 2022, Vol. 42
2022, Vol. 42文章信息
- 魏识广, 叶万辉, 练琚愉, 李林, 周景钢
- WEI Shiguang, YE Wanhui, LIAN Juyu, LI Lin, ZHOU Jinggang
- 南亚热带常绿阔叶林群落及其卫星样地物种多样性特征
- Species diversity characteristics of lower subtropical evergreen broad-leaved forest community and its satellite plots
- 生态学报. 2022, 42(11): 4515-4523
- Acta Ecologica Sinica. 2022, 42(11): 4515-4523
- http://dx.doi.org/10.5846/stxb202105071190
-
文章历史
- 收稿日期: 2021-05-07
- 网络出版日期: 2022-02-10





2. 广西师范大学珍稀濒危动植物生态与环境保护教育部重点实验室, 桂林 541006;
3. 广西漓江流域景观资源保育与可持续利用重点实验室, 桂林 541006;
4. 中国科学院华南植物园退化生态系统植被恢复与管理重点实验室, 广州 510650;
5. 中国科学院核心植物园植物生态中心, 广州 510650
2. Guangxi Normal University, Key Laboratory of Ecology of Rare and Endangered Species and Environmental Protection, Ministry of Education, Guilin 541006, China;
3. Guangxi Key Laboratory of Landscape Resources Conservation and Sustainable Utilization in Lijiang River Basin, Guilin 541006, China;
4. Key Laboratory of Vegetation Restoration and Management of South China Botanical Garden, Chinese Academy of Sciences, Guangzhou 510650, China;
5. Center of Plant Ecology, Core Botanical Gardens, Chinese Academy of Sciences, Guangzhou 510650, China
21世纪以来, 随着人类经济社会的飞速发展, 人类对生境的干扰导致多样性的损失日益加剧, 生物多样性的研究在全球范围内也备受关注[1]。森林动态大样地是生物多样性科学综合研究平台[2]。近年来, 随着对森林大型固定样地数据的不断深入获取, 生物多样性的研究也得到了长足的发展, 相关研究成果极大地推动了对群落物种共存机制的认知[3]。
20世纪60年代, 生态学家Robert H. Whittaker提出了α、β和γ多样性用来表征不同尺度群落的物种多样性[4]。α多样性检测特定群落或区域的物种丰富程度;β多样性检测不同群落间由于环境作用导致的物种组成差异或群落分化的程度;而γ多样性表征包括许多群落的大区域总的物种丰富度[5]。此后多样性测度的方法日趋完善, 并得到广泛应用, 特别是针对特定群落多样性水平评价时, α多样性备受推崇[6]。α多样性与群落其他特征的相关性研究也备受关注:有研究发现人工林物种多样性的增加有利于林分生物量的提高[7];对植物群落α多样性与地形特征的关系研究中发现中海拔区域的多样性高于高海拔和低海拔区域[8]。随着生物多样性监测范围的不断扩大, 采用β多样性指数比较样地间的多样性也引起关注[5, 9]。有研究用β多样性指数检测显示:西双版纳望天树样地内的群落组成和结构变化很大, 任意两固定样地间均有50%以上的植物物种不相同[10]。利用β多样性指数对湖南省八达贡山的环境异质性分析发现:不同地点的环境异质性值存在非常显著的差异[11]。
近年来, 围绕生物多样性的形成机制和影响因素也开展了一系列研究。如, 研究发现群落类型和海拔是中亚热带古田山森林群落α和β多样性的主要影响因子, 生境过滤等机制对该区域的森林物种多样性格局起着主要作用[12];隔离度显著影响岛屿植物的β多样性[13];坡向影响到秦岭松栎林群落生物多样性分布格局[14];以Jaccard物种相异性指数为指标发现群落物种在空间上的更替是环境限制和扩散限制综合作用的结果[15]。而目前在高生物多样性的南亚热带森林群落的此类研究还有待加强。
鼎湖山的南亚热带常绿阔叶林群落DHS大样地属于中国森林生物多样性监测网络成员, 迄今为止已经15年连续监测数据, 对其群落结构、多样性组成以及多样性维持机制方面开展了研究[16-18]。由于鼎湖山自然保护区植被的独特性(全球同纬度带上遍布沙漠), 以及其超过400年的保护历史, 造就了此区域内群落类型多样, 深入研究发现即使是20 hm2南亚热带常绿阔叶林群落中也存在不同发育阶段的斑块[19]。为了拓展和补充该区域多样性的监测范围, 在20 hm2南亚热带常绿阔叶林群落周围建立了5个1hm2卫星样地。本研究首次以鼎湖山20 hm2南亚热带地带性常绿阔叶林群落大样地结合其不同海拔和林型的卫星样地群落作为研究对象, 分别从种面积关系、α多样性和β多样性几个角度进行分析比对, 旨在探索南亚热带典型大样地群落与其卫星样地的多样性状况的异同性, 全面深入探索南亚热带森林区域多样性特征与分布格局, 为该区域森林群落的多样性保护和可持续发展提供科学依据。
1 研究地概况鼎湖山(以下简称, DHS)国家级自然保护区(112°30′39″-112°33′41″E, 23°09′21″-23°11′30″N)属于科技部重点野外台站, 对于鼎湖山的详细介绍见文献[20]。本研究选取20 hm2的南亚热带常绿阔叶林群落, 及其周围约2000 hm2范围内5个1 hm2的不同林型卫星样地群落为研究对象, 各样地位置及类型见表 1。各卫星样地与大样地的相对位置见图 1。样地调查按照CTFS(Center of Tropical Forest Sciences) 在热带森林生物多样性长期监测研究的方法来实施[21]。采用DHS大样地及其卫星样地2015年度的调查数据进行统计计算和分析。
| 样地号 Plot No. |
林型 Forest type |
样地尺度 Sample size/m |
海拔 Altitude/m |
原点北纬 Northern latitude |
原点东经 East longitude |
| 0 | 南亚热带常绿阔叶林(鼎湖山大样地) | 400×500 | 230-470 | 23°10′8.7″ | 112°32′2.1″ |
| 1 | 南亚热带山地常绿阔叶林 | 100×100 | 602-660 | 23°10′34″ | 112°31′30″ |
| 2 | 南亚热带沟谷雨林 | 80×125 | 90-133 | 23°10′15″ | 112°32′29″ |
| 3 | 南亚热带常绿阔叶林 (荷木林Schima superba forset) |
100×100 | 182-256 | 23°08′22″ | 112°30′49″ |
| 4 | 南亚热带常绿针叶林 (马尾松林Pinus massoniana forest) |
100×100 | 38-93 | 23°09′58″ | 112°33′28″ |
| 5 | 南亚热带针阔混交林 | 100×100 | 50-85 | 23°09′41″ | 112°32′36″ |

|
| 图 1 DHS大样地与及其卫星样地的相对位置 Fig. 1 Relative positions between DHS large-scale plot and satellite plots 0为DHS(鼎湖山Ding Hu Shan)大样地, 1-5分别为卫星样地的编号 |
通过R里的geoxy函数把6个样地的四个角点的经纬度转换为直角坐标系的x值与y值, 然后输出并绘制为矩形框图(图 1)。geoxy函数默认以最小经纬度样地的西南角点作为平面直角坐标的原点。6个样地中, 3号卫星样地的经纬度最小, 故它的西南角点为图 1直角坐标的零点。各卫星样地均为边长为100 m的正方形, DHS大样地X方向(东西方向)长度为400 m, Y方向(南北方向)为500 m。
2.2 种面积曲线绘制上述六个样地中, 20 hm2 DHS样地的数据经过核对检验后载入R工作空间, 从20 m到400 m每隔20 m取一个尺度总共20个尺度作为取样方框的边长, 每个尺度上随机位置方形取样各8次, 统计每次所取的物种数其均值。5个卫星样地数据, 则从5 m到100 m每次增加5 m共20个尺度, 直至取样完1 hm2样地全部植株, 每个尺度上随机方形取样8次, 相应面积所对应的物种数取自于随机取样的平均值。6个样地统计的系列数据对应到X-Y坐标图绘制为种-面积曲线。
2.3 多样性计算依次选取3种α多样性指数:Shannon-Weiner指数[22]、Simpson指数[23]与Heip均匀度指数[24];选取3个β多样性指数:Jaccard指数[25]、Sorenson指数[25]、Bray-Curtis指数[26];用于表征群落物种多样性状况的6个多样性指数详见表 2。
| ID | 指标 Index |
公式 Formula |
参数描述 Parameter description |
表征说明 Description of characterization |
| 1 | Shannon-Weiner指数 Shannon-Weiner index |
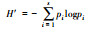 |
任意两个个体为不同物种的概率p时, pi为各个物种i的相对多度, S为目标样地中的总物种数 | 指数值越大表征群落的α多样性越高 |
| 2 | Simpson指数D Simpson index |
D=1-∑pi2 | ||
| 3 | Heip均匀度指数 Heip uniformity index |
J=(e-∑pilogpi-1)/(S-1) | ||
| 4 | Jaccard指数 Jaccard index |
Cj=j/(x+y-j) | j为两样地共有的物种数, x与y分别为两样地的物种数 | 指数值越大表征两个群落的相似性越高, 共有种越多, β多样性就越低 |
| 5 | Sorenson指数 Sorenson index |
Cs=2j/(x+y) | ||
| 6 | Bray-Curtis指数 Bray-Curtis index |
CN=2jN/(N1+N2) jN= ∑min(j N1, j N2) |
N1为样地1里所有个体数之和, N2为样地2所有个体数之和, jN为每个共有种在两个样地中个体数较少者之和[27] | 指数值越大表征两个群落相似性越高, 差异性越小, 即β多样性越低 |
上述所有数据处理均在R3.5上编程实现[28]。
3 结果与分析 3.1 物种概况及种面积关系各样地的物种概况见表 3, 其中的个体数量不含分枝数。20 hm2大样地共有80937个体, 分属于177物种。1 hm2卫星样地3号的物种数最多, 为96个物种;其次为1号和2号;4号和5号低于1-3号样地, 其中4号样地物种数最少, 为41种。5个卫星样地总个体数差异较大, 排序为:1号>3号>5号>4号>2号样地。1号南亚热带山地常绿阔叶林总个体数4050株, 明显多于其它卫星样地。2号南亚热带沟谷雨林个体数最少, 仅有1997株, 比1号样地少2053株, 甚至比4号马尾松林少371株。由于2号样地中分布着许多大树和巨树, 42株树木胸径DBH(Diameter at Breast Height)大于40 cm;有16株胸径大于60 cm, 其中锥栗(Castanopsis chinensis)9株;5株胸径超过100 cm的个体都是建群种锥栗(DBH分别为:119.4 cm、119.7 cm、165.6 cm、178 cm、184 cm), 因此2号样地的胸高断面积远大于其它样地。
| Plot No. | 个体数 Individuals |
丰富度 Richness |
平均胸径/cm Mean DBH (Diameter at Breast Height) |
胸高断面积/(m2/hm2) Basal area at breast height |
| 0 | 80937 | 177 | 4.98 | 25.28 |
| 1 | 4050 | 90 | 6.21 | 19.95 |
| 2 | 1997 | 88 | 8.20 | 36.17 |
| 3 | 3068 | 96 | 5.38 | 20.42 |
| 4 | 2368 | 41 | 6.41 | 19.98 |
| 5 | 2766 | 55 | 6.38 | 17.93 |
20 hm2大样地的种面积曲线显示(图 7):随着面积增大, 物种数不断增多, 特别是0-5 hm2范围增加的最快, 5 hm2时出现拐点, 物种数达到128, 是整个样地物种数的72.3%;20 hm2时仍然呈现轻微上升趋势, 说明20 hm2的典型样地物种并未达到完全饱和, 周围的卫星样地可以有效补充代表该区域物种。卫星样地的种面积曲线显示:4号南亚热带常绿针叶林(马尾松林)物种在1 hm2面积时物种达到饱和(图 5), 而其余四块样地物种均保持明显上升趋势, 充分体现了鼎湖山南亚热带森林群落高多样性特征。
3.2 α多样性比较物种丰富度S、Shannon-Weiner指数、Simpson指数和物种均匀度Heip指数表征各样地的物种多样性见表 4。比较发现:DHS大样地的多样性较高。综合Shannon-Weiner指数和Simpson指数结果发现:DHS样地多样性水平仅次于卫星样地2号南亚热带沟谷雨林和3号南亚热带常绿阔叶林。而表征物种个体数之间差异的物种均匀度Heip指数的计算结果表明DHS大样地物种均匀度较小, 仅高于5号南亚热带针阔混交林。
| 样地号 Plot No. |
物种丰富度S Species richness S |
Shannon-Weiner指数 Shannon-Weiner index |
Simpson指数 Simpson index |
Heip指数 Heip index |
| 0 | 177 | 3.167 | 0.901 | 0.129 |
| 1 | 90 | 2.909 | 0.902 | 0.195 |
| 2 | 88 | 3.254 | 0.934 | 0.286 |
| 3 | 96 | 3.187 | 0.925 | 0.244 |
| 4 | 41 | 2.417 | 0.860 | 0.255 |
| 5 | 55 | 1.891 | 0.672 | 0.104 |
卫星样地之间的比较结果表明(表 4):4号和5号卫星样地的丰富度明显低于其它类型样地, 其中4号物种丰富度最低。以荷木为优势种的3号样地的物种丰富度最高, 其次是1号和2号样地。体现物种数和相对多度综合作用的Shannon-Weiner指数结果显示:2号样地的物种多样性最高, 其次是3号样地, 继而是1号和4号样地, 最后是5号样地。而主要统计优势种影响力的Simpson指数, 也表现出与Shannon-Weiner指数相同的规律。体现群落均匀度Heip指数计算结果显示:2号样地的物种个体分布均匀度最大, 其次是4号样地, 继而是3号和1号样地, 最后是5号样地的针阔混交林的物种个体分布最不均匀, 这与针阔混交林植株聚集度较高有关。
考虑到DHS大样地面积远大于其卫星样地, 故综合比较面积相同的卫星样地的α多样性指数, 多样性指数计算结果显示(表 4):2号, 3号和1号卫星样地明显高于4号和5号卫星样地, 其中2号沟谷雨林样地Shannon-Weiner指数、Simpson指数及Heip指数的数值皆在5个样地中最大, 说明南亚热带沟谷雨林在各个样地群落中物种是最多样化的。4号和5号卫星样地均属于低海拔样地, 4号卫星样地南亚热带常绿针叶林(马尾松林)的物种丰富度最低, 但其海拔跨度(38-93m)大于5号卫星样地(50-85 m), 因此物种多样性高于5号卫星样地。
3.3 β多样性比较采用Jaccard、Sorenson和Bray-Curtis指数表征β多样性又称生境间的多样性, 表征群落之间由于生境差异造就的物种组成的相异性。3个指数统计显示(表 5):DHS大样地与其几个卫星样地的物种之间存在差异, 与3号卫星样地物种相似度最高(Jaccard指数和Sorenson指数最大), Bray-Curtis指数也较高, 高相似度的主要原因是两者植被类型较为接近;其次是2号卫星样地, DHS大样地与1号和5号卫星样地的物种差异逐渐增大;DHS大样地与其4号卫星样地的Jaccard指数和Sorenson指数最小, Bray-Curtis指数较低, 说明它们之间的物种组成差异相较于其他样地之间是最大的。
| 样地号 Plot No. |
Jaccard指数 Jaccard index |
Sorenson指数 Sorenson index |
Bray-Curtis指数 Bray-Curtis index |
||||||||||||||
| 1 | 2 | 3 | 4 | 5 | 1 | 2 | 3 | 4 | 5 | 1 | 2 | 3 | 4 | 5 | |||
| 0 | 0.335 | 0.395 | 0.429 | 0.141 | 0.234 | 0.502 | 0.566 | 0.601 | 0.248 | 0.379 | 0.084 | 0.046 | 0.071 | 0.051 | 0.064 | ||
| 1 | 0.245 | 0.319 | 0.149 | 0.283 | 0.506 | 0.484 | 0.260 | 0.441 | 0.301 | 0.643 | 0.290 | 0.187 | |||||
| 2 | 0.383 | 0.122 | 0.233 | 0.554 | 0.217 | 0.378 | 0.629 | 0.044 | 0.119 | ||||||||
| 3 | 0.234 | 0.291 | 0.380 | 0.450 | 0.248 | 0.455 | |||||||||||
| 4 | 0.297 | 0.458 | 0.841 | ||||||||||||||
卫星样地之间的差异为见表 5, 3个指数的计算结果均显示:1号、2号、3号样地之间差异较小, 2号和3号样地之间的物种相似度更高:Jaccard指数最大(0.383)和Sorenson指数同样在卫星样地间最高(0.554), Bray-Curtis指数也仅次于1号和3号之间(0.643), 屈居第二(0.629), 说明2号和3号卫星样地的群落结构很接近。
4号与1、2、3号卫星样地物种差异较大, 其中4号与2号卫星样地群落差异最大, 相似度Bray-Curtis值只有0.044。各个卫星样地之间的物种差异主要是海拔差异和群落小生境的不同而导致的。4号与5号样地的物种差异最小, 在4号样地与其他卫星样地中的相似度最高(Jaccard指数、Sorenson指数和Bray-Curtis指数都最大), 其中Bray-Curtis指数达到0.841, 即4号与5号卫星样地物种相似度高达84%。
4 讨论 4.1 种个体和种面积变化规律20 hm2大样地三次调查结果统计表明2005年到2010年期间乔木总多度有一定幅度的增加, 增加了6812株。可能与样地内的倒木死亡形成多个林窗有关, 林窗对森林生态系统结构和功能短期的调节作用比较明显[29]。林窗对群落组成、功能结构、物质循环、能量流动和生物多样性维持起着重要作用, 是群落演替的主要驱动力[30]。经过更新演替, 2015年DHS大样地的总个体数出现了爆发式增长, 比5年前增加2792株, 比10年前甚至增加了9604株。进一步探索群落更新演替的趋势还需要后期持续调查监测结果来分析。
卫星样地间物种数和个体数的差异主要是由其生境类型和海拔不同造成的。南亚热带山地常绿阔叶林(1号样地) 海拔远高于其他几块卫星样地, 其总个体数也遥遥领先, 其次是3号样地南亚热带常绿阔叶林(荷木林)。而南亚热带沟谷雨林由于位于低海拔区域, 受人为干扰较大, 其植株个体数在卫星样地中是最少的。也是因为样地中大树和巨树分布较多, 在群落中占到绝对优势, 导致相同面积上生长的个体数最少。
除了4号样地种面积曲线在1 hm2时物种趋于直线外, 其他卫星样地物种在1 hm2时均保持明显上升趋势, 甚至大样地在20 hm2时种面积曲线仍然呈现轻微上升趋势(图 2-7)。这也说明了对物种丰富的鼎湖山南亚森林群落生物多样性进行研究时还需要结合考虑周边卫星样地情况, 与大样地物种存在较大差异的卫星样地会是大样地物种更新的重要来源。

|
| 图 2 样地1种面积曲线图 Fig. 2 Species-area curve of plot 1 |

|
| 图 3 样地2种面积曲线图 Fig. 3 Species-area curve of plot 2 |

|
| 图 4 样地3种面积曲线图 Fig. 4 Species-area curve of plot 3 |

|
| 图 5 样地4种面积曲线图 Fig. 5 Species-area curve of plot 4 |

|
| 图 6 样地5种面积曲线图 Fig. 6 Species-area curve of plot 5 |

|
| 图 7 大样地种面积曲线图 Fig. 7 Species-area curve of large-scale plot |
根据多样性指数和均匀度指数的计算结果, 结合表 1各样地的海拔范围, 发现除了2号样地(虽然2号样地总物种个体数最低, 然而由于大量大树和巨树的存在导致物种均匀度提升, 从而使得多样性指数计算结果最大), 其它卫星样地呈现出多样性与海拔高度正相关的规律, 这与其他区域的相关研究结论不尽相同。海南岛不同海拔高度青皮林群落的多样性与海拔高度呈相反变化趋势[31]。白龙江干旱河谷不同坡向主要灌丛群落随着海拔的升高, 不同坡向物种数表现为先增加后减少的趋势, 即中海拔多样性最高[32]。对林窗植物多样性及其海拔动态研究显示:低海拔林窗中植物多样性最高, 其次为中海拔, 高海拔林窗中多样性中低海拔之间存在显著差异[33]。鼎湖山地处南亚热带区域, 水热条件充沛, 属低海拔丘陵区, 最高峰鸡笼山海拔1000m, 而南亚热带山地常绿阔叶林样地所处的最高海拔为660m。低海拔区域受一定的人为干扰, 相对区域内较高海拔样地其光热条件处于劣势, 因此低海拔样地多样性也较低。前期鼎湖山样带的研究也发现:随着海拔升高, 水、热、光等条件随海拔升高而趋于优化, 多样性也逐渐增大[34]。
同一区域内不同群落α多样性的大小也与其群落演替阶段密切相关。黑石顶森林群落演替系列α多样性的尺度效应研究表明:阔叶林样带比混交林样带在群落组成结构上具有更高的空间变异[35]。鼎湖山样带的研究发现:乔木层物种多样性指数是中生性阔叶林>针阔混交林>阳生性阔叶林[36]。针阔叶混交林是我国南亚热带针叶林向地带性常绿阔叶林演替的中间林分类型, 故丰富度处于中间, 本研究也发现了各个不同演替阶段群落物种丰富度的规律为:沟谷雨林、常绿阔叶林(1, 2, 3号样地)>针阔混交林(5号样地)>针叶林(4号样地)。
β多样性度量群落间物种组成的变化, 由物种更替和丰富度差异两种过程决定[3]。同一区域内处于相同或相近演替阶段的群落其物种相似度高, 相反处于演替阶段两极的群落物种相似度最低。南亚热带常绿阔叶林(DHS大样地)与其3号卫星样地(南亚热带常绿阔叶林-荷木林)处于相近演替阶段, 因此物种相似度最高:Sorenson指数检测出有60%的物种相同(表 5);与其4号卫星样地南亚热带常绿针叶林演替阶段相差最远, 物种差异性也最大:Bray-Curtis指数检测出仅有5%的物种相同;DHS大样地与其1、2、5号卫星样地的物种相似性都介于上述与3号和4号卫星样地之间。卫星样地之间的物种相似性也存在差异。因此, 这些与典型大样地物种各有差异的不同海拔和林型的卫星样地有力地促进了南亚热带典型森林大样地群落的物种更新和提高多样性。
5 结论鼎湖山物种资源丰富, 公认为北回归线上的“绿色明珠”, 对典型植物群落及其卫星样地物种多样性状况进行探索是一项非常有意义的研究工作。文中通过种面积关系、α多样性和β多样性的计算和分析比较, 得出以下几个主要结论:
(1) 20 hm2南亚热带常绿阔叶林(DHS大样地)经过10年的更新演替, 总个体数出现了爆发式增长。1 hm2卫星样地的物种数表现出:南亚热带常绿阔叶林(荷木林)最多, 南亚热带常绿针叶林(马尾松林)物种数最少。卫星样地之间总个体数差异较大。
(2) 种面积曲线表明:20 hm2 DHS大样地的物种并未达到完全饱和, 周围的卫星样地可以有效补充代表该区域物种, 卫星样地也是自然保护区多样性的重要组成部分。
(3)α多样性研究发现:南亚热带常绿阔叶林(DHS大样地) 的α多样性较高, 多样性水平仅次于卫星样地2号南亚热带沟谷雨林和3号南亚热带常绿阔叶林(荷木林)。同一区域内不同群落丰富度的大小与其群落演替阶段密切相关。
(4) 南亚热带常绿阔叶林(DHS大样地) 与其3号卫星样地南亚热带常绿阔叶林(荷木林)处于相近演替阶段, 因此物种相似度最高;与其4号卫星样地南亚热带常绿针叶林(马尾松林)演替阶段相差最远, 物种差异性也最大。卫星样地之间比较:4号南亚热带常绿针叶林(马尾松林)与5号南亚热带针阔混交林的演替阶段相近, 物种相似度最高, Bray-Curtis指数表明两者具有84%的相似度。
综上所述, 若要获取鼎湖山国家级自然保护区整个区域群落的多样性特征, 研究不能仅聚焦到该区域南亚热带常绿阔叶林典型大样地(DHS大样地), 还需统筹考虑其周边辐射范围不同海拔和林型地卫星样地群落。通过联合研究多个卫星样地与20 hm2的DHS大样地构成的复合群落系统, 即可给南亚热带常绿阔叶林区域森林群落的多样性特征与分布格局以一个精准画像, 为区域生物多样性保护工作提供更加科学的理论依据。
| [1] |
马克平. 生物多样性科学的热点问题. 生物多样性, 2016, 24(1): 1-2. |
| [2] |
马克平. 森林动态大样地是生物多样性科学综合研究平台. 生物多样性, 2017, 25(3): 227-228. |
| [3] |
马克平, 徐学红. 中国森林生物多样性监测网络有力支撑生物群落维持机制研究. 中国科学: 生命科学, 2020, 50(4): 359-361. |
| [4] |
Whittaker R H. Vegetation of the Siskiyou Mountains, Oregon and California. Ecological Monographs, 1960, 30(3): 279-338. DOI:10.2307/1943563 |
| [5] |
斯幸峰, 赵郁豪, 陈传武, 任鹏, 曾頔, 吴玲兵, 丁平. Beta多样性分解: 方法、应用与展望. 生物多样性, 2017, 25(5): 464-480. |
| [6] |
马克平, 刘玉明. 生物群落多样性的测度方法Ⅰα多样性的测度方法(下). 生物多样性, 1994, 2(4): 231-239. DOI:10.3321/j.issn:1005-0094.1994.04.009 |
| [7] |
温远光, 陈放, 刘世荣, 梁宏温, 元昌安, 朱宏光. 广西桉树人工林物种多样性与生物量关系. 林业科学, 2008, 44(4): 14-19. |
| [8] |
宋玲玲, 田青, 李宗杰. 岷山摩天岭北坡东南部植物群落α多样性与地形特征的关系. 甘肃农业大学学报, 2017, 52(3): 67-74. |
| [9] |
陈圣宾, 欧阳志云, 徐卫华, 肖燚. Beta多样性研究进展. 生物多样性, 2010, 18(4): 323-335. |
| [10] |
汤明华. 西双版纳望天树林的群落组成和结构特征研究. 西部林业科学, 2007, 36(2): 26-32. DOI:10.3969/j.issn.1672-8246.2007.02.005 |
| [11] |
熊芳, 汪政科, 朱忠保, 朱林峰, 熊四清. 八大公山多样性分析与物种分布格局研究. 林业科学, 1997, 33(S1): 20-30. |
| [12] |
翁昌露, 张田田, 巫东豪, 陈声文, 金毅, 任海保, 于明坚, 罗媛媛. 古田山10种主要森林群落类型的α和β多样性格局及影响因素. 生物多样性, 2019, 27(1): 33-41. |
| [13] |
王金旺, 魏馨, 陈秋夏, 李效文, 杨升. 温州沿海小型海岛植物丰富度和β多样性及其影响因子. 生态学报,, 2017, 37(2): 523-540. |
| [14] |
吴昊. 秦岭松栎林群落物种丰富度特征及其环境解释. 生态环境学报, 2017, 26(6): 931-938. |
| [15] |
蒙洋, 邱月, 张亮, 王翠玲, 臧振华, 张学耀, 申国珍, 闫彩凤, 陈全胜. 地理距离、海拔和气候差异对独龙江流域维管植物群落物种空间相异性的影响. 生物多样性, 2017, 25(12): 1313-1320. DOI:10.17520/biods.2017076 |
| [16] |
叶万辉, 曹洪麟, 黄忠良, 练琚愉, 王志高, 李林, 魏识广, 王章明. 鼎湖山南亚热带常绿阔叶林20公顷样地群落特征研究. 植物生态学报, 2008, 32(2): 274-286. |
| [17] |
Li L, Huang Z L, Ye W H, Cao H L, Wei S G, Wang Z G, Lian J Y, Sun I F, Ma K P, He F L. Spatial distributions of tree species in a subtropical forest of China. OIKOS, 2009, 118(4): 495-502. DOI:10.1111/j.1600-0706.2009.16753.x |
| [18] |
Wei S G, Li L, Walther B A, Ye W H, Huang Z L, Cao H L, Lian J Y, Wang Z G, Chen Y Y. Comparative performance of species-richness estimators using data from a subtropical forest tree community. Ecological Research, 2010, 25(1): 93-101. DOI:10.1007/s11284-009-0633-2 |
| [19] |
练琚愉, 陈灿, 黄忠良, 曹洪麟, 叶万辉. 鼎湖山南亚热带常绿阔叶林不同成熟度群落特征的比较. 生物多样性, 2015, 23(2): 174-182. |
| [20] |
黄忠良, 孔国辉, 魏平. 鼎湖山植物物种多样性动态. 生物多样性, 1998, 6(2): 116-121. |
| [21] |
Condit R. Tropical Forest Census Plots: Methods and Results from Barro Colorado Island, Panama and a Comparison with Other Plots. Berlin: Springer-Verlag, 1998.
|
| [22] |
Shannon C E, Weaver W. The Mathematical Theory of Communication. Urbana: University of Illinois Press, 1949.
|
| [23] |
Simpson E H. Measurement of diversity. Nature, 1949, 163(4148): 688-688. DOI:10.1038/163688a0 |
| [24] |
Heip C. A new index measuring evenness. Journal of the Marine Biological Association of the United Kingdom, 1974, 54(3): 555-557. DOI:10.1017/S0025315400022736 |
| [25] |
马克平, 刘灿然, 刘玉明. 生物群落多样性的测度方法Ⅱβ多样性的测度方法. 生物多样性, 1995, 3(1): 38-43. |
| [26] |
Bray J R, Curtis J T. An ordination of the upland forest communities of southern Wisconsin. Ecological Monographs, 1957, 27(4): 325-349. DOI:10.2307/1942268 |
| [27] |
林国俊, 黄忠良, 竺琳, 欧阳学军. 鼎湖山森林群落β多样性. 生态学报, 2010, 30(18): 4875-4880. |
| [28] |
R Core Team. R: A Language and Environment for Statistical Computing. Vienna, Austria: R Foundation for Statistical Computing, 2018.
|
| [29] |
柳泽鑫, 张璐, 区余端, 孙东, 苏志尧. 冰灾后粤北常绿阔叶林粗死木质残体的组成与结构. 西南林业大学学报, 2011, 31(4): 18-23. |
| [30] |
唐凤华, 全文选, 李朝婵, 钱沉鱼, 许塔艳, 杨荞安. 天然林林窗与自然更新研究进展. 西部林业科学, 2018, 47(4): 95-101. |
| [31] |
郝清玉, 刘淑菊, 钟琼芯, 辛琨, 刘强. 海南岛不同海拔高度青皮林群落结构与多样性. 林业资源管理, 2012(4): 48-54. |
| [32] |
金毅, 陈建华, 米湘成, 任海保, 马克平, 于明坚. 古田山24 ha森林动态监测样地常绿阔叶林群落结构和组成动态: 探讨2008年冰雪灾害的影响. 生物多样性, 2015, 23(5): 610-618. |
| [33] |
陈力, 刘国华, 刘丹, 石松林. 亚高山针叶林不同大小林窗植物多样性及其海拔动态. 中南林业科技大学学报, 2017, 37(10): 90-97. |
| [34] |
李林, 周小勇, 黄忠良, 魏识广, 史军辉. 鼎湖山植物群落α多样性与环境的关系. 生态学报, 2006, 26(7): 2301-2307. |
| [35] |
王永繁, 余世孝, 黄向, 刘蔚秋. 黑石顶森林群落演替系列α多样性的尺度效应. 中山大学学报: 自然科学版, 2002, 41(3): 68-71, 76-76. |
| [36] |
史军辉, 黄忠良, 周小勇, 欧阳学军, 李炯, 张池. 鼎湖山森林群落多样性垂直分布格局的研究. 生态学杂志, 2005, 24(10): 1143-1146. |