | 126.67 | 0.93 | 68.02 | 0.92 | ||
| 土革螨科Ologamasidae | 62.50 | 5.62 | 46.67 | 0.34 | 54.58 | 0.74 | ||
| 滑珠甲螨科Damaeolidae | 106.67 | 0.78 | 53.33 | 0.72 | ||||
| 肉食螨科Cheylrtidae | 103.13 | 9.27 | 51.56 | 0.70 | ||||
| 洼甲螨科Camisiidae | 9.38 | 0.84 | 93.33 | 0.69 | 51.35 | 0.70 | ||
| 副大翼甲螨科Parakalumnidae | 100.00 | 0.73 | 50.00 | 0.68 | ||||
| 寄螨科Parasitidae | 100.00 | 0.73 | 50.00 | 0.68 | ||||
| 寄螨目Parasiformes | 9.38 | 0.84 | 86.67 | 0.64 | 48.02 | 0.65 | ||
| 足翅甲螨科Podopterotegaeidae | 86.67 | 0.64 | 43.33 | 0.59 | ||||
| 若螨Nymph | 3.13 | 0.28 | 80.00 | 0.59 | 41.56 | 0.56 | ||
| 真伊螨科Eviphididae | 80.00 | 0.59 | 40.00 | 0.54 | ||||
| 直卷甲螨科Archoplophoridae | 66.67 | 0.49 | 33.33 | 0.45 | ||||
| 步甲螨科Carabodidae | 6.25 | 0.56 | 60.00 | 0.44 | 33.13 | 0.45 | ||
| 礼服甲螨科Trhypochthoniidae | 25.00 | 2.25 | 40.00 | 0.29 | 32.50 | 0.44 | ||
| 短甲螨科Brachychthoniidae | 25.00 | 2.25 | 40.00 | 0.29 | 32.50 | 0.44 | ||
| 盲甲螨科Malaconothridae | 3.13 | 0.28 | 60.00 | 0.44 | 31.56 | 0.43 | ||
| 隐颚螨科Cryptognathidae | 40.63 | 3.65 | 20.00 | 0.15 | 30.31 | 0.41 | ||
| 尖棱甲螨科Ceratozetidae | 53.33 | 0.39 | 26.67 | 0.36 | ||||
| 真卷甲螨科Euphthiracaridae | 46.67 | 0.34 | 23.33 | 0.32 | ||||
| 赫甲螨科Hermanniidae | 40.00 | 0.29 | 20.00 | 0.27 | ||||
| 植绥螨科Phytoseiidae | 3.13 | 0.28 | 33.33 | 0.24 | 18.23 | 0.25 | ||
| 垂盾甲螨科Scutoverticidae | 6.25 | 0.56 | 26.67 | 0.20 | 16.46 | 0.22 | ||
| 平脊甲螨科Autognetidae | 31.25 | 2.81 | 15.63 | 0.21 | ||||
| 异珠足甲螨科Heterobelbidae | 9.38 | 0.84 | 20.00 | 0.15 | 14.69 | 0.20 | ||
| 跗线螨科Tarsonmidae | 28.13 | 2.53 | 14.06 | 0.19 | ||||
| 镰螯螨科Tydeidae | 3.13 | 0.28 | 20.00 | 0.15 | 11.56 | 0.16 | ||
| 叶螨科Tetranychidae | 20.00 | 0.15 | 10.00 | 0.14 | ||||
| 绒螨科Trombidiidae | 20.00 | 0.15 | 10.00 | 0.14 | ||||
| 异小黑螨科Xenocaligonellidae | 20.00 | 0.15 | 10.00 | 0.14 | ||||
| 双革螨科Digamasellidae | 20.00 | 0.15 | 10.00 | 0.14 | ||||
| 隐爪螨科Nanorchestidae | 12.50 | 1.12 | 6.67 | 0.05 | 9.58 | 0.13 | ||
| 甲螨亚目Oribatida | 3.13 | 0.28 | 13.33 | 0.10 | 8.23 | 0.11 | ||
| 罗甲螨科Lohmanniidae | 3.13 | 0.28 | 13.33 | 0.10 | 8.23 | 0.11 | ||
| 鲜甲螨科Cepheidae | 3.13 | 0.28 | 13.33 | 0.10 | 8.23 | 0.11 | ||
| 四奥甲螨科Quadroppiidae | 13.33 | 0.10 | 6.67 | 0.09 | ||||
| 德之甲螨科Tokunocepheidae | 13.33 | 0.10 | 6.67 | 0.09 | ||||
| 卷甲螨科Phthiracaridae | 13.33 | 0.10 | 6.67 | 0.09 | ||||
| 邦甲螨科Banksinomidae | 13.33 | 0.10 | 6.67 | 0.09 | ||||
| 真足螨科Eupodidae | 13.33 | 0.10 | 6.67 | 0.09 | ||||
| 胭螨科Rhodacaridae | 13.33 | 0.10 | 6.67 | 0.09 | ||||
| 表刻螨科Epicriidae | 13.33 | 0.10 | 6.67 | 0.09 | ||||
| 菌板鳃甲螨科Mycobatidae | 12.50 | 1.12 | 6.25 | 0.08 | ||||
| 前气门亚目Prostigmata | 12.50 | 1.12 | 6.25 | 0.08 | ||||
| 耳头甲螨科Otocepheidae | 3.13 | 0.28 | 6.67 | 0.05 | 4.90 | 0.07 | ||
| 长须螨科Stigmaeidae | 9.38 | 0.84 | 4.69 | 0.06 | ||||
| 沙足甲螨科Eremobelbidae | 6.67 | 0.05 | 3.33 | 0.05 | ||||
| 树缝甲螨科Arborichthoniidae | 6.67 | 0.05 | 3.33 | 0.05 | ||||
| 木单翼甲螨科Xylobatidae | 6.67 | 0.05 | 3.33 | 0.05 | ||||
| Tetracondylidae | 6.67 | 0.05 | 3.33 | 0.05 | ||||
| Ameronothridae | 6.67 | 0.05 | 3.33 | 0.05 | ||||
| 喜螨科Terpnacaridae | 6.67 | 0.05 | 3.33 | 0.05 | ||||
| (虫穴)螨科Zerconidae | 6.67 | 0.05 | 3.33 | 0.05 | ||||
| 足角螨科Podocinidae | 6.67 | 0.05 | 3.33 | 0.05 | ||||
| 粉螨亚目Acaridida | 6.67 | 0.05 | 3.33 | 0.05 | ||||
| 古甲螨科Palaeacaridae | 6.25 | 0.56 | 3.13 | 0.04 | ||||
| Haplochthoniidae | 3.13 | 0.28 | 1.56 | 0.02 | ||||
| 缰板鳃甲螨科Chamobatidae | 3.13 | 0.28 | 1.56 | 0.02 | ||||
| 球缝甲螨科Sphaerochthoniidae | 3.13 | 0.28 | 1.56 | 0.02 | ||||
| 高壳甲螨科Liodidae | 3.13 | 0.28 | 1.56 | 0.02 | ||||
| 滕岛螨科Teneriffidae | 3.13 | 0.28 | 1.56 | 0.02 | ||||
| 跳螨科Sphacrolichidae | 3.13 | 0.28 | 1.56 | 0.02 | ||||
| 弹尾目Collembola(18.70%) | ||||||||
| 等节跳科Isotomidae | 59.38 | 5.34 | 1646.67 | 12.09 | 853.02 | 11.58 | ||
| 疣跳科Neanuridae | 3.13 | 0.28 | 340.00 | 2.50 | 171.56 | 2.33 | ||
| 棘跳科Onychiuridae | 15.63 | 1.40 | 200.00 | 1.47 | 107.81 | 1.46 | ||
| 圆跳科Sminthuridae | 3.13 | 0.28 | 180.00 | 1.32 | 91.56 | 1.24 | ||
| 长角跳科Entomobryidae | 18.75 | 1.69 | 160.00 | 1.17 | 89.38 | 1.21 | ||
| 球角跳科Hypogastruridae | 68.75 | 6.18 | 53.33 | 0.39 | 61.04 | 0.83 | ||
| 鳞跳科Tomoceridae | 6.67 | 0.05 | 3.33 | 0.05 | ||||
| 昆虫纲Insecta(10.28%) | ||||||||
| 长角沼甲科幼虫Ptilodactylidae | 3.13 | 0.28 | 6.67 | 0.05 | 4.90 | 0.07 | ||
| 隐翅甲科幼虫Staphylinidae larvae | 3.13 | 0.28 | 33.33 | 0.24 | 18.23 | 0.25 | ||
| 缨甲科成虫Ptiliidae adult | 3.13 | 0.28 | 46.67 | 0.34 | 24.90 | 0.34 | ||
| 隐翅甲科成虫Staphylinidae adult | 3.13 | 0.28 | 60.00 | 0.44 | 31.56 | 0.43 | ||
| 步甲科成虫Scarabidae adult | 20.00 | 0.15 | 10.00 | 0.14 | ||||
| 盘甲科成虫Discolomidae adult | 20.00 | 0.15 | 10.00 | 0.14 | ||||
| 薪甲科成虫Lathridiidae adult | 6.67 | 0.05 | 3.33 | 0.05 | ||||
| 象甲科幼虫Curculionidae larvae | 6.67 | 0.05 | 3.33 | 0.05 | ||||
| 拟球甲科幼虫Corylophidae larvae | 3.13 | 0.28 | 1.56 | 0.02 | ||||
| 粗角叩甲科幼虫Throscidae larvae | 3.13 | 0.28 | 1.56 | 0.02 | ||||
| 拟步甲科幼虫Tenebrionidae larvae | 6.67 | 0.05 | 3.33 | 0.05 | ||||
| 古瘦隐甲科幼虫Archeocrypticidae larvae | 6.67 | 0.05 | 3.33 | 0.05 | ||||
| 花萤科幼虫Cantharidae larvae | 6.67 | 0.05 | 3.33 | 0.05 | ||||
| 金龟甲科幼虫Scarabaeidae larvae | 6.67 | 0.05 | 3.33 | 0.05 | ||||
| 萤科幼虫Lampyridae larvae | 6.67 | 0.05 | 3.33 | 0.05 | ||||
| 叩甲科幼虫Elateridae larvae | 20.00 | 0.15 | 10.00 | 0.14 | ||||
| 球蕈甲科幼虫Leiodidae larvae | 6.67 | 0.05 | 3.33 | 0.05 | ||||
| 蚁甲科成虫Pselaphidae adult | 6.67 | 0.05 | 3.33 | 0.05 | ||||
| 苔甲科成虫Scydmaenidae adult | 13.33 | 0.10 | 6.67 | 0.09 | ||||
| 蝇科Muscidae | 3.13 | 0.28 | 13.33 | 0.10 | 8.23 | 0.11 | ||
| 寡脉蝇科Asteiidae | 26.67 | 0.20 | 13.33 | 0.18 | ||||
| 舞虻科Empididae | 6.67 | 0.05 | 3.33 | 0.05 | ||||
| 长足虻科Dolichopodidae | 6.67 | 0.05 | 3.33 | 0.05 | ||||
| 小黑螨科Caligonellidae | 6.67 | 0.05 | 3.33 | 0.05 | ||||
| 实蝇科Tephritidae | 6.67 | 0.05 | 3.33 | 0.05 | ||||
| 蚤蝇科Phoridae | 13.33 | 0.10 | 6.67 | 0.09 | ||||
| 鼓翅蝇科Sepsidae | 6.67 | 0.05 | 3.33 | 0.05 | ||||
| 缟蝇科Lauxaniidae | 6.67 | 0.05 | 3.33 | 0.05 | ||||
| 花蝇科Anthomyiidae | 25.00 | 2.25 | 12.50 | 0.17 | ||||
| 蠓科Bibionidae | 18.75 | 1.69 | 9.38 | 0.13 | ||||
| 粪蚊科Scatopsidae | 133.33 | 0.98 | 66.67 | 0.91 | ||||
| 摇蚊科Chironomidae | 146.67 | 1.08 | 73.33 | 1.00 | ||||
| 蚁科Formicidae | 12.50 | 1.12 | 553.33 | 4.06 | 282.92 | 3.84 | ||
| 蜂类Bee | 13.33 | 0.10 | 6.67 | 0.09 | ||||
| 半翅目Hemiptera | 6.67 | 0.05 | 3.33 | 0.05 | ||||
| 土蝽科Cydnidae | 13.33 | 0.10 | 6.67 | 0.09 | ||||
| 同翅目Homoptera | 6.25 | 0.56 | 133.33 | 0.98 | 69.79 | 0.95 | ||
| 蚜虫Aphid | 6.67 | 0.05 | 3.33 | 0.05 | ||||
| 啮目Psocoptera | 3.13 | 0.28 | 1.56 | 0.02 | ||||
| 虱啮科Liposcelididae | 6.67 | 0.05 | 3.33 | 0.05 | ||||
| 管蓟马科Phlaeothripidae | 33.33 | 0.24 | 16.67 | 0.23 | ||||
| 离蛹Free pupa | 6.67 | 0.05 | 3.33 | 0.05 | ||||
| 其他Others(0.47%) | ||||||||
| 古蚖目Eosentomata | 3.13 | 0.28 | 1.56 | 0.02 | ||||
| 球马陆科Glomeridae | 6.67 | 0.05 | 3.33 | 0.05 | ||||
| 钩马陆科Harpagophoridae | 6.67 | 0.05 | 3.33 | 0.05 | ||||
| 石蜈蚣目Lithobiomorpha | 6.67 | 0.05 | 3.33 | 0.05 | ||||
| 幺蚣科Scolopendrellidae | 26.67 | 0.20 | 13.33 | 0.18 | ||||
| 苦伪蝎科Chernetidae | 20.00 | 0.15 | 10.00 | 0.14 | ||||
| 总个体数Individual number | 356 | 2043 | 2399 | |||||
| 总类群数Group number | 64 | 116 | 136 | |||||
PCA排序分析结果表明, 各季节洞穴与自然林小型土壤节肢动物群落组成结构差异明显(图 1)。不同季节, 影响小型土壤节肢动物群落结构的主要类群有不同。春季, 在PC1轴上影响群落结构的主要类群是若甲螨科和厚厉螨科(Pachylaelapidae);夏季, 在PC1轴上影响群落结构的主要类群是盾珠甲螨科(Suctobelbidae)和尾足螨科(Uropodidae);秋季, 在PC1轴上影响群落结构的主要类群是尾足螨科;冬季, 在PC1轴上影响群落结构的主要类群是蚁科(Formicidae)。

|
| 图 1 小型土壤节肢动物主成分分析排序图 Fig. 1 Principal component analysis (PCA) ordination diagrams of soil microarthropod |
各季节均以自然林小型土壤节肢动物类群数、密度和Shannon-Wiener指数(H′)高于洞穴, 但洞穴和自然林小型土壤节肢动物类群数、密度和Shannon-Wiener指数(H′)季节变化有不同(图 2)。Repeated measures ANOVA分析显示, 洞穴小型土壤节肢动物类群数、密度和Shannon-Wiener指数(H′)与自然林差异显著(P < 0.01), 季节变化显著影响小型土壤节肢动物类群数、密度和Shannon-Wiener指数(H′)(P < 0.05), 交互作用显著影响类群数和Shannon-Wiener指数(H′)(P < 0.05)。One-way ANOVA结果表明, 自然林小型土壤节肢动物类群数(F=24.56, P < 0.01)、密度(F=21.30, P < 0.05)和Shannon-Wiener指数(H′)(F=29.51, P < 0.05)季节波动明显, 洞穴小型土壤节肢动物类群数、密度和Shannon-Wiener指数(H′)无显著季节差异。

|
| 图 2 小型土壤节肢动物多样性(平均值±标准误) Fig. 2 Diversity of soil microarthropod (Mean±SE) 小写字母不同表示同一生境不同季节间差异显著 |
不同季节洞穴螨类、跳虫和昆虫密度均低于自然林(图 3)。Repeated measures ANOVA分析表明, 洞穴与自然林螨类、跳虫和昆虫密度差异显著(P < 0.05), 季节变化对螨类密度影响显著(P < 0.05), 对跳虫和昆虫密度影响不明显, 交互作用对螨类、跳虫和昆虫密度影响均不明显。One-way ANOVA结果显示, 自然林螨类密度存在显著的季节差异(F=28.17, P < 0.001)。

|
| 图 3 不同小型土壤节肢动物类群个体密度(平均值±标准误) Fig. 3 The dynamic in individual density of different groups (Mean±SE) |
各生境土壤化学性质如表 3所示。洞穴与自然林土壤pH值、全磷和全钾差异显著(P<0.05), 其余无显著不同。
| 洞穴 Cave |
自然林 Natural forest |
t | P | |
| pH值 | 6.89±0.17 | 5.75±0.27 | 3.54 | <0.05 |
| SOM/(g/kg) | 41.87±10.8 | 62.52±11.22 | -1.95 | >0.05 |
| TN/(g/kg) | 4.15±1.03 | 3.66±0.38 | -0.51 | >0.05 |
| TP/(g/kg) | 13.51±0.81 | 8.48±0.80 | 5.26 | <0.01 |
| TK/(g/kg) | 3.67±0.63 | 0.62±0.09 | 2.83 | <0.05 |
| AN/(mg/kg) | 1066.49±304.21 | 321.91±42.27 | 1.21 | >0.05 |
| AP/(mg/kg) | 1063.83±271.07 | 122.40±18.16 | 1.03 | >0.05 |
| AK/(mg/kg) | 72.48±26.29 | 15.69±2.58 | 2.20 | >0.05 |
| SOM: 有机质Soil organic matter; TN: 全氮Total nitrogen; TP: 全磷Total phosphorus; TK: 全钾Total potassium; AN: 有效氮Available nitrogen; AP: 有效磷Available phosphorus; AK: 有效钾Available potassium | ||||
环境因子与小型土壤节肢动物相关性分析结果(表 4)表明, 小型土壤节肢动物类群数和Shannon-Wiener指数(H′)与pH值、全磷、全钾和光照强度显著相关(P < 0.05), 密度与pH值、全磷、有机质和光照强度显著相关(P < 0.05), 土壤跳虫和昆虫密度与pH值和有机质显著相关(P < 0.05), 土壤螨类密度与pH值、全氮和有机质显著相关(P < 0.05)。
| 类群数 Group number |
密度 Density/ (个/m2) |
多样性指数 Shannon- Wiener index(H′) |
螨类密度 Density of mite/ (个/m2) |
跳虫密度 Density of springtail/ (个/m2) |
昆虫密度 Density of insect/ (个/m2) |
|
| pH | -0.90** | -0.95** | -0.89** | 0.95** | 0.93** | 0.93** |
| TN | 0.34 | 0.53 | 0.30 | -0.76* | -0.71 | -0.74 |
| AN | -0.37 | -0.20 | -0.40 | -0.12 | -0.11 | -0.10 |
| TP | -0.88** | -0.78* | -0.88** | 0.51 | 0.48 | 0.51 |
| AP | -0.30 | -0.11 | -0.33 | -0.19 | -0.14 | -0.17 |
| TK | -0.74 | -0.65 | -0.75 | 0.42 | 0.43 | 0.44 |
| AK | -0.67 | -0.57 | -0.67 | 0.31 | 0.29 | 0.33 |
| SOM | 0.74 | 0.86* | 0.72 | -0.96** | -0.97** | -0.96** |
| 光照强度Light intensity | 0.98** | 0.92** | 0.99** | -0.75 | -0.75 | -0.75 |
| *表示显著水平为P < 0.05, **表示显著水平为P < 0.01 | ||||||
RDA分析显示, 第一轴和第二轴对小型土壤节肢动物多样性与土壤因子关系的解释率为91.14%, 且所有轴(F=16.54, P < 0.05)和第一轴(F=64.09, P < 0.05)在统计学上达显著水平(图 4)。小型土壤节肢动物类群数、密度、Shannon-Wiener指数(H′)、螨类密度、跳虫密度和昆虫密度与土壤有机质正相关, 与pH值、有效钾和有效磷负相关。蒙特卡洛检验显示, 土壤pH值是影响小型土壤节肢动物多样性最主要的土壤因子(P < 0.01)。

|
| 图 4 群落多样性与土壤因子之间的冗余分析 Fig. 4 Redundance analysis (RDA) of community diversity explained by soil factors GNSM: 小型土壤节肢动物类群数Group number of soil microarthropod; DSM: 小型土壤节肢动物密度Density of soil microarthropod; H′: 小型土壤节肢动物多样性指数Shannon-Wiener index; DM: 螨类密度Density of mite; DS: 跳虫密度Density of springtail; DI: 昆虫密度Density of insect |
生多样性的高低能反映生态系统稳定性和食物网的复杂程度[27-28]。洞穴平均Shannon-Wiener指数(H′)为0.27, 不仅低于本研究中的自然林, 且低于喀斯特地区大多数的生态系统类型, 如火烧迹地(1.43—2.18)[10], 草地(0.53), 灌木丛(0.88), 灌木林(0.87)[29], 农田(2.47)和撂荒地(2.28)[30], 表明喀斯特洞穴生态系统具有稳定性差, 食物网简单的特征。
本研究结果表明, 洞穴小型土壤节肢动物群落结构与自然林差异明显, 优势类群明显不同。从自然林到洞穴, 优势类群等节跳科密度由1646.67个/m2显著下降到59.38个/m2(P < 0.05), 在群落中的比例随之大幅度降低(12.09% 到5.23%);自然林常见类群奥甲螨科(Oppiidae)个体密度(200.00个/m2)与洞穴(231.25个/m2)接近(P>0.05), 但在洞穴中的比例(20.79%)较自然林(1.74%)大幅度提升, 转变为优势类群。可以看出, 奥甲螨科对洞穴环境的适应能力明显强于等节跳科, 食性不同可能是二者空间选择存在差异的原因。奥甲螨科为杂食性, 而等节跳科为菌食性[31], 杂食性土壤动物食源广泛, 较菌食性土壤动物对环境变化具有更强的适应能力[32-33], 所以奥甲螨科在洞穴这样相对严酷的环境中仍有较高的优势地位。朱新玉等[34]也撰文指出杂食性是节肢动物对不稳定环境适应的表现。
独有类群对环境条件有强烈的依赖性, 只在特定的环境出现, 所以生境间独有类群的多寡对小型土壤节肢动物群落组成结构差异有重要影响。自然林和洞穴共有类群为45个, 独有类群分别为71个和19个, 分别占洞穴和自然林小型土壤节肢动物总类群数的61.21%和29.69%, 说明洞穴和自然林小型土壤节肢动物群落组成差异较大, 自然林小型土壤节肢动物群落组成较为复杂, 而洞穴小型土壤节肢动物群落组成相对简单。土壤动物群落组成和结构受植物群落物种组成和凋落物质量和含量的影响[35-36]。洞穴与自然林无论是植物的组成, 还是凋落物的质量和数量都表现出较大的不同(表 1), 导致小型土壤节肢动物群落组成在不同空间产生明显差异。
本研究发现, 小型土壤节肢动物类群数、密度和多样性与土壤全磷和pH值显著正相关, 与光照强度显著负相关, 表明土壤全磷、pH值和光照强度对小型土壤节肢动物群落有重要影响。光是植物的重要能量来源, 也是植物生长的基本条件之一, 光照强度的降低不利于植物的生长和生物量的积累[37-38], 但土壤动物分布受植被的影响[39-40], 植物多样性越高, 意味着环境异质性越高, 能为土壤动物提供更丰富的小生境和多样化的食物资源, 从而促进土壤动物多样性的提高[41-42]。洞穴环境光照不足, 在黑暗带甚至不能进行光合作用, 制约了植物的生长分布, 食物资源贫乏和栖息环境的严酷性限制了小型土壤节肢动物种类和数量的增长, 小型土壤节肢动物类群数、密度和多样性因此较低。自然林光照充足, 植物多样性高, 能为小型土壤节肢动物提供多样化的食物资源和相对良好的栖息环境, 小型土壤节肢动物类群数、密度和多样性也较高。表明光照可以通过改变植物生长从而间接影响小型土壤节肢动物空间分布。
适量的磷元素对土壤节肢动物有利, 过多则会产生不利影响[43]。洞穴小型土壤节肢动物类群数、密度和多样性均显著低于自然林, 显示了洞穴过高的土壤全磷含量确实对小型土壤节肢动物产生了明显的负面作用。相关研究表明, 微酸性和中性条件更适宜于大多数土壤动物的繁衍生息[44-45]。本研究发现, 在研究区酸性土壤环境中, 低土壤酸度的洞穴环境对小型土壤节肢动物有明显的抑制作用(表 3和表 4), 与何振等[46]的研究结果一致。相比之下, 土壤酸性较高的自然林更有利于小型土壤节肢动物的生存。RDA分析也表明土壤节肢动物类群数、密度和多样性与低pH值环境密切相关, 螨类、跳虫和昆虫密度在土壤酸性较高的自然林更为丰富(图 3, 图 4)。张洪芝等[47]对青藏东缘若尔盖高寒草甸中小型土壤动物进行研究时也有类似发现, 并认为这是土壤动物长期生活于低温环境, 形成了与环境相适应的生理机制和低温度忍耐限度所致。喀斯特小型土壤节肢动物在长期的演化历程中可能已经形成独特的适应机制, 对环境变化的响应具有特殊性。
洞穴内湿度较高, 土壤在夏季和秋季也保持了较高湿度, 在弱光带和黑暗带甚至可达到过饱和状态。与此同时, 洞道在向内部延伸的过程中不仅土层厚度会逐渐降低, 土壤质地还会趋于硬化。土壤水分处于饱和或过饱和状态会对生活在其间的一些土壤节肢动物产生致死作用[48]或影响其繁殖活动[49], 而土壤的硬化板结, 则能使土壤孔隙度降低, 紧密度提升, 孔隙中所能提供的氧气减少, 土壤动物会因呼吸作用受阻而密度降低[50-52]。对于像跳虫这类对孔隙度敏感的土壤节肢动物, 孔隙度的降低还意味着其生存空间的减少, 由于不能进行掘穴活动, 为了保护体表蜡质层, 跳虫通常会避免在低孔隙土壤环境中活动[53]。尾足螨个体数量也与土壤孔隙有相关性, 其个体数量可随孔隙体积的缩小而降低[54]。不仅如此, 低孔隙土壤环境还能阻碍微生物的生长[55], 在上行效应的作用下, 菌食性土壤节肢动物数量会因缺少食物而下降, 进而影响群落整体的密度。以上分析说明, 光照强度低, 植被稀少, 土壤条件差是洞穴土壤节肢动物类群数、密度和多样性维持在较低水平的重要原因。
自然林和洞穴小型土壤节肢动物季节动态有不同, 自然林小型土壤节肢动物类群数、密度和多样性有显著季节变化, 洞穴小型土壤节肢动物类群数、密度和多样性受季节变化的影响不明显。已有的研究表明植被[56]、温度[57]和降水[58]是引起土壤动物季节性变化的重要原因。秋季气温和水分条件适宜, 是自然林凋落物增加的高峰期, 营养源的增加能促进土壤动物多样性提升[59], 一些昆虫也会在此阶段进入土壤冬眠[60], 使秋季自然林小型土壤节肢动物类群、密度和多样性显著高于其他季节。虽然洞穴植物稀少, 对土壤动物季节变化影响力有限[61], 但洞穴的温湿度较稳定[62], 所以洞穴小型土壤节肢动物季节稳定性高。
4 结论奥甲螨科对洞穴适应能力强, 等节跳科在自然林最为丰富。洞穴小型土壤节肢动物群落组成结构与自然林明显不同, 且类群数、密度和Shannon-Wiener指数(H′)差异显著。洞穴小型土壤节肢动物类群数、密度和Shannon-Wiener指数(H′)季节稳定性高, 而自然林小型土壤节肢动物类群数、密度和Shannon-Wiener指数(H′)季节敏感性强。土壤酸碱度是影响螨类、跳虫和昆虫密度以及小型土壤节肢动物多样性的主要环境因子。研究表明, 土壤因子和光照强度的不同使小型土壤节肢动物在洞穴和自然林呈差异化分布。洞穴生态系统小型土壤节肢动物种类少, 密度低, 多样性差, 生态系统稳定性弱, 应该加强保护, 避免不合理的开发利用。
致谢: 感谢茂兰国家级自然保护区管理局在调查采样过程中给予的协助, 感谢贵阳学院陈欢、吴骄、瞿明兵和邹龙秋等同学协助野外调查采样和室内分离工作。| [1] |
邓实群. 贵州省荔波县洞穴动物及其生态学研究[D]. 贵阳: 贵州师范大学, 2005.
|
| [2] |
韩康. 应用稳定同位素研究贵州施秉下洞和木老唐洞的营养级和食物网[D]. 贵阳: 贵州师范大学, 2014.
|
| [3] |
班凤梅. 洞穴滴水地球化学变化特征及其与土壤过程的联系——以北京石花洞为例[D]. 南京: 南京农业大学, 2007.
|
| [4] |
邵元虎, 张卫信, 刘胜杰, 王晓丽, 傅声雷. 土壤动物多样性及其生态功能. 生态学报, 2015, 35(20): 6614-6625. |
| [5] |
Bender S F, Wagg C, Heijden M M A V. An Underground Revolution: Biodiversity and Soil Ecological Engineering for Agricultural Sustainability. Trends in Ecology & Evolution, 2016, 31(6): 440-452. |
| [6] |
Neher D A. Soil community composition and ecosystem processes Comparing agricultural ecosystems with natural ecosystems. Agroforestry Systems, 1999, 45: 159-185. DOI:10.1023/A:1006299100678 |
| [7] |
Sharma N, Parwez H. Population density and diversity of Soil mites (Order: acarina) in agroforestry habitat: Relationship to Soil temperature and Soil moisture. International Journal of Applied Environmental Sciences, 2017, 12: 1449-1460. |
| [8] |
Barros E, Pashanasi B, Constantino R, Lavelle P. Effects of land-use system on the soil macrofauna in western Brazilian Amazonia. Biology & Fertility of Soils, 2002, 35(5): 338-347. |
| [9] |
薛娟, 魏雪, 何先进, 吴鹏飞. 高寒草甸生态系统蚁丘对小型土壤节肢动物群落的影响. 生态学报, 2021, 41(4): 1613-1624. |
| [10] |
杨大星, 杨茂发, 徐进, 尚小丽. 马尾松人工林火烧迹地不同恢复阶段中小型土壤节肢动物多样性. 生态学报, 2013, 33(8): 2531-2544. |
| [11] |
杨大星, 杨茂发, 尚小丽, 师沛琼. 黔南喀斯特不同火烧迹地土壤动物群落特征比较. 西南农业学报, 2012, 25(6): 2190-2197. DOI:10.3969/j.issn.1001-4829.2012.06.047 |
| [12] |
陈浒, 金道超, 张燕. 喀斯特小流域栓皮栎林土壤螨类群落结构特征. 应用生态学报, 2018, 29(5): 1667-1676. |
| [13] |
王鹏举, 陈浒, 周政, 林丹丹, 巫仁霞, 朱佳运. 梵净山常绿落叶阔叶混交林土壤螨类群落结构研究. 土壤, 2018, 50(4): 687-695. |
| [14] |
杨大星, 杨茂发. 黔南不同撂荒地土壤节肢动物群落特征. 贵州农业科学, 2016, 44(6): 149-154. DOI:10.3969/j.issn.1001-3601.2016.06.040 |
| [15] |
杨大星, 杨茂发, 徐进. 生态恢复方式对喀斯特土壤节肢动物群落特征的影响. 贵州农业科学, 2013, 41(2): 91-94. DOI:10.3969/j.issn.1001-3601.2013.02.026 |
| [16] |
杨大星, 李灿, 杨茂发. 黔南喀斯特土壤螨类对火烧和垦殖的响应. 应用与环境生物学报, 2019, 25(6): 1335-1343. |
| [17] |
张智英, 张亮, 李玉辉, 何晓岚. 云南石林喀斯特景区不同生境大型土壤动物多样性研究. 林业科学研究, 2005, 18(6): 601-705. |
| [18] |
Raschmanová N, Miklisová D, Kováč L, Šustr V. Community composition and cold tolerance of soil Collembola in a collapse karst doline with strong microclimate inversion. Biologia, 2015, 70(6): 802-811. DOI:10.1515/biolog-2015-0095 |
| [19] |
Raschmanová N, Miklisová D, Kováč L'. Soil Collembola communities along a steep microclimatic gradientin the collapse doline of the SilickáIadnica Cave, Slovak Karst. Biologia, 2013, 68(3): 470-478. DOI:10.2478/s11756-013-0172-8 |
| [20] |
杨大星, 李灿, 王春. 洞穴内外中小型土壤节肢动物群落特征及其对环境因子的响应. 生态学杂志, 2021, 40(10): 3184-3194. |
| [21] |
黎道洪, 罗蓉, 陈浒. 贵州大洞口内若干动物群落研究. 生态学报, 2001, 21(1): 126-130. |
| [22] |
尹文英. 中国土壤动物检索图鉴. 北京: 科学出版社, 1998.
|
| [23] |
Balogh P, Balogh J. The Oribatid Mites Genera of the World(Vol.1 & 2). Budapest: The Hungarian National Museum Press, 1992.
|
| [24] |
Krantz G W, Walter D E. A Manual of Acarology(Third edition). Texas: Texas Tech University Press, 2009.
|
| [25] |
鲁如坤. 土壤农业化学分析法. 北京: 中国农业出版社, 2000.
|
| [26] |
Oksanen J, Blanchet F G, Friendly M, Kindt R, Legendre P, McGlinn D, Minchin P R, O'Hara R B, Simpson G L, Solymos P, Stevens M H H, Szoecs E, Wagner H. vegan: Community Ecology Package. R package version 2.5-6. 2019. https://CRAN.R-project.org /package=vegan.
|
| [27] |
Kardol P, Newton J S, Martijn Bezemer T, Maraun M, van der Putten W H. Contrasting diversity patterns of soil mites and nematodes in secondary succession. Acta Oecologica, 2009, 35: 603-609. DOI:10.1016/j.actao.2009.05.006 |
| [28] |
吴东辉, 张柏, 陈鹏. 吉林中、西部平原区土壤弹尾虫群落结构的比较. 昆虫学报, 2005, 48(6): 935-942. DOI:10.3321/j.issn:0454-6296.2005.06.020 |
| [29] |
李宜蓉, 龙健, 李娟, 刘灵飞, 廖洪凯, 王显, 杨睿. 退化喀斯特生态恢复过程对土壤微节肢动物群落多样性的影响. 农业环境科学学报, 2021, 40(2): 310-318. |
| [30] |
杨大星, 叶婷, 杨文佳, 李灿. 撂荒地植被恢复过程中中小型土壤节肢动物群落动态. 西南农业学报, 2020, 33(7): 1517-1527. |
| [31] |
张阿娟, 张健, 李金金, 刘志刚, 张丹桔. 轮伐期前后不同密度巨桉(Eucalyptus grandis)人工林土壤动物群落结构特征. 生态学报, 2020, 40(3): 808-821. |
| [32] |
林英华, 张夫道, 刘海东, 苏化龙. 百望山土壤动物群落结构在枸树落叶分解中的变化. 动物学杂志, 2005, 40(3): 60-66. DOI:10.3969/j.issn.0250-3263.2005.03.012 |
| [33] |
李艳红, 罗承德, 杨万勤, 胡杰, 吴福忠. 桉-桤混合凋落物分解及其土壤动物群落动态. 应用生态学报, 2011, 22(4): 851-856. |
| [34] |
朱新玉, 刘洁, 贺振, 吴东辉. 耕作方式对玉-豆轮作地表节肢动物多样性及其营养结构的影响. 生态学报, 2018, 38(20): 7242-7253. |
| [35] |
李红月, 殷秀琴, 马辰, 郭玉梅. 长白山地丘陵区不同土地利用方式土壤动物群落生态分布特征. 土壤学报, 2017, 54(4): 1018-1028. |
| [36] |
张安宁, 刘任涛, 刘佳楠, 赵娟, 常海涛, 陈蔚. 干旱风沙区柠条枯落物对土壤节肢动物群落的影响. 生态学杂志, 2020, 39(7): 2383-2391. |
| [37] |
马银山, 杜国祯, 张世挺. 光照强度和肥力变化对垂穗披碱草生长的影响. 生态学报, 2014, 34(14): 3908-3916. |
| [38] |
陈圣宾, 宋爱琴, 李振基. 森林幼苗更新对光环境异质性的响应研究进展. 应用生态学报, 2005, 16(2): 365-370. DOI:10.3321/j.issn:1001-9332.2005.02.034 |
| [39] |
Wu P F, Liu S R, Liu X L. Composition and spatio-temporal changes of soil macroinvertebrate communities in the biodiversity hotspot of the northern Hengduanshan Mountains, China. Plant Soil, 2012, 357: 321-338. DOI:10.1007/s11104-012-1166-y |
| [40] |
李小涵, 刘世荣, 魏雪, 吴鹏飞. 南亚热带6种人工林小型土壤节肢动物群落多样性. 生态学杂志, 2021, 40(5): 1458-1468. |
| [41] |
Vilardo G, Tognetti P M, González-Arzac A, Yahdjian L. Soil arthropod composition differs between old-fields dominated by exotic plant species and remnant native grasslands. Acta Oecologica, 2018, 91: 57-64. DOI:10.1016/j.actao.2018.06.003 |
| [42] |
Loranger-Merciris G, Imbert D, Bernhard-Reversat F, Ponge J F, Lavelle P. Soil fauna abundance and diversity in a secondary semi-evergreen forest in Guadeloupe (Lesser Antilles) influence of soil type and dominant tree species. Biology and Fertility of Soils, 2007, 44: 269-276. DOI:10.1007/s00374-007-0199-5 |
| [43] |
韩雪梅, 李丹丹, 梁子安, 陈云峰, 胡诚. 北方常见农业土地利用方式对土壤螨群落结构的影响. 生态学报, 2013, 33(16): 5026-5034. |
| [44] |
钱复生, 王宗英. 水东枣园土壤动物与土壤环境的关系. 应用生态学报, 1994, 6(1): 42-48. |
| [45] |
唐静, 袁访, 宋理洪. 施用生物炭对土壤动物群落的影响研究进展. 应用生态学报, 2020, 31(7): 2473-2480. |
| [46] |
何振, 赵琴, 李迪强, 李密. 八大公山土壤跳虫群落结构与多样性的季节动态. 林业科学研究, 2018, 31(3): 68-77. |
| [47] |
张洪芝, 吴鹏飞, 杨大星, 崔丽巍, 何先进, 熊远清. 青藏东缘若尔盖高寒草甸中小型土壤动物群落特征及季节变化. 生态学报, 2011, 31(15): 4385-4397. |
| [48] |
黄丽荣, 张雪萍. 大兴安岭寒温带地区中小型土壤动物群落特征. 应用与环境生物学报, 2008, 14(3): 388-393. DOI:10.3321/j.issn:1006-687X.2008.03.020 |
| [49] |
邓晓保, 邹寿青, 付先惠, 姚天全, 盛才余, 白智林. 西双版纳热带雨林不同土地利用方式对土壤动物个体数量的影响. 生态学报, 2003, 23(1): 130-138. DOI:10.3321/j.issn:1000-0933.2003.01.017 |
| [50] |
Kevan P G, Forbes B, Kevan S M, Behan-Pelletier V. Vehicle tracks on high Arctic tundra: their effects on the soil, vegetation and soil arthropods. Journal of Applied Ecology, 1995, 32: 655-667. DOI:10.2307/2404660 |
| [51] |
Niwranski K, Kevan P G, Fjellberg A. Effects of vehicle disturbance and soil compaction on Arctic collembolan abundance and diversity on Igloolik Island, Nunavut, Canada. European journal of soil biology, 2002, 38: 193-196. DOI:10.1016/S1164-5563(02)01142-1 |
| [52] |
杨旭, 林琳, 张雪萍, 张利敏, 沙迪. 松嫩平原典型黑土耕作区中小型土壤动物时空分布特征. 生态学报, 2016, 36(11): 3253-3260. |
| [53] |
Heisler C, Kaiser E. Influence of agricultural traffic and crop management on collembola and microbial biomass in arable soil. Biology and Fertility of Soils, 1995, 19: 159-165. DOI:10.1007/BF00336153 |
| [54] |
Koehler H H. Predatory mites (Gamasina, Mesostigmata). Agriculture Ecosystems & Environment, 1999, 74: 395-410. |
| [55] |
Roithmeier O, Pieper S. Influence of Enchytraeidae (Enchytraeus albidus) and compaction on nutrient mobilization in an urban soil. Pedobiologia, 2009, 53: 29-40. DOI:10.1016/j.pedobi.2009.02.001 |
| [56] |
周育臻, 吴鹏飞. 贡嘎山东坡森林小型土壤节肢动物群落多样性与时空分布. 生态学杂志, 2020, 39(2): 586-599. |
| [57] |
张蕊, 李佳, 高梅香. 玉米秸秆处理方式对冬季黑土农田蜱螨目和弹尾目群落结构的影响. 生态学报, 2020, 40(22): 8315-8325. |
| [58] |
廖崇惠, 李健雄, 杨悦屏, 张振才. 海南尖峰岭热带林土壤动物群落——群落结构的季节变化及其气候因素. 生态学报, 2003, 23(1): 139-147. |
| [59] |
黄玉梅, 李向, 张丹桔, 邓楚璇, 罗春兰, 罗紫薇, 沈金亮, 解文峰. 成都市温江区不同栽植年限园林植物土壤动物群落特征. 应用生态学报, 2020, 31(11): 3859-3868. |
| [60] |
李娜, 张雪萍, 张利敏. 三种温带森林大型土壤动物群落结构的时空动态. 生态学报, 2013, 33(19): 6236-6245. |
| [61] |
吴鹏飞, 杨大星. 若尔盖高寒草甸退化对中小型土壤动物群落的影响. 生态学报, 2011, 31(13): 3745-3757. |
| [62] |
徐承香, 黎道洪, 梅杰. 贵州大水洞和马狼洞内动物群落结构与部分环境因子的相关性研究. 贵州师范大学学报(自然科学版), 2010, 28(2): 1-9. DOI:10.3969/j.issn.1004-5570.2010.02.001 |